While it is clear that environmental factors play a significant role in the development of obesity, research work over the last decades has clearly documented a genetic contribution to obesity-related phenotypes as well. Obesity and its related traits represent the archetype of a common complex phenotype. The configuration of multiple genes can range from polygenic (i.e. many genes with a relatively small contribution) to oligogenic (i.e. few genes with large measurable effects often expressed on a residual polygenic background)Reference Loktionov1. Indeed, it is this oligogenic architecture that justified all current efforts to map genes for complex phenotypes.
Prior to the mid-1990s, most of the work on genetics of human obesity had been limited to demonstrating that variation in a wide range of obesity-related phenotypes could be attributable to varying degrees to the effects of genes2. Quantitative genetics analyses (twins, siblings and family studies) have shown significant heritability, individual’s chances of being obese are increased when having obese relatives, with estimates of heritability generally ranging from 30% to 70%. Different sets of monozygotic twins overfed showed differences in the degree to which excess calories were stores of fat, but the tendency towards increased adiposity within each set of twins were very similar, indicating that genetic factors play a major role in the individual susceptibility to gain weight in a specific environmentReference Farooqi3–Reference Hunter5.
From the late 1980s through the early 1990s, segregation analysis was utilised in an effort to dissect further the genetic component underlying human obesity by attempting to detect the effects of anonymous major genes on obesity-related phenotypes. A number of these studies have reported consistent evidence of an anonymous major gene effect accounting for as much as 40% of the variation in body mass index (BMI) or fat mass in a variety of populations2.
By the early to mid-1990s, researchers began to focus on the goal of identifying the specific genes involved. Initially, such efforts to identify specific genes influencing complex traits such as those associated to obesity relied on the use of a priori selected candidates genes. Such candidates genes are selected on the basis of their perceived role or function in biochemical pathways relevant to the specific phenotype of interest (e.g. the structural gene for a circulating protein). The conservation of hypothalamic pathways in rodents and humans has certainly aided in choosing suitable candidates genes. All the spontaneously occurring obesity mutations in mice either have been found to harbour functionally relevant mutations in humans too or have led to the identification of a system/pathway in which other genes were found to be mutated. The tools recently developed allow easy determination of most gene polymorphisms from a blood sample, especially single-nucleotide polymorphism (SNP). This advancement opens a new era where research work devoted to study the interaction among diets, metabolic variables, disease risk factors and gene polymorphism can be carried out. According to the last available version of the Gene Obesity Map (2004) there are more than 600 genes, markers and chromosomal regions involved in body weight regulation and obesity developmentReference Perusse, Rankinen, Zuberi, Chagnon, Weisnagel and Argyropoulos6.
Genetics of body weight regulation
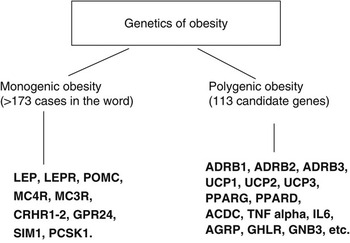
Body weight regulation and stability depends upon an axis with three interrelated components: food intake, energy expenditure and adipogenesis, although there are still many unknown features concerning fuel homoeostasis and energy balance. There are 358 studies on obese humans reporting positive associations with 113 candidate genes. Among them, 18 genes are supported by at least five positive studiesReference Perusse, Rankinen, Zuberi, Chagnon, Weisnagel and Argyropoulos6 (Fig. 1). Here we will summarise a number of mutations in genes encoding factors regulating food/energy intake and factors implicated in energy expenditure and adiposity.

Fig. 1 Genetics of obesity
Genes encoding factors regulating food/energy intake
It was generally accepted that hypothalamic and brain stem centres are involved in the regulation of food intake and energy balance but information on the relevant regulatory factors and their genes was scarce before the last decadeReference Loktionov1. Insulin remained the only candidate for the key role in body weight regulation for a long time. The discovery of leptin is one of the most exciting findings of the last decade. This cytokine-like peptide mainly expressed by adipocytes is now believed to be a key regulator of fat metabolism and energy intake. Leptin is the product of human homologue of mouse ‘obese’ gene, whose homozygous mutation caused hereditary obesity in mice (monogenic).
