


INTRODUCTION
In marine invertebrates, the reproductive cycle is defined as the set of physiological events that occur in the gonad (i.e. each gonad development stage), at intervals of cyclic time (Giese & Pierse, Reference Giese, Pearse, Giese and Pierse1977). However, in marine bivalves gametogenesis (the process of development and maturation of sex cells into gametes) is regulated by exogenous factors, and among the most studied are temperature and food availability in the water. These two variables, together with endogenous factors, such as the genetic and hormonal load, determine the reproductive cycle of an organism, resulting in a pattern of reproduction for a population (Seed, Reference Seed and Bayne1976; Giese & Pearse, Reference Giese, Pearse, Giese and Pierse1977; MacDonald & Thompson, Reference MacDonald and Thompson1986; Jaramillo & Navarro, Reference Jaramillo and Navarro1995; Thorarinsdóttir & Gunnarsson, Reference Thorarinsdóttir and Gunnarsson2003). The reproductive cycles in marine bivalves may reflect a latitudinal pattern because physical factors change with latitude (i.e. temperature) (Giese, Reference Giese1959), therefore affecting reproduction and recruitment of intertidal animals (Broitman et al., Reference Broitman, Navarrete, Smith and Gaines2001; Connolly et al., Reference Connolly, Menge and Roughgarden2001). It is for this reason that the reproductive season can be annual, semi-annual or continuous, according to the ripe/spawning stages. According to this, in both hemispheres, species living at medium (30–60°) and high (>60°) latitudes show annual reproductive cycles with just one spawning occurring mainly in spring–summer, while species living at low latitudes and close to the equator (<30°) have several spawning periods, which can occur on a semi-annual or continuous periodicity (Rand, Reference Rand1973; Clarke, Reference Clarke1987; Jaramillo & Navarro, Reference Jaramillo and Navarro1995). However, more recent studies indicate the presence of reproductive asynchrony in bivalves along their distribution range (Uribe et al., Reference Uribe, Oliva, Aguilar, Yamashiro and Riascos2012), therefore the latitudinal effect on gametogenesis still is not very clear.
Along the Chilean coast several publications have described biogeographic provinces and ecoregions (Strub et al., Reference Strub, Mesías, Montecino, Rutllant and Salinas1998; Spalding et al., Reference Spalding, Fox, Allen, Davidson, Ferdaña, Finlayson, Halpen, Jorge, Lombana, Lourie, Martin, Mcmanus, Molnar, Recchia and Robertson2007; Cardenas et al., Reference Cardenas, Castilla and Viard2009). These studies take into consideration the dominant biogeographic forcing agents defining the ecoregions, which vary from one location to another but include isolation, upwelling, nutrient inputs, freshwater influx, temperature regimes, exposure, sediments, currents and bathymetric or coastal complexity (Spalding et al., Reference Spalding, Fox, Allen, Davidson, Ferdaña, Finlayson, Halpen, Jorge, Lombana, Lourie, Martin, Mcmanus, Molnar, Recchia and Robertson2007). According to these studies our sampled locations for P. purpuratus fall within the Central Chile and Araucanian ecoregions and within the Peruvian Province (20–30oS) and the Intermediate Area (30–40oS).
