


Introduction
Extensive systems to produce heavy pigs (slaughtered at 140 to 180 kg body weight (BW) and 18 to 25 months of age) are typically located in some regions of the Mediterranean area. Their low productivity encourages current tendencies to shorten their productive cycle by intensification of management during the growing and early stages of the finishing period. The nutritional requirements of the Iberian pig (Sus mediterraneus), a slow-growing, obese porcine breed, are not well defined and seem to differ from those of conventional (standard performance) or high-performing pigs. Nieto et al. (Reference Nieto, Miranda, García and Aguilera2002) demonstrated that the optimal dietary protein to energy ratio was considerably lower in Iberian pigs growing from 15 to 50 kg BW than in pigs with a greater genetic potential for protein deposition (PD) (Campbell et al., Reference Campbell, Taverner and Curic1985; Kyriazakis and Emmans, Reference Kyriazakis and Emmans1992). Maximal PD (74 g/day) was achieved with a diet supplying 6.86 g digestible ideal crude protein (CP) per MJ metabolisable energy (ME) fed close to ad libitum intake. Although conflicting results have been reported (Siebrits et al., Reference Siebrits, Kemm, Ras and Barnes1986; Whittemore et al., Reference Whittemore, Tullis and Emmans1988; Möhn and De Lange, Reference Möhn and De Lange1998), the relationship between PD and energy intake may change as the animal increases BW. To test this hypothesis in the growing-fattening Iberian pig (50 to 100 kg BW) a wide range of dietary protein to energy ratios were assayed. In this way an appropriate balance between ideal CP and energy concentrations can be established and the response of the animals to changes in energy or amino acid (AA) supply can be predicted. To achieve our objective, the optimal supply of dietary ideal CP leading to maximum daily rates of PD (PDmax) and the marginal efficiency of body PD (Fuller et al., Reference Fuller, Franklin, McWilliam and Pennie1995), i.e., the change in daily PD per unit change in daily ME at restricted intakes (ΔPD:ΔME; g/MJ), were considered. A second objective was to establish reliable estimates of energy utilisation in the growing-fattening pig. This information, together with corresponding data on carcass quality as the main source of edible products, is relevant for application in the practical feeding of heavier animals prior to their extensive management. At this respect, in a previous paper we have reported that both the protein to energy ratio of the diet and the level of feeding affected carcass quality (Barea et al., Reference Barea, Nieto, Lara, García, Vilchez and Aguilera2006). Also, altering the feed intake resulted in significant changes in carcass and viscera as proportions of empty BW.
Material and methods
Animals and experimental design
As described in the previous paper (Barea et al., Reference Barea, Nieto, Lara, García, Vilchez and Aguilera2006), three groups of purebred castrated male Iberian pigs of Silvela strain were used. They were from the same herd of pigs used in our previous experiment (Nieto et al., Reference Nieto, Miranda, García and Aguilera2002). The protein and energy content of the standard diet given to the experimental pigs before the beginning of the experiments were similar to our previous study (Nieto et al., Reference Nieto, Miranda, García and Aguilera2002). In total, 81 Iberian castrates, 49.9 ± 0.0 kg initial BW, were randomly assigned to 4 diets containing 145 to 70 g ideal CP per kg dry matter (DM) and three feeding levels (FL) in a factorial arrangement of treatments with six to seven individually housed pigs per treatment. Due to the availability of only 28 individual pens, the experiment was conducted in three replicates, each with two or three pigs per treatment. When establishing the ideal CP for dietary treatments, it was assumed that the PDmax value for the growing-fattening pig would be somewhat lower than the rate previously observed in the younger pig (Nieto et al., Reference Nieto, Miranda, García and Aguilera2002). In that study, the PDmax was obtained with a diet providing 6.86 g digestible ideal CP per MJ ME, a value which is within the range of the values used in the present experiment. The range of FL was chosen to allow an accurate estimation of the efficiency of dietary energy utilisation, while the higher allowances are usual in a practical feeding.