The mechanisms participating in the effects of leptin and other peptides on food intake and body weight regulation are now becoming clearer. Certain areas of the hypothalamus are rich in specific receptors binding regulatory peptides and triggering central regulatory mechanisms. Factors acting at the central nervous system level include neuropeptide Y (NPY), corticotropin-releasing hormone, proopiomelanocortin, α-melanocyte-stimulating hormone, agouti-related protein, melanin-concentrating hormone, cocaine- and amphetamine-regulated transcript and other molecules. Interaction between them involving complex neuronal mechanisms eventually influence the behaviour and provide important links with neuroendocrine regulation of other vital functions of the organism2.
Studies in humans have failed to find leptin or any other mutant gene to be the unique ‘obesity gene’. Conversely, multifactorial patterns involving actions of numerous polymorphic gene products now look more likely. Evidence is accumulating that most of these genes encoding central peptide factors as well as their receptors (leptin receptors, melanocortin receptors, NPY receptors) are polymorphic. Dominant inheritance of obesity conferred by missense, nonsense and frameshift mutations in the melanocortin 4 receptor (MC4R) gene has been extensively reported in many populations including French, English, German, American, Italian and Spanish individuals2, Reference Kublaoui and Zinn7–Reference Lubrano-Berthelier, Dubern, Lacorte, Picard, Shapiro and Zhang10. It has been estimated that 1–6% of extremely obese individuals harbour functionally relevant MC4R mutations. More than 70 mutations of the MC4R, 57 non-synonymous, 5 nonsense and 10 frameshift mutations, have been reported, many of them associated with dominant inheritance of obesityReference Kublaoui and Zinn7, Reference Hinney, Bettecken, Tarnow, Brumm, Reichwald and Lichtner9. Functional studies showed that many of the missense mutations also lead to a loss-of-function of the MC4R. Meanwhile, other mutations (i.e. Thr-11-Ser, Arg-18-Cys) and two polymorphisms (Val-103-Ile, Ile-251-Leu) did not modify the function of the MC4R in vitro 2.
NPY is released from the arcuate hypothalamic nucleus in fasting or in hypoglycaemia situations, its secretion being inhibited after food intake. The Leu7Pro polymorphism in the NPY gene appears to be implicated in lipid metabolism regulation. Some works reported that carriers of the Pro7 allele had higher NPY levels and also body fatness11.
A number of peptides synthesised along the gastrointestinal tract also affect food intake. They include ghrelin (orexigenic peptide mainly produced in the stomach), cholecystokinin (produced in the small intestine acting as a short-term satiety signal) and peptide YY3-36 (produced in the colon and suppressing appetite for up to 12 hours). Exploration of these signalling pathways has started and it is becoming clear that polymorphism in relevant genes may have important functional consequences. For the ghrelin receptor gene, two SNPs were reported: Ala204Glu and Phe279Leu, which selectively impair the constitutive activity of the receptor in humans leading to short stature and obesity that apparently develop during pubertyReference Holst and Schwartz12.
Moreover, the identification of relevant genes related to food preferences has just started. A novel family of 40–80 human and rodent G protein-coupled receptors expressed in taste receptor cells of tongue and palate epithelia has been identified. Taste 2 receptors (T2Rs) have been shown to function as bitter taste receptor and T1Rs as putative receptor for sweet taste. There is no information on polymorphism in the T1R family genes while some SNPs in T2R have been reportedReference Hinrichs, Wang, Bufe, Kwon, Budde and Allen13, Reference Kim, Jorgenson, Coon, Leppert, Risch and Drayna14. Rapid progress has been made in this field to elucidate the genetic mechanism controlling formation of food preferences.
Genes encoding factors implicated in energy expenditure
The adaptive thermogenesis in humans is closely related to the active mobilisation of lipids from fat tissues and demand special interest in relation to obesity. Central neural pathways responsible for the food intake and energy expenditure regulation are tightly interconnected. The peripheral transmission of central commands to the fat stores is mediated by the sympathetic nervous system. β-adrenoceptor gene families (ADRB2, ADRB3, ADRB1) are intensively studied candidate genes in the obesity field for their participation in energy expenditure regulation.