The lack of both time series and large sampling distribution makes it more difficult to understand the effects of large environmental change (i.e. climate change) on the reproduction of marine invertebrates. Temperature is one of the variables that have been changing in recent decades in the aquatic environment (Levitus et al., Reference Levitus, Antonov, Boyer and Stephens2000; IPCC, 2013). In this context, there are several phenological and biogeographic studies on marine life (Edwards & Richardson, Reference Edwards and Richardson2004; Parmesan, Reference Parmesan2007; Mueter & Litzow, Reference Mueter and Litzow2008; Cardenas et al., Reference Cardenas, Castilla and Viard2009), but only few studies that emphasize the effects on gametogenesis. Although there are experimental studies on the effect of thermal increase on gametogenesis, recent data indicate that in the southern Pacific Ocean, water temperature has been decreasing considerably (Baumann & Doherty, Reference Baumann and Doherty2013), probably caused by an intensification in coastal upwelling as a result of increasing wind-stress linked to global climate change (Bakun, Reference Bakun1990; Bakun et al., Reference Bakun, Field, Redondo-Rodriguez and Weeks2010; Sydeman et al., Reference Sydeman, García-Reyes, Schoeman, Rykaczewski, Thompson, Black and Bograd2014). Garreaud & Falvey (Reference Garreaud and Falvey2009) indicated that the largest southerly wind increase occurs between 37–41°S during spring and summer, which expands the favourable upwelling conditions in that region. This has also been suggested by England et al. (Reference England, McGregor, Spence, Meehl, Timmermann, Cai, Sen Gupta, McPhaden, Purich and Santoso2014), therefore giving the possibility to lead to a regional cooling even as the world as a whole continues to warm up. In this sense, the real effect of global climate change on reproductive patterns of molluscs is still unknown in those areas where the water temperature has decreased. This phenomenon could create a reproductive vulnerability for the species exposed to these water temperature changes, since the SST is the most important abiotic factor influencing marine life (Clarke, Reference Clarke2003; Lawrence & Soame, Reference Lawrence and Soame2004).
The species Perumytilus purpuratus inhabits the intertidal rocky shore from South-east Pacific (Ecuador, Perú, Chile) rounding the southern in the South-west Atlantic (Cape Horn, Chile) until La Lobería, Argentina (41°8′S 63°10′W) (Zagal & Hermosilla, Reference Zagal and Hermosilla2001; Prado & Castilla, Reference Prado and Castilla2006). In Chile it is a key species forming extensive and dense three-dimensional matrices where up to 92 species of invertebrates inhabit (Guiñez, Reference Guiñez1996; Prado & Castilla, Reference Prado and Castilla2006; Guiñez et al., Reference Guiñez, Pita, Pérez, Briones, Navarrete, Toro and Presa2016). Therefore, this is an excellent model species to test the effect of latitude and temperature on its reproductive pattern.
Perumytilus purpuratus is a gonochoric species (separate sexes) with external fertilization. Sexual maturity is achieved at about 8–10 mm of total length (Lozada & Reyes, Reference Lozada and Reyes1981). The reproductive cycle is annual in El Tabo, Chile (33°27′S). The maturation period begins in April and ends in August (winter) (Lozada & Reyes, Reference Lozada and Reyes1981), but it is semi-annual (several spawning periods) during January and July in Taltal (25°23′S) and Huasco (28°27′S), Chile (Oyarzún et al., Reference Oyarzún, Toro, Jaramillo, Guiñez, Briones and Astorga2010). However, it is not understood whether these variations in the bivalve reproductive season have a latitudinal pattern or whether it is a species-specific population trait.
The objective of this study was to determine the reproductive cycle using qualitative and quantitative methods in seven populations of P. purpuratus distributed in a latitudinal gradient of ~2400 km of Chilean coastline, over a period of 2 years, in order to estimate the effect that latitude and temperature have had on gametogenesis, using information on SST data for the last decade. Such information allows us to understand and predict the consequences of temperature changes in the sea on the reproduction of this important intertidal mussel species of the South American coast.
MATERIALS AND METHODS
Sampling site
Perumytilus purpuratus adults (20–37 mm) were randomly collected from seven intertidal rocky coastal areas in Chile: Iquique (20°14′S), Antofagasta (23°42′S), Caleta Bolfin (23°51′S), Taltal (25°23′S), Montemar (32°57′S), Tumbes (36°38′S) and Pucatrihue (40°32′S) (Figure 1, Table 1). Sampling of reproductively active individuals in each place was conducted on a monthly basis from September 2010 to October 2012, seeking to do it at the same time. Individuals were kept alive in seawater at low temperature during their transfer to the Histology Laboratory of the Instituto de Ciencias Marinas y Limnológicas (UACh).

Fig. 1. Reproductive cycle (set of gonad development stages) of Perumytilus purpuratus from the coast of Chile based on samples collected monthly from September 2010 to October 2012. The reproductive cycles at Huasco (from Oyarzún et al., Reference Oyarzún, Toro, Jaramillo, Guiñez, Briones and Astorga2010) and Tabo (based on data from Lozada & Reyes, Reference Lozada and Reyes1981) are also shown. Methodology, GSI, gonadosomatic index; GVF, Gamete Volume Fraction; Histology, histological observations.
