



1. Introduction
Curiosity and creativity are considered fundamental drives in human behavior (e.g., Boden, Reference Boden2004; Kobayashi, Ravaioli, Baranès, Woodford, & Gottlieb, Reference Kobayashi, Ravaioli, Baranès, Woodford and Gottlieb2019). Curiosity has been defined in the past as a motivation to explore uncertain environments (Li et al., Reference Li, Huo, Zhuang, Song, Wang, Ren and Qiu2019; Litman, Reference Litman2005), and as a state by which one is rewarded by new knowledge and novel experiences (Kashdan & Silvia, Reference Kashdan, Silvia, Lopez and Snyder2009; Litman & Jimerson, Reference Litman and Jimerson2004; Litman & Spielberger, Reference Litman and Spielberger2003). According to the classical definition of Berlyne (Reference Berlyne1966), curiosity is “the condition of discomfort due to an inadequacy of information that motivates specific exploration” (p. 26). Others referred to curiosity as a natural exploratory process aimed at gaining new information (Tian, Silva, & Liu, Reference Tian, Silva and Liu2021), which is thought to induce other essential behaviors for exploring novel, surprising, or intriguingly complex stimuli (Kidd & Hayden, Reference Kidd and Hayden2015). These different definitions all converge to seeing curiosity as a state by which one seeks novelty.
As to creativity, it has been operationalized in the past as an active behavior and as a personal trait, and in both contexts, it has been consensually defined as the creation, or ability to create something that is both novel and useful (Runco & Acar, Reference Runco and Acar2012; Sternberg & Lubart, Reference Sternberg and Lubart1996). To delineate our discussion within existing research, here we predominantly focus on curiosity as a state and on creativity as a behavior or action and their underlying cognitive processes, as they are controllably operationalized and measured in a lab, in hope that the theoretical framework proposed here may apply to curiosity and creativity “in the wild.”
The crux of our proposal is that curiosity and creativity converge on novelty-seeking mechanisms, whereby curiosity is a novelty-seeking state in which we more readily attend and absorb novelty in the world, screening what will eventually be learned and consolidated, in creativity we seek novel recombinations of stored knowledge. Within this framework, we further propose that because curiosity facilitates attention, categorization, and consolidation of new information, it serves creativity, which uses stored representations in memory to generate novel ones.
Curiosity has been conceptually associated with creativity in the past (Day & Langevin, Reference Day and Langevin1969; Maw & Maw, Reference Maw and Maw1970), and several empirical studies have investigated this potential relationship. These studies indicated a positive correlation between curiosity and creativity measures in adults (Hardy, Ness, & Mecca, Reference Hardy, Ness and Mecca2017; Vidler & Karan, Reference Vidler and Karan1975), high school students (Karwowski, Reference Karwowski2012), and children (Rubenstein, Reference Rubenstein2000). Hagtvedt, Dossinger, Harrison, and Huang (Reference Hagtvedt, Dossinger, Harrison and Huang2019) provided an initial demonstration of a predictive link between curiosity and creativity by showing that inducing curiosity increased creativity score in a subsequent idea generation task, supporting the possibility of a causal link between the two. Similarly, a recent small-scale meta-analysis of 10 studies concluded that there was a positive correlation between self-reported curiosity and both self-reported and rated creativity (Schutte & Malouff, Reference Schutte and Malouff2019). Thus, although curiosity and creativity are typically studied separately, these few reports provide initial support to the link between them.
In this article, we link curiosity and creativity by showing the covariations and reciprocal interactions between them in diverse contexts and conditions. After substantiating this link, we propose a novelty-seeking model (NSM) that underlies them. Finally, we show how one's state of mind (SoM; Herz, Baror, & Bar, Reference Herz, Baror and Bar2020) determines the manifestation of novelty seeking.
2. The cognitive underpinning of curiosity and creativity
Curiosity and creativity are complex cognitive constructs that involve several psychological mechanisms and brain networks. In this section, we show how curiosity and creativity act similarly across multiple domains, reflecting their proposed connection. We focus on aspects such as openness to experience and to uncertainty, as well as on key functions of human cognition such as attention, memory, and cognitive control – all of which were found to be involved in curiosity and creativity, independently and in a similar way. To account for these dynamics, we follow the SoM framework (Herz et al., Reference Herz, Baror and Bar2020) as a global concept. This framework proposes that the mind is a multidimensional construct that changes according to circumstances, influencing our subjective experience of the environment. To explain how the dynamics of different dimensions relate to one another, the SoM framework suggests that dimensions change along a continuum ranging from narrow to broad. Broad thinking entails broad perception, global attention, exploratory behavior, and positive mood. Conversely, narrow thinking entails narrowed perception, local attention, exploitatory behavior, and negative mood. We suggest that the global SoM and its related dimensions exert coordinated effects on curiosity and creativity, most directly facilitated by an exploratory SoM. Hereafter, we aim to integrate the literature from both domains to support the unified nature of curiosity and creativity. It should be noted that despite our efforts to allocate equal attention to curiosity and creativity, empirical research in the field of curiosity has remained relatively scarce in comparison to the extensive body of research on creativity. That said, we posit that the integration of literature on curiosity and creativity presented herein has the potential to yield reciprocal advantages and catalyze advancements in both domains.
2.1 Openness to experience
The SoM dimension most relevant for the understanding of curiosity and creativity is openness to experience, which determines the way we approach novel information. Openness to experience is traditionally considered to be a personality trait, part of the “big five” model of personality structure (Digman, Reference Digman1997), which is tightly related to both curiosity (Silvia & Christensen, Reference Silvia and Christensen2020; Silvia & Sanders, Reference Silvia and Sanders2010) and creativity (Beaty & Silvia, Reference Beaty and Silvia2012; Li et al., Reference Li, Li, Huang, Kong, Yang, Wei and Liu2015; McCrae, Reference McCrae1996). Openness to experience is the most consistent personality predictor of different forms of creativity: creative thought (Puryear, Kettler, & Rinn, Reference Puryear, Kettler and Rinn2017), self-perception (Karwowski & Lebuda, Reference Karwowski and Lebuda2016), activity (Batey & Furnham, Reference Batey and Furnham2006), and achievement (Feist, Reference Feist1998). At the same time, curiosity is a central component of openness to experience, which represents a core attribute of the trait (Silvia & Christensen, Reference Silvia and Christensen2020).
According to the SoM framework, beyond being a personality trait, openness to experience is also a dynamic state along the exploration/exploitation continuum, and whether we explore or exploit depends on the specific current context (Herz et al., Reference Herz, Baror and Bar2020). In an exploratory SoM, people attend the environment with a wider scope, are more learning-oriented, and are attuned more to sensory input than to familiar knowledge in memory (Herz et al., Reference Herz, Baror and Bar2020; Hills et al., Reference Hills, Todd, Lazer, Redish, Couzin, Bateson and Wolfe2015). This mode has been shown to increase the likelihood of making a thought breakthrough (Andriopoulos & Lewis, Reference Andriopoulos and Lewis2009). On the other hand, in an exploitatory SoM, people rely more on what they already know and on a priori expectations, less open to novelty and surprises, and gravitate more to the details than to the “big picture” (Schwartenbeck, FitzGerald, Dolan, & Friston, Reference Schwartenbeck, FitzGerald, Dolan and Friston2013). Interestingly, it has been further suggested that novelty seeking is affected by the tension between exploration and exploitation, such that increased exploitation reduces the chance for novel responses in a free associations task (Baror & Bar, Reference Baror and Bar2016).
An exploratory orientation enables the acquisition of new knowledge and identification of novel associations between concepts (Acar & van den Ende, Reference Acar and van den Ende2016; Baror & Bar, Reference Baror and Bar2016), which are required for both curiosity and creativity. Curiosity has been conceptualized in the past as an exploratory decision-making strategy, in which the choice made is not associated with any external rewards (Davis, Settlage, & Harlow, Reference Davis, Settlage and Harlow2012; Harlow, Harlow, & Meyer, Reference Harlow, Harlow and Meyer1950; Kidd & Hayden, Reference Kidd and Hayden2015). This relationship was further suggested to be bidirectional, such that higher levels of interest in an activity, which indicate curiosity, predict in turn exploratory actions (Berlyne, Reference Berlyne1966; Kashdan & Silvia, Reference Kashdan, Silvia, Lopez and Snyder2009).
Exploration has also been found to be involved in creative thinking (Carroll, Reference Carroll2011; Picciuto & Carruthers, Reference Picciuto, Carruthers, Paul and Kaufman2014), creative problem solving (Ruscio & Amabile, Reference Ruscio and Amabile1999), and improvisation (Martín, Ric, & Hristovski, Reference Martín, Ric and Hristovski2015). Exploration was argued to favor creativity by facilitating the sampling of different stimuli (Laureiro-Martínez, Brusoni, & Zollo, Reference Laureiro-Martínez, Brusoni and Zollo2010), eventually increasing the repertoire of possible responses, thus enhancing novel thoughts and actions (Blikstein, Silveira Gomes, Teruo Akiba, & Schneider, Reference Blikstein, Silveira Gomes, Teruo Akiba and Schneider2017). Similarly, demand for exploration was argued to increase as the desire for originality increases (Madjar, Greenberg, & Chen, Reference Madjar, Greenberg and Chen2011), further linking originality with exploration. In that sense, considering that creativity requires both originality and usefulness, it seems that exploration is especially associated with the “novel” and “original” rather than “useful” component in creativity. This point is further exemplified in recent work by Steele, Hardy, Day, Watts, and Mumford (Reference Steele, Hardy, Day, Watts and Mumford2021) who examined whether exploration and exploitation are related to creativity in different ways. The authors found that although exploration was positively related to the novelty of the product, exploitation was related to the usefulness of the product.
Exploitatory behavior was argued to promote monitoring, control, and persistence (Steele et al., Reference Steele, Hardy, Day, Watts and Mumford2021), allowing one's performance to be evaluated and refined (Ford, Weissbein, Smith, Gully, & Salas, Reference Ford, Weissbein, Smith, Gully and Salas1998). More specifically, according to reinforcement learning models, although exploration enables a wider search for a greater range of information, the narrower search in exploitation is more efficient and takes full advantage of existing information (Harada, Reference Harada2020). Along similar lines, Ruscio and Amabile (Reference Ruscio and Amabile1999) gave participants a novel structure-building task: Half of them received heuristic instructions for the task that aimed to increase exploratory behavior, and the other half received algorithmic instructions (a step-by-step demonstration for building a sample structure) that aimed to increase exploitatory behavior. The heuristic instructions led to a flexible experimentation with the task materials, as well as a tendency to use learned techniques in a less rigid fashion. Interestingly, participants receiving the algorithmic instructions exhibited greater confidence and speed during the task, but they were less likely to engage in exploratory behavior or to deviate from the sample structure. These results further demonstrate the different contribution of exploration and exploitation to the creative process. Both exploration and exploitation appear to be necessary for creativity, but their corresponding contribution is exerted at different phases of the creative process (Harada, Reference Harada2020).
One should take into account that exploration and exploitation strategies present trade-offs (Lavie, Stettner, & Tushman, Reference Lavie, Stettner and Tushman2010; March, Reference March1991), which must be managed during the process (Andriopoulos & Lewis, Reference Andriopoulos and Lewis2009). Throughout acts of creativity or curiosity, people alternate between an exploratory SoM, which is necessary for novelty seeking, and an exploitatory SoM, which is necessary for further refinement and elaboration (March, Reference March1991; Mehlhorn et al., Reference Mehlhorn, Newell, Todd, Lee, Morgan, Braithwaite and Gonzalez2015). In creativity, exploration enables generating original ideas, whereas exploitation is needed for further elaboration and evaluation. A similar process may take place in curiosity. Curiosity is typically identified with exploration of the environment for novelty, but exploitation may be needed for selecting what information to gather for further learning and consolidation. Although the dynamic of the shifts between exploration and exploitation requires further investigation, it seems that both creativity and curiosity rely on the intricate balance between exploration and exploitation (Herz et al., Reference Herz, Baror and Bar2020; Schad, Lewis, Raisch, & Smith, Reference Schad, Lewis, Raisch and Smith2016; Smith & Lewis, Reference Smith and Lewis2011).
Clearly, openness to experience is not dependent solely on the balance between exploration and exploitation. It is rather a multidimensional construct (Christensen, Kenett, Cotter, Beaty, & Silvia, Reference Christensen, Kenett, Cotter, Beaty and Silvia2018; Silvia & Christensen, Reference Silvia and Christensen2020), and some of its facets (such as the need of novelty; Joy, Reference Joy2004; Lynn & Snyder, Reference Lynn, Snyder, Snyder and Lopez2002) might even mediate the link between curiosity and creativity.
