



1. Introduction
In a recent article, Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) attempted to analyse the taxonomy and morphological plasticity of the earliest known paradoxidine trilobites from the Moroccan Atlas ranges and the Iberian Chains of Spain. Their study is almost entirely based on a morphometric analysis limited to linear measurements and landmark-based geometric morphometrics. They conclude that the Spanish and Moroccan specimens examined in the study, and which have been assigned to at least six different species, are all referable to a single taxon, Acadoparadoxides mureroensis (Sdzuy, Reference Sdzuy1958). Consequently, the detailed biostratigraphic scenario and evolutionary development portrayed in Geyer & Vincent (Reference Geyer and Vincent2015; Fig. 1) is not supportable.
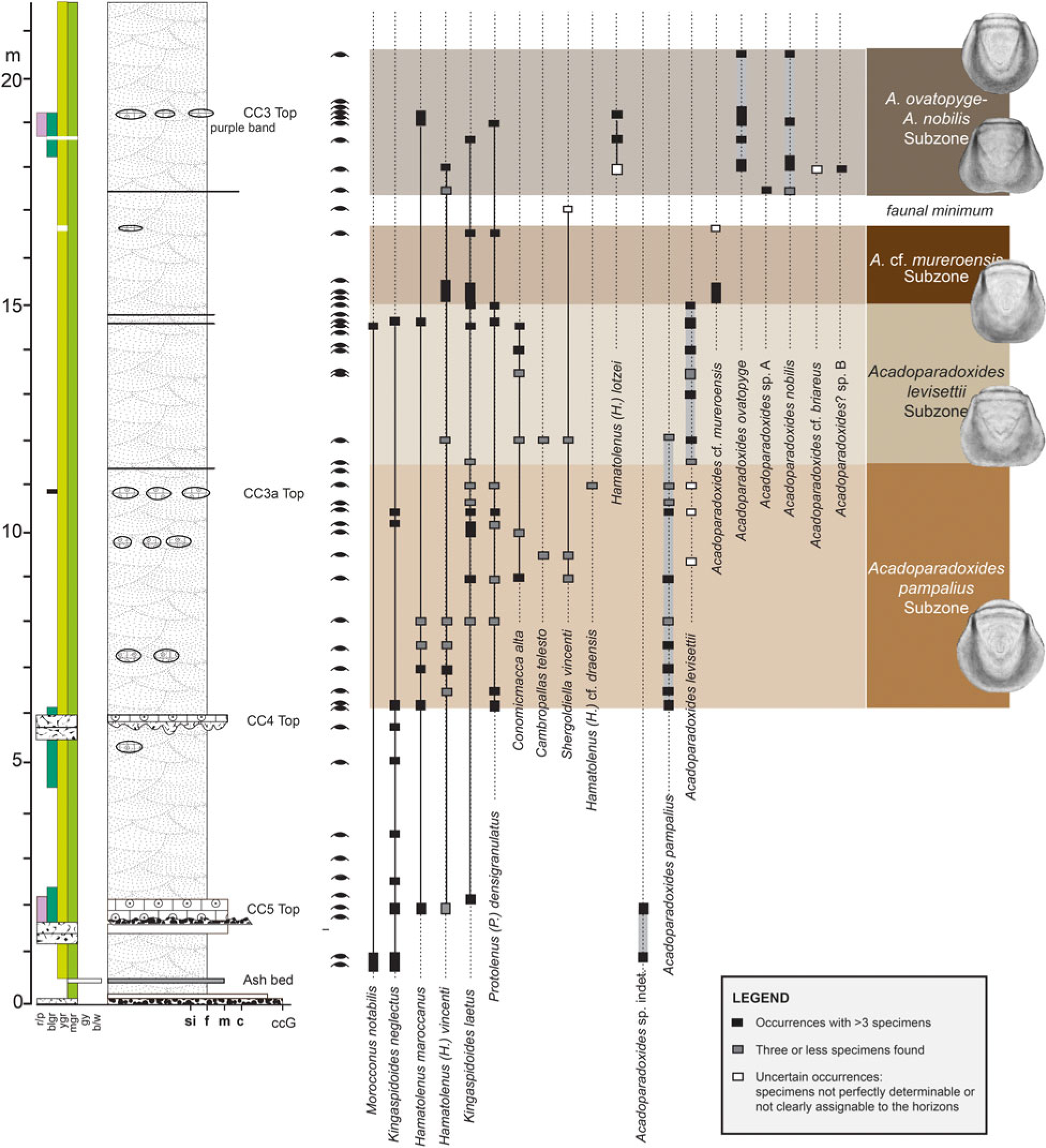
Fig. 1. Stratigraphic occurrence of Acadoparadoxides species and some index species of other trilobites in the lower part of the Brèche à Micmacca Member, Jbel Wawrmast Formation in the Tarhoucht area, Jbel Ougnate region, eastern Anti-Atlas, Morocco (based on data from Geyer & Vincent, Reference Geyer, Landing, Heldmaier, Geyer and Landing2015). Thickness in metres above the base of the Jbel Wawrmast Formation. Left column refers to rock colours (r/p – red or purple; blgr – blue-green; ygr – yellow-green; mgr – middle grey; gy – grey; b/w – buff/white). Lithologies: si – siltstone; f – fine-grained sandstone; m – medium-grained sandstone; c – coarse-grained sandstone; ccG – conglomerate. Fossiliferous horizons with trilobite remains indicated to the left of the stratigraphic columns.
We demonstrate herein that the conclusions of Álvaro et al.’s study are inappropriate, because a seeming inability to demonstrate morphologic differences, without other taxonomically important data, does not prove conspecificity. Álvaro et al.’s data are inadequate to support robust conclusions. Indeed, ignoring stratigraphic information leads to false and misleading results. This, in turn, suggests that the sole use of morphometric methods and a lack of attention to detailed stratigraphic distributions and associations of key sclerites of taxa may lead to the simplistic creation of poorly defined taxa and incorrect biostratigraphic correlations, as suggested earlier by Geyer (Reference Geyer1998). In addition, it must be emphasized that A. mureroensis is an inadequately known form that should not be used as a comparative standard in taxonomic work.
2. Paradoxides biostratigraphy
Despite being accepted as index fossils for the traditional middle Cambrian (Brøgger, Reference Brøgger1886), Paradoxides group trilobites (Paradoxidinae) have long been recognized as having a remarkably diachronous first occurrence both between and interregionally along Cambrian palaeocontinents and even different regions (e.g. Geyer, Reference Geyer1998; Sundberg et al. Reference Sundberg, Geyer, Kruse, McCollum, Pegel’, Żylińska and Zhuravlev2016). The Moroccan Atlas ranges have successions that help reconstruct the earliest evolution of Paradoxides s.l. Among the prime candidates for the earliest occurrence of the Paradoxidinae are the oldest species of Acadoparadoxides known from the Moroccan Atlas ranges, the Iberian Chains in Spain and the Holy Cross Mountains in southern Poland. All three regions feature typical West Gondwana-type lithological successions that more or less clearly record the stratigraphic truncation(s) of the lower–middle Cambrian boundary interval regression–transgression history (erroneously correlated with and termed the ‘Hawke Bay Event’, Palmer & James, Reference Palmer and James1980; see discussion in Landing & Webster, Reference Landing and Webster2018). Recognition of the precise correlation of the strata in the three regions (and their correlation into other Cambrian continents) is thus difficult, as detailed in Sundberg et al. (Reference Sundberg, Geyer, Kruse, McCollum, Pegel’, Żylińska and Zhuravlev2016). Highly resolved intercontinental correlation of this interval requires a stratigraphical standard for these strata, which indeed can be recognized in the eastern Anti-Atlas of Morocco.