Table 1. Summary of gametogenic cycles and spawning periods of intertidal bivalve P. purpuratus in the analysed populations (i) Villalón (Reference Villalón1965), (ii) Oyarzún et al. (Reference Oyarzún, Toro, Jaramillo, Guiñez, Briones and Astorga2010), (iii) Lozada (Reference Lozada1964), Lozada & Reyes (Reference Lozada and Reyes1981).

Gonad dissection
To excise mussel meat from its shell, a knife was carefully run between the mantle and the lip of each valve, detaching the mantle from the shell. The byssal threads were completely removed from the byssal gland to avoid later difficulties in tissue sectioning. A piece of the middle region of the gonad of each specimen was extracted using a scalpel and then fixed in 5% formalin during 10 days and subsequently transferred to 50% ethanol. The cross-section was obtained such that the dorsal-ventral aspect passes through the digestive gland and gills at an angle across the body and such that ventral edge of the cross-section was slightly towards the posterior-ventral margin (Kim et al., Reference Kim, Ashton-Alcox and Powell2006). Each cross-section was placed in a tissue cassette and processed immediately after dissection.
Histological procedure
The samples were dehydrated using a series of ascending alcohols (50, 70, 80, 90, 95 and 100°). They were then cleared in butanol and infiltrated and embedded in paraffin, and cuts were sectioned at 7 μm thick, transverse to the axis of the gonad lobe. Sections were stained with haematoxylin & eosin using the progressive method (Humason, Reference Humason1962; Howard & Smith, Reference Howard and Smith1983).
Classification of gonad development stages
The description of the reproductive cycle represented the division of gametogenesis of P. purpuratus at different gonad development stages (GDS), in a qualitative way. The methodology was based on microscopic observations of histological sections, using the criteria and categories of Oyarzún et al. (Reference Oyarzún, Toro, Jaramillo, Guiñez, Briones and Astorga2010). Each gonad (female and male) was assigned to one of the following four GDS: (1) development, (2) mature, (3) spawning and (4) spent. It showed the dominant GDS in each month, which was obtained by calculating the greater percentage in each month.
Analysis of gamete volume fraction
Whereas the gonad of the mytilids is homogeneous in relation to the distribution of germ cells during the reproductive season (Lowe et al., Reference Lowe, Moore and Bayne1982; Oyarzún et al., Reference Oyarzún, Toro, Jaramillo, Guiñez, Briones and Astorga2010, Reference Oyarzún, Toro, Jaramillo, Guiñez, Briones and Astorga2011), the gonad fraction used in reproduction was quantitatively determined through stereometry (Weibel & Gomez, Reference Weibel and Gomez1962; Weibel, Reference Weibel1969). In order to estimate the gamete volume fraction (GVF), a representative histological section from each of the sampled females was chosen. A full sequence of the gonad was captured through digital photography. Then the number of oocytes was counted in each of the different stages of development, and also the attached gonad tissue using CPCe 4.1 software. The total hit points in each category was expressed as a percentage, thus obtaining the percentage of coverage for each type of oocyte. The criteria used to identify the oocyte stages (previtellogenic, vitellogenic and mature) and gonad tissues were based on Oyarzún et al. (Reference Oyarzún, Toro, Jaramillo, Guiñez, Briones and Astorga2010).
Analysis of gonadosomatic index
In order to estimate the gonadosomatic index (GSI), between 40 to 50 individuals were sampled every month in each locality. Each specimen had the soft tissue of the valve removed, determining sex and complete fresh weight (CFW) (±0.01 g). Both gonad lobes were dissected and weighted (±0.01 g) to determine gonad fresh weight (GFW). In these mussels gonad tissue is fully differentiated from the soma tissue.
The GSI was obtained as
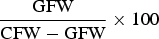 $\displaystyle{{{\rm GFW}} \over {{\rm CFW} - {\rm GFW}}} \times 100$
.
$\displaystyle{{{\rm GFW}} \over {{\rm CFW} - {\rm GFW}}} \times 100$
.