2.2 Tolerance for uncertainty
Uncertain environments provide significant amounts of new information. As human beings, we seek a sense of certainty, which helps us plan and be prepared for the future (i.e., exploitation SoM). At other times and states (exploration SoM; see Herz et al., Reference Herz, Baror and Bar2020, for a review), we are tolerant for surprises and seek novelty.
Research has suggested that curiosity facilitates information seeking and interest in the unknown, despite perceived potential negative consequences (Hsee & Ruan, Reference Hsee and Ruan2016; Kashdan & Silvia, Reference Kashdan, Silvia, Lopez and Snyder2009). In a similar fashion, curious individuals are more likely to see problems as challenges to be solved rather than as insurmountable setbacks (Denneson, Smolenski, Bush, & Dobscha, Reference Denneson, Smolenski, Bush and Dobscha2017). Thus, although some may attempt to minimize uncertainty by refraining from novelty altogether, the state of curiosity, almost by definition, serves as a mechanism for minimizing uncertainty by approaching new knowledge – a notion supported by recent studies (Blanchard, Hayden, & Bromberg-Martin, Reference Blanchard, Hayden and Bromberg-Martin2015; van Lieshout, Vandenbroucke, Müller, Cools, & de Lange, Reference van Lieshout, Vandenbroucke, Müller, Cools and de Lange2018, Reference van Lieshout, de Lange and Cools2019). In this fashion, curiosity might serve as an approach–avoidance regulatory function (Kashdan & Silvia, Reference Kashdan, Silvia, Lopez and Snyder2009), influencing one's tendency to withstand or engage in certain activities based on an uncertainty appraisal. The avoidance system constrains the breadth of attention (Derryberry & Tucker, Reference Derryberry, Tucker, Neidenthal and Kitayama1994; Friedman & Förster, Reference Friedman and Förster2010), and the approach system promotes broad associative thought (Chermahini & Hommel, Reference Chermahini and Hommel2012). Because curiosity is associated with increased exploration, it may sustain an approach orientation (Gasper & Middlewood, Reference Gasper and Middlewood2014), ultimately increasing the probability to engage in creative activities (e.g., de Dreu, Nijstad, & Baas, Reference de Dreu, Nijstad and Baas2010; Flaherty, Reference Flaherty2005; Friedman & Förster, Reference Friedman and Förster2010; Jauk, Reference Jauk2019) that relate to novelty seeking and openness to experience as well (Baas, Nijstad, Koen, Boot, & de Dreu, Reference Baas, Nijstad, Koen, Boot and de Dreu2020).
Interestingly, curiosity is associated with being more comfortable with anxiety (Kashdan et al., Reference Kashdan, Farmer, Adams, Ferssizidis, McKnight and Nezlek2013), allowing risk-taking behavior, which has long been acknowledged as integral to creativity (Dewett, Reference Dewett2007; Eisenman, Reference Eisenman2013; Feist, Reference Feist1998; Runco, Reference Runco2015; Simpson Steele, Reference Simpson Steele2015; Sternberg & Lubart, Reference Sternberg and Lubart1992). Taking sensible risks was postulated to be a prerequisite for creativity, as it promotes breakthroughs and innovativeness (Baas, Koch, Nijstad, & de Dreu, Reference Baas, Koch, Nijstad and de Dreu2015; Sternberg & Lubart, Reference Sternberg and Lubart1992), further linking uncertainty with exploration and novelty in creativity. Similarly, uncertainty was found to enhance the generation of ideas (Audia & Goncalo, Reference Audia and Goncalo2007), whereas avoiding uncertainty may restrain individuals from generating novel ideas (Erez & Nouri, Reference Erez and Nouri2010). More directly, Mueller, Melwani, and Goncalo (Reference Mueller, Melwani and Goncalo2012) found that having high tolerance for uncertainty was associated with greater chances of recognizing a creative idea, compared with having low tolerance for uncertainty. Together, it is not unlikely that curiosity, which is considered a state in which uncertainty-related anxiety is reduced, may ultimately result in enhancing creativity. Specifically, curiosity may help transforming situations of uncertainty from being experienced as threatening to provide a fruitful ground for the generation of novel ideas.
2.3 Attention to the novel
Curiosity and creativity are both tightly and reciprocally linked to the scope of attention. In some sense, curiosity directs our attentional “spotlight” toward what we perceive as potentially interesting (Gottlieb, Oudeyer, Lopes, & Baranes, Reference Gottlieb, Oudeyer, Lopes and Baranes2013; Kidd & Hayden, Reference Kidd and Hayden2015). In creativity, similar attentional processes take place, broadly considering representations that would potentially facilitate a novel combination (Benedek, Reference Benedek, Jung and Vartanian2018; Chun, Golomb, & Turk-Browne, Reference Chun, Golomb and Turk-Browne2010). Here we propose that curiosity and creativity rely on shared attentional mechanisms. Supporting this notion, Gross, Araujo, Zedelius, and Schooler (Reference Gross, Araujo, Zedelius and Schooler2019) recently demonstrated that both curiosity and creativity are characterized by exploratory eye movements. Curiosity was associated with faster anticipatory gaze shifts to the expected location of the answer in trivia tasks (Baranes, Oudeyer, & Gottlieb, Reference Baranes, Oudeyer and Gottlieb2015), and wider saccadic exploration of visual scenes was associated with trait curiosity (Risko, Anderson, Lanthier, & Kingstone, Reference Risko, Anderson, Lanthier and Kingstone2012). The level of curiosity was even predicted by gaze patterns using machine learning methods (Baranes et al., Reference Baranes, Oudeyer and Gottlieb2015).
Although the link between curiosity and attention is quite clear, in creativity this link is more complex, because of the two-stage conceptualization of creativity that begins with broad idea generation and concludes with a narrow selection based on usefulness or appropriateness. Creativity was found to relate to different types of attention such as broad attentional scope (Ansburg & Hill, Reference Ansburg and Hill2003), focused attention (Nusbaum & Silvia, Reference Nusbaum and Silvia2011), “leaky” or defocused attention, that is, attention that allows “irrelevant” information to be noticed (Carson, Higgins, & Peterson, Reference Carson, Higgins and Peterson2003; Martindale, Reference Martindale and Sternberg1999; Mendelsohn & Griswold, Reference Mendelsohn and Griswold1964; Rawlings, Reference Rawlings1985), and flexible attention, that is, the ability to switch between focused and defocused attention (Vartanian, Martindale, & Kwiatkowski, Reference Vartanian, Martindale and Kwiatkowski2007; Zabelina & Robinson, Reference Zabelina and Robinson2010). Defocused attention enables some information to “leak in,” whereas focused attention is needed to screen out interfering stimulation for further elaboration (Benedek, Reference Benedek, Jung and Vartanian2018). Similarly to how both exploration and exploitation contribute to creativity at different stages, attentional processes seem to show a similar two-phase dynamic influence on creativity. As proposed above, we suggest that curiosity predominantly shares the broad attentional scope with the first stage of idea generation in creativity.
Notably, previous studies suggested that individuals with attention-deficit/hyperactivity disorder (ADHD) are more creative (White & Shah, Reference White and Shah2006, Reference White and Shah2016). As ADHD is associated with leaky attention and with distraction by irrelevant stimuli (Baird et al., Reference Baird, Smallwood, Mrazek, Kam, Franklin and Schooler2012; Carson et al., Reference Carson, Higgins and Peterson2003; Zabelina, Saporta, & Beeman, Reference Zabelina, Saporta and Beeman2016), the question is how the attentional scope of individuals with ADHD relates to their curiosity (Kidd & Hayden, Reference Kidd and Hayden2015). It is challenging for individuals with ADHD to stay focused and to withhold their response because their likelihood of being attracted by task-irrelevant stimuli is higher. These attentional fluctuations and leaky attention may subserve the link between curiosity and creativity in general. For instance, a certain degree of distractibility was found to improve flexibility in the generation of ideas (Baird et al., Reference Baird, Smallwood, Mrazek, Kam, Franklin and Schooler2012; Carson et al., Reference Carson, Higgins and Peterson2003), and processing of irrelevant stimuli was suggested to expand the associative network, resulting in original combinations of information (Boot, Nevicka, & Baas, Reference Boot, Nevicka and Baas2017). It was suggested that although leaky attention per se may result is some forms of attention disorders and/or psychopathology, high cognitive control would serve as a protective factor, and together with leaky attention, would support creative achievements (Zabelina, Reference Zabelina, Jung and Vartanian2018). In other words, what may seem an attentional deficit of pathological distractibility may in fact subserve curiosity by allowing attention to be directed to possibly irrelevant yet novel information, and creativity as well if subsequent cognitive control is then applied to narrow the scope of attention toward a creative achievement (Zabelina, Reference Zabelina, Jung and Vartanian2018).
2.4 Memory consolidation
Here we propose that creativity and curiosity are further linked through their parallel interaction with memory consolidation. Research shows that curiosity has an influence on consolidation and category-clustering processes. Induced curiosity seems to enhance memory performance (Kang et al., Reference Kang, Hsu, Krajbich, Loewenstein, McClure, Wang and Camerer2009), and people who are highly open to experience and curious show more efficient recall abilities, and generate more associative responses (Christensen et al., Reference Christensen, Kenett, Cotter, Beaty and Silvia2018; Gruber, Gelman, & Ranganath, Reference Gruber, Gelman and Ranganath2014; Kang et al., Reference Kang, Hsu, Krajbich, Loewenstein, McClure, Wang and Camerer2009). Similarly, with regard to creativity, Beaty and Kenett (Reference Beaty and Kenett2020) suggested that individuals who are high in openness to experience have a richer and more interconnected semantic memory. Thus, it seems that by increasing the chance to notice novel information that might be overlooked or coded as irrelevant by others (i.e., defocused attention; Zabelina, Reference Zabelina, Jung and Vartanian2018, or diffused attention), curiosity and openness to experience also support richer consolidation processes. The increased interconnectedness can later subserve creativity, when attempting to generate novel ideas from existing knowledge, in promoting richer and broader associations. For example, it was found that creative writers showcase the ability to use richer sets of representations in a given context (Andreasen & Powers, Reference Andreasen and Powers1975). These findings are in line with the SoM framework, according to which exploration is interlinked to a broad scope of associative thinking, attention, and perception.
Although curiosity improves memory of novel information, creativity intimately relies on the reorganization of existing knowledge (Lewis, Knoblich, & Poe, Reference Lewis, Knoblich and Poe2018) and involves goal-directed memory retrieval (Madore, Thakral, Beaty, Addis, & Schacter, Reference Madore, Thakral, Beaty, Addis and Schacter2019). Previous studies demonstrated that original ideas usually emerge from combining distal domains of knowledge (Miron-Spektor, Gino, & Argote, Reference Miron-Spektor, Gino and Argote2011; Mumford, Baughman, Supinski, & Maher, Reference Mumford, Baughman, Supinski and Maher1996; Simonton, Reference Simonton2003). Furthermore, it was suggested that novel ideas do not emerge ex nihilo but are thought to arise from meaningful variations and recombination of available knowledge (Campbell, Reference Campbell1960). Studies show that during creative tasks, like many other mental activities, task-relevant information is derived from long-term memory (Beaty, Benedek, Silvia, & Schacter, Reference Beaty, Benedek, Silvia and Schacter2016a; Benedek et al., Reference Benedek, Jauk, Fink, Koschutnig, Reishofer, Ebner and Neubauer2014a). Interestingly, the induction of episodic memory (i.e., brief training in recalling details of a recent event) prior to beginning a creativity task was found to enhance creative performance (Madore, Addis, & Schacter, Reference Madore, Addis and Schacter2015).