The β2-adrenergic receptor gene (ADRB2) encodes a major lipolytic receptor protein in human fat cells. Two common polymorphisms of the ADRB2 gene, characterised by an amino acid replacement of arginine by glycine in codon 16 (Arg16Gly) and glutamine by glutamic acid in codon 27 (Gln27Glu), have been explored in several diseases such as hypertension and obesity2, Reference Macho-Azcarate, Marti, Gonzalez, Martinez and Ibanez15–Reference Corbalan, Marti, Forga, Martinez-Gonzalez and Martinez18. A relationship between the Arg16Gly polymorphism and an altered function of the ADBR2 has been reported leading to a decreased agonist sensitivity. Meanwhile, the Gln27Glu variant was also found to be linked to obesity in some populations. In men, the 27Glu allele has been associated with increased BMI and subcutaneous fat and with elevated leptin and triglycerides levels, while in women, the 27Glu variant was reported to be linked to increased BMI, body fat mass and waist-to-hip ratioReference Macho-Azcarate, Marti, Gonzalez, Martinez and Ibanez15–Reference Corbalan, Marti, Forga, Martinez-Gonzalez and Martinez18. However, other studies in Caucasians (Danish men, Austrian women and German subjects) found no association between the Gln27Glu variant of the ADRB2 gene and obesity2, Reference Perusse, Rankinen, Zuberi, Chagnon, Weisnagel and Argyropoulos6.
The β3-adrenergic receptor (ADRB3) protein plays a role in adipocyte metabolism mediating the rate of lipolysis in response to catecholamines and their agonists have potential anti-diabetes and anti-obesity properties2, Reference Marti, Corbalan, Martinez-Gonzalez and Martinez19–Reference Ochoa Mdel, Marti and Martinez22. A common polymorphism in this gene, characterised by an amino acid replacement of tryptophan by arginine at position 64 (Trp64Arg), has been identified and may be linked to lower lipolytic activity and account for lipid accumulation in the adipose tissue. A number of articles have reported the relationship between the Trp64Arg variant of the ADRB3 and obesity-related phenotypes. With regard to BMI, more than nine studies have shown a statistically significant association between BMI and the Trp64Arg polymorphism in a variety of populations from 134 to 856 subjects. Besides, two meta-analyses examining the effect of this mutation on BMI have been published for Caucasian populations. One includes 2447 subjects and the significant BMI difference among carriers and non-carriers of the mutation is 0.30Reference Ochoa Mdel, Marti and Martinez22. The second includes 7399 subjects but the results are negative for the associationReference Ochoa Mdel, Marti and Martinez22. This polymorphism has been associated with abdominal/visceral fat obesity in several populations such as Caucasians and Japanese subjects. Similarly, several studies carried out among Mexican American, Japanese and Caucasian women have shown that carriers of the Arg allele had a higher BMI and lower reduction in visceral fat after weight loss2, Reference Park, Kim and Lee23. Some authors, however, failed to reproduce the finding on β-adrenoceptors gene variants and further confirmation is required.
More recently, variants of the ADRB1 gene have been studied in relation to obesity. The ADRB1 is also considered a potential candidate gene for obesity because of its role in catecholamine-induced energy homoeostasis. Stimulation of ADRB1, a member of G-protein-coupled receptors, mediates energy expenditure and lipolysis in adipose tissue. In the C-terminal intracellular G-protein-coupling domain of ADRB1, a polymorphism Arg389Gly has been identified, with the Arg389 allele vs. the Gly389 allele displaying enhanced adenylate cyclase activity in vitro Reference Li, Chen, Srinivasan, Boerwinkle and Berenson24, Reference Linne, Dahlman and Hoffstedt25.