It is worth considering that in some molluscs GSI is not a good indicator of spawning or other gametogenic stages. However, Oyarzún et al. (Reference Oyarzún, Toro, Jaramillo, Guiñez, Briones and Astorga2010) showed that for this species, the GSI is a reliable indicator of maturity, mainly because of the low water content of soft tissues. According to their results, the GSI of males and females did not differ significantly.
Sea surface temperature (SST)
Daily sea surface temperature data were taken from the NOAA AVHRR OISST dataset (NOAA optimum interpolation 1/4 degree daily sea surface temperature analysis; http://www.ncdc.noaa.gov/sst/) for two periods: January 2003 to December 2012 (only in the locality of Taltal) and September 2010 to September 2012 (all localities). This product uses Advanced Very High Resolution Radiometer (AVHRR) infrared satellite and in situ SST from ships and buoys. The data include a large-scale adjustment of satellite biases with respect to the in situ SST using an empirical orthogonal teleconnection algorithm. SST data were extracted from the grid square closest to the sampling stations.
In order to know if there is a trend in SST we applied the following procedure: the daily SST from Taltal station was converted to monthly average; then a time series of the SST anomalies was also established. Finally, the time series of SST anomalies was adjusted with a linear trend between 2003 and 2012.
Statistical analysis
Results of the gonadosomatic index (GSI) and gamete volume fraction (GVF) were expressed as the mean ± SE. To evaluate the GSI as a gametic indicator and its relation to the variable surface temperature, the Pearson correlation coefficient was estimated between the variables (a) percentage of mature oocytes and gonadosomatic index (GSI) and (b) gonadosomatic index with SST (Sokal & Rohlf, Reference Sokal and Rohlf1995). The relationship between the number of spawnings per year and the latitude of each population was analysed using a simple linear regression model. Differences of GSI with respect to the time (months) for each location were performed by one-way ANOVA and a Tukey post hoc analysis (α = 0.05). The normality presumptions of the data were tested using the Kolmogorov–Smirnov test corrected by Lilliefors; and the Levene test was used for the presumptions of variance homogeneity. We used the Box–Cox method to normalize the data (GSI) which were not a normal distribution (Box & Cox, Reference Box and Cox1964). All the analyses were carried out using the statistical program STATISTICA v.7.0 (Stat Soft, Inc).
A test for reproductive status (GSI) by geographic distance was carried out. Geographic distances were determined in kilometres from the latitudinal and longitudinal coordinates using Haversine geodesic distances (Sinnott, Reference Sinnott1984). The resemblance matrix (reproductive) was generated using the Manhattan distance based on monthly averages GSI. Correlation between matrices of pairwise geographic and reproductive status was estimated using Mantel tests as implemented in R (Package vegan) (R Development Core Team, 2008).
RESULTS
Gonad development stages by locality
The populations that showed semi-annual cycles (Iquique, Antofagasta and Montemar) reported two cycles for a period of about 1 year (Figure 1; Table 1). These populations were characterized by a short gametogenic development (1–3 months), mainly during autumn/winter (Iquique = April–June and October; Antofagasta = January and June; Montemar = June–August and December). This type of reproductive pattern was characterized by brief or no recess gametogenic stages.
The populations that showed annual cycles (Caleta Bolfin, Taltal, Tumbes and Pucatrihue) had a longer gonad development (up to 7 months) during summer, autumn and part of winter, and long spawning events between late winter and spring (Caleta Bolfin = April–May; Taltal = January–June; Tumbes = October–December; Pucatrihue = February–July (Figure 1; Table 1).
Variations in GVF
Between the 2 years analysed, no differences were observed in the gametogenesis of P. purpuratus (Figure 2). Previtellogenic, vitellogenic and mature oocytes were found in all samples. However, the mature oocytes were dominant (between 15–82%). The percentage of mature oocytes had sharp decreases – spawning (e.g. Iquique = July–August 2011; Taltal = July–September 2011; Montemar = September–November 2011; Tumbes = January–February 2011; and Pucatrihue = September–October 2011; Figure 2).

Fig. 2. Gamete Volume Fraction (GVF) of previtellogenic, vitellogenic and mature oocytes in females of the bivalve Perumytilus purpuratus in populations of (A) Iquique, (B) Taltal, (C) Montemar, (D) Tumbes and (E) Pucatrihue (see Figure 1), between September 2010 and September 2012. Greyshading indicates winter season.