In the same vein, according to the associative theory of creativity (Mednick, Reference Mednick1962), the specific structure of semantic memory, characterized by “flat” (broader associations) instead of “steep” (few, common associations) associational hierarchies, is necessary for accessing remote concepts and weaker connections. The more remote these concepts are, the more creative the new combination will be. In line with this theory, recent network-science studies (e.g., Baronchelli, Ferrer-i-Cancho, Pastor-Satorras, Chater, & Christiansen, Reference Baronchelli, Ferrer-i-Cancho, Pastor-Satorras, Chater and Christiansen2013; Karuza, Thompson-Schill, & Bassett, Reference Karuza, Thompson-Schill and Bassett2016) suggested that for highly creative people, more distant concepts appear closer in their associative networks. These studies demonstrated that associative networks marked by shorter path lengths and increased interconnectivity between concepts tend to characterize creative thinkers (Benedek et al., Reference Benedek, Kenett, Umdasch, Anaki, Faust and Neubauer2017; Gray et al., Reference Gray, Anderson, Chen, Kelly, Christian, Patrick and Lewis2019; Kenett, Anaki, & Faust, Reference Kenett, Anaki and Faust2014). The short path lengths indicate a faster diffusion of information and smaller distances between concepts with fewer mediating associations (Li, Kenett, Hu, & Beaty, Reference Li, Kenett, Hu and Beaty2021). Successful consolidation of interlinks in memory, as curiosity seems to promote, would enable connecting nodes in a novel manner and the shortening of path lengths in the network, eventually resulting in a more efficient and flexible network (Kenett, Reference Kenett, Kapoula, Volle, Renoult and Andreatta2018). It is important to note here that although research has predominantly focused on semantic networks, it is likely that episodic memory is involved as well (e.g., Madore et al., Reference Madore, Addis and Schacter2015, Reference Madore, Thakral, Beaty, Addis and Schacter2019). More specifically, Duszkiewicz, McNamara, Takeuchi, and Genzel (Reference Duszkiewicz, McNamara, Takeuchi and Genzel2019) suggested that although novel experiences that share some commonality with past ones (“common novelty”) promote semantic memory formation via systems memory consolidation, experiences that bear only a minimal relationship to past experiences (“distinct novelty”) trigger strong initial memory consolidation in the hippocampus, resulting in vivid and long-lasting episodic memories. Curiosity may alter the way episodic experiences are encoded, ultimately changing the way these memories are accessible for subsequent creative combinations.
To conclude, we propose that curiosity influences memory encoding in a way that critically facilitates access to memory during creativity. We further propose that the relationship between curiosity/creativity and memory is bidirectional: Curiosity boosts consolidation of novel information and results in more interconnected memories, and the more interconnected the memory is, the higher the probability of the creative process to occur. A further function for this relationship is associative flexibility (Benedek, Könen, & Neubauer, Reference Benedek, Könen and Neubauer2012b), which uses control processes to shift between contexts and concepts in memory, and reach more distinct and remote networks (Marron et al., Reference Marron, Lerner, Berant, Kinreich, Shapira-Lichter, Hendler and Faust2018), as we describe next.
2.5 Cognitive control: Balancing competing demands
Creativity and curiosity are complex cognitive constructs comprised of competing demands, tensions, and trade-offs. As mentioned above, both contain elements that seem at odds with one another, such as exploration–exploitation, focused–defocused attention, and originality–usefulness, which must be simultaneously accommodated during the process (Lewis, Reference Lewis2000). To manage these inherent tensions, both curiosity and creativity employ cognitive control processes (Benedek & Fink, Reference Benedek and Fink2019; Cervera, Wang, & Hayden, Reference Cervera, Wang and Hayden2020; Chrysikou, Reference Chrysikou, Jung and Vartanian2018, Reference Chrysikou2019), which include updating the content in working memory, shifting between tasks and mental sets, and inhibition functions (Miyake & Friedman, Reference Miyake and Friedman2012). These functions allow performance to be optimized through dynamic adjustments in attention allocation, response selection, and maintenance of task-related goals (Botvinick, Carter, Braver, Barch, & Cohen, Reference Botvinick, Carter, Braver, Barch and Cohen2001).
Studies show that executive abilities optimize both creativity (Beaty & Silvia, Reference Beaty and Silvia2012; Benedek, Jauk, Sommer, Arendasy, & Neubauer, Reference Benedek, Jauk, Sommer, Arendasy and Neubauer2014b) and curiosity (Foley, Kelly, Mhatre, Lopes, & Gottlieb, Reference Foley, Kelly, Mhatre, Lopes and Gottlieb2017; Lau, Ozono, Kuratomi, Komiya, & Murayama, Reference Lau, Ozono, Kuratomi, Komiya and Murayama2020). For example, high working-memory capacity was found to promote the fluency and originality components of creativity by supporting goal maintenance (de Dreu, Nijstad, Baas, Wolsink, & Roskes, Reference de Dreu, Nijstad, Baas, Wolsink and Roskes2012). In line with that, highly creative participants showed higher attentional flexibility by engaging more cognitive control during the attentional switch (Zabelina & Ganis, Reference Zabelina and Ganis2018). Shifting was suggested to allow management of the trade-off between competing interests during curiosity tasks (Cervera et al., Reference Cervera, Wang and Hayden2020) and the selection and implementation of more effective task strategies during creativity tasks (Beaty & Silvia, Reference Beaty and Silvia2012; Benedek, Franz, Heene, & Neubauer, Reference Benedek, Franz, Heene and Neubauer2012a). Interestingly, task switching has recently been found to enhance both divergent and convergent forms of creative thinking, by reducing cognitive fixation, which is characterized by the inability to shift away from an undesired thought or idea (Lu, Akinola, & Mason, Reference Lu, Akinola and Mason2017).
Inhibitory control might benefit both creativity and curiosity by facilitating the suppression of interference from common and inappropriate response tendencies, as was documented in creativity research (Beaty & Silvia, Reference Beaty and Silvia2012; Benedek et al., Reference Benedek, Franz, Heene and Neubauer2012a; Chrysikou, Reference Chrysikou2019), and by deemphasizing the demand for immediate reward in favor of indirect benefits of information, as was suggested in curiosity research (Cervera et al., Reference Cervera, Wang and Hayden2020). Thus, although not yet investigated directly, inhibitory control might suppress affinity to the mundane when surveying our environment curiously similarly to the suppression of mundane ideas in creativity.
In addition to steering and optimization, cognitive control plays a special role in creativity as it is responsible for the evaluation and appraisal of the novelty and usefulness of stimuli and ideas in context, prioritizing information that is task relevant (Chrysikou, Reference Chrysikou2019). An increasing number of studies hold that the evaluation phase plays an important role in the creative process (for a review, see Kleinmintz, Ivancovsky, & Shamay-Tsoory, Reference Kleinmintz, Ivancovsky and Shamay-Tsoory2019). For example, Benedek et al. (Reference Benedek, Nordtvedt, Jauk, Koschmieder, Pretsch, Krammer and Neubauer2016) tested participants' evaluation accuracy in differentiating between common, inappropriate, and creative ideas, and found that accuracy was positively correlated with creative skills.
Some behavioral studies have highlighted the importance of evaluation processes in curiosity as well, suggesting that curiosity relies on the appraisal of one's ability and resources to resolve the challenges raised by the recognition of an information gap (Noordewier & van Dijk, Reference Noordewier and van Dijk2016; Silvia, Reference Silvia2005, Reference Silvia2006). In the context of curiosity, evaluation refers to novelty appraisal (including incongruity, complexity, unexpectedness, obscurity, and uncertainty; Berlyne, Reference Berlyne1960). This is followed by an evaluation of coping potential, such as a person's ability to comprehend a new, complex stimulus (Scherer, Reference Scherer2001; Silvia, Reference Silvia2005). Therefore, stimuli evaluated as novel, yet potentially comprehensible, are experienced as interesting. When evaluation determines that the information is worth pursuing, it is proposed to reinforce the novelty seeking by coupling this information with signals from the dopaminergic system (Lau et al., Reference Lau, Ozono, Kuratomi, Komiya and Murayama2020), a reward mechanism that is further detailed in the next section.
In summary, flexible regulation, supported by cognitive control mechanisms(Hommel, Reference Hommel2015), seems to be crucial for both creativity and curiosity. It allows optimizing performance by managing the multiple tensions that creativity and curiosity are wrought of, as well as alternating between competing strategies. Moreover, cognitive control plays an important role in evaluating stimuli and ideas during the curious/creative process, selecting which information to focus on in curiosity, and which ideas are appropriate in creativity.
2.6 Motivation and reward: The novelty-seeking drive
Although for task-related behaviors the goal of a task is known in advance and can be quantified in terms of extrinsic rewards, the motivation for curiosity (Loewenstein, Reference Loewenstein1994; Oudeyer, Kaplan, & Hafner, Reference Oudeyer, Kaplan and Hafner2007) and creativity (Grant & Berry, Reference Grant and Berry2011; Joy, Reference Joy2004; Kaufman & Beghetto, Reference Kaufman and Beghetto2009) seems to be predominantly intrinsic. Intrinsic motivation reflects an interest in and enjoyment of an activity for the sake of the activity itself, rather than for its instrumental value (Deci, Reference Deci1971; Ryan & Deci, Reference Ryan and Deci2000). One innate motivation for such a strategy is the brain's information-seeking drive that intrinsically urges organisms to explore their environments (Anselme, Reference Anselme2010; Bromberg-Martin & Hikosaka, Reference Bromberg-Martin and Hikosaka2011). It has been suggested that knowledge itself might act as an intrinsic reward (Brydevall, Bennett, Murawski, & Bode, Reference Brydevall, Bennett, Murawski and Bode2018; Charpentier & O'Doherty, Reference Charpentier and O'Doherty2018; Ligneul, Mermillod, & Morisseau, Reference Ligneul, Mermillod and Morisseau2018). Generating creative ideas, just as acquiring novel information, or satisfying our drive to learn something new, activates the reward system and is characterized by an increase in dopamine levels (Takeuchi et al., Reference Takeuchi, Taki, Sassa, Hashizume, Sekiguchi, Fukushima and Kawashima2010). It is therefore possible that high levels of dopamine, which is thought to be the “neuromodulator of exploration,” lower the behavioral threshold for engagement in creative activities (Jauk, Reference Jauk2019) and in information seeking (Bromberg-Martin & Hikosaka, Reference Bromberg-Martin and Hikosaka2009; Gottlieb, Lopes, & Oudeyer, Reference Gottlieb, Lopes and Oudeyer2016; Redgrave, Gurney, & Reynolds, Reference Redgrave, Gurney and Reynolds2008). To explain such behaviors and the high degree of motivation associated with them, it seems necessary to assume that the brain generates intrinsic rewards related to learning or acquiring information (Berlyne, Reference Berlyne1960; Gottlieb et al., Reference Gottlieb, Oudeyer, Lopes and Baranes2013). Studies show that external rewards are not necessary to elicit curiosity-driven behavior, as reward-related dopaminergic circuits can be activated by information independently of extrinsic rewards. For example, human infants naturally explore new environments regardless of physical rewards (Berlyne & Slater, Reference Berlyne and Slater1957; Kreitler, Zigler, & Kreitler, Reference Kreitler, Zigler and Kreitler1984).
Indeed, higher curiosity has been associated with increased activation of reward regions in the brain (Gruber et al., Reference Gruber, Gelman and Ranganath2014; Jepma, Verdonschot, van Steenbergen, Rombouts, & Nieuwenhuis, Reference Jepma, Verdonschot, van Steenbergen, Rombouts and Nieuwenhuis2012; Kang et al., Reference Kang, Hsu, Krajbich, Loewenstein, McClure, Wang and Camerer2009; van Lieshout et al., Reference van Lieshout, Vandenbroucke, Müller, Cools and de Lange2018). Similarly, increased activation of reward regions was also documented during creative problem solving (e.g., Oh, Chesebrough, Erickson, Zhang, & Kounios, Reference Oh, Chesebrough, Erickson, Zhang and Kounios2020), and dopaminergic medications were found to enhance divergent thinking among Parkinson's patients (Faust-Socher et al., Reference Faust-Socher, Kenett, Cohen, Hassin-Baer and Inzelberg2014; Garcia-Ruiz, Castrillo, & Desojo, Reference Garcia-Ruiz, Castrillo and Desojo2019). Associations between dopamine levels and creativity were also reported to have a genetic basis (Murphy, Runco, Acar, & Reiter-Palmon, Reference Murphy, Runco, Acar and Reiter-Palmon2013). In line with this, studies show that novel information induces dopamine release in the hippocampus, triggering memory consolidation and boosting memory persistence (Duszkiewicz et al., Reference Duszkiewicz, McNamara, Takeuchi and Genzel2019; Tulving & Kroll, Reference Tulving and Kroll1995; Wittmann, Bunzeck, Dolan, & Düzel, Reference Wittmann, Bunzeck, Dolan and Düzel2007). It is therefore not unlikely that curiosity as well as creative thinking trigger reward-related neural changes.
