
Contents
Research Article
One-relator surface groups
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 467-474
-
- Article
- Export citation
Homotopy skein modules of orientable 3-manifolds
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 475-488
-
- Article
- Export citation
Quantum groups and representations of monoidal categories
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 261-290
-
- Article
- Export citation
On ideals and sublattices in linear lattices and F-lattices
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 79-87
-
- Article
- Export citation
-
The main aim of this paper is to study the role of the assumptions in the following
Theorem 0. Every ideal I of codimension 1 in an F-lattice X is closed†.
For X a Banach lattice this result appears in [11], ii.5·3, corollary 3. The proof carries over to the case where X is an. F-lattice, i.e. a topological linear lattice the topology of which is metrizable and complete, with the help of a result due to Klee ([10], theorem v.5·5; see also [3], theorem 16·6). We first note that the assumption of metric completeness is essential (Example 2). We then extend Theorem 0 to the case of ideals of finite codimension (Theorem 1), and show that non-closed ideals exist whenever X is infinite-dimensional (Corollary 2). Our main result commenting on Theorem 0 is, however, the existence of dense sublattices of arbitrary codimension between 1 and
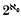 in every infinite-dimensional F-lattice (Theorem 6).
in every infinite-dimensional F-lattice (Theorem 6).
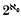 in every infinite-dimensional
in every infinite-dimensional A calculus for rational tangles: applications to DNA recombination
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 489-515
-
- Article
- Export citation
Degenerate curves and harmonic analysis
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 89-96
-
- Article
- Export citation
Localization of seif-homotopy equivalences inducing the identity on homology
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 291-297
-
- Article
- Export citation
Orientable surfaces in the 4-sphere associated with non-orientable knotted surfaces
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 299-306
-
- Article
- Export citation
-
Let F be a closed connected and non-orientable surface smoothly embedded in the 4-sphere S4 with normal Euler number e(F) = 0. We note that if e(F) = 0, then the non-orientable genus n
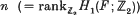 is even (ef. [7]) and the tubular neighbourhood N(F) of F in S4 which is a D2-bundle over F has a trivial I-subbundle. Let τ be a trivial I-subbundle of N(F) and let τ* = F × I ⊂ N(F) be its orthogonal I-subbundle which is twisted. Then
is even (ef. [7]) and the tubular neighbourhood N(F) of F in S4 which is a D2-bundle over F has a trivial I-subbundle. Let τ be a trivial I-subbundle of N(F) and let τ* = F × I ⊂ N(F) be its orthogonal I-subbundle which is twisted. Then 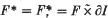 is a closed connected genus n – 1 orientable surface smoothly embedded in S4 and doubly covers F. We call this surface a doubled surface of F in S4 (associated with τ). If a trivial I-subbundle τ is given, then we see that the knot type of F* ⊂ S4 is uniquely determined.
is a closed connected genus n – 1 orientable surface smoothly embedded in S4 and doubly covers F. We call this surface a doubled surface of F in S4 (associated with τ). If a trivial I-subbundle τ is given, then we see that the knot type of F* ⊂ S4 is uniquely determined.
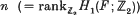 is even (ef. [
is even (ef. [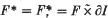 is a closed connected genus
is a closed connected genus The sum of Rademacher functions and Hausdorff dimension
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 97-103
-
- Article
- Export citation
-
For 0 < α < 1, let
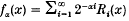 for 0 ≤ x < 1, where
for 0 ≤ x < 1, where 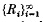 is the sequence of Rademacher functions. We give a class of fα so that their graphs have Hausdorff dimension 2 − α. The result is closely related to the corresponding unsolved question for the Weierstrass functions.
is the sequence of Rademacher functions. We give a class of fα so that their graphs have Hausdorff dimension 2 − α. The result is closely related to the corresponding unsolved question for the Weierstrass functions.
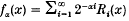 for 0 ≤ x < 1, where
for 0 ≤ x < 1, where 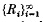 is the sequence of Rademacher functions. We give a class of
is the sequence of Rademacher functions. We give a class of Characteristic classes of Galois representations
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 517-522
-
- Article
- Export citation
-
Let K be a field with char if K≠2 and let Ks denote the separable closure of K and GK the Galois group of the extension Ks/K. If K⊂L is a finite extension and ρ:GL↦Or(R) a (continuous) real representation of GL we have a map ρ:BGL→BO which is used to define Stiefel–Whitney classes wi(ρ) = ρ*(wi). In general if f is any element of H*(BO; ℤ/2) we denote by f(ρ) the characteristic class ρ*(f). Now let
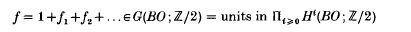
be a genus (see e.g. [9]), for example the total Stiefel–Whitney class w = 1+w1+w2 + … Let K⊂L and ρ be as above and let
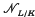 denote the multiplicative transfer (see e.g. [3, 5, 2, 14, 15]). Our principal result is a generalization of theorem 1 of [3]
denote the multiplicative transfer (see e.g. [3, 5, 2, 14, 15]). Our principal result is a generalization of theorem 1 of [3]
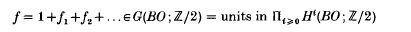
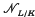 denote the multiplicative transfer (see e.g. [3,
denote the multiplicative transfer (see e.g. [3, The Hausdorff dimension of certain sets of singular n-tuples
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 105-110
-
- Article
- Export citation
Copies of l∞ in an operator space
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 523-526
-
- Article
- Export citation
Periodic orbits and fixed points of a C1 orientation-preserving embedding of D2
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 307-310
-
- Article
- Export citation
The uniform compactification of a locally compact abelian group
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 527-538
-
- Article
- Export citation
-
In recent years, the Stone-Čech compactification of certain semigroups (e.g. discrete semigroups) has been an interesting semigroup compactification (i.e. a compact right semitopological semigroup which contains a dense continuous homomorphic image of the given semigroup) to study, because an Arens-type product can be introduced. If G is a non-compact and non-discrete locally compact abelian group, then it is not possible to introduce such a product into the Stone-Čech compactification βG of G (see [1]). However, let UC(G) be the Banach algebra of bounded uniformly continuous complex functions on G, and let UG be the spectrum of UC(G) with the Gelfand topology. If f∈ UC(G), then the functions f and fy defined on G by
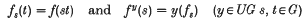
are also in UC(G).
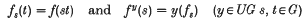
A Markov chain identity and monotonicity of the diffusion constants for a random walk in a heterogeneous environment
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 111-126
-
- Article
- Export citation
Character-pure locally compact abelian groups and their dual groups
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 311-315
-
- Article
- Export citation
Annihilators in JB-algebras
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 317-323
-
- Article
- Export citation
Toeplitz operators associated with analytic crossed products
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 539-549
-
- Article
- Export citation
Word problems related to periodic solutions of a non-autonomous system
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 127-151
-
- Article
- Export citation
-
In this paper, we develop an abstract formulation of a problem which arises in the investigation of the number of limit cycles of systems of the form
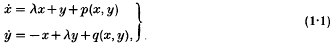
where p and q are homogeneous polynomials. This is part of the much wider study of Hilbert's sixteenth problem, in which information is sought about the number of limit cycles of systems of the form
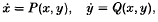
where P and Q are polynomials, and their possible configurations.
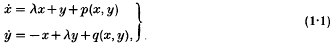
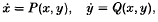
Three-dimensional constitutive equations for rigid/perfectly plastic granular materials
-
- Published online by Cambridge University Press:
- 24 October 2008, pp. 153-169
-
- Article
- Export citation