Diets and feeding
Four diets of similar ME concentration were formulated to contain 145, 120, 95, and 70 g ideal CP per kg DM. These diets were prepared by mixing a protein-free diluting mixture and a high-protein (HP) diet (Table 1). The HP diet was formulated to provide an optimal pattern of amino acids (Table 2), according to the ideal protein concept (Agricultural Research Council (ARC), 1981). The diluting mixture was made to match the macronutrient content of the HP diet. In this way, the relative proportions of nutrients in the experimental diets would remain constant, except for the replacement of CP by starch, which causes a slight increase in ME concentration. However, this increase in the energy value was considered irrelevant because the key factor affecting PD in the pig is the amount of digestible ideal CP per unit of ME, which resulted to be 8.05, 6.50, 5.20 and 3.65 g/MJ. Each of the four diets was offered at three FL (0.95, 0.80 and 0.60 × ad libitum intake) daily in two equal meals. Water was available ad libitum.
Table 1 Composition and chemical analysis of the high-protein (HP) diet

† Provided the following per kg diet: retinol, 3.38 mg as retinyl acetate; cholecalciferol, 56.3 μg; dl-α-tocopherol, 2.52 mg as dl-α-tocopheryl acetate; menadione, 1.5 mg as menadione sodium bisulfite; thiamine, 0.15 mg; riboflavin, 3 mg; pyridoxine, 0.15 mg; cyanocobalamin, 15 μg; folic acid, 15 μg; nicotinic acid, 22.5 mg; d-pantothenic acid, 15 mg as calcium pantothenate; Mn, 15 mg as MnSO4.4H2O; Fe, 75 mg as FeSO4.7H2O; Zn, 120 mg as ZnO; I, 450 μg as KI; Cu, 60 mg as CuSO4.5H2O; Co, 300 μg as CoSO4.7H2O.
‡ l-Lys HCl containing 78.8% l-Lys; l-Thr and l-His, added as free base, 98.5 and 100% l-Thr and L-His, respectively.
§ Mixed with HP diet to prepare experimental diets. Containing (g/kg, as fed): maize starch, 883.1; cellulose, 64.2; dibasic calcium phosphate, 29; corn oil (containing butylated hidroxytoluene to provide 0.125 g per kg diluting mixture), 15.7; common salt, 5; vitamin/mineral pre-mix, 3. Analysed chemical composition (g/kg): total N, < 0.5; dry matter, 878.
Table 2 Nutrient composition of the experimental diets obtained by dilution of a high-protein (HP) diet with a diluting mixture based on maize starch

† TDP: true digestible protein; TDLys: true digestible lysine; TDAA/TDLys: true digestible amino acid relative to TDLys; DP: apparent digestible ideal CP; DM: dry matter.
The ad libitum intake was defined as what the pig would consume when given sole access to its respective diet for two periods of 60 min/day, and it was estimated previously by offering similar diets to those used in the present experiment to individually housed castrated male Iberian pigs (50 to 85 kg BW) in a room maintained at 19.0 ± 1.5°C (Nieto et al., Reference Nieto, Lara, García, Gómez, Zaldive, Cruz, Pariente, Moreno and Aguilera2001). Based on the BW of the pig, the daily allowance was determined weekly prior to feeding using the formula:
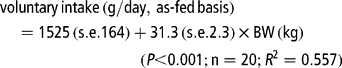
Pig performance and digestibility measurements
On arrival, pigs were group-housed and given ad libitum access to a barley-soybean meal-based diet containing 165 g ideal CP per kg DM and approximately 14.5 MJ ME per kg DM until they reached 50 kg. Pigs were then housed in individual pens (2 m2) located in a environmentally controlled room (19.0 ± 1.5°C) until they reached 100 kg when they were slaughtered. During the experiment, the pigs were weighed once per week before feeding and for each animal the daily feed allowance for the following week was adjusted accordingly. Digestibility trials were conducted at approximately 75 kg BW, as described in Barea et al. (Reference Barea, Nieto, Lara, García, Vilchez and Aguilera2006). Apparent energy digestibility [d(E)] and metabolisability [m(E)] were calculated as digestible energy (DE): gross energy (GE) and ME:GE ratios.