Although the positive impact of intrinsic motivation on creativity and curiosity has been highlighted, extrinsic motivation, whenever an activity is performed to attain some separable outcome (Ryan & Deci, Reference Ryan and Deci2000), has been less investigated in this context (Amabile, Reference Amabile1995; Anderson, Potočnik, & Zhou, Reference Anderson, Potočnik and Zhou2014). Nevertheless, given the tensions curiosity and creativity bear, aiming to fill an information gap or seeking a solution to a specific problem both might also involve extrinsic motivation. For example, curiosity was suggested to involve both the intrinsic pleasure of learning as well as extrinsically regulated concerns about the accuracy or fit of newly gathered information (Ryan & Deci, Reference Ryan and Deci2000). Extrinsic motivation has an incremental effect on creativity as well, especially when the reward is contingent on creativity (Eisenberger, Pierce, & Cameron, Reference Eisenberger, Pierce and Cameron1999; Eisenberger & Rhoades, Reference Eisenberger and Rhoades2001). It is reasonable to assume that intrinsic and extrinsic motivation could synergistically benefit creativity and curiosity. Intrinsic motivation may be essential for the novelty component (Amabile, Reference Amabile1993), just like exploratory SoM, whereas extrinsic motivation can help to ensure perseverance and elaboration, similar to exploitatory SoM.
We suggest that the underlying motivation of curiosity and creativity is novelty seeking, and just like the satisfaction of basic biological needs such as thirst or hunger, the hunger for knowledge (or novelty) is satisfying (Biederman & Vessel, Reference Biederman and Vessel2006; Lau et al., Reference Lau, Ozono, Kuratomi, Komiya and Murayama2020; Murayama, Reference Murayama2022). From an evolutionary perspective, rewarding curiosity and creativity may be a long-term investment. Unlike the satisfaction of biological needs, here, the brain invests resources in satisfying mental needs that might turn out to be profitable in the future (Oh et al., Reference Oh, Chesebrough, Erickson, Zhang and Kounios2020).
To summarize, thus far we provided a body of evidence suggesting that shared cognitive mechanisms may link curiosity and creativity. We demonstrated how both are tightly related to SoM and are governed by dynamic trade-offs between exploration and exploitation, which are flexibly tuned by cognitive control. We further demonstrated how curiosity and creativity and their reinforcing dynamics are related to memory and attention. Finally, we postulated that both are driven by a shared entity of novelty seeking.
3. The neuroscience of curiosity and creativity
In this section, we review empirical evidence from neuroscience research, indicating that shared brain regions are involved in both creativity and curiosity. We first discuss dual-process models of human cognition and the role of spontaneous and controlled processes in creativity and curiosity. Then we focus on three major brain networks: the default mode network (DMN), the executive control network (ECN), and the salience network (SN), and their role in curiosity and creativity. Finally, we integrate existing findings to propose modes of interplay between these three networks.
3.1 Spontaneous and controlled processes
What are the processes that may induce us to become curious about a certain stimulus? How are creative ideas generated in our mind? One of the oldest and most ubiquitous approaches in the effort to understand thinking is to distinguish two types of processes that together comprise the broader sphere of cognition (Barr, Beaty, & Seli, Reference Barr, Beaty, Seli, Preiss, Cosmelli and Kaufman2020). Dual-process models of cognition assume two modes of cognitive processing, which are typically named type 1 and type 2 thinking processes (e.g., Evans, Reference Evans2007; Stanovich, Reference Stanovich1999). Type 1 processes are described as automatic, rapid, effortless, nonconscious, and associative in nature, in which a stimulus (either external or internal) elicits associations to relevant information in long-term memory (Benedek & Jauk, Reference Benedek, Jauk, Fox and Christoff2018). Type 2 processes are described as controlled, analytic, slow, conscious, and effortful, and are related to working-memory processing (Kahneman, Reference Kahneman2011).
Other models classify thinking as either spontaneous or controlled (Christoff, Irving, Fox, Spreng, & Andrews-Hanna, Reference Christoff, Irving, Fox, Spreng and Andrews-Hanna2016). Spontaneous cognition (also referred to as undirected or self-generated thinking; Christoff, Reference Christoff2012) can be defined as thinking processes that are stimulus-independent or are non-deliberative and less prone to conscious guidance. This mode involves rapid information retrieval from episodic and semantic memory that is mostly unconscious (Sowden, Pringle, & Gabora, Reference Sowden, Pringle and Gabora2015) and is typically linked to the DMN (e.g., Fox & Christoff, Reference Fox, Christoff, Fleming and Frith2014). On the other hand, controlled cognition (also referred to as goal-directed thought), refers to thoughts that are stimulus-driven or deliberative and is guided by top-down processes (Marron & Faust, Reference Marron and Faust2019), is typically ascribed to the ECN (e.g., Seeley et al., Reference Seeley, Matthews, Crawford, Gorno-Tempini, Foti, Mackenzie and Miller2008). This mode requires cognitive control for maintaining task focus, developing mental strategies, task monitoring, and evaluating outcomes (Benedek & Jauk, Reference Benedek, Jauk, Fox and Christoff2018; Chrysikou, Reference Chrysikou2019).
Dual-process models have a long tradition in creativity research (for a recent review, see Sowden et al., Reference Sowden, Pringle and Gabora2015), yet these models are lacking in curiosity research. Traditionally, creativity is depicted by a two-staged process (Basadur, Reference Basadur1995; Martindale, Reference Martindale and Sternberg1999; Mayseless, Aharon-Peretz, & Shamay-Tsoory, Reference Mayseless, Aharon-Peretz and Shamay-Tsoory2014; Sowden et al., Reference Sowden, Pringle and Gabora2015): the generation phase, where a combination of remote associations is activated in a novel manner, and a subsequent evaluation phase, in which ideas are logically valued. Spontaneous and controlled processes are associated with generative and evaluative functions, respectively, which are thought to interact in the forging of creative ideas (Benedek & Jauk, Reference Benedek, Jauk, Fox and Christoff2018). In line with this, Guilford (Reference Guilford1956) originally distinguished between convergent and divergent thinking, whereby although divergent thinking underlies the generation phase, convergent thinking guides goal-directed reasoning and related mental operations, and involves enhanced evaluative processes (see Zhang, Sjoerds, & Hommel, Reference Zhang, Sjoerds and Hommel2020 for the different neurocognitive mehcanisms involved in each).
Theoretical accounts of creative cognition, hence, acknowledge the relevance of both spontaneous and controlled processes, which can be loosely mapped to the conceptualizations of type 1 and type 2 thinking, or undirected and goal-directed thought (Benedek & Jauk, Reference Benedek, Jauk, Fox and Christoff2018). Accordingly, we suggest that curiosity can be conceptualized using similar terms: We propose that curiosity first begins with divergent and spontaneous exploration of novel information in a nonrestrictive manner, just like we spontaneously generate as many ideas as we can in creativity. Subsequently, a more controlled selection is activated, guided by our estimation of relevance and interest, resulting in convergence to a certain stimulus to be examined more closely and possibly stored in memory. We propose that spontaneous and controlled modes of cognitive processing, mediated by DMN and ECN, underlie the trade-offs between exploration and exploitation and between defocused and focused attention, respectively, interchanging during the processes of curiosity and creativity. We further postulate that the dynamic shift between the networks is governed by the SN, which functions as a switchboard. Evidence for the involvement of these brain networks in creative thought and curiosity is reviewed next.
3.2 DMN: The initiation of novelty seeking
What are the neural mechanisms and dynamics driving affinity toward the novel? We suggest that novelty detection or generation is triggered by expectancy violation and involve spontaneous thoughts that are manifested by novel neuronal firing. In other words, novel ideas and new information elicit a neural activity pattern that has never occurred before and results in a cascade of neural responses across the novelty network (Kafkas & Montaldi, Reference Kafkas and Montaldi2018; Murty, Ballard, Macduffie, Krebs, & Adcock, Reference Murty, Ballard, Macduffie, Krebs and Adcock2013). This unique pattern precedes the initiation of curiosity and creativity and is manifested by a slow uprising phase of a spontaneous fluctuation in cortico-hippocampal circuits. This is followed by low-level activation spread in relevant networks generated by any new content (Noy et al., Reference Noy, Bickel, Zion-Golumbic, Harel, Golan, Davidesco and Malach2015) and ends with dopamine release in the hippocampus (Duszkiewicz et al., Reference Duszkiewicz, McNamara, Takeuchi and Genzel2019; Wittmann et al., Reference Wittmann, Bunzeck, Dolan and Düzel2007).
Slow anticipatory buildup of spontaneous fluctuations, also previously known as “readiness potential” (Kornhuber & Deecke, Reference Kornhuber and Deecke1965), has recently been suggested as the driving mechanism of the entire range of voluntary behaviors (Moutard, Dehaene, & Malach, Reference Moutard, Dehaene and Malach2015) and more specifically was observed prior to creative idea generation (Broday-Dvir & Malach, Reference Broday-Dvir and Malach2021). When such a fluctuation crosses the activation threshold, a spontaneous mental event can emerge (Norman, Raccah, Liu, Parvizi, & Malach, Reference Norman, Raccah, Liu, Parvizi and Malach2021). Using this account, being in a novelty-seeking mode raises the neural activity above the decision threshold and explains how certain stimulation can attract our attention and be perceived as novel and interesting. Thus, we suggest that large-scale ongoing spontaneous (also termed resting state) activity throughout the cortex plays a role in novelty detection.
The primary neural network that has been suggested to sustain both spontaneous thought (Andrews-Hanna, Smallwood, & Spreng, Reference Andrews-Hanna, Smallwood and Spreng2014; Beaty et al., Reference Beaty, Benedek, Silvia and Schacter2016a; Buckner, Andrews-Hanna, & Schacter, Reference Buckner, Andrews-Hanna and Schacter2008; Jung, Mead, Carrasco, & Flores, Reference Jung, Mead, Carrasco and Flores2013; Raichle et al., Reference Raichle, MacLeod, Snyder, Powers, Gusnard and Shulman2001) and associative and predictive processing (Bar, Aminoff, Mason, & Fenske, Reference Bar, Aminoff, Mason and Fenske2007; Stawarczyk, Bezdek, & Zacks, Reference Stawarczyk, Bezdek and Zacks2021) is the default mode network (DMN). The DMN includes the medial prefrontal cortex (mPFC), the posterior cingulate cortex (PCC/precuneus), and the bilateral inferior parietal lobes (IPL; Gusnard & Raichle, Reference Gusnard and Raichle2001). These areas have been reported to be involved in information integration (Binder, Desai, Graves, & Conant, Reference Binder, Desai, Graves and Conant2009), memory retrieval (Ciaramelli, Grady, & Moscovitch, Reference Ciaramelli, Grady and Moscovitch2008; Sugiura, Shah, Zilles, & Fink, Reference Sugiura, Shah, Zilles and Fink2005), mind-wandering (Christoff, Gordon, Smallwood, Smith, & Schooler, Reference Christoff, Gordon, Smallwood, Smith and Schooler2009; Mason et al., Reference Mason, Norton, van Horn, Wegner, Grafton and Macrae2007), generation of associative predictions (Bar et al., Reference Bar, Aminoff, Mason and Fenske2007; Baror, Aminoff, & Bar, Reference Baror, Aminoff, Bar, Gilead and Ochsner2021), and bottom-up processing (Cabeza, Ciaramelli, Olson, & Moscovitch, Reference Cabeza, Ciaramelli, Olson and Moscovitch2008). The DMN demonstrates increased activation during a resting state (Gusnard & Raichle, Reference Gusnard and Raichle2001) and is thought to support cognitive processes that draw upon stored episodic or semantic knowledge (Andrews-Hanna, Reidler, Sepulcre, Poulin, & Buckner, Reference Andrews-Hanna, Reidler, Sepulcre, Poulin and Buckner2010), such as the generation of novel conceptual combinations (Volle, Reference Volle, Jung and Vartanian2018; Zabelina & Andrews-Hanna, Reference Zabelina and Andrews-Hanna2016). Thus, the DMN constitutes a plausible candidate for mediating novelty-seeking initiation.