Whereas β-adrenoceptors participate in the regulation of adaptive thermogenesis as a component of sympathetic responses, uncoupling proteins (UCPs) are involved in the modulation of heat-generating uncoupled respiration at the mitochondrial level. They represent a family of carrier proteins localised in the inner layer of mitochondrial membranesReference Ochoa Mdel, Marti and Martinez22. There are different members: UCP1, mostly expressed in brown adipose tissue, has a role in thermogenesis, UCP2 is ubiquitously present in any tissue and UCP3 is mainly expressed in the skeletal muscle and brown adipose tissue. Their putative role as ‘uncoupling proteins’ has been intensively explored; like UCP1, UCP2 mediates mitochondrial proton leak releasing energy stores as heat and thereby affecting energy metabolism efficiencyReference Ochoa Mdel, Marti and Martinez22. The actual functions for UCP2 and UCP3 proteins are still under investigation. It has been proposed that uncoupling proteins act as regulators of energy metabolism, they being fatty acid transmembrane transporters in the mitochondria facilitating proton exchangeReference Ochoa Mdel, Marti and Martinez22.
Moreover, a number of human studies indicated the relationship between UCP polymorphisms and exercise efficiency, resting energy expenditure, substrate oxidation, energy metabolism, BMI, obesity risk, type 2 diabetes risk, leptin, fat accumulation, body weight changes, physical activity and so on2. These observations have led to the consideration of UCP2 and UCP3 as candidate genes for obesity, given their function in the regulation of fuel metabolism.
Several UCP2 gene variants have been described: a G/A mutation in the promoter region −866G/A, a valine for alanine substitution at amino acid 55 in exon 4 (Ala55Val) and a 45 base pair insertion/deletion in the untranslated region of exon 8Reference Ochoa Mdel, Marti and Martinez22, Reference Marti, Corbalan, Forga, Martinez-Gonzalez and Martinez26, Reference Zurbano, Ochoa, Moreno-Aliaga, Martinez and Marti27. The association between these polymorphisms of the UCP2 and various aspects of obesity have been intensively studied. From the literature, it seems that allele G in the promoter region of UCP2 increases obesity risk while it affords relative protection for type 2 diabetesReference Ochoa Mdel, Marti and Martinez22. Meanwhile the Ala55Val polymorphism has shown to be associated with increased exercise efficiencyReference Marti, Corbalan, Forga, Martinez-Gonzalez and Martinez26. However, results concerning the exon 8 insertion allele of the UCP2 gene have been inconsistent. While no association with obesity was observed in a number of studies conducted in several populations, significant associations between the exon 8 insertion of the UCP2 gene and BMI or fat mass or presence of obesity were found with P values of 0.01, 0.001, 0.002 and 0.005Reference Ochoa Mdel, Marti and Martinez22, Reference Zurbano, Ochoa, Moreno-Aliaga, Martinez and Marti27. A sixth study reported an association of the exon 8 insertion allele with sleeping metabolic rate (P = 0.007)Reference Marti, Corbalan, Forga, Martinez-Gonzalez and Martinez26, Reference Zurbano, Ochoa, Moreno-Aliaga, Martinez and Marti27.
There are also several UCP3 gene variants. In linkage studies, some of them have been associated to a higher obesity riskReference Ochoa Mdel, Marti and Martinez22. Specifically, the −55C/T polymorphism in the promoter region of this gene has been associated with an elevated BMI, an increased level of adiposity or a greater waist-to-hip ratioReference Alonso, Marti, Corbalan, Martinez-Gonzalez, Forga and Martinez28. However, other authors have not found any relationship between this polymorphism and a higher risk of obesity or changes in the metabolic rateReference Alonso, Marti, Corbalan, Martinez-Gonzalez, Forga and Martinez28. Even some studies have reported an inverse correlation with BMI and the presence of −55C/T polymorphismReference Alonso, Marti, Corbalan, Martinez-Gonzalez, Forga and Martinez28.