The fluctuations between previtellogenic and vitellogenic oocytes were similar. When the number of mature oocytes decreased, the previtellogenic oocytes were increased (e.g. Iquique = August 2011; Taltal = September 2011; Montemar = November 2011; Tumbes = July 2011; and Pucatrihue = January 2012; Figure 2).
The atresic oocytes (lysis of cytoplasmic structure and membranes in mature oocytes) were observed in mussels from Tumbes during the summer of 2012 (March–April). This phenomenon matches a sharp decrease in the number of mature oocytes in the female gonad (Figure 2D).
Variations in GSI
The percentage of gonad relative to total tissue (GSI) ranged between 4–24% (Figure 3). Correlation between the variables percentage of mature oocytes and gonadosomatic index was positive and significant (r = 0.72, N = 1025, P < 0.05). Thus, when the gonad somatic index had a sharp decrease (about 5%), it was considered that a gamete emission had occurred (spawning). In Figure 3 this is represented by a dashed line.

Fig. 3. Gonadosomatic index (mean ± SE) for Perumytilus purpuratus in the locations of (A) Iquique, (B) Antofagasta, (C) Caleta Bolfin, (D) Taltal, (E) Montemar, (F) Tumbes and (G) Pucatrihue (see Figure 1), between September 2010 and November 2012. Dotted line corresponds to the period of spawning (both sexes) corroborated against histological cuts. The vertical lines mark the annual period.
The two-way ANOVA showed significant differences for the gonad index among localities (ANOVA; F[6,11] = 145.29, P < 0.05) and months (ANOVA; F[6,11] = 91.26, P < 0.05). The post hoc test showed that during winter months (southern hemisphere) all locations had significantly higher values (May–June–July) with the exception of mussels from Tumbes, where this occurred during January (summer). Therefore, in all localities spawning was during the winter months, except for individuals from Montemar, which had important spawning in autumn (Figure 3).
Moreover, nearby populations have different reproductive conditions. Antofagasta and Caleta Bolfin (distance ~30 km) showed significant differences in GSI values (ANOVA; F[11,1] = 68.65, P < 0.05), mainly between October and February. Even though both populations showed a decrease in gonadal tissue in September (spawning), there was no spawning during summer in Caleta Bolfin, as was found in mussels from Antofagasta (Figure 3B, C).
Temperature (SST) and reproduction
There is a link between the reproductive cycles and fluctuations in SST. The localities that showed annual cycles had lower fluctuations than those localities with semi-annual cycles (Figure 4). Also, a negative linear correlation (r = −0.23, N = 131, P < 0.05) was obtained between the variables gonadosomatic index and SST in the populations analysed. Thus, when gamete emissions occur, SSTs are increased (~1°C).

Fig. 4. Annual average (±SD) sea surface temperature (SST) from October 2010 to November 2012.
The SST anomalies obtained for the locality of Taltal from 2002 to 2012, showed a tendency to decrease (Figure 5B). The mussels of this locality showed a change in the gametogenic pattern from semi-annual (between 2007 and 2008; data extracted from Oyarzún et al., Reference Oyarzún, Toro, Jaramillo, Guiñez, Briones and Astorga2010) to annual (between 2010 and 2012; data from this study) (Figure 5).

Fig. 5. Time series in the Taltal station (Chilean coast) between 2003 and 2012. (A) Daily Sea surface temperature (SST); Grey, Warm phase of El Niño–Southern Oscillation (ENSO) derived from Oceanic Niño Index obtained from NOAA's Climate Prediction Center; (B) Monthly SST anomalies. Semi-annual cycle extracted from Oyarzún et al. (Reference Oyarzún, Toro, Jaramillo, Guiñez, Briones and Astorga2010); Annual cycle data from this study.
Latitudinal analysis
The reproductive isolation did not correlate with geographic distance (Mantel's test: r = 0.03, P > 0.05). Although there was not a latitudinal pattern regarding reproductive cycles, there was a significant correlation between the number of spawns per year and the latitude of a population (r = −0.79, P < 0.05). As latitude increases (to the south), the amount of spawning along the Chilean coast decreases (Figure 6).