Indeed, activation of key regions of the DMN was repeatedly found during the generation of creative ideas (e.g., Beaty et al., Reference Beaty, Benedek, Wilkins, Jauk, Fink, Silvia and Neubauer2014, Reference Beaty, Benedek, Silvia and Schacter2016a; Gonen-Yaacovi et al., Reference Gonen-Yaacovi, de Souza, Levy, Urbanski, Josse and Volle2013; Ivancovsky, Kleinmintz, Lee, Kurman, & Shamay-Tsoory, Reference Ivancovsky, Kleinmintz, Lee, Kurman and Shamay-Tsoory2018; Jung et al., Reference Jung, Mead, Carrasco and Flores2013; Mayseless, Eran, & Shamay-Tsoory, Reference Mayseless, Eran and Shamay-Tsoory2015). Furthermore, activation of the DMN, together with the hippocampus, is thought to be involved in the production of novel combinations of associations (Bar et al., Reference Bar, Aminoff, Mason and Fenske2007; Beaty et al., Reference Beaty, Benedek, Silvia and Schacter2016a; Bendetowicz, Urbanski, Aichelburg, Levy, & Volle, Reference Bendetowicz, Urbanski, Aichelburg, Levy and Volle2017; Madore et al., Reference Madore, Thakral, Beaty, Addis and Schacter2019). Although less investigated, several neuroimaging studies point to the involvement of the DMN in curiosity as well (Li et al., Reference Li, Huo, Zhuang, Song, Wang, Ren and Qiu2019). For example, van Lieshout et al. (Reference van Lieshout, Vandenbroucke, Müller, Cools and de Lange2018) found that curiosity was related to increased activity in the IPL. In line with our hypothesis, it was suggested that during curious state, prior knowledge and novel information are integrated by the mPFC (Ligneul et al., Reference Ligneul, Mermillod and Morisseau2018). Further exploration of the involvement of the DMN in curiosity may be fruitful in unlocking the neural correlates of curiosity, illuminating the neural circumstances associated with the state of being curious in which novel knowledge is more readily obtained.
Interestingly, associations between DMN and openness to experience were found in several studies (e.g., Adelstein et al., Reference Adelstein, Shehzad, Mennes, DeYoung, Zuo, Kelly and Castellanos2011; Beaty et al., Reference Beaty, Kaufman, Benedek, Jung, Kenett, Jauk and Silvia2016b; DeYoung, Reference DeYoung, Mikulincer, Shaver, Cooper and Larsen2015). For example, Beaty et al. (Reference Beaty, Kaufman, Benedek, Jung, Kenett, Jauk and Silvia2016b) demonstrated that openness predicts increased DMN efficiency, which is thought to reliably measure the network integrity, and considered to reflect efficiency in information processing in the network. In this context, the ability to engage the neurocognitive resources of the DMN efficiently may account for the tendency of highly open individuals to be drawn to novel stimuli. Taken together, we propose that the initiation of creativity and curiosity is manifested by a spontaneous activity in networks such as the DMN which is later evolve to a unique neural firing in subsequent networks.
3.3 SN: Evaluation and selection of new information
After attending to the novel, we evaluate the relevance of the detected novel information. The evaluation of potential relevance does not directly map onto cognitive control but is rather a complex process that also relies on valance-valuation judgments ascribed to the SN (Kleinmintz et al., Reference Kleinmintz, Ivancovsky and Shamay-Tsoory2019). The SN includes the bilateral anterior cingulate cortex (ACC) and the anterior insula (AN), which are associated with assigning valance to relevant stimuli (Uddin, Reference Uddin2015). The SN has close connections with regions involved in emotion and motivation, such as the amygdala, the ventral tegmental area (VTA), and the caudate (Menon, Reference Menon and Toga2015). Thus, the SN is thought to determine whether the stimulus will be approached or avoided, and what emotional valance is associated with it (Barford, Fayn, Silvia, & Smillie, Reference Barford, Fayn, Silvia and Smillie2018; Chrysikou, Reference Chrysikou, Jung and Vartanian2018; Xia, Touroutoglou, Quigley, Feldman Barrett, & Dickerson, Reference Xia, Touroutoglou, Quigley, Feldman Barrett and Dickerson2017). It has been suggested that fronto-insular nodes function as iterative “relevance detectors” of both external and internal information (perceptual and episodic information, respectively), which if deemed relevant becomes available in working memory for ECN processing (Chrysikou, Reference Chrysikou, Jung and Vartanian2018). Therefore, the SN may contribute to the detection of relevant stimuli (Uddin, Reference Uddin2015) within the DMN, such as candidate ideas in creativity (Jung et al., Reference Jung, Mead, Carrasco and Flores2013) and novel information in curiosity (Li et al., Reference Li, Huo, Zhuang, Song, Wang, Ren and Qiu2019).
Induction of curiosity was found to activate the AN and the ACC (Jepma et al., Reference Jepma, Verdonschot, van Steenbergen, Rombouts and Nieuwenhuis2012). Insular activity was also found to relate to curiosity relief (i.e., the sense that curiosity was satisfied), as activity there seems to increase linearly with the amount of information gained after curiosity was satisfied with the desired information (van Lieshout et al., Reference van Lieshout, Vandenbroucke, Müller, Cools and de Lange2018). The insula is also engaged in various creative processes, such as creative idea production, divergent thinking, and visual creativity (Beaty, Benedek, Barry Kaufman, & Silvia, Reference Beaty, Benedek, Barry Kaufman and Silvia2015; Wu et al., Reference Wu, Miller, Adhimoolam, Zackey, Khan, Ketelle and Miller2015). For example, increased gray matter volume in the insula was suggested to be associated with higher creativity (Takeuchi et al., Reference Takeuchi, Taki, Sassa, Hashizume, Sekiguchi, Fukushima and Kawashima2010).
Considering that both curiosity and creativity seem to be driven by novelty, we suggest that the SN determines the threshold for them to be triggered. Similar to the perception of signals and in line with the signal detection theory in psychophysics (e.g., Peterson, Birdsall, & Fox, Reference Peterson, Birdsall and Fox1954; Tanner & Swets, Reference Tanner and Swets1954), it is possible that the more curious and creative you are, the higher your sensitivity to novel stimuli would be; thus, more candidate information will cross the threshold for further elaboration. We suggest here that SN activity regulates the threshold for novelty detection.
3.4 ECN: Top-down monitoring and evaluation of novelty
It has been suggested that actions that are associated with new information require control, whereas actions that have low uncertainty are habitual and can be performed with little control-related resources (Cavanagh & Frank, Reference Cavanagh and Frank2014; Fan, Reference Fan2014). Under this claim, curiosity and creativity, which both involve engagement with novelty, are predicted to require controlled processing. The ECN is a vast, frontal network known to involve several control-related processes. For example, inhibition is mediated by the ventro-lateral prefrontal cortex (vlPFC, also known as the inferior frontal gyrus; IFG), working memory is mediated by the dorsolateral prefrontal cortex (dlPFC), error detection is related to the medial prefrontal cortex (mPFC), updating is associated with the temporoparietal junction (TPJ), and switching involves the interaction between several fronto-parietal regions (Chrysikou, Reference Chrysikou, Jung and Vartanian2018, Reference Chrysikou2019; Kleinmintz et al., Reference Kleinmintz, Ivancovsky and Shamay-Tsoory2019). It was further suggested that increased ECN activity can support response combination and selection by evaluating the novelty of the generated responses (Chrysikou, Reference Chrysikou2019). Interestingly, during evaluation of novelty, higher activations were found in the ECN, as opposed to evaluation of appropriateness, in which increased activity was found in regions related to memory, emotion, and motivation (Huang, Tang, Sun, & Luo, Reference Huang, Tang, Sun and Luo2018), suggesting that evaluation of novelty is related to cognitive control processes, whereas the evaluation of appropriateness may be related to different mechanisms (Sowden et al., Reference Sowden, Pringle and Gabora2015).
Curiosity might also involve regions associated with information evaluation required to accomplish its functions (Tian et al., Reference Tian, Silva and Liu2021). Indeed, some initial evidence demonstrates how the ECN might play a role in curiosity as well. Kang et al. (Reference Kang, Hsu, Krajbich, Loewenstein, McClure, Wang and Camerer2009) found that activity in the caudate and in the IFG during a trivia task was associated with self-reported curiosity. The key analyses focused on activations during the anticipatory period after participants had received a question, but before they were given the answer. The anticipatory period parallels the evaluation phase to some degree, as participants had to assign a value to the question and evaluate their interest in finding out the answer.
The involvement of cognitive control in creativity is studied to a greater extent than the involvement of cognitive control in curiosity but its role is debatable. Neuroscientific evidence contributes to this ongoing discussion. Activation across regions in the ECN was found during various types of creative thought such as divergent thinking tasks, musical improvisation, and creative problem solving (Chen, Beaty, & Qiu, Reference Chen, Beaty and Qiu2020; Jung et al., Reference Jung, Mead, Carrasco and Flores2013; Perchtold et al., Reference Perchtold, Papousek, Koschutnig, Rominger, Weber, Weiss and Fink2018). Overall, ECN recruitment appears to be a function of whether creative cognition is constrained to meet task-specific goals (Beaty et al., Reference Beaty, Benedek, Silvia and Schacter2016a). For example, it was found that under conditions of high semantic constraints, in which one is primed with mundane rather than creative associations, the ability to nonetheless perform creative production of associations is related to connectivity patterns between ECN regions and DMN regions (Beaty, Christensen, Benedek, Silvia, & Schacter, Reference Beaty, Christensen, Benedek, Silvia and Schacter2017). Nevertheless, lesion studies and neurostimulation studies found that decreased activity of the IFG, which results in reduced inhibitory control, seems to lead to increased creative production (Mayseless et al., Reference Mayseless, Aharon-Peretz and Shamay-Tsoory2014; Miller, Ponton, Benson, Cummings, & Mena, Reference Miller, Ponton, Benson, Cummings and Mena1996, Reference Miller, Boone, Cummings, Read and Mishkin2000; Seeley et al., Reference Seeley, Matthews, Crawford, Gorno-Tempini, Foti, Mackenzie and Miller2008).
It is important to note that inhibition in the creative process pertains to the suppression of mundane ideas in favor of remote and original ones. Inhibition that constricts the scope of our semantic search to begin with would be detrimental for creativity. We propose that applying inhibitory control in a creativity-facilitating manner would require executive functions. As suggested elsewhere (Benedek & Fink, Reference Benedek and Fink2019), a slight reduction of cognitive control may support certain creative performances, by adjusting the balance of controlled and spontaneous processes toward preferable conditions for the given task (Benedek & Jauk, Reference Benedek, Jauk, Fox and Christoff2018; Chrysikou, Weber, & Thompson-Schill, Reference Chrysikou, Weber and Thompson-Schill2014). In line with our assumption, direct investigation of the involvement of cognitive control during the creative process by neuroscientific means would shed more light on its debatable role, as the contribution of ECN might change across the different phases of the creative process.
Interestingly, highly creative participants showed increased coupling of DMN regions with the ECN during idea generation as well as other creative activities such as musical improvisation and poetry generation (Beaty et al., Reference Beaty, Benedek, Silvia and Schacter2016a). This connectivity pattern points to a mechanism where controlled and spontaneous cognitive processes interact in creative cognition (Benedek & Fink, Reference Benedek and Fink2019; Zabelina & Andrews-Hanna, Reference Zabelina and Andrews-Hanna2016). In this context, although the DMN may provide self-generated information via episodic retrieval, the ECN directs and monitors the integration of this information (Beaty, Seli, & Schacter, Reference Beaty, Seli and Schacter2019). Similar to the interaction between these processes in creativity, it is reasonable to expect that cooperation between the DMN and ECN would benefit curiosity as well. Future studies should directly examine the role of cognitive control in curiosity and its interaction with the DMN.
Taken together, we suggest that cognitive control plays a key “gatekeeper” role both in curiosity and in creativity. Nevertheless, although some cognitive control is needed to attend novel stimuli around us as well as generate novel ideas, excessive control might hinder creativity (Runco & Basadur, Reference Runco and Basadur1993) and curiosity. The brain network dynamics, described next, determine the balance between spontaneous and controlled processes during curiosity and creativity, ultimately governing their maturation.
3.5 Brain network dynamics
It has previously been suggested that the interaction between the DMN and ECN reflects goal-directed, self-generated cognition, with the DMN involved in generative processes and the ECN in guiding, constraining, and modifying these processes to meet task goals (Beaty et al., Reference Beaty, Benedek, Silvia and Schacter2016a; Ellamil, Dobson, Beeman, & Christoff, Reference Ellamil, Dobson, Beeman and Christoff2012; Mok, Reference Mok2014). The interplay between the networks may reflect the cyclic motion between spontaneous and controlled processes during the curiosity/creativity process.