Genes encoding factors implicated in adipogenesis
The last group of genes acting in connection with the peripheral regulation of energy expenditure comprises the transcription factors leading to adipogenesis and adipocytes differentiation. The key actor is peroxisome proliferator-activated receptor γ, particularly the adipose-specific isoform PPARG2. In a meta-analysis examining the Pro12Ala polymorphism in 19 136 subjects, a positive association with BMI was foundReference Ochoa Mdel, Marti and Martinez22. In our study, the frequency of the Ala allele, similar to other Caucasian populations, was higher in obese subjects (allelic frequency 0.13) than in controls (0.08), suggesting that this polymorphism was associated with obesityReference Marti, Corbalan, Martinez-Gonzalez, Forga and Martinez29. There is also information on the functional role of PPARG gene variants. Some mutant proteins appear to have a reduced activityReference Ochoa Mdel, Marti and Martinez22.
Monogenic obesity
The clinical features of human subjects with leptin (or leptin receptor), proopiomelanocortin (POMC), MC4R and proprotein convertase 1 (PC1) deficiencyReference Zurbano, Ochoa, Moreno-Aliaga and Marti30 are often associated with an obese phenotype.
Congenital human leptin deficiency has been identified in subjects showing severe early-onset obesity (8 years and 86 kg, or 2 years and 29 kg) with intense hyperphagia and undetectable levels of serum leptin due to a frameshift mutation in the ob gene (deletion G133) in a homozygosis, which resulted in a truncated protein not secretedReference Farooqi3. Children with leptin deficiency had also profound abnormalities in T-cell number and function consistent with high rates of infection and childhood mortality from infections. Leptin therapy in these subjects has a major effect on appetite with normalisation of hyperphagia and reductions body weight. Leptin receptor-deficient subjects were also found, with the phenotype being similar to those with leptin deficiency. The birth weight was normal, but a rapid weight gain was seen in the first months of life, with severe hyperphagia and aggressive behaviour when food was deniedReference Farooqi3, Reference Zurbano, Ochoa, Moreno-Aliaga and Marti30. Basal temperature and resting metabolic rate were normal and they were normoglycemic with mildly elevated plasma insulin as reported for leptin-deficient subjects. But specifically, they also debut with mild growth retardation and impaired basal and stimulated growth hormone secretionReference Farooqi3.
Homozygous and heterozygous subjects for mutations in POMC have been found. In neonatal life these subjects showed adrenocorticotropic hormone (ACTH) deficiency (the POMC gene encoded ACTH and other peptides), the children have red hair and pale skin due to the lack of melanocyte-stimulating hormone (MSH) action at the melanocortin-1 receptors in skin and hair folliclesReference Farooqi3. The POMC deficiency is associated with hyperphagia and early-onset obesity due to the lack of activation of the melanocortin-4 receptor.
Since 1998 many groups have reported at least 70 mutations in MC4R mostly associated with severe early-onset obesityReference Kublaoui and Zinn7–Reference Lubrano-Berthelier, Dubern, Lacorte, Picard, Shapiro and Zhang10. Other clinical features of MC4R mutation carriers are hyperphagia, accelerated linear growth in children and marked increase in bone mineral density. Probands with homozygous MC4R mutations show more severe obesity than their heterozygous relatives; thus, the mode of inheritance is codominantReference Farooqi3. The prevalence of pathogenic MC4R mutations varied, being the highest in obese adults selected for severe childhood obesity, suggesting that both factors, severe obesity and early age of onset, may be markers of MC4R mutations. It has been also shown that pathogenic MC4R mutations are more prevalent in northern European populations than in the Mediterranean or even Asian populations. The figures for prevalence are always quite low, for example 68 obese patients with pathogenic MC4R mutations out of 2000 or even one mutation in a sample of 3000 obese patients in another published studyReference Kublaoui and Zinn7. Functional analyses of these mutations allow us to classify them on the basis of their effects on receptor signalling. For instance, some authors demonstrated that mutations that caused intracellular retention of the receptor in vitro were associated with earlier age of onset and greater severity of obesity than other mutations.
The extremely low prevalence of pathogenic MC4R mutations in the general population underlies the fact that the obesity epidemic is not an epidemic of new mutations.
Subject carriers of PC1 mutations mainly have severe early-onset obesity, impaired prohormone processing and hypocortisolaemiaReference Farooqi3. Another clinical feature is small intestine dysfunction, which may result from an erroneous maturation of propeptides within the PC1-secreting cells along the gutReference Farooqi3.