Comparative slaughter measurements
At 100 kg BW the pigs were electrically stunned and bled, and blood was collected. After emptying the gut, carcass and non-carcass parts were weighed separately. Viscera and blood were kept at − 20°C until analysis. The eviscerated carcass with head, feet and tail were chilled overnight and weighed. Four components were obtained for each pig: carcass, head (together with feet and tail), viscera and blood. The carcass without head was split longitudinally. After weighing both halves, they were sealed in plastic bags and kept at − 20°C until analysis. The right half of the carcass and the rest of components were cut into small pieces and ground in a mincer (Talleres Cato, Sabadell, Spain) separately, homogenised in a cutter (Talleres Cato, Sabadell, Spain), and sub-samples were taken for freeze-drying and subsequent analysis. Aliquot samples were analysed for DM content, CP (total N × 6.25), GE and ash. Body fat was calculated from total body GE assuming an energy content of 23.85 and 39.75 kJ/g for protein and fat, respectively.
To predict body composition of the experimental pigs at the start of the trial, the data from 18 additional pigs were obtained from the previous experiment carried out in Iberian pigs growing from 15 to 50 kg BW (Nieto et al., Reference Nieto, Miranda, García and Aguilera2002). These pigs were offered diets containing 175 and 156 g ideal CP per kg DM with 14.4 and 14.8 MJ ME per kg DM, respectively, at 0.80 and 0.95 × ad libitum. Small and not significant differences in whole-body composition due to the treatments imposed were observed. Pigs were slaughtered before initiating the present experiment. The relationships between live and empty BW at slaughter and the analysed body composition obtained both in the previous experiment were used to estimate the initial body composition of pigs used in the present experiment. Total body composition was calculated from the chemical composition of the four body components and their respective weights. Increases in protein (determined as total nitrogen (N) retention), energy, fat and minerals were then calculated as the difference between final measured composition of experimental pigs and estimated initial composition assessed from the initial group. The experimental protocol was approved by the Bioethical Committee of the Spanish Council for Scientific Research (CSIC), Spain.
Chemical analysis
All analyses were performed in duplicate. The DM content of feeds and faeces was determined (Association of Official Analytical Chemists (AOAC), 1990). Whenever an analysis was made on a freeze-dried material, the DM content of an aliquot sample was determined in a ventilated oven to estimate the residual moisture content after freeze-drying, and the corresponding result was expressed on a DM basis. Total lipids in feeds were determined by ether extraction (AOAC, 1990). Acid-detergent fibre content in feed samples was analysed by the method of Goering and Van Soest (Reference Goering and Van Soest1970). Total N in feed, urine, freeze-dried samples of body components (except blood), and faeces was determined by the Kjeldahl procedure. After hydrolysis with 6 mol/l HCl plus 1% phenol in sealed, evacuated tubes at 110°C for 24 h, amino acids in feed samples was determined by HPLC according to the Waters Pico Tag method (Cohen et al., Reference Cohen, Meys and Tarvin1989). Cysteine and methionine were determined as cysteic acid and methionine sulphone, respectively, after oxidation with performic acid before hydrolysis with 6 mol/l HCl (Moore, Reference Moore1963). The total ash content of feed and freeze-dried body components samples was determined (AOAC, 1990). The GE content of feed, freeze-dried body components, and faecal samples was measured in an adiabatic bomb calorimeter (Gallenkamp Autobomb CBA 305, Loughborough, UK). Urine was freeze-dried in a polyethylene sheet of known energy value and their GE values were obtained by difference.
Statistical analysis
The data were subjected to an analysis of variance using the GLM procedure of Statistical Analysis Systems Institute (1990). The individual pig was used as the experimental unit. The effects of dietary ideal CP, FL, block (or replication) and all possible interactions were included in the initial statistical model. With no significant block effect or no treatment by block interactions, these terms were deleted from the statistical model for subsequent analyses. Orthogonal polynomials were used to determine linear and quadratic effects of the dietary ideal CP and the FL on response criteria.