The SN is thought to be sensitive to bottom-up salience (Abraham, Reference Abraham2019) and to mediate interactions between internally and externally oriented attention (Menon & Uddin, Reference Menon and Uddin2010), and it is therefore possible that it is involved in tuning the balance between the DMN and ECN during the curiosity or creativity processes, depending on what is necessary in the current context (Abraham, Reference Abraham2019). In fact, the SN has been shown to moderate DMN–ECN interactions during creative idea production (Beaty et al., Reference Beaty, Christensen, Benedek, Silvia and Schacter2017; Goulden et al., Reference Goulden, Khusnulina, Davis, Bracewell, Bokde, McNulty and Mullins2014), and high functional connectivity between the three networks was found among highly creative individuals (Beaty et al., Reference Beaty, Benedek, Silvia and Schacter2016a). Similarly, curiosity was found to be related to the functional connectivity of these three networks (Li et al., Reference Li, Huo, Zhuang, Song, Wang, Ren and Qiu2019).
In sum, creativity and curiosity are proposed to arise from the interplay between spontaneous and controlled processes, expressed by coupling of the DMN, ECN, and SN. Each of these networks is ascribed to a different and essential aspect of both. The DMN contributes to the integration of existing knowledge with novel information through flexible retrieval and search processes. The SN is thought to filter useful and novel candidate information and forward it to the ECN that evaluates and constrains this stream toward a specific goal by using flexible inhibition mechanisms. The dynamic interaction between these networks, together with subsequent memory and reward circuits, may postulate the underlying mechanism of curiosity and creativity. In the next section, we integrate the neurocognitive evidence from both fields into a unified model of novelty seeking.
4. The novelty-seeking model (NSM)
Based on the literature reviewed above, we propose a unified model that can encompass both curiosity and creativity, pointing to a similar involvement of generative and evaluative processes in both. This is in line with the dual-process models described above (e.g., Sowden et al., Reference Sowden, Pringle and Gabora2015). The proposed NSM is characterized by four distinct phases: affinity, activation, evaluation, and commitment (Fig. 1). These same four phases are proposed to be at the basis of creativity, curiosity, and all other novelty-seeking behaviors: First, attraction to a stimulus or a problem. Second, activation of the mental operations required to pursue it. Third, an iterative process of evaluating merit and relevance, and fourth, consolidation of what has crossed the threshold of interest in memory. We elaborate on each of these phases, after we describe our working terminology first.
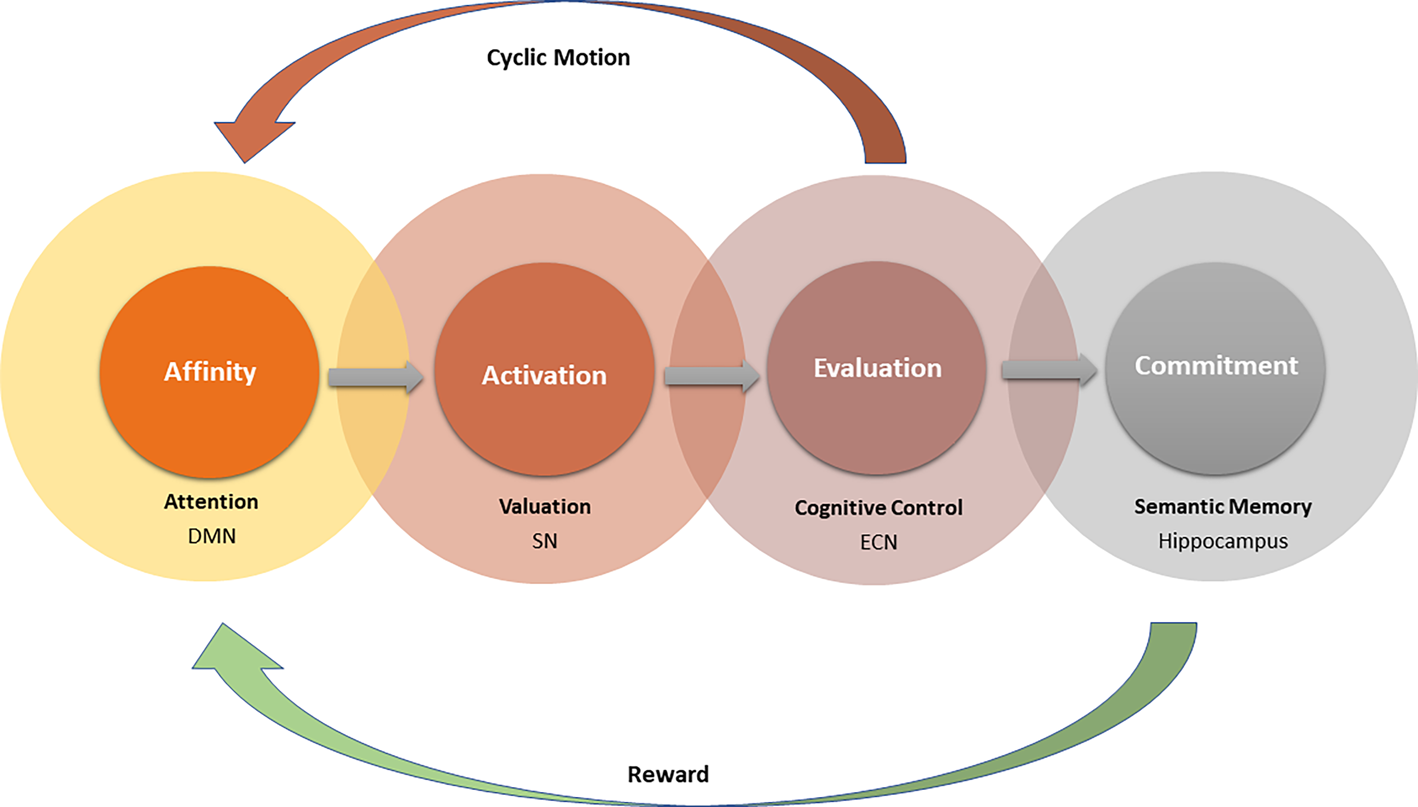
Figure 1. Four-phase model of the NSM. The process is initiated by an affinity toward novel internal or external stimulation and results in generation of novel combinations in a semantic network. The affinity phase relies on DMN activation as well as on attentional orientation. The candidate combinations potentially cross a relevance threshold in the activation phase, based on their saliency. This is followed by more deliberate and controlled processes in the evaluation phase, as each selected combination is being assessed. The process ends with commitment to memory, as a novel interlink is created in the semantic network, and then rewarded by the dopaminergic system.
Affinity: Any act of curiosity or creativity starts with affinity to a certain stimulation. Stimulation is the catalyst that triggers the process, be it an external stimulus or an internal idea. This affinity may be driven internally following a goal-directed search, such as an attempt to find some solution, or externally through a spontaneous encounter with something that attracts our attention and violates previous expectations. Affinity to stimulation depends on openness (SoM) and on the availability of mental resources. Because this phase relies on spontaneous processes and on search processes through semantic and autobiographical memory, among others, it is presumed to be mediated by the DMN (Beaty et al., Reference Beaty, Benedek, Silvia and Schacter2016a; Benedek et al., Reference Benedek, Schües, Beaty, Jauk, Koschutnig, Fink and Neubauer2018; Madore et al., Reference Madore, Thakral, Beaty, Addis and Schacter2019). We further suggest that affinity is expressed in curiosity as a unique scan path of the external environment (Krajbich, Lu, Camerer, & Rangel, Reference Krajbich, Lu, Camerer and Rangel2012), and in creativity as a unique pattern of “random walks” (Abbott, Austerweil, & Griffiths, Reference Abbott, Austerweil and Griffiths2015, or shorter path length; Kenett et al., Reference Kenett, Anaki and Faust2014) in the associative network. In curiosity, this is the stage by which the scope of attention directs the search toward the novel (Gottlieb, Cohanpour, Li, Singletary, & Zabeh, Reference Gottlieb, Cohanpour, Li, Singletary and Zabeh2020). In creativity, this is the phase where the generation of novel stimulation is initiated, creating unique combinations in mind by integrating existing knowledge with novel internal or external information. The integration of external and internal stimuli is supported by inner–outer attentional fluctuations (Kucyi, Hove, Esterman, Matthew Hutchison, & Valera, Reference Kucyi, Hove, Esterman, Matthew Hutchison and Valera2017), which are suggested to take place in the DMN during the affinity phase.
Activation: The affinity phase ends with a set of prioritized representations, which initiates the spread of activation in the semantic network (the spreading-activation theory; Collins & Loftus, Reference Collins and Loftus1975). This spreading continues until an intersection between two concept nodes is found and a novel combination is generated in the network. Combination is the integration of existing knowledge with novel internal or external information. In creativity we combine remote associations in a novel manner, whereas in curiosity the combinations are made between prior knowledge and a novel stimulus to satisfy an information gap. In the activation phase, new combinations potentially cross a relevance threshold, depending on their saliency. Saliency is determined by various aspects such as novelty (Foley, Jangraw, Peck, & Gottlieb, Reference Foley, Jangraw, Peck and Gottlieb2014), personal relevance (e.g., goal, experience), SoM, perceptual saliency (e.g., brightness or color), or emotional saliency (e.g., a crying baby or a car accident). If the process is initiated voluntarily, the relevance threshold would reflect the level of interest, and if it is goal-directed, the threshold would reflect the goodness of fit between the stimulation and the goal.
The activation phase generates combinations, and those that cross the threshold are candidates for evaluation. We suggest that the activation phase is mediated by the salience network (SN), which determines the threshold and is responsible for the switching between the generative and evaluative processes (Abraham, Reference Abraham2019). The novelty of the stimulation is the key element according to which the combinations are evaluated and selected in later stages, together with its usefulness.
Evaluation: Each combination that has crossed the threshold is evaluated for its originality and usefulness. As opposed to the last two phases, evaluation is a systematic and deliberate process, using top-down information to examine the goodness of fit of the candidate combinations to the initial goal. There is a cyclic motion between the generation and the evaluation of combinations such that, although common or deviant combinations are rejected, novel and appropriate combinations receive further examination and elaboration. This phase is crucial for both curiosity and creativity as it enables us to narrow down the candidate combinations and to deeply explore them one by one. The evaluation phase relies on the valuation, monitoring, and selection of combinations, and these are commonly viewed as cognitive control processes (Benedek et al., Reference Benedek, Schües, Beaty, Jauk, Koschutnig, Fink and Neubauer2018; Chrysikou, Reference Chrysikou, Jung and Vartanian2018; Ellamil et al., Reference Ellamil, Dobson, Beeman and Christoff2012; Kleinmintz et al., Reference Kleinmintz, Ivancovsky and Shamay-Tsoory2019). The involvement of cognitive control is essential for preventing overbursts of stimulation and to efficiently direct the available resources. Otherwise, no commitment through consolidation takes place.
Commitment: The process results, if the novel information is deemed worthy, in consolidation and categorization. Unlike previous models, we suggest that the NSM continues beyond the evaluation phase and ends with commitment to memory. The commitment phase is expressed by consolidating a novel combination in memory, that is, integration of novel stimuli into preexisting representations and thus the creation of new representations (Ranganath & Rainer, Reference Ranganath and Rainer2003; Schomaker & Meeter, Reference Schomaker and Meeter2015). This is more than mere consolidation; we commit to a combination we generated in previous stages, willing to invest our mental resources in further learning and elaboration. In curiosity, we commit to consolidating novel information in memory, whereas in creativity, we commit to the novel idea. The commitment phase ends with a call for action – elaboration and execution of an idea or further exploration of the novel stimulus. The process results in broadening of the associative network and scope of thinking and is mediated by the hippocampus. Hippocampal activity is reinforced by reward mechanisms that raise the chances for its persistence and reoccurrence (Duszkiewicz et al., Reference Duszkiewicz, McNamara, Takeuchi and Genzel2019; Kafkas & Montaldi, Reference Kafkas and Montaldi2018). The richer the memory structure is, the higher the efficacy of finding and establishing novel combinations (Gray et al., Reference Gray, Anderson, Chen, Kelly, Christian, Patrick and Lewis2019). We, therefore, propose that the NSM originates and ends in memory.
5. The NSM within the context of dynamic changes in SoM
The novelty-seeking process contains inherent tensions, such as exploration–exploitation, focused–defocused attention, and originality–usefulness, which influence the form that curiosity and creativity will take. Here, we account for these trade-offs by linking the different types of curiosity and creativity with the dynamic and overarching SoM (Herz et al., Reference Herz, Baror and Bar2020) that shapes them.