Fig. 6. Number of annual spawning in the analysed populations (latitude) of the bivalve Perumytilus purpuratus from September 2010 to November 2012. IQ, Iquique; AN, Antofagasta; CB, Caleta Bolfin; TT, Taltal; MO, Montemar; TU, Tumbes; PU, Pucatrihue (see Figure 1).
DISCUSSION
On a latitudinal gradient, the reproductive cycle of P. purpuratus did not follow the description found in the literature, with only annual cycles at high latitudes (Rand, Reference Rand1973; Clarke, Reference Clarke1987), even if there is a latitudinal gradient in seawater temperature along the Chilean coast (Garcés-Vargas & Abarca-del-Río, Reference Garcés-Vargas and Abarca-del Río2012). Unfortunately, studies of the latitudinal variation on the reproduction of molluscs are scarce in the literature and inconclusive because the dominant factor regulating reproductive cycles (biotic or abiotic) is still unknown (Clarke, Reference Clarke1987; Hilbish & Zimmerman, Reference Hilbish and Zimmerman1988). Nevertheless, apparently local environmental conditions are those that determine the reproductive cycles since there is no correlation between reproduction status and geographic distance (Uribe et al., Reference Uribe, Oliva, Aguilar, Yamashiro and Riascos2012). This is evident in our results when comparing nearby localities. The contrasting pattern between Antofagasta and Caleta Bolfin populations is remarkable. The Antofagasta sampling site is located inside Antofagasta Bay (23°42′16.74″S 70°25′33.53″W), while Caleta Bolfín is found 30 km south of Antofagasta Bay (23°51′43.40″S 70°30′50.21″W). This last location is under the direct influence of one of the most important and permanent upwelling currents in northern Chile (upwelling events occurring year round) (Strub et al., Reference Strub, Mesías, Montecino, Rutllant and Salinas1998; Escribano et al., Reference Escribano, Rosales and Blanco2004). Coastal localized upwelling causes large drops in SST at scales of days, and may be accompanied by increased delivery of nutrients and invertebrate larvae to benthic habitats (Graham & Largier, Reference Graham and Largier1997; Shanks et al., Reference Shanks, Largier, Brink, Brubaker and Hooff2000), so cold upwelled waters and permanent upwelling influence could be determining the annual reproductive cycle at Caleta Bolfin. The Antofagasta Bay is a semi-enclosed, southward-facing bay containing warmer waters with temperatures averaging 2–3°C more than along the adjacent upwelling coasts from outside the bay (Piñones et al., Reference Piñones, Castilla, Guiñez and Largier2007). These results suggest that in the two analysed populations the reproductive pattern is regulated by local conditions that characterize the particular areas (i.e. local temperature and/or local oceanographic conditions). A study on another important seawater variable described the effects of salinity on bivalve reproduction (Ascencio et al., Reference Ascencio, Enríquez, Martínez and Aldana2016), showing that in female oysters (Crassostrea virginica) of different sizes, it does not have any effects on their reproduction. Moreover, Zweng et al. (Reference Zweng, Reagan, Antonov, Locarnini, Mishonov, Boyer, Garcia, Baranova, Johnson, Seidov, Biddle, Levitus and Mishonov2013) showed only reduced changes in salinity along the locations sampled (20–40°S), showing in situ averages (1955–2012), which ranged between 35 to 33.25 psu. Mackas et al. (Reference Mackas, Strub, Thomas, Montecino, Robinson and Brink2006) describes that along the Chilean coast, the locations of major coastal freshwater inputs are found at over 40oS latitude.