Regression analyses
Linear regressions were calculated for each experimental diet to relate rate of gain (g/day) and ME intake. For this purpose the average value of the whole feeding period for each pig was used. All experimental diets resulted in homogeneous regression coefficients and a composite regression was developed for the pooled data. The marginal efficiency of body PD (ΔPD:ΔME, g/MJ) was estimated by linear regressions with individual data from the different dietary treatments. All diets resulted in homogeneous regressions. The intercept of the composite regression calculated on pooled data did not differ significantly from zero, and a new regression forced to the origin was constructed.
To predict energy maintenance requirements (MEm) and calculate net efficiencies of energy utilisation in the growing animal (kg), ME intake (kJ/kg BW0.75 per day) was related to the total retained energy (RE, kJ/kg BW0.75 per day) by linear regression equations using ME intake as either an independent or dependent variable. Separate regressions were calculated within the dietary ideal CP content. All diets showed homogeneous regression coefficients, and pooled equations were established accordingly. Estimates of the partial efficiency of the ME utilisation for PD (kp) and fat deposition (kf) were calculated by a multiple regression equation (Kielanowski, Reference Kielanowski1965), using ME intake (kJ/kg BW0.75 per day) as a dependent variable and energy retained as protein and fat (REp and REf, respectively, kJ/kg BW0.75 per day) as independent variables. An equation was derived with the pooled data for the four dietary treatments.
Results
All pigs were in good health during the whole trial. No feed refusals were observed except for some pigs on the feeding level closer to ad libitum. Any refusals were collected each morning, dried to a constant weight and weighed to calculate the actual consumption.
Feed intake and digestibility measurements
Feed intake was not affected by the dietary ideal CP content and averaged 2.58 kg DM per day (Table 3). There were linear (P < 0.01) and quadratic (P < 0.01) effects of the FL on feed intake. Average daily DE and ME intakes were 29.2 and 28.7, 38.6 and 37.9, and 45.4 and 44.6 MJ for the three FL, respectively. The digestibility and metabolisability of energy increased linearly (P < 0.001) by decreasing the ideal CP content of the diet (Table 3). The decreases in d(E) and m(E) were partially due to the dilution of the HP diet with a slightly higher digestible protein-free mixture. The change in the value was slightly higher for m(E), which increased from 0.772 to 0.852 by decreasing the dietary ideal CP content from 145 to 70 g/kg DM. On average, the energy density of the diets increased from 13.94, 14.29, 14.56 and 14.83 MJ ME per kg DM by the reduction in the ideal CP content. Linear (P = 0.01 and P < 0.001) and quadratic (P < 0.05 and P < 0.01 respectively) effects of the FL on d(E) and m(E) were observed, with the 0.60 × ad libitum intake resulting in the highest d(E) and m(E) ratios. The ME:DE ratio increased linearly with decreasing the ideal CP content of the diet (P < 0.001), but it remained close to 0.98 regardless of the feeding allowance.
Table 3 Effects of dietary protein content (DPC) and level of feeding (FL) on the digestibility of energy (d(E)) and nitrogen (d(N)) and metabolisability of energy (m(E)) in Iberian pigs growing from 50 to 100 kg body weight (mean values for 20 or 21 pigs per DPC group)

† L = linear contrast; Q = quadratic contrast. DPC × FL interactions were not significant (P>0.05).
Apparent digestible N:N intake (or apparent digestibility of CP) was unaffected by the dietary ideal CP. But, it increased linearly from 0.756 to 0.791 by decreasing the FL (P < 0.001).
Growth performance
Average daily gain (ADG), gain:feed and gain:ME intake are shown in Table 4. ADG increased (linear and quadratic, P < 0.001) by decreasing the dietary CP content. As expected, ADG increased with the FL (linear, P < 0.001; quadratic, P < 0.05). Gain:feed and gain:ME intake improved by decreasing the ideal CP content in the diet (linear, P < 0.001 and P < 0.05, respectively; quadratic P < 0.001 for both variables). The diet supplying 95 g ideal CP per kg DM resulted in a relative greater gain:ME intake than those containing 145 or 70 g ideal CP per kg DM. Increasing the FL improved linearly gain:feed intake and gain:ME intake (P < 0.001). No interactions were observed.
Table 4 Effects of dietary protein content (DPC) and level of feeding (FL) on the performance and protein deposition of Iberian pigs growing from 50 to 100 kg body weight (mean values for 20 or 21 pigs per DPC group)

† L = linear contrast; Q = quadratic contrast. DPC × FL interactions were not significant (P>0.05).