5.1 Different types of creativity and curiosity and their attribution to SoM
It is increasingly acknowledged that curiosity is comprised of two types: feelings of general interest (also known as diversive curiosity), which motivate diverse exploration, and feelings of deprivation (also known as specific curiosity), which promote specific exploration and aim to solve specific problems (Litman, Reference Litman2008). Although interest is motivated by positive feelings and by the opportunity to learn something new, deprivation, on the other hand, can be seen as a need that drives us to reduce the feeling of uncertainty by acquiring missing information (Litman, Reference Litman2008). Following Litman's discrimination, it is important to note that on the continuum of interest–deprivation, deprivation may arise out of a need to fill the gap and to reduce uncertainty, whereas interest is less related to anxiety and more to creativity. As proposed above, we hypothesize that in an exploratory SoM, curiosity would expand the scope of our stream of thoughts and enable creativity to arise in an uncertain environment. On the other hand, in an exploitatory SoM, the same information might be perceived as overwhelming and invoke anxiety, so curiosity will take the form of deprivation, promoting narrow and specific exploration, thus leaving no room for new ideas to arise. This hypothesis is partially supported by a recent study (Lauriola et al., Reference Lauriola, Litman, Mussel, de Santis, Crowson and Hoffman2015) that showed a positive correlation between interest, fun seeking, positive expectancies, and risk taking, which orients participants toward exploration, and vice versa. Exploitation was positively correlated with thoughtful evaluation and concern over negative outcomes and potential risks, orienting participants toward caution regarding knowledge search.
Creativity, as well, involves two modes of thinking: convergent and divergent. Convergent thinking (CT) refers to a single solution to a given problem, as opposed to divergent thinking (DT), which refers to the extrapolation of many possible responses to an initial stimulus (Guilford, Reference Guilford1967). DT requires combining information in novel ways, linking remote associates or transforming information into unexpected forms, CT involves manipulation of existing knowledge by applying more logical search, recognition, and decision-making techniques to derive the best solution (Cropley, Reference Cropley2006). These two modes of thinking are necessary to produce novel ideas and are shifted during the creative process, as described above (Guilford, Reference Guilford1956).
We suggest that the subtypes of curiosity and creativity are parallel: Open-ended novelty-seeking curiosity is similar to DT, as multiple options are acceptable. Goal-directed novelty-seeking curiosity, on the other hand, includes the specific/deprivation type, which is similar to CT when the focus is on a single option in a predetermined task. Some support for this notion comes from a recent study that measured these subtypes of curiosity and creativity by using lab-based tasks (Koutstaal, Kedrick, & Gonzalez-Brito, Reference Koutstaal, Kedrick and Gonzalez-Brito2022). The authors found a positive correlation between the novelty of the questions that participants generated during a curiosity Q&A task and the originality of their responses on a DT task. Notably, performance in both tasks was positively correlated with the trait-based interest type but not with the deprivation type. On the other hand, the extent to which participants sought out missing information regarding the presented stimuli in the curiosity task (referred to by the authors as “gap-related information foraging”) positively correlated with CT tasks. In line with that, in a recent meta-analysis, that examined the relationship between the two (Schutte & Malouff, Reference Schutte and Malouff2019), the authors suggested that the exploration dimension of curiosity may be more relevant for creativity than the deprivation sensitivity dimension.
Another important distinction that should be made is between the two components of creative ideas, that is, originality and usefulness (Amabile, Reference Amabile1993; Stein, Reference Stein1953; Sternberg & Lubart, Reference Sternberg, Lubart and Sternberg1999). Although both are essential for creativity (otherwise, ideas would be either non-original or inappropriate), they require different abilities. Originality refers to breaking existing frames, whereas usefulness refers to the practical qualities, acceptance by other people, and adherence to cultural norms (Morris & Leung, Reference Morris and Leung2010). It is noteworthy that creative people have been found to excel in both dimensions of creativity (Miron-Spektor & Erez, Reference Miron-Spektor and Erez2017) and that originality and usefulness are also interdependent, reinforcing each other during the creative process (Lewis, Reference Lewis2000). According to Miron-Spektor and Erez (Reference Miron-Spektor and Erez2017), “Novel insights can help improve product usefulness, and considering usefulness issues can inspire novel ideas” (p. 4). Thus, to produce a creative solution, problem solvers must also find a way to manage usefulness after establishing novelty (Amabile, Reference Amabile1996; Berg, Reference Berg and Berg1991; Litchfield, Reference Litchfield2008).
We suggest that the close relationship between these different types of curiosity and creativity are best captured by the SoM framework (Fig. 2). Although exploration results in diversity, exploitation results in specificity. Therefore, DT, diversive curiosity, and originality orientation would be related to exploration, whereas CT, specific curiosity, and usefulness would be associated with exploitation. It is important to note that one is rarely at either extreme of this SoM continuum, but rather dynamically alters between the different modes based on situational demands.
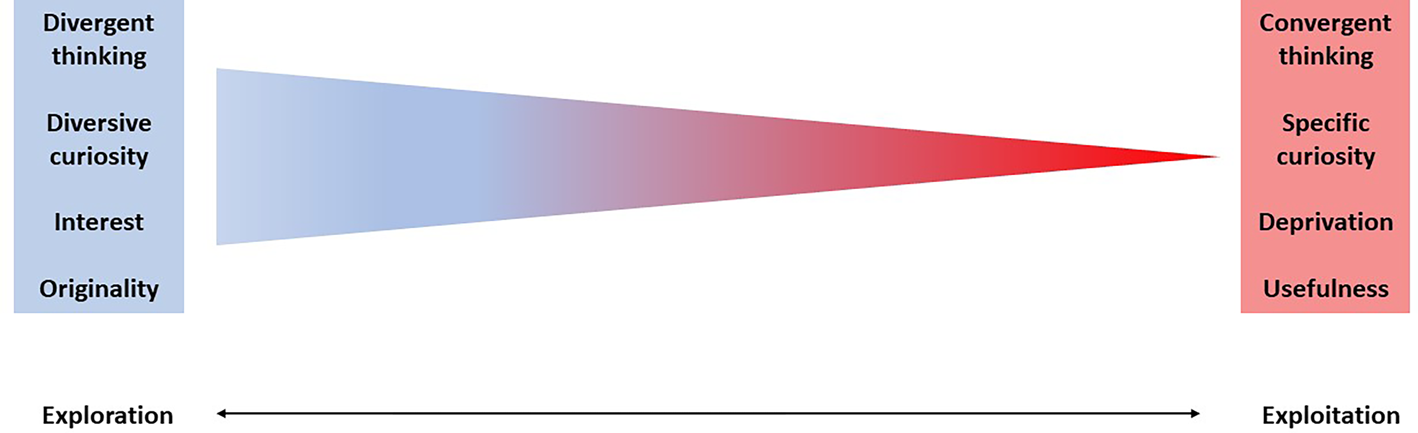
Figure 2. Effects of SoM on the NSM. Illustration of the continuum of SoM ranging from broad to narrow in relation to the NSM. At the left endpoint broad SoM consists of an exploratory disposition that results in diversity; thus, it is associated with divergent thinking, diverse curiosity, and an originality orientation. At the right endpoint, narrow SoM consists of an exploitatory disposition, resulting in specificity. Therefore, convergent thinking, specific curiosity, and a usefulness orientation are associated with exploitation.
5.2 Flexible control dynamics
Much like the trade-off between exploration–exploitation, adaptive levels of control might be indicative of cognitive flexibility and benefit novelty seeking. Therefore, the ability to flexibly switch between inhibition/disinhibition and exploration/exploitation might lead to an optimal novelty-seeking performance, that is, generating/consolidating novel and useful knowledge.
We suggest that cognitive control tunes the flow of stimulation by screening which input will enter, and what ideas will arise. In this framework, disinhibition results in an overflow of stimulation that may not be translated into an output that is a product of creativity or curiosity, whereas hyper-inhibition may lead to a stand-still when we stick to existing knowledge and a premature closure of ideas that could otherwise be further developed (Runco & Basadur, Reference Runco and Basadur1993). The interaction between these two factors results in different types of curiosity and creativity. These manifestations could be explained using the matrix portrayed in Figure 3. The position on this two-axis matrix determines the quality and quantity of the NSM. The cross-points between the axes determine the expression the NSM will take. Once activated, the degree to which each type of curiosity/creativity is experienced and behaviorally expressed varies according to individual differences and task constraints. At the extremes, hyper-inhibition × exploitation would result in a stand-still state when we stick with our previous knowledge, whereas disinhibition × exploration results in an overflow of information and bizarre ideas.
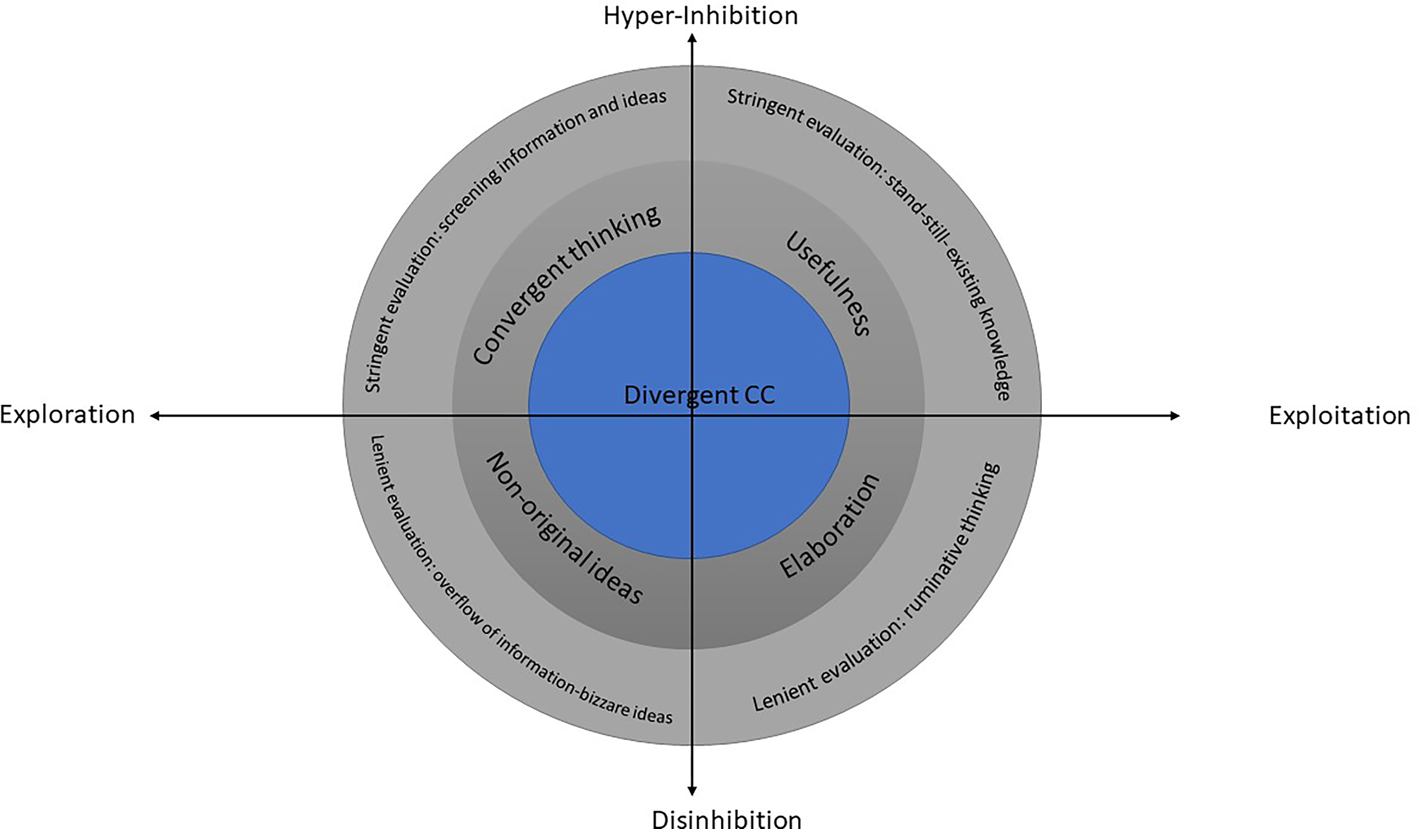
Figure 3. Different types of curiosity and creativity, as manifested by the SoM × inhibitory control matrix. The interaction between SoM and inhibitory control results in different types of curiosity and creativity. The position on this two-axis matrix determines the quality and quantity of the NSM. The central circle represents a balanced interaction between the two factors, and results in an ideal expression of divergence as one moves toward the endpoints of the continuum.