The number of spawning events per year is higher in populations inhabiting low latitudes (Figure 4). That is, the latitude affects the reproductive output, regulating the strategy of progeny and probably optimizing indirectly the larval survival, because phytoplankton abundance (food) at high latitudes (Southern Chile) is seasonal (mainly in spring–summer) (Toro et al., Reference Toro, Paredes, Villagra and Senn1999). In comparison, at low latitudes the phytoplankton concentration is relatively constant during the year (Daneri et al., Reference Daneri, Dellarossa, Quiñones, Jacob, Montero and Ulloa2000). This is a reason why populations and numbers of individuals of P. purpuratus are more abundant towards the northern coast of Chile (Broitman et al., Reference Broitman, Navarrete, Smith and Gaines2001). Furthermore, in all populations sampled there was a mass spawning in late winter. However, the species that co-inhabit with Perumytilus purpuratus (i.e. Mytilus spp., Choromytilus choros and Aulacomya atra) have spring–summer spawning (Garrido, Reference Garrido1996; Oyarzún et al., Reference Oyarzún, Toro, Jaramillo, Guiñez, Briones and Astorga2011). We consider that a broadcast gamete emission during winter is probably a reproductive strategy for larval survival (avoiding predation of larvae), furthermore Garrido (Reference Garrido1996) reported that P. purpuratus has a lecithotrophic or facultative larvae, which could make them independent of the amount of picoplankton during early larval stages.
A single spawning per year means that there is a concentration of the annual reproductive effort in a single event. Therefore, the success of recruitment of each cohort can vary from year to year depending on the interaction with the physical environment that occurs between spawning and settlement (Calvo et al., Reference Calvo, Morriconi, Orlear and Boschi1998). This has been observed in inter-annual variability during the recruitment of such species, caused by oceanographic changes such as the El Niño–Southern Oscillation (ENSO) (Navarrete et al., Reference Navarrete, Broitman, Wieters, Finke, Venegas and Sotomayor2002). In this context, the populations that have one spawning per year are more vulnerable to environmental changes. Lawrence & Soame (Reference Lawrence and Soame2004) suggested that major environmental changes (i.e. global warming), when decoupling and altering the relationship between temperature and photoperiod, is likely to produce consequences in the development stages. A drastic change in reproductive timing of these populations can have consequences such as: (1) difficulties in gonad maturation, (2) unstable spawning season and (3) decrease in the abundance of this species. Thus, the species that inhabit the P. purpuratus constructed matrices (e.g. polychaetes and small arthropods) would be indirectly affected. Undoubtedly, populations that are most vulnerable are those inhabiting high latitudes (e.g. Southern Chile) (Figure 4). Therefore, further studies are needed to take this into account.
There is no clarity in the effect of temperature on the reproductive cycle in molluscs (Jeffs et al., Reference Jeffs, Dunphy and Wells2002; Peck et al., Reference Peck, Powell and Tyler2007; Fearman & Moltschaniwskyj, Reference Fearman and Moltschaniwskyj2010). However, our data indicate that in localities with smaller fluctuations of SST, the mussels developed annual cycles, and in localities with greater fluctuations, the individuals developed semi-annual cycles (Figure 5). In addition, our analyses indicate that gamete emissions occur when temperatures increase on average ~1°C in a short time period, that is, a few days prior to spawning. Hence, seawater temperatures have an effect on reproductive cycles, because bivalves exposed to cold temperatures, with minimal seasonal changes, would exhibit slow developmental stages (Peck et al., Reference Peck, Powell and Tyler2007), since low temperatures cause a slow cellular activity, mainly due to inactivation of proteins (Van Der Have, Reference Van Der Have2002). There is a seasonal pattern in the incorporation of lipids within the cytoplasm of the oocytes, which is mainly associated with temperature (Angel-Dapa et al., Reference Angel-Dapa, Rodriguez-Jaramillo, Caceres-Martinez and Saucedo2010). Lipids are known to play a key role in maximizing the rate of hatched eggs (fecundity), rate of larval growth and survival and rate of settled post-larvae (reproductive success) (Gallager et al., Reference Gallager, Mann and Saaki1986; Fraser, Reference Fraser1989; Palacios et al., Reference Palacios, Racotta, Arjona-Leyva, Marty, Coz, Moal and Samain2007).