‡ Approaching significance (P < 0.1).
§ PD, protein deposition.
‖ ApDN, apparent digestible nitrogen.
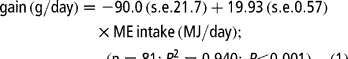
Equation [1] is a composite regression developed for the rate of gain and ME intake based on the pooled data irrespective of the dietary ideal CP because all diets resulted in homogeneous slopes. The regression coefficient shows an average increase of 19.93 g gain per MJ increase in ME intake, equivalent to an energy cost of 50.2 kJ ME/g gain (1/19.93):
Protein deposition
Mean estimated initial body composition at 50 kg BW was 137.1 ± 1.3, 324.9 ± 6.3, 32.9 ± 0.8 and 502.2 ± 5.4 g/kg empty BW for protein, fat, ash and water, respectively. There were no interactions between the dietary ideal CP content and the FL for PD variables (Table 4). There was a quadratic response (P < 0.05) in the rate of PD as dietary ideal CP content decreased. However, the decrease in the FL reduced linearly the rate of PD (P < 0.001). Nitrogen retained (obtained by comparative slaughter measurements):N intake and N retained:apparent digestible N improved linearly with the decrease in the dietary ideal CP content (P < 0.001). Increasing the FL from 0.60 to 0.80 or 0.95 × ad libitum reduced N retained:N intake and N retained:apparent digestible N in a quadratic pattern (P < 0.05 for both variables). Protein deposition:ME intake ranged between 1.23 and 1.44. There were tendencies for quadratic responses in PD:ME intake of dietary ideal CP content and FL (P = 0.078 and 0.058, respectively).
The pooled equation [2], obtained with data from all diets, shows a marginal efficiency of body PD (ΔPD:ΔME; g/MJ) of 1.34 g. This indicates that within the range of the dietary ideal CP content investigated, each additional MJ in ME at restricted intakes was associated with 1.34 g of PD in the Iberian pig growing from 50 to 100 kg BW:
Energy utilisation
Data from energy retention and their partition into protein and fat deposition expressed in kJ/kg BW0.75 per day are presented in Table 5. No interactions between the dietary ideal CP content or the FL were noted. Retained energy increased linearly (P < 0.001) by decreasing the ideal CP content in the diet and showed linear (P < 0.001) and quadratic responses on increasing the FL (P < 0.01). Retained energy as protein increased in a quadratic pattern by decreasing the dietary ideal CP content (P < 0.05), and linearly (P < 0.001) with increasing the FL. Energy deposition as fat increased linearly (P < 0.001) by decreasing the dietary ideal CP content and increased linearly (P < 0.001) and quadratically (P < 0.01) as the FL increased. The overall efficiency of utilisation of the ME for energy gain (RE:ME) improved linearly (P < 0.001) as the dietary ideal CP content decreased, whereas it increased linearly as the FL increased (P < 0.001). The proportion of the energy retained as protein or fat to the total energy retained (REp:RE or REf:RE, respectively) was linearly influenced by both factors (P < 0.01 for dietary protein content and FL). The effect was to increase the proportion of fat deposition in replacement of PD with decreasing the supply of dietary ideal CP and with increasing the FL. Mean values for REp:RE and REf:RE were 7.4% (s.e. 0.61) and 92.6% (s.e. 0.61) respectively.
Table 5 Effects of dietary protein content (DPC) and level of feeding (FL) on the energy balance of Iberian pigs growing from 50 to 100 kg body weight (BW) (mean values for 20 or 21 pigs per DPC group)

† L = linear contrast; Q = quadratic contrast. DPC × FL interactions were not significant.
‡ RE, total retained energy.
§ REp, energy retained as protein.
‖ REf, energy retained as fat.