3. Acadoparadoxides in the eastern Anti-Atlas of Morocco
Detailed description of the basal part of the Jbel Wawrmast Formation in the Tarhoucht area, eastern Anti-Atlas, and comparison with other sections in southern Morocco has been done by Geyer & Vincent (Reference Geyer and Vincent2015). This work not only shows the abrupt appearance of the genus Acadoparadoxides in nearly monofacial successions, but also illustrated a rapid diversification of this genus (Fig. 1).
The thick Jbel Wawrmast Formation, with its lower Brèche à Micmacca Member (Geyer & Landing, Reference Geyer, Landing, Geyer and Landing2006), is generally composed of a fairly monotonous succession of greenish-grey, fine-grained sandstones with episodic carbonate beds, layers of carbonate nodules and/or coarser-grained sandstone beds deposited in shallow-marine environments. The depositional history of the Jbel Wawrmast Formation in the central and eastern Anti-Atlas is clearly reflected by parasequences dominated by siliciclastic sediments with episodic carbonate layers (W. Heldmaier, unpub. Ph.D. thesis, Univ. Würzburg, 1998). These parasequences are visible as colour cycles that reflect a shoaling-upward development within an oxygen-stratified marine environment. The thickness of a parasequence, or colour cycle, ranges from decimetres to several tens of metres. In addition, some of the red-coloured intervals yield volcaniclastic sand grains useful as a lithostratigraphic basis for correlation and which help confirm a chronostratigraphic utility of the colour cycles. The rocks of each subunit within the parasequences are otherwise devoid of other significant variations. However, the succession of parasequences can be traced over most of the central and eastern Anti-Atlas and provide a basis for detailed regional correlation.
Transtensional and small-scale block faulting of the region took place prior to or at the onset of the deposition of the Jbel Wawrmast Formation (Landing et al. Reference Landing, Geyer and Heldmaier2006). Thus, regional or local topography influenced the fine-scale stratigraphy. Nevertheless, the cycle tops with their carbonate-rich horizons can be taken as isochrones that allow precise correlation of the lower Jbel Wawrmast Formation. Detailed cycle-by-cycle correlation over several hundred kilometres across the entire Palaeozoic Souss Basin is possible because of the lateral persistence of the colour cycles and their reliability to indicate the time of deposition.
The lower part of the Jbel Wawrmast Formation with its superb exposures in the Tarhoucht area, eastern Anti-Atlas, was studied by Geyer & Vincent (Reference Geyer and Vincent2015). This interval is characterized by five or six shoaling-up parasequences (Fig. 2). Their tops are marked by carbonate-rich intervals. The upper part of the Jbel Wawrmast Formation above these parasequences is divisible into three intervals.

Fig. 2. Parasequences/colour cycles and their boundaries in the Bou Tiouit section, Tarhoucht area, and the Zizaoun Tazemamt section (see also Fig. 3). Small photo shows abandoned part of the quarry in the Assemame section, with ‘Level 4’ of Álvaro et al. (Reference Álvaro, Esteve and Zamora2018; see Fig. 3) being the c. 30 cm thick cut at the base, and top of parasequence 3 formed by a calcareous horizon at 32.6 m. Bou Tiouit section on left with distribution of Acadoparadoxides species in lower part of the Morocconus notabilis Zone. Photos provided by T. Vincent.
The Jbel Ougnate area in the eastern Anti-Atlas has some quarries exploited for giant trilobites (with Acadoparadoxides briareus Geyer, Reference Geyer1993; e.g. Geyer et al. Reference Geyer, Landing, Heldmaier, Geyer and Landing1995). These quarries are located in the extraordinarily thick, lowest energy depositional settings of the shoaling-up sequences. More than half of the formation belongs to a single (albeit relatively long lasting) biostratigraphic zone, termed the Morocconus notabilis Zone that is equivalent to more than 80 m thickness in the Tarhoucht area (Geyer, Reference Geyer1990b; Geyer et al. Reference Geyer, Landing, Heldmaier, Geyer and Landing1995; Geyer & Landing, Reference Geyer and Landing2004). This interval brackets more than the stratigraphic ranges of the Acadoparadoxides species discussed herein. The well-known A. briareus is quarried mainly from strata c. 60–70 m above the base of the Jbel Wawrmast Formation. These levels yield a similar faunal assemblage, but slight differences in the somewhat depauperate assemblage have been described (Geyer et al. Reference Geyer, Landing, Heldmaier, Geyer and Landing1995).
Based on thorough stratigraphic collecting, Geyer & Vincent (Reference Geyer and Vincent2015) were able to demonstrate the successive occurrence of seven distinct species/forms of Acadoparadoxides in the lower two-thirds of the Jbel Wawrmast Formation within a pile of c. 70 m of strata in the Bou Tiouit section and neighbouring sections. Particularly interesting is the lowest occurrence and diversification of Acadoparadoxides in the lower third of the Jbel Wawrmast Formation. This part of the formation can be subdivided into five parasequences, in which at least six different species of Acadoparadoxides can be identified. The stratigraphic distribution of Acadoparadoxides starts with Acadoparadoxides pampalius Geyer & Vincent, Reference Geyer and Vincent2015 in the lower third of parasequence 3. This species is succeeded by Acadoparadoxides levisettii Geyer & Vincent, Reference Geyer and Vincent2015 in a discrete overlying interval. Acadoparadoxides cf. mureroensis (Sdzuy, Reference Sdzuy1958) is recorded only in a short interval. There is finally a joint occurrence of Acadoparadoxides ovatopyge Geyer & Vincent, Reference Geyer and Vincent2015, Acadoparadoxides nobilis Geyer, Reference Geyer1998, an unnamed species and possibly A. briareus Geyer, Reference Geyer1993 high in parasequence 3 (Figs 1, 2). Acadoparadoxides briareus and another, undescribed species are found higher in parasequences 2 and 1. They show a maximum abundance in an interval that was extensively quarried for extraordinarily large trilobites in the initial stage of the eastern Anti-Atlas trilobite bonanza in the 1990s (Geyer, Reference Geyer1993).