The present study observed a change in the reproductive pattern of mussels from Taltal, from a semi-annual cycle to an annual reproductive cycle, in combination with a decrease in SST at this location. Considering the effect of temperature on the reproductive cycle, it is possible that the reduced seawater temperature could be a factor affecting the change of the reproductive pattern (Figure 5). Thus, these results are the first record of a change in the reproductive cycle in marine invertebrates as a result of a decrease in the SST caused by global climate change (England et al., Reference England, McGregor, Spence, Meehl, Timmermann, Cai, Sen Gupta, McPhaden, Purich and Santoso2014; Sydeman et al., Reference Sydeman, García-Reyes, Schoeman, Rykaczewski, Thompson, Black and Bograd2014; Ancapichún & Garcés-Vargas, Reference Ancapichún and Garcés-Vargas2015). There was also a change in the reproductive cycle, increasing the stage of gonad development from 3 to 7 months and showing longer spawning periods. In the literature there is also evidence of changes in reproductive cycles of P. purpuratus in Antofagasta and Tabo (Lozada, Reference Lozada1964; Villalón, Reference Villalón1965; Lozada & Reyes, Reference Lozada and Reyes1981; Uribe et al., Reference Uribe, Oliva, Aguilar, Yamashiro and Riascos2012). Based on these data, it is suggested that a decrease in water temperature would induce more prolonged cycles during some seasons (i.e. bi-annual). Although there is evidence of a change in the reproductive timing pattern, the effects on the survival and abundance of recruits, as well as the extent of endogenous factors of these mussels populations (i.e. genetic load), being able to sustain the current patterns remain unknown. Unfortunately, we have no reproductive data with a greater temporal extent that will allow us to corroborate the reproductive cycle in the other locations. Therefore, we recommend continuing to assess reproductive traits in these localities.
Recent literature showed evidence of a poleward shift of marine organisms in response to climate change (Rivadeneira & Fernández, Reference Rivadeneira and Fernández2005; Przeslawski et al., Reference Przeslawski, Falkner, Ashcroft and Hutchings2012; Poloczanska et al., Reference Poloczanska, Brown, Sydeman, Kiessling, Schoeman, Moore, Brander, Bruno, Buckley, Burrows, Duarte, Halpern, Holding, Kappel, O'Connor, Pandolfi, Parmesan, Schwing, Thompson and Richardson2013) due to ocean warming, which is causing marine species to change their breeding patterns and migration due to an advancement of the arrival of spring in the sea (Poloczanska et al., Reference Poloczanska, Brown, Sydeman, Kiessling, Schoeman, Moore, Brander, Bruno, Buckley, Burrows, Duarte, Halpern, Holding, Kappel, O'Connor, Pandolfi, Parmesan, Schwing, Thompson and Richardson2013). However, seawater warming has not been uniform around oceans. In some regions SST has cooled, such as the coast of Chile (Figure 5) (Baumann & Doherty, Reference Baumann and Doherty2013; England et al., Reference England, McGregor, Spence, Meehl, Timmermann, Cai, Sen Gupta, McPhaden, Purich and Santoso2014; Ancapichún & Garcés-Vargas, Reference Ancapichún and Garcés-Vargas2015). This recent cooling trend in the eastern Pacific sea surface temperature (Garreaud & Falvey, Reference Garreaud and Falvey2009; England et al., Reference England, McGregor, Spence, Meehl, Timmermann, Cai, Sen Gupta, McPhaden, Purich and Santoso2014; Sydeman et al., Reference Sydeman, García-Reyes, Schoeman, Rykaczewski, Thompson, Black and Bograd2014; Ancapichún and Garcés-Vargas, Reference Ancapichún and Garcés-Vargas2015), is caused by an intensification of upwelling-favourable winds in eastern boundary current systems (Sydeman et al., Reference Sydeman, García-Reyes, Schoeman, Rykaczewski, Thompson, Black and Bograd2014). The reproductive changes produced by the seawater cooling are unknown; however, the present study is one of the first to provide some evidence on the consequences that this change in seawater temperature could have on marine bivalve reproduction. Moreover, Helmuth et al. (Reference Helmuth, Mieszkowska, Moore and Hawkins2006) indicated that changes in the intertidal biota due to climate change are occurring faster than changes in terrestrial species. Therefore, there is a need to continue this kind of study, since reproductive and phenological changes should be the first biological effects of permanent environmental changes.
ACKNOWLEDGEMENTS
We acknowledge the histological support from the Instituto de Ciencias Marinas y Limnológicas.
FINANCIAL SUPPORT
FONDECYT No. 1101007 (RG) and FONDECYT 1120419 (JET) financed this study. POC was supported by a CONICYT PhD scholarship.