Equations [3] and [4] were constructed with data from all diets to predict the energy requirement for maintenance and the net efficiency of energy utilisation in the finishing animal:
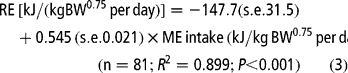
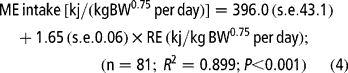
Equation [3] indicates a value of 0.545 for the net efficiency of utilisation of dietary energy for growth (kg), and estimates the ME requirement for maintenance (MEm) as 272 kJ/kg BW0.75 per day. Equation [4] estimates MEm as 396 kJ/kg BW0.75 per day. The regression coefficient of this equation shows an energetic cost for growth of 1.65 kJ ME per kJ RE and a kg value of 0.606 (1/1.65). Estimates of the partial efficiency of the ME utilisation for PD and fat deposition were calculated by the following multiple regression equation:
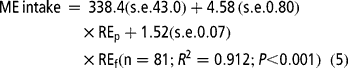
Equation [5] estimates MEm as 338 kJ/(kg BW0.75 per day) and k p and k f as 0.218 (1/4.58) and 0.658 (1/1.52), respectively. From the coefficients of the energy retained as protein and fat, energy costs were estimated to be 109 (4.58 × 23.85) and 60 (1.52 × 39.75) kJ ME per g for protein and fat, respectively.
Composition of gain
The empty BW to BW ratio was not affected by experimental treatments and the mean was 0.973 (Table 6). The average rates of deposition of fat, ash, water and energy in the empty BW of Iberian pigs growing from 50 to 100 kg BW are also presented in Table 6. Body fat deposition and energy retention increased linearly (P < 0.001) with each decrease in the dietary ideal CP content. Similarly to PD, water retention showed a quadratic increase (P < 0.01) by decreasing the dietary ideal CP. The rates of deposition of fat, ash, water and energy were linearly increased (P < 0.001) as the FL increased. The mean whole-body chemical composition (g/kg) at approximately 100 kg empty BW was 108.8 ± 2.7, 465.6 ± 9.9, 31.2 ± 2.1 and 392.0 ± 8.1 for protein, fat, ash and water, respectively. Between 50 and 100 kg, the chemical composition of 1 kg gain averaged 77.6 ± 4.9, 592.2 ± 19.0, 28.7 ± 2.9 and 284.0 ± 15.0 g for protein, fat, ash and water, respectively.
Table 6 Effects of dietary protein content (DPC) and level of feeding (FL) on the rates of deposition of fat, ash, water and energy in the empty BW of Iberian pigs growing from 50 to 100 kg body weight (BW) (mean values for 20 or 21 pigs per DPC group)

† L = linear contrast; Q = quadratic contrast. DPC × FL interactions were not significant.
‡ Approaching significance (P < 0.1).
Discussion
In the current experiment, the response of Iberian pigs (50 to 100 kg BW) to changes in energy intake at different ideal CP concentrations was studied. Expressed on a metabolic BW basis, average ME intakes were 1.15, 1.52 and 1.80 MJ/(kg BW0.75 per day), respectively, for 0.60, 0.80 and 0.95 × ad libitum FL. Taking into account our estimate for MEm (396 kJ/kg BW0.75 per day) calculated from equation 4, these energy intakes provided 2.9, 3.8 and 4.5 times the maintenance energy requirements for this period of growth.