This biostratigraphic succession and the stacked occurrences with differential acmes or singular occurrences of named Acadoparadoxides species was based primarily on the Bou Tiouit section near Tarhoucht (or Taghoucht) in the Jbel Ougnate area (Geyer et al. Reference Geyer, Landing, Heldmaier, Geyer and Landing1995; W. Heldmaier, unpub. Ph.D. thesis, Univ. Würzburg, 1998). The reproducibility of this sequence was proven by other short sections in the region, such as that on the western and eastern flanks of Jbel Tazderout or Jbel Mialbene (Geyer & Vincent, Reference Geyer and Vincent2015), and subsequently confirmed by additional unpublished data recorded by T. Vincent in 2016–2018 in the Tarhoucht and Assemame areas.
The Acadoparadoxides material collected by Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) came from the Assemame quarry near Assemame (or Asemmam) in the Alnif area, c. 10 km SSE of Tarhoucht. Four fossil levels were reported from the quarry, with a similar distribution of trilobites (Zamora et al. Reference Zamora, Álvaro, Clausen, Esteve, Devaere, Clausen and Álvaro2014; Álvaro et al. Reference Álvaro, Esteve and Zamora2018). However, Acadoparadoxides is only reported from sample horizon ‘Level 4’, an interval described as being ‘30 cm thick’ at the Assemame quarry (see Fig. 2). Zamora et al. (Reference Zamora, Álvaro, Clausen, Esteve, Devaere, Clausen and Álvaro2014) and Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) figured a total of eight cranidia and ten pygidia of Acadoparadoxides from ‘Level 4’, reported as A. mureroensis (Álvaro et al. Reference Álvaro, Esteve and Zamora2018, fig. 3). Of these, six pygidia (Álvaro et al. Reference Álvaro, Esteve and Zamora2018, fig. 17f, g, k–n) show characters typical of a species referred to as A. cf. mureroensis in Geyer & Vincent (Reference Geyer and Vincent2015). The remaining four pygidia (Álvaro et al. Reference Álvaro, Esteve and Zamora2018, fig. 17a, b, e, h) are typical for A. levisettii. The thorax with attached pygidium shown in Álvaro et al. (Reference Álvaro, Esteve and Zamora2018, fig. 17l) is again typical for A. cf. mureroensis, and the pygidium is distinct from the broader, parabolic outline of the pygidia known in A. levisettii and A. pampalius (see descriptions below). This suggests that an interval equivalent to the thin A. cf. mureroensis band known from Bou Tiouit in the Tarhoucht area was sampled at Assemame, with the Assemame quarry reaching down into the top of the underlying range of A. levisettii.
The A. cf. mureroensis band is located at 15.0–15.3 m above the base of the Jbel Wawrmast Formation at Bou Tiouit (Geyer & Vincent, Reference Geyer and Vincent2015; Figs 1–3). Reinvestigation of the succession at Zizaoun Tazemamt in the Assemame area by T. Vincent (unpub. data) indicates that this horizon lies at c. 28.0–28.5 m (centred at 28.2 m) above the base of the Brèche à Micmacca Member in the Assemame section (not at 18 m as indicated in Álvaro et al. Reference Álvaro, Esteve and Zamora2018, fig. 3). Thus, this collection is within colour cycle/parasequence 3, whose top is at c. 32–33 m (Fig. 3). Thus, this only reported horizon with Acadoparadoxides in the Assemame quarry section coincides perfectly with the horizon slightly above the middle part of colour cycle/parasequence 3 in the Bou Tiouit section (at c. 15.0 to 15.3 m; Geyer & Vincent, Reference Geyer and Vincent2015, figs 7, 8) that yielded A. levisettii and A. cf. mureroensis (Fig. 3). The Acadoparadoxides material from horizon 4 at Assemame can be positively identified on the basis of the occurrence of these two species (see also additional data provided by T. Vincent at https://sites.google.com/view/acadoparadoxides on the ‘Geometric analysis of cranidia’ page, or alternatively at goo.gl/X2JCnm). The rest of the fauna is consistent with this biostratigraphic level as recorded in the Tarhoucht area.
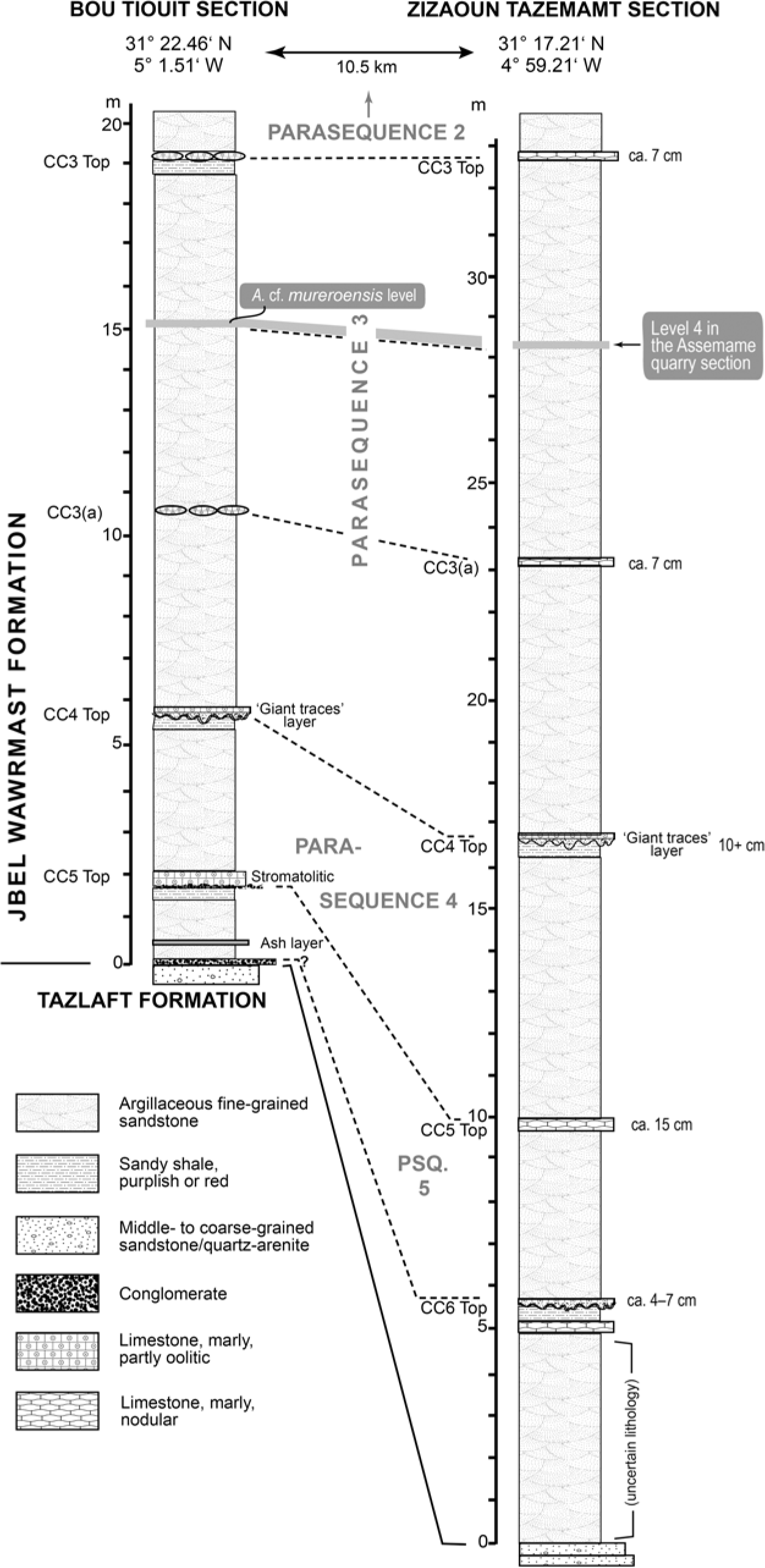
Fig. 3. Stratigraphic correlation between the Bou Tiouit and Zizaoun Tazemamt sections, Tarhoucht and Assemame areas, Jbel Ougnate region, eastern Anti-Atlas, Morocco, with reference to parasequences/colour cycles introduced by W. Heldmaier (unpub. Ph.D. thesis, Univ. Würzburg, 1998) and Geyer & Vincent (Reference Geyer, Landing, Heldmaier, Geyer and Landing2015), with simplified lithologies. Based on unpublished field data from T. Vincent (2016–2018). Abbreviations: CC3 – colour cycle 3; CC4 – colour cycle 4; CC5 – colour cycle 5; CC6 – colour cycle 6.