When we are somewhere in the middle along this continuum, the NSM is expressed in various ways. Exploration × inhibitory control will result in a CT mode, which requires the elimination of incorrect solutions, yet it also finds the right creative solution. Exploitation × inhibitory control will result in usefulness: Generating practical solutions and looking for specific information. Exploitation × disinhibition will result in the elaboration of existing ideas, and exploration × disinhibition might result in non-original ideas. The balanced condition will result in DT and diversive curiosity (interest). Using the matrix, one can better understand why it is not a matter of either–or, but rather a dynamic interplay between “opposing” forces that are altered throughout the process and determine the different manifestations of the NSM. This is based on the relative weight and balance between the forces and is influenced by the context demands and available resources. If so, flexibility is key to enhance both curiosity and creativity.
5.3 Clinical perspective
Our framework provides the basis for characterizing various psychopathologies in relation to the NSM. Although much has already been written about the relationship between creativity and psychopathology (e.g., Abraham, Reference Abraham2014; Carson, Reference Carson2011; Herz et al., Reference Herz, Baror and Bar2020; Jung, Grazioplene, Caprihan, Chavez, & Haier, Reference Jung, Grazioplene, Caprihan, Chavez and Haier2010; Post, Reference Post1994; Simonton, Reference Simonton2003), research into curiosity and psychopathology is still lacking. The suggested matrix, with the SoM × inhibition axes, has potential clinical implications. At one extreme of the matrix, schizophrenic individuals are characterized by hyper-exploration × disinhibition, which results in delirium and bizarre ideas, rather than in original ones. Considering that such individuals also suffer from a decreased level of semantic processing (Pomarol-Clotet, Oh, Laws, & McKenna, Reference Pomarol-Clotet, Oh, Laws and McKenna2008) and impaired episodic memory (Goldberg, Keefe, Goldman, Robinson, & Harvey, Reference Goldberg, Keefe, Goldman, Robinson and Harvey2010), both of which are crucial for associative thinking, could further explain why the NSM takes an extreme and pathological manifestation in such cases.
At the other extreme, and far from the myth of the “tormented yet creative artist,” depression can be characterized by exploitation × hyper-inhibition, resulting in a narrowed scope of associative thinking (perseveration of an extremely narrow SoM; Herz et al., Reference Herz, Baror and Bar2020) that might be expressed as pathological ruminations (repetitive thought patters, e.g., Nolen-Hoeksema, Reference Nolen-Hoeksema2000). In this case, increased self-referential processing (i.e., the tendency to experience stimuli as strongly related to one's own person; Northoff et al., Reference Northoff, Heinzel, de Greck, Bermpohl, Dobrowolny and Panksepp2006; Wright & Beck, Reference Wright and Beck1983), together with a high probability of reliance upon top-down processing (Herz et al., Reference Herz, Baror and Bar2020), leaves no resources for exploring the environment, and results in a lower motivation for novelty seeking. In line with this suggested conceptualization, bipolar affective disorder is characterized by alterations between exploration–exploitation and inhibition–disinhibition. Therefore, the critical point for the NSM to occur, in that instance, would be in the transition between the depressive and manic episodes. Indeed, bipolar disorder was suggested to interweave creative accomplishments (see Johnson et al., Reference Johnson, Murray, Fredrickson, Youngstrom, Hinshaw, Bass and Salloum2012, for a review), and higher rates of the disorder have been found among famous artists, musicians, and writers (Goodwin & Jamison, Reference Goodwin and Jamison2007; Ludwig, Reference Ludwig1992; Rothenberg, Reference Rothenberg2001).
Lastly, it is important to note that although curiosity and creativity are part of the same process, the emphasis can be switched from one entity to the other (when you are creative, you are x% curious and vice versa). Therefore, people can be highly curious, but not necessarily creative. An example of such a case is Asperger's syndrome, in which individuals have high-specific curiosity to such an extent that they can memorize a specific field of knowledge by heart, but they lack the flexibility to be creative. Considering that high-functioning autistic individuals show a deficit in tasks that require cognitive switching, whereas their cognitive inhibition is intact (Kleinhans, Akshoomoff, & Delis, Reference Kleinhans, Akshoomoff and Delis2005) supports this notion and, according to our matrix, implies that Asperger's individuals can be characterized by hyper-inhibition × exploitation. Therefore, the NSM depends on the ability to flexibly alternate between different levels of cognitive mechanisms. Although these cases might challenge our framework, they set the stage for future in-depth studies that should test the inter-relations between the dimensions of curiosity and creativity in various conditions and populations. Future studies should design tasks that directly measure the unified process rather than each construct separately.
6. Implications and future directions
In this article, we postulated that a unified process of novelty seeking may be shared in underlying both creativity and curiosity. We provided a modified model (NSM), which expands previous dual-process models, and demonstrated how curiosity and creativity similarly evolve through each of the four stages of the model. We suggest that the NSM begins with an affinity toward the novel that is manifested by a specific pattern of neural firing, initiated by the novelty of response. Using this framework, one can imagine how attentional fluctuations can tilt the system toward a specific stimulus or idea at a random moment in time. In this manner, an attentional spotlight is directed to what we perceive as novel in curiosity, whereas in creativity, similar attentional processes are applied in order to generate novel concepts along the network of stored representations. Diverse thinking enlarges the spotlight, and more information is obtained. By expanding our knowledge, our memory becomes more interconnected, and we are more likely to reach remote associations. It is a loop of feeding and generating: The more combinations cross the novelty threshold, the more interconnected our memory becomes, which will eventually raise the chances that the NSM will be initiated once again.
This is a cyclical process in which one walks through all the stages to be creative or curious. Iterations of the process may result in greater affinity via expansion of memory, and this iterative process potentially explains the reinforcing nature of both curiosity and creativity. That said, it should be noted that although we propose shared mechanisms for curiosity and creativity, and although evidence points to the possibility that curiosity may set the stage for creative thinking, empirical evidence to support a causal relationship between the two is still very scarce (Gross, Zedelius, & Schooler, Reference Gross, Zedelius and Schooler2020). It may be that curiosity is an impetus for problem identification and information gathering, upon which further processes leading to creativity then build. On the other hand, it might be that creative behavior positively engages individuals so that they want to know more, resulting in greater curiosity (Schutte & Malouff, Reference Schutte and Malouff2020). To date, very few studies have experimentally manipulated curiosity to look for causal effects. The aforementioned studies are correlational and rely on self-reported trait measures of curiosity. Indeed, one issue that currently limits curiosity research is the lack of behavioral measures that assess curiosity as a psychological state (Gross et al., Reference Gross, Zedelius and Schooler2020). This is especially prominent in comparison to creativity research, where a vast selection of tasks may result in inconsistent results. Future studies should design such behavioral tasks and induce, for example, a curious state prior to creative thinking tasks, in order to shed light on the causal relationship between the two.
Following the SoM framework (Herz et al., Reference Herz, Baror and Bar2020), we further suggested that although the flexible alterations between exploratory and exploitatory SoMs are essential for both creativity and curiosity, they do not have equal utility. Both behaviors stem mainly from exploratory orientation and its interlinked dimensions, such as defocused attention and bottom-up processing. Exploitatory SoM on the other hand, associated with top-down control, balances and directs the process, but novelty seeking that drives curiosity and creativity is primarily encapsulated in an exploratory SoM.
The theoretical framework suggested here, according to which curiosity and creativity are two manifestations of a unified process, has several implications. Acknowledging the underlying mechanisms of the NSM, and experimentally manipulating them, may help to enhance curiosity and creativity by need. This can be tested experimentally in behavioral methods, such as cognitive stimulation, that would influence the processing resources needed to unleash novelty-seeking behavior. Furthermore, targeting the neural mechanisms underlying the NSM by means of direct neural stimulation could potentially enhance novelty-seeking behavior as well. This notion was already tested in creativity using transcranial direct current stimulation (tDCS; see Lucchiari, Sala, & Vanutelli, Reference Lucchiari, Sala and Vanutelli2018, for a review), transcranial magnetic stimulation (TMS; e.g., Flaherty, Reference Flaherty2005), and neurofeedback (Gruzelier, Reference Gruzelier2014), but to date, such efforts in the field of curiosity are still to come.
Our efforts to link curiosity and creativity are wrought of some limitations. First and foremost, to delineate our discussion within existing research, here we referred to both curiosity and creativity in a narrow context of which they are controllably operationalized and measured in the lab. From the same reasons, here we mainly embraced a neurocognitive perspective. Although we are aware that both are multifaceted and entail much more than the context described in the current article (educational and developmental perspectives, artistic or Big C creativity, personality traits, to mention a few) we hope that the theoretical framework proposed here may apply to curiosity and creativity “in the wild.” As more detailed discussion regarding other facets of curiosity and creativity are beyond the scope of this article, we encourage future research to investigate their relationship in different contexts and perspectives.
Second, the shared cognitive faculties and brain networks involved in creativity and curiosity included in the NSM are involved in many aspects of cognition. Similar to other complex constructs, although these mechanisms are not solely underlying curiosity and creativity, we aimed to conceptually establish a link between the two by showing how those faculties change in tandem. Once the link between curiosity and creativity has been created, we hope that it will serve as a foundation for further theorizing about the nature of their unified process.
7. Concluding remarks
It may be common knowledge that curious people are more creative, but surprisingly, no systematic investigation has been conducted so far to explain why and how this may be true. Here, we reviewed cognitive and neural mechanisms that might explain the intuitive link between curiosity and creativity and, for the first time, provide a systematic explanation for their relationship. We propose that both are manifestations of a unified process that underlies novelty seeking, as illustrated by a novelty-seeking model. We claim that both have a reinforcing nature, the more curious and creative you are, the higher your sensitivity to novel stimuli would be. Lastly, we demonstrated how our state of mind determines which form curiosity and creativity will take. We believe that integrating curiosity and creativity under one perspective can, in turn, facilitate greater intrafield understanding of both processes.
Financial support
T. I.'s work in this article was supported by a Joy Ventures postdoctoral fellowship grant. M. B.'s work was supported by Israel Science Foundation Grant 673/17 and by the Sagol Family.
Competing interest
None.




Target article
A shared novelty-seeking basis for creativity and curiosity
Related commentaries (29)
A developmental account of curiosity and creativity
A shared “optimal-level of arousal”: Seeking basis for creativity and curiosity
An extension of the novelty-seeking model: Considering the plurality of novelty types and their differential interactions with memory
Be curious: Strategic curiosity drives creativity
Beyond novelty: Learnability in the interplay between creativity, curiosity and artistic endeavours
Breaking down (and moving beyond) novelty as a trigger of curiosity
Commentary on creativity and curiosity
Computational models of intrinsic motivation for curiosity and creativity
Creativity is motivated by novelty. Curiosity is triggered by uncertainty
Curiosity is more than novelty seeking
Curious? The relationship between curiosity and creativity is likely NOT novelty
Dissecting the neuroanatomy of creativity and curiosity: The subdivisions within networks matter
Distinct neurocognitive pathways underlying creativity: An integrative approach
Expanding horizons in reinforcement learning for curious exploration and creative planning
Exploratory exploitation and exploitative exploration: The phenomenology of play and the computational dynamics of search
Getting curiouser and curiouser about creativity: The search for a nuanced model
Ignoring the role of reiterative processing and worldview transformation leads to exaggeration of the role of curiosity in creativity
Is a wandering mind a novelty-seeking mind? The curious case of incubation
Mindfulness, curiosity, and creativity
Models need mechanisms, but not labels
Mood regulation as a shared basis for creativity and curiosity
Novelty seeking is neither necessary nor sufficient for curiosity or creativity, instead both curiosity and creativity may reflect an epistemic drive
Novelty seeking might underlie curiosity and the novelty dimension of creativity, but not the usefulness dimension
On the dual nature of creativity: Same same but different?
Prediction error minimization as a common computational principle for curiosity and creativity
Question-asking as a mechanism of information seeking
The costs of curiosity and creativity: Minimizing the downsides while maximizing the upsides
The creativity of architects
Toward a causal model of curiosity and creativity
Author response
A shared novelty-seeking basis for creativity and curiosity: Response to the commentators