The rate of growth and PD depended on the FL, and also on the ideal CP content (Table 4). As a result of the low potential for lean tissue deposition in the Iberian pig, the efficiency of utilisation of the dietary ideal CP protein was rather poor, indicating that substantial amounts of the ideal CP provided were used for lipid gain. At fixed energy intakes, the efficiency with which dietary protein was deposited in tissues, expressed either as NR:NI or NR:ApDN, varied inversely with the supply of dietary ideal CP. The result is in agreement with Nieto et al. (Reference Nieto, Miranda, García and Aguilera2002). The maximum PD in Iberian pigs growing from 50 to 100 kg BW was obtained with the diet supplying 95 g ideal CP per kg DM. Taking into account our average apparent digestibility for CP, this diet provided 5.17 g digestible protein per MJ ME. This CP level also showed the highest PD (71 g/day) when offered at 0.95 × ad libitum. This feeding regimen resulted in ADG of 854 g/day and RE of 21.4 MJ/day. It should be pointed out that the PDmax was obtained with 95% ad libitum, and therefore, might slightly underestimate the PDmax for these pigs. Nieto et al. (Reference Nieto, Miranda, García and Aguilera2002) used Iberian pigs growing from 15 and 50 kg BW and obtained PDmax of 74 g/day when the diet containing 129 g ideal CP per kg DM (providing 6.86 g digestible ideal CP per MJ ME) was offered close to ad libitum intake. Both values of PDmax are basically the same for the two periods of growth. This observation is in agreement with results of, among others, Whittemore et al. (Reference Whittemore, Tullis and Emmans1988), Quiniou et al. (Reference Quiniou, Dourmad and Noblet1996), and Möhn and De Lange (Reference Möhn and De Lange1998) who found no significant effect of the stage of growth on PDmax, but in contrast with the increasing pattern of the response of PD to live weight up to a peak rate found by others (Thorbek et al., Reference Thorbek, Chwalibog and Henckel1984; Tauson et al., Reference Tauson, Chwalibog, Jakobsen and Thorbek1998; Siebrits et al., Reference Siebrits, Kemm, Ras and Barnes1986). The maximum capacity for PD obtained in the present work is far lower than values found in lean and conventional genotypes at a similar BW range (>150 g/day; Quiniou et al., Reference Quiniou, Dourmad and Noblet1996) and similar to the average value of 75.6 g/day in Meishan castrated fed close to ad libitum and growing from 20 to 107 kg BW (Noblet et al., Reference Noblet, Karege, Dubois and Van Milgen1999).
Our results show clearly that the rate of PD was increased as the FL increased even when most of the diets were not limiting in ideal CP supply. The relationship between PD and ME intake was linear and similar for all the ideal CP levels tested (equation [2]). The low marginal efficiency of PD at restricted intakes (1.34 g PD per MJ increase in ME intake) is associated with the low PDmax found in the Iberian pig at this stage of growth. This observation confirms the suggestion that the decrease in PD due to restriction in energy allowance would be greater in pigs of higher genetic potential for lean tissue deposition (Campbell and Taverner, Reference Campbell and Taverner1988; Rao and McCracken, Reference Rao and McCracken1991; Quiniou et al., Reference Quiniou, Dourmad and Noblet1996). In Iberian pigs growing from 15 to 50 kg BW, Nieto et al. (Reference Nieto, Miranda, García and Aguilera2002) reported a marginal efficiency for PD of 2.81 g PD per MJ ME with the diet that provided the optimal protein concentration for growth. These results are in the line with previous observations by Black et al. (Reference Black, Campbell, Williams, James and Davies1986) and Bikker (Reference Bikker1994) who reported a decrease in marginal efficiency of PD with BW.
In the present experiment, the increase in the rate of lipid deposition with energy intake averaged 13.05 g/MJ ME (12.79 g/MJ DE). This estimate is somewhat below the range of values reported in the literature for pigs of different genotypes growing from 45 to 100 kg BW (13.3 to 15.0 g/MJ DE; Campbell and Taverner, Reference Campbell and Taverner1988; Bikker, Reference Bikker1994; Quiniou et al., Reference Quiniou, Dourmad and Noblet1996), indicating a comparatively higher energetic cost of lipid deposition in the body of the Iberian pig. On average, 0.926 of the energy retained was deposited as fat, of which a considerable portion may have been synthesised from dietary amino acids, and, therefore, with a low energetic efficiency. This may explain the low estimates for the net efficiency of energy gain (k g = 0.545 (equation [3]) and 0.606 (equation [4])) and the high energetic cost of fat deposition (k f = 0.658 (equation [5])). Nieto et al. (Reference Nieto, Miranda, García and Aguilera2002) obtained an estimate of 0.582 for k g in Iberian pigs growing from 15 to 50 kg BW.
We failed to obtain a reliable estimate of the maintenance energy requirements. It is well known that MEm estimates are highly dependent on the statistical model used and show a strong correlation with the variables included in the model (Noblet et al., Reference Noblet, Karege, Dubois and Van Milgen1999). From equation [3], a very low and unacceptable value is derived. Our estimate of 396 kJ/kg BW0.75 per day for MEm calculated by equation 4 is slightly lower than the value obtained in our previous study conducted with Iberian pigs growing from 15 to 50 kg BW (422 kJ/kg BW0.75 per day; Nieto et al., Reference Nieto, Miranda, García and Aguilera2002) and far lower than the preferred estimate of 458 by the ARC (1981) and the mean estimate of 443 kJ/kg BW0.75 per day by National Research Council (NRC, 1998). A decrease in the energy requirement for maintenance is expected because a considerable increase of lipid deposition takes places in pigs growing between 50 and 100 kg (327 g/kg at 50 kg empty BW (Nieto et al., Reference Nieto, Miranda, García and Aguilera2002) and 466 g/kg at 100 kg empty BW). Meanwhile, the contribution of metabolically active body compartments such as gastro-intestinal tract and liver to whole body size may become less (total viscera (g/kg empty BW), 118 g at 50 kg BW (Nieto et al., Reference Nieto, Lara, García, Vílchez and Aguilera2003) v. 98 g at 100 kg BW (Barea et al., Reference Barea, Nieto, Lara, García, Vilchez and Aguilera2006)). Noblet et al. (Reference Noblet, Karege, Dubois and Van Milgen1999) and Van Milgen and Noblet (Reference Van Milgen and Noblet1999) concluded that body muscle and, even more importantly, visceral mass explain a large part in the variation of MEm. Higher maintenance energy requirements in lean genotypes are supported by the findings of Thorbek et al. (Reference Thorbek, Chwalibog and Henckel1984), Campbell and Taverner (Reference Campbell and Taverner1988) and Rao and McCracken (Reference Rao and McCracken1991).
Our estimates of the partial efficiency of use of ME for protein and fat deposition (0.218 and 0.658, respectively) were associated with a low MEm. In the experiment conducted with growing Iberian pigs at an earlier stage of growth, Nieto et al. (Reference Nieto, Miranda, García and Aguilera2002) obtained values of 0.303 and 0.812 for k p and k f, respectively, along with a comparatively higher estimate of MEm. Our present estimations indicate higher energy costs of protein and fat deposition in the Iberian pig in comparison with literature values (ARC, 1981; NRC, 1998). The increased energy cost of synthesising urea to metabolise dietary amino acids in excess of the requirements, and the high protein turn-over rate observed in muscle tissue of the Iberian pig (Rivera-Ferre et al., Reference Rivera-Ferre, Aguilera and Nieto2005) may explain the low energy efficiency of PD.
In the present experiment, at 100 kg BW the empty body of the Iberian pig contained (g/kg) 109 and 466 g protein and fat, respectively. Gains of 78 and 592 g protein and fat per kg, respectively, were deposited on average in the body of the animal between 50 and 100 kg BW, underlining important compositional differences compared with conventional or modern genotypes of pigs.
In conclusion, in the present experiment the protein and energy requirements of Iberian pigs growing from 50 to 100 kg BW are defined. The results support earlier findings that the genotype has marked effects on protein and energy metabolism of growing pigs. The results of the present and previous studies indicate that Iberian pigs growing from 15 to 50 kg and 50 to 100 kg BW have a similar rate of PD. The information is extremely valuable to define feeding strategies, which are economically favourable and environmentally acceptable to achieve optimal growth and maximum efficiency of nutrient utilisation for this Mediterranean pig breed, compatible with obtaining optimal quality products.
Acknowledgements
This study was supported by Spanish MCYT grant no. AGF99-0903. R. Barea was recipient of a FPI grant from the Spanish Ministry of Education and Science. We thank Mrs E. Colmenero and Mr F. Funes for skillful technical assistance and Sánchez Romero Carvajal Jabugo S.A. (Seville) and Sucesores de Miguel Vílchez Riquelme S.A. (Granada) for their helpful collaboration.
Part of present results were presented in Barea R, Nieto R and Aguilera JF 2003. The effect of protein:energy ratio of the diet and feed intake on protein and energy metabolism of Iberian pigs growing from 50 to 100 kg. Progress in research on energy and protein metabolism. EAAP publ. no. 109, pp. 121–124. Rostock-Warnemünde, Germany.