Syndromic forms of obesity
There are about 30 rare syndromes caused by discrete genetics defects or chromosomal abnormalities with obesity as a clinical feature in association with mental retardation, dysmorphic features and organ-specific abnormalities (i.e. pleiotropic syndromes)Reference Farooqi3, Reference Bell, Walley and Froguel4. At least four syndromes show severe hyperphagia and/or other signals of hypothalamic dysfunction, indicating a plausible origin at the level of the central nervous system. The most frequent of these syndromes is Prader-Willi syndrome (prevalence 1/25 000) characterised by obesity, hyperphagia, hypotonia, mental retardation, short stature and hypogonadiotropic hypogonism. It is usually caused by lack of the paternal segment 15q11.2-q12, either through deletion of the paternal critical segment (75%) or through loss of the entire paternal chromosome 15 with the presence of two maternal homologues in 22% of patients (uniparental maternal disomy). One suggested mediator of the obesity phenotype is ghrelin, the stomach-secreted peptide that increased appetite by interacting with POMC/CART (cocaine- and amphetamine-regulated transcript) and NPY hypothalamic neurons whose levels are high in Prader-Willi syndrome patientsReference Farooqi3, Reference Bell, Walley and Froguel4.
The loss of the single-minded homologue 1 (SIM1) gene in chromosome 6 has been associated with hyperphagia in syndromic obesity. This gene encoded a protein which is a regulator of neurogenesis. In humans, deletion or disruption of the SIM1 region results in either ‘Prader-Willi-like’ phenotype or an early-onset obesity linked to hyperphagiaReference Farooqi3, Reference Bell, Walley and Froguel4.
Albright’s hereditary osteodystrophy is an autosomal dominant disorder due to mutations in GNASI, which encodes for α-subunit of the stimulatory G protein (Gs α). Maternal transmission of GNASI mutations leads to Albright’s hereditary osteodystrophy (obesity, short stature, round facies, ectopic tissue ossification) plus resistance to several hormones – such as parathyroid hormone – which activate Gs in their target tissues, while paternal transmission leads only to Albright’s hereditary osteodystrophy phenotype (pseudopseudohypoparathyroidism)Reference Farooqi3, Reference Bell, Walley and Froguel4.
The origin of obesity is more complex in Bardet-Biedl syndrome (prevalence of BBS < 1/100 000). It is an autosomal recessive syndrome characterised by central obesity (75%), polydactily, learning disabilities, rod–cone dystrophy, hypogonadism and renal abnormalities. The Bardet-Biedl syndrome is a genetically heterogenous disorder that is known to map to at least eight loci, seven of which have now been identified at the molecular level (mutations in BBS1–BBS11 genes)Reference Farooqi3, Reference Bell, Walley and Froguel4, Reference Chiang, Beck, Yen, Tayeh, Scheetz and Swiderski31.
The molecular causes that underlie the aetiology of syndromic obesity are far more complex than that for monogenic obesity. Positional genetic strategies have led to the recent identification of several causative mutations responsible for these syndromes; however, in many cases the defective gene product is a ubiquitously expressed protein of unknown function.
Genetic and dietary influences at the present time
The interaction of functional gene polymorphisms with environmental factors (gene–environment interactions) plays a substantial role in obesity risk. Gene–environment interaction implies that in combination the effect of genotype and environment deviates from the additive or multiplicative effects of the two factors. A simple statistical approach to the analysis of gene–environment interaction when dealing with a quantitative trait (e.g. BMI) is to use the statistical test analysis of covariance, with the quantitative trait being entered as the dependent variable, genotype and environmental factors being entered as main effects and with an interaction term between the genotypic and environmental factors. If this term is statistically significant, the implication is that there is a greater or lesser additive effect and an interaction is suggested. However, this outcome could be the result of chance alone, which highlights the importance of confirmatory findings. If the P value for the interaction term is not significant, the implication is that there is a lack of interaction, but it could also simply reflect the lack of power in the study to detect an effect2, Reference Hunter5.