The entire succession in the Assemame quarry area has been re-examined recently by T. Vincent. His unpublished data confirm a perfect match with the succession in the Tarhoucht area except for slight modifications of the thicknesses and the presence of an additional parasequence at the base of the Jbel Wawrmast Formation at Assemame. This lowest parasequence records an earlier onset of the deposition following the hiatus between the Tazlaft and Jbel Wawrmast formations (Fig. 3).
The limestone bed that caps colour cycle 3(a) and marks the joint A. pampalius–A. levisettii overlap at 10.8 m in the Bou Tiouit section in the Tarhoucht area forms a small terrace at 23.2 m in the Assemame quarry section but is obscured by debris cascading from the ‘Level 4’ excavations above it (Fig. 2). The levels at the Assemame quarry below the quarried level are largely covered by scree and loose blocks dislodged by quarrying, and collecting of fossils is difficult. The section shown in Figure 3 was measured and studied at c. 150 m to the west of the quarry. It is possible that additional species of Acadoparadoxides as described from the Tarhoucht area can be collected in these rocks below level 4 of the Assemame quarry.
4. Taxonomy of the early species of Acadoparadoxides
The early–middle Cambrian boundary interval (provisional Cambrian Epoch 2–Miaolingian boundary interval) poses difficult problems for intercontinental and even regional correlation owing to its extreme endemism and local habitat control of trilobites and other taxa (Geyer, Reference Geyer2005; Sundberg et al. Reference Sundberg, Geyer, Kruse, McCollum, Pegel’, Żylińska and Zhuravlev2016). Therefore, it is puzzling that the majority of the few species of Acadoparadoxides (and some species of Eccaparadoxides) described before 1960 have been supposedly identified from rocks of different Cambrian continents, such as Avalonia, Baltica, Siberia and West Gondwana, and have often served for correlation based on those identifications (e.g. Orłowski, Reference Orłowski1959, Reference Orłowski1964, Reference Orłowski1985; Repina, Reference Repina1969; Solov’ev, Reference Solov’ev1969, Reference Solov’ev1981; T. P. Fletcher, unpub. Ph.D. thesis, Univ. Cambridge, 1972; Savitskiy et al. Reference Savitskiy, Evtushenko, Egorova, Kontorovich and Shabanov1972; Egorova et al. Reference Egorova, Shabanov, Rozanov, Savitskiy, Chernysheva and Shishkin1976; Rozanov et al. Reference Rozanov, Repina, Appolonov, Shabanov, Zhuravlev, Pegel’, Fedorov, Astashkin, Zhuravleva, Egorova, Chugaeva, Dubinina, Ermak, Esakova, Sundukov, Sukhov and Zhemchuzhnikov1992; Fletcher et al. Reference Fletcher, Theokritoff, Lord and Zeoli2005). Subsequently, a more careful examination revealed that, in most cases, the often considerable deformation of sclerites and seemingly ‘large morphological changes’ during ontogeny were used in these reports to assign specimens to well-known species without sufficient criteria. In addition, the relatively large size of Acadoparadoxides individuals leads to an infrequent preservation of well-preserved specimens compared with smaller trilobite species.
Inadequate bases for taxonomic conclusions are not limited to these earlier reports. We conclude that four principal procedural problems can be demonstrated that make the specific conclusions by Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) questionable. These problems are explained below.
4.a. Disregard of stratigraphic ranges
Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) failed to take into account the stratigraphic ranges of Acadoparadoxides species. Stratigraphic ranges can provide one of the bases for the recognition of different species. Indeed, without a detailed recording of their range, the specimens of Acadoparadoxides from the critical c. 60 m of section at Bou Tiouit and neighbouring localities could be interpreted as a single species, or better, as two species with unusually large morphological plasticity. However, careful investigation of all available data unmistakably shows that some of the particular cranidial morphotypes occur only with particular pygidial morphotypes.
As an example, the relatively ‘unspecific’ type of cranidia of A. levisettii occurs only with typical pygidia with a sub-triangular outline (i.e. with a straight or slightly indented median section at the posterior margin) between 12 and 15 m of the Bou Tiouit section and coeval intervals of parasequence 3. No pygidia with a convex curvature of the posterior margin have been found in this stratigraphic interval, whereas below this interval the pygidia all have a short straight median section dominated by a convex curvature of the posterior margin and are typical of A. pampalius. The pygidium of A. pampalius is indeed quite variable, but this variation can be attributed in part to ontogenetic size differences and an often stronger distortion of the available specimens by comparison to the high number of specimens known from A. levisettii.
Specimens of both pygidial morphotypes occur at approximately metre 12 (Fig. 1) and appear to indicate a limited stratigraphic overlap of both species. There is also seemingly a ‘transitional phase’ for these two species where no confident identification is possible for several of the specimens. Despite the transition in pygidial morphology, well-preserved cranidia of both species can be distinguished confidently. It is thus remarkable that typical pygidia of A. levisettii appear in the succession of the Tarhoucht area before typical cranidia of A. levisettii, which might be expected in a case of sympatric speciation.
Pygidia of A. nobilis are morphologically close to those of A. levisettii. However, the pygidia of A. levisettii are distinguished by their stratigraphic occurrence from those of A. nobilis, which occur higher in the sections. This is confirmed by an interval in which this ‘indented posterior margin morphotype’ does not occur, although the rock succession is monofacial and richly fossiliferous (Fig. 1). Despite their morphological similarity, the cranidia of A. nobilis can be distinguished from those of A. levisettii in having a more evenly expanding glabella (not detected in the morphometric measurements by Álvaro et al. Reference Álvaro, Esteve and Zamora2018), an anterior border that expands less distinctly from the axial line to the facial suture and slightly broader palpebral lobes.
From the stratigraphic occurrence of the specimens, the sequential appearance of the species portrayed by Geyer & Vincent (Reference Geyer and Vincent2015) is evident. Even more convincingly, species with a very similar morphology such as A. pampalius and A. ovatopyge on the one hand, or A. levisettii and A. nobilis on the other, are separated stratigraphically by ranges of species that can be clearly differentiated. This is shown by the species’ ranges in Figure 1. This close relationship of stratigraphic range and the succession of morphologically distinct sclerites of earlier named Acadoparadoxides species was not included in the study by Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) as a test of their proposed synonymy of a number of species with A. mureroensis as they interpreted that taxon.
Ranges of overlapping species indeed feature the overlap of similar sclerites, and confident determinations may be difficult in these intervals, as at 11.5–12.2 m in the Bou Tiouit section. This is not surprising because it depicts successive transitional morphologies of the relevant species through time. This sequential appearance in turn appears to portray the evolution within Acadoparadoxides. These overlapping ranges may not show an abrupt change in morphological characters. A number of intricate morphological analyses in other fossil groups show similar patterns of complicated multidirectional development in the ammonoid genus Sutneria from the upper Oxfordian to the lower Tithonian (Geyer, Reference Geyer1969) or the Late Cretaceous foraminiferan Gaudryina–Spiroplectinata plexus (Grabert, Reference Grabert1959). Application of simple morphometric methods to these fossils without the information provided by stratigraphic succession would allow the conclusion that these fossils represent only one or few ‘species’ when their stratigraphic range is ignored.
The apparent high frequency of appearances and disappearances in stratigraphic ranges of the Acadoparadoxides species is less dramatic than it may be assumed. Although radiometric dates are lacking, sequence stratigraphic analysis (W. Heldmaier, unpub. Ph.D. thesis, Univ. Würzburg, 1998) suggests that the overall time of the Jbel Wawrmast Formation deposition was not short. The ‘subzones’ bracketed by the appearances of the different species of Acadoparadoxides in the Tarhoucht area (Fig. 1) may be comparable to those recorded by the succession of the Bergeroniellinae that defines the Bergeroniellus micmacciformis, Bergeroniellus gurarii, Bergeroniellus asiaticus, Bergeroniaspis ornata and Bergeroniellus ketemensis zones and its additional subzones in the Botoman and lower Toyonian of the Siberian Platform (e.g. Astashkin et al. Reference Astashkin, Pegel, Repina, Rozanov, Shabanov, Zhuravlev, Sukhov and Sundukov1991; Rozanov et al. Reference Rozanov, Repina, Appolonov, Shabanov, Zhuravlev, Pegel’, Fedorov, Astashkin, Zhuravleva, Egorova, Chugaeva, Dubinina, Ermak, Esakova, Sundukov, Sukhov and Zhemchuzhnikov1992, Reference Rozanov, Khomentovsky, Shabanov, Karlova, Varlamov, Luchinina, Pegel’, Demidenko, Parkhaev, Korovnikov and Skorlotova2008), or even longer than the c. 32 subzones that have been discriminated in the Olenus to Acerocare zones of the Furongian of Scandinavia (e.g. Westergård, Reference Westergård1947; Henningsmoen, Reference Henningsmoen1957, Reference Henningsmoen1958).
4.b. Morphometrics
The morphometric analysis by Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) is based on linear measurement and landmark-based geometric morphometrics. The methods applied are described in some detail and require no further explanation here. These are more or less standard techniques, although several problems undercut their analysis.
Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) used both linear measurements and geometric morphometrics. It needs to be emphasized that the apparent similarity of morphologies claimed by Álvaro et al. (Reference Álvaro, Esteve and Zamora2018, fig. 5) exists mainly in the bivariate scatterplots that depict relative proportions related to glabellar length, but are clearly less convincing in the results of the principal components analyses (PCA) (Álvaro et al. Reference Álvaro, Esteve and Zamora2018, tables 3–5, fig. 6). Indeed, their scatterplots would have enveloped not only all data points for the species they dealt with, but also all species of Acadoparadoxides and most species of Eccaparadoxides, although proportional differences in the different angles to the main axis are not depicted.

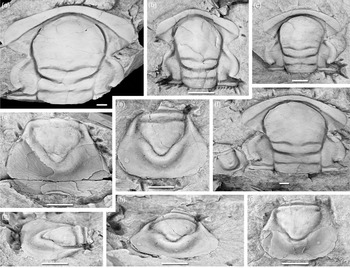
Fig. 4. Acadoparadoxides pampalius Geyer & Vincent, Reference Geyer, Landing, Heldmaier, Geyer and Landing2015. (a) MMUW 2013A-016, paratype, small cranidium, dorsal view, Bou Tiouit section, 7.0 m; (b) MMUW 2013A-178, paratype, cranidium, dorsal view, Bou Tiouit section, 6.2 m; (c) MMUW 2013A-184a, small cranidium, dorsal view, Bou Tiouit section, 6.2 m; (d) MMUW 2013A-002, paratype, immature incomplete carapace, dorsal view, Bou Tiouit section, 5–7 m; (e) MMUW 2013A-020, holotype, pygidium, dorsal view, Bou Tiouit section, c. 9.0 m; (f, j) MMUW 2013A-010, paratype, pygidium, dorsal and oblique posterior views, Bou Tiouit section, 7.0 m; (g, i, k) MMUW 2013A-024, paratype, pygidium, dorsal, lateral and oblique posterior views, Bou Tiouit section, 10.7 m; (h) MMUW 2013A-013, paratype, pygidium, dorsal view, Bou Tiouit section, 7.0 m; (l) MMUW 2013A-183a, paratype, pygidium, posterior view, Bou Tiouit section, 10.5 m. Scale bars equal 5 mm.

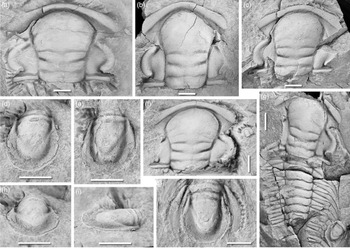
Fig. 5. Acadoparadoxides levisettii Geyer & Vincent, Reference Geyer, Landing, Heldmaier, Geyer and Landing2015. Typical cranidia and pygidia. (a) MMUW 2013A-153, paratype, nearly gerontic cranidium, dorsal view, Jbel Tazderout, pit A; (b) MMUW 2013A-152, paratype, cranidium, dorsal view, Jbel Tazderout, pit A; (c) MMUW 2013A-143, paratype, nearly complete cranidium, dorsal view, Jbel Tazderout, pit A; (d) MMUW 2013A-164, paratype, pygidium, largely exfoliated, with broad ventral doublure covered by distinct terrace ridges; dorsal and oblique lateral views, Jbel Tazderout, pit A; (e, g, h) MMUW 2013A-140, paratype, pygidium, slightly compressed dorsoventrally; dorsal, lateral and posterior views, Jbel Tazderout, pit A; (f) MMUW 2013A-145ac, paratypes, large cranidium, pygidium and fragment of thoracic segment; dorsal view, Jbel Tazderout, pit A; (j) MMUW 2013A-159a, paratype, pygidium, dorsal view, Jbel Tazderout, pit A. Scale bars equal 5 mm.

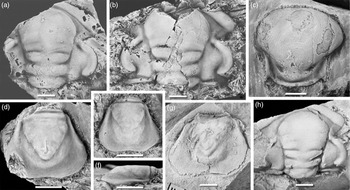
Fig. 6. Acadoparadoxides cf. mureroensis (Sdzuy, Reference Sdzuy1958). Typical cranidia and pygidia. (a) MMUW 2013A-173a, nearly complete cranidium, dorsal view, Bou Tiouit section, 15.0 m; (b) MMUW 2013A-051, nearly complete cranidium, dorsal view, Bou Tiouit section, 15.0 m; (c) MMUW 2013A-059, nearly complete cranidium, dorsal view, Bou Tiouit section, 15.3 m; (d, h, i) MMUW 2013A-058, pygidium, dorsal, posterior and oblique lateral views, Bou Tiouit section, 15.0–15.35 m; (e) MMUW 2013A-056, pygidium, dorsal view, Bou Tiouit section, 15.0–15.35 m; (f) MMUW 2013A-060a, cranidium, dorsal view, Bou Tiouit section, 15.3 m; (g) MMUW 2013A-045a, partial carapace with dislocated pygidium, moult ensemble, dorsal view, Bou Tiouit section, 15.0 m; (j) MMUW 2013A-044, detail of disarticulated posterior part of thorax with pygidium attached to posteriormost thoracic segment, dorsal view, Bou Tiouit section, 15.0 m. Scale bars equal 5 mm.
In addition, the particular sclerite features that were measured may be questioned. None of the scatterplots compare the frontal area width (tr.) with the width (tr.) of the cranidium across the palpebral lobes as a distinctive character, although both measurements were done, as is evident in the dataset in Álvaro et al. (Reference Álvaro, Esteve and Zamora2018). This proportion is regarded as an important feature in the description of the species by Geyer & Vincent (Reference Geyer and Vincent2015). For example, it distinguishes A. levisettii from the other relevant species of Acadoparadoxides discussed in Geyer & Vincent (Reference Geyer and Vincent2015). Although obscure, the data points in Álvaro et al. (Reference Álvaro, Esteve and Zamora2018, fig. 6e) indicate a strong overlap with the other species and a relatively isolated data point cloud for A. levisettii specimens. However, the situation is further obscured by data points for Spanish specimens assigned to A. mureroensis that are difficult to recognize and suffer from the uncertainties of the taxonomic validity of this species (discussed in Section 5). The material of A. mureroensis from Spain which the authors used to compare with the material from Morocco consisted of ‘the type material of A. mureroensis […] supplemented by 13 cranidia and 7 pygidia of the same species from the type locality’ (Álvaro et al. Reference Álvaro, Esteve and Zamora2018, p. 4). However, only one of the specimens is figured (a pygidium in their fig. 15), and this is the only item from the online Supplementary Material whose catalogue number has been provided. For none of these specimens has the sampling level been provided. As we know from Lotze (Reference Lotze1929) and Sdzuy (Reference Sdzuy1961), the origin of the type material is vague, and its recorded origin (‘Fp. 15’) refers to an interval of at least 17 m at the type section (see discussion in Section 5).
In the pygidia, the maximum width (tr.) was taken without considering its position relative to the length axis (which is for example more posterior in A. nobilis). This choice of the analysed parameters seems to have obscured the differences detected between the groups. The PCA plots for the pygidia (Álvaro et al. Reference Álvaro, Esteve and Zamora2018, fig. 6g–i) illustrate a distinct separation of the pygidia found in the Assemame quarry from those of A. ovatopyge and A. nobilis, which is a basis for concluding that the latter do not occur in the quarry. In addition, the pygidial landmark scores (Álvaro et al. Reference Álvaro, Esteve and Zamora2018, table 10) indicate that the Assemame quarry material is different from not only A. ovatopyge and A. nobilis, but also from A. pampalius, which is also not found in the quarry. This landmark study discriminates between the pygidia of the studied Acadoparadoxides species better than linear measurements, despite the tectonic distortion.
Furthermore, the box plots in some of the graphs are not inclusive so that the data presented are inconclusive in a strict sense. As an example, the dots for specimens in Álvaro et al. (Reference Álvaro, Esteve and Zamora2018, fig. 10a, c) are not included with two specimens and appear to be outliers (PC2 near 0.15).
Generally, morphological analyses of very similar species may lead to two differing results that may not be relevant. On the one hand, even different populations of the same species may have slightly different measures. On the other hand, not detecting distinct morphotypes (taxa) using only linear measurements is nothing surprising. Even the members of different genera of the Paradoxididae often cannot be distinguished using only classic morphometrics, although they can be easily described as undoubtedly different taxa based on their qualitative features. An example of such a case was discussed by Żylińska & Nowicki (Reference Żylińska and Nowicki2017): it was impossible to detect differences between Hydrocephalus polonicus (Czarnocki, Reference Czarnocki1927) and Acadoparadoxides slowiecensis (Czarnocki, Reference Czarnocki1927) by using solely linear measurements, despite the obvious differences that occur between these two species (e.g. the specimens illustrated in Orłowski, Reference Orłowski1985, pl. 5).
Classic morphometrics in the Paradoxididae may be useful in describing some aspects of distinguishable taxa (e.g. ontogenetic development), but clearly it is not a technique that is sensitive enough to be functional for solving taxonomic issues. Paradoxidids are known to undergo considerable changes in the proportions of the cranidia and its parts during ontogeny (e.g. specimens illustrated in Westergård, Reference Westergård1936, pl. 5), and these modifications can only be assessed if the size distributions of the investigated species are carefully considered. Thus, data derived from classic morphometrics cannot be readily taken as a proof of conspecificity.
Geometric morphometrics is a more sensitive technique, and is able to describe morphology in a more detailed way. In short, it should be more sensitive in detecting potential differences between the analysed groups. On the other hand, the outcome of geometric morphometric analysis can be distorted in several ways. First of all, not taking tectonic deformation into account may be risky: although in classic morphometrics the amount of deformation present in the sample may be negligible, geometric morphometrics is more vulnerable to such distortion. In a recent study, Nowicki & Żylińska (Reference Nowicki and Żylińska2018) have tested the utility of geometric morphometric analyses on species of Acadoparadoxides from lowermost middle Cambrian strata of the Holy Cross Mountains, Poland. Nowicki & Żylińska (Reference Nowicki and Żylińska2018) noted a study by Angielczyk & Sheets (Reference Angielczyk and Sheets2007), which concluded a comprehensive validation of numerical retrodeformation techniques in geometric morphometrics. The latter authors recognized PCA as a method that does not successfully remove tectonic variance from the dataset, with the major problem of PCA application being the distribution of tectonic variance between several components. Removal of only one component does not significantly improve the outcome, whereas removal of more than one component also results in throwing out part of the variance that is not connected with deformation.
According to the results of Nowicki & Żylińska (Reference Nowicki and Żylińska2018), the removal of a principal component that is best correlated with the length:width proportions does not improve the outcome when the axial symmetry is not restored in the specimens before the procedure. A procedure known as symmetrization improves the results, but can be safely applied only to specimens in which the deformation is relatively simple, without a high amount of noise added to the deformation, which would compromise the apparent significance of those analyses.
In fact, the Moroccan specimens are affected by a rather complex deformation. In such case, several factors can limit the output of the morphometrics, including bending of the symmetry axis (e.g. Álvaro et al. Reference Álvaro, Esteve and Zamora2018, fig. 16e: note the asymmetry of the frontal lobe) and compaction-related crushing of the carapaces (e.g. Álvaro et al. Reference Álvaro, Esteve and Zamora2018, fig. 16a, c versus fig. 16e, f). Thus, without developing some new retrodeformation techniques, it is impossible to remove the effect of such complex deformation. The amount of variation added to the sample by the deformation may be too high when the sample contains very similar specimens so that the original variation of the sample can be described as relatively low.
Secondly, Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) ignored almost all of the entire anterior part of the cranidia, and designated only five landmarks in this region (1, 2, 3, 9, 10). The curves between those landmarks could be described using semilandmarks. Lack of description of the anterior border and the insufficient coverage of the anterior branch of the facial suture may result in a misleading outcome of the analysis. Moreover, Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) applied type III landmarks in their analysis (Bookstein, Reference Bookstein1991): landmarks 4 and 7 on the cranidium and landmark 4 on the pygidium are defined as the extreme points of the palpebral lobe (cranidium) and the margin of the pygidium, respectively. Such landmarks do not represent homologous locations on the elements, thus their usage is incorrect (Bookstein, Reference Bookstein1991). Another related issue occurs in both cranidia and pygidia. Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) have used landmarks/semilandmarks only from one-half of the carapace elements. Such selection results in an artificial transition of the landmark position variance from the marginal landmarks/semilandmarks to the axial ones. In a set affected by deformation, this problem may be essential (Nowicki & Żylińska, Reference Nowicki and Żylińska2018). In addition, the landmark analyses do not reflect differences in the dorsoventral component (relief), which for example serves to indisputably discriminate the pygidia of A. pampalius from similar pygidia of A. levisettii or A. ovatopyge.
All these problems result in an indefinite outcome, where the variance derived from deformation and issues related to the correct location of landmarks obscure the true diversity of the sample.
Application of morphometrics (classic and geometric) by Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) does not detect differences between the specimens, but this fact does not indicate any proof of conspecificity. It does indicate rather that the applied technique was not able to detect potential differences. As recently emphasized by Nowicki & Żylińska (Reference Nowicki and Żylińska2018), not rejecting the null hypothesis in such studies cannot be taken as their confirmation, particularly as the analysis in Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) suffers from several problems presented above that undermine its results. In contrast, morphological differences have been described in Geyer & Vincent (Reference Geyer and Vincent2015) in a classic way, but these were not discussed in any detail in Álvaro et al. (Reference Álvaro, Esteve and Zamora2018).
Finally, a number of principal problems need to be addressed that shed some doubt on the significance of the results. The huge morphological plasticity that is seen in ‘convincingly identified’ specimens of A. mureroensis is entirely based on specimens from Spain, and this fact is discussed in some detail below. By contrast, the specimens identified as ‘Acadoparadoxides mureroensis’ from the Assemame quarry exhibit a remarkably low amount of variation (Álvaro et al. Reference Álvaro, Esteve and Zamora2018, fig. 16) when compared with the morphological disparity of other species described by Geyer & Vincent (Reference Geyer and Vincent2015). This is obviously caused by the fact that only part of colour cycle/parasequence 3 was quarried at Assemame so that this fauna is equivalent to the fauna recorded in the upper part of cycle/parasequence 3 in the Tarhoucht area (see Fig. 2).
4.c. Recognition of specific character sets
A large number of trilobites, particularly from the Cambrian, show a mosaic pattern of characters, in which the morphological features occur in different and variable combinations in closely related genera and species. This phenomenon leads to complications in recognizing and classifying taxa when the stratigraphic occurrence does not clearly display morphological trends (e.g. morphoclines, etc.) or in studies in which stratigraphic occurrence of distinct sclerites is not recorded or considered not pertinent to taxonomic evaluation.
These complications are often exceedingly difficult to overcome. One of the basic requirements is an evaluation of the ranking of key characters, which will lead to weighted application. The validity of what may initially be regarded as an arbitrary decision can then be tested by stratigraphic information. In the case of the Acadoparadoxides species dealt with herein, a number of characters occur in different combinations and allow distinction of the species, rather than species being identifiable by one or more unique distinctive characters. However, several characters can be distinguished as being of critical significance for the recognition of taxa. Examples of these include the differential developments of the facial suture and the shape of the posterior pygidial margin. In particular, the course of the anterior branch of the facial suture differs considerably between species, mainly depending on the angle at which the suture develops from the anterior margin of the ocular suture. This angle determines the point at which the suture meets the anterior cephalic margin and, thus, also determines the abaxial margin of the anterior border and, consequently, whether or not this abaxial margin of the anterior border lies in a more abaxial position than the most abaxial point of the internal branch of the ocular suture.
Additional important character sets that are not addressed in the analyses performed by Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) are the dorsoventral component, i.e. the relief, of some portions of the carapace such as the convexity of the palpebral area with its posteroproximal swelling, the concavity of the pleural fields in the pygidium or the expression of vestiges of segmentation in the pygidial axis. In addition, the anterior border and the palpebral lobes are developed differently between the species. A distinctly flattened dorsal face of the anterior border is developed in A. levisettii, A. nobilis and A. ovatopyge, whereas the anterior border is slightly flattened close to the facial suture in A. pampalius and convex in sagittal and exsagittal directions in A. cf. mureroensis.
Detailed comparisons between the species/forms of Acadoparadoxides from the eastern Anti-Atlas and a comprehensive table listing their characters and character states has been presented in Geyer & Vincent (Reference Geyer and Vincent2015, fig. 12). A brief differential characterization of these species and forms presented in the following paragraphs focuses on the characters of the cranidium and the pygidium.
Specimens dealt with below are housed in the repositories of the Naturmuseum Senckenberg, Frankfurt am Main (SMF), the Mineralogisches Museum of the Würzburg University (MMUW) and the Bayerische Staatssammlung für Paläontologie und Geologie, München (PIW).
4.c.1. Acadoparadoxides pampalius (Fig. 4)
Glabella slightly expanding and relatively slender in the posterior part; anterior cephalic margin more or less evenly curved, its dorsal face flattened near the suture. Thoracic segments with pronounced pleural spines. Pygidium sub-rounded to slightly sub-hexagonal in outline, posterior margin usually with low curvature throughout or rarely with short straight median section; pygidial axis of only 55–60% pygidial length, with vestiges of segmentation; pleural fields sunken between rhachis and lateral border so that the pygidium has a slightly cup-shaped morphology.
This is the only species of Acadoparadoxides known from the Moroccan Cambrian with a cup- or bowl-shaped pygidium, in which the lateral margin forms a slightly raised border. This distinguishes the pygidium from otherwise often similar pygidia seen in A. cf. mureroensis or A. ovatopyge. In addition, the pygidia of A. pampalius are generally broader (larger ratio of pygidial maximum width/sagittal length) than those of A. cf. mureroensis or A. ovatopyge and with the maximum transverse width lying slightly more posteriorly. The species is also characterized by its generally slender posterior section of the glabella with a slight expansion towards the anterior, and the flattened surface of the anterior border helps to distinguish the species from A. levisettii and A. ovatopyge with their overall flattened border surface. The cranidial width across the anterior border is always exactly equal to that across the palpebral lobes, which reliably distinguishes the cranidia of A. pampalius from those of A. levisettii and A. nobilis. All specimens known of A. pampalius are relatively small, and the available material indicates that the species did not attain the large size of later species such as A. levisettii. Therefore, its sclerites are only comparable in size to the small- to medium-sized sclerites from larger species in morphometric diagrams.
4.c.2. Acadoparadoxides levisettii (Fig. 5)
Glabella sub-parallel or rarely slightly expanding in the posterior part; anterior cephalic margin more or less evenly curved, its dorsal face flattened throughout. Thoracic segments with falcate pleural ends. Pygidium sub-hexagonal to sub-triangular in outline, posterior margin with distinct straight median section; pygidial axis of 60–68% pygidial length, with vestiges of segmentation in its anterior part; pleural fields sloping ventrally from axis, lateral border flat or with low convexity.
Acadoparadoxides levisettii has a relatively large morphological plasticity (which is correctly shown by the PCA plots of Álvaro et al. Reference Álvaro, Esteve and Zamora2018), but the overwhelming majority of specimens are characterized by a number of distinctive features and do not show much variation, particularly in the pygidium. Among these features are the relatively limited expansion of the glabella in the anterior versus posterior sections in large specimens; the flat dorsal surface of the anterior border with a comparatively large expansion of the exsagittal width towards the facial suture; relatively narrow fixigenae when compared with those of most other species of Acadoparadoxides in the Moroccan sections; and a sub-triangular outline of the pygidium with broadly curved posterolateral margins and a straight or nearly straight median section of the posterior margin. The cranidial width across the anterior border is always greater than that across the palpebral lobes, which distinguishes cranidia of A. levisettii from those of A. pampalius, A. cf. mureroensis and A. ovatopyge.
4.c.3. Acadoparadoxides cf. mureroensis (Fig. 6)
Glabella with sub-parallel lateral margins in the posterior part; anterior cephalic margin more or less evenly curved or sub-arcuate, with its dorsal face sagittally and exsagittally convex throughout. Thoracic segments with falcate pleural ends. Pygidium sub-triangular to sub-oval in outline, posterior margin with shallow curvature or (rarely) almost straight in median section; pygidial axis long, of 72–82% pygidial length, with vestiges of segmentation in the anterior half; pleural fields sloping ventrally from axis, lateral border flat or with low convexity.
The cranidium of A. cf. mureroensis is difficult to identify with certainty when dorsoventrally compressed. However, well-preserved specimens are characterized by and differ from similar species in the sub-parallel lateral margins in the posterior part of the glabella; the sagittally and exsagittally convex dorsal surface of the anterior border with a generally sub-arcuate curvature and a relatively minor growth in exsagittal breadth towards the facial suture; and more strongly elevated palpebral lobes than in the other species. The cranidial width between the sutures across the anterior border is either equal to or less than those that cross the palpebral lobes, which distinguishes the cranidia of A. cf. mureroensis from those of A. levisettii and A. nobilis.
The pygidium of A. cf. mureroensis can be easily distinguished from those of A. levisettii and A. nobilis by its longitudinally sub-triangular or sub-ovate outline that lacks the conspicuous posterolateral extensions seen in the latter species. There is a similarity of the shape of the pygidium with that of A. pampalius, but the pygidial axis is longer in A. cf. mureroensis than in the other discussed species described herein from Morocco.
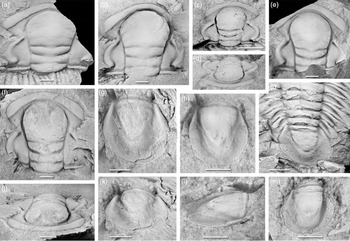
4.c.4. Acadoparadoxides nobilis (Fig. 7)
Glabella with sub-parallel lateral margins posteriorly; anterior cephalic margin more or less evenly curved, its dorsal face distinctly flattened throughout. Pygidium sub-triangular in outline, posterior margin indented; pygidial axis moderately long, of 62–74% pygidial length, occasionally with vestiges of segmentation in the anterior part; pleural fields slope ventrally from axis, lateral border flat or with low convexity.
Acadoparadoxides nobilis is probably the easiest species to recognize of those discussed herein, and is known from very well-preserved material from the High Atlas as well (Geyer, Reference Geyer1998). The cranidium of this species has an anterior border with a flat dorsal surface and with a fairly distinct kink at its posterior margin where it meets the weakly developed facial line curving forwards from the eye ridges. The palpebral lobes are weakly subdivided into a narrower adaxial and a wider abaxial part as seen in the specimens from the Anti-Atlas. The cranidial width between the facial sutures across the anterior border is greater than that across the palpebral lobes; this feature clearly distinguishes cranidia of A. nobilis from those of A. pampalius, A. cf. mureroensis and A. ovatopyge. The hypostomata of A. nobilis have a large anterior lobe so that the posterior lobe is shorter than in any of the hypostomata known from other species of Acadoparadoxides discussed herein. The pygidium is distinctly sub-triangular with a slightly indented median part of the posterior margin and with a pygidial axis that is shorter than that in the other species described from Morocco. This pygidium clearly falls out of the cluster illustrated in the PCA plots of Álvaro et al. (Reference Álvaro, Esteve and Zamora2018, fig. 10).
4.c.5. Acadoparadoxides ovatopyge (Fig. 8)

Fig. 7. Acadoparadoxides nobilis Geyer, Reference Geyer1998. Typical cranidia and pygidia. (a) PIW 97II66, paratype, incomplete cranidium, dorsal view; near Afourigh, Ouneïn area, High Atlas, sample horizon X 211; (b) MMUW 2013A-193, incomplete slightly crushed cranidium with bilobate palpebral lobes and caeca on fixigenae, dorsal view, near Afourigh, Ouneïn area, High Atlas, sample horizon X 211; (c) PIW 97II63, paratype, hypostome, dorsal view, near Afourigh, Ouneïn area, High Atlas, sample horizon X 211; (d) PIW 97II67, paratype, pygidium, dorsal view, near Afourigh, Ouneïn area, High Atlas, sample horizon X 211; (e, f) PIW 97II144, paratype, pygidium