From a mechanistic viewpoint, interaction suggests that at the molecular level the effect or byproducts of the environmental insult modifies the molecular function of the product of the gene under observationReference Marti, Razquin and Martinez33–Reference Moreno-Aliaga, Santos, Marti and Martinez37. The higher obesity risk will come from the co-existence of both genetic and environmental influences at a high scale for a given population. On one hand individuals inherited a number of gene variants in key loci and on the other they adopt a different position on the environmental spectrum of risk by the lifestyle choices they make (e.g. low-fat vs. high-fat diets, high vs. low levels of physical activity, etc.) Thus, while the environmental factors are modifiable, the genetic factors are notReference Marti, Moreno-Aliaga, Hebebrand and Martinez32. Understanding the gene–environment relationship is one of the big challenges that is being faced.
Methodological aspects on the study of genetics of obesity
A complementary strategy to the candidate gene approach for the identification of obesity genes is the use of genome-wide linkage scans. This strategy involves the genotyping of families using polymorphic markers positioned across the whole genome followed by calculation of the degree of linkage of the marker to a disease trait. This approach does not rely on any pre-existing knowledge of the genes related to the disease. For the study of obesity, genome-wide linkage scans have been applied: in families, representative of general population and also in families chosen because they have an around obese proband. This latter approach is very useful for linkage analysis. In fact, the first genome scans for obesity located a quantitative trait locus (QTL) for leptin levels and fat mass at 2p21 in Mexican-Americans for the San Antonio Family Heart Study. Subsequently, more than 204 quantitative trait loci for obesity-related phenotypes were found on all but chromosome Y from more than 50 genome-wide scans reported since 1997Reference Bell, Walley and Froguel4, Reference Perusse, Rankinen, Zuberi, Chagnon, Weisnagel and Argyropoulos6. Most of the genome-wide scans are based primarily on BMI measurements. Several of these QTLs have been confirmed in various studies, at least 38 QTLs showed evidence of linkage in two studiesReference Perusse, Rankinen, Zuberi, Chagnon, Weisnagel and Argyropoulos6.
Nowadays, the availability of OMICs technology represents a major event for studying the genetics of obesityReference Moreno-Aliaga, Marti, Garcia-Foncillas and Martinez38–Reference Kaput43. The different biomarkers at the transcript, protein and metabolite level will hopefully develop into indicative and predictive sets of molecules, their diagnostic power cannot be uncoupled from pre-determined genetic disposition. Human genetic differences appear at the level of SNPs, copy number polymorphisms and the specific combinations of alleles (haplotypes). On top of these levels of sequence variability, epigenetic phenomena, such as DNA methylation, histone acetylation and RNA interference, add to the complexity of individually different gene regulation. Besides, the high throughput offers the opportunity to genotype more than 10 000 samples a day and enables genomewide association studiesReference Moreno-Aliaga, Marti, Garcia-Foncillas and Martinez38–Reference Kaput43. DNA hybridisation arrays represent a new tool to study the association of several different polymorphisms with the development of obesity. Gene chips containing 500 000 SNPs are already available allowing the identification of possible mutations in known and unknown genes related to obesityReference Herbert, Gerry, McQueen, Heid, Pfeufer and Illig41. This technology constitutes the appropriate tool for approaching the study of those combinations of genes and mutations that are implicated in the development of obesity in humans, as well as for establishing how environmental factors affects the onset of obesity in individuals with a specific genetic background.
Acknowledgements
Sources of funding: Financial support from the University of Navarra (LE/97), Government of Navarra (Department of Health) and Spanish Government (Health and Education Departments) is gratefully acknowledged concerning the current review.
Conflict of interest declaration: Authors declare that they have no conflict of interest in relation with this material.
Authorship responsibilities: J.A.M., M.J.M. and A.M. contributed with funds from the aforementioned organism to obtain some of the reviewed results. Also, they participated in data collection, analysis and interpretation as well as in the manuscript preparation and discussion. L.E. was substantially involved in the inclusion of the scientific contents and bibliographical search as well as in the careful reading and discussion of the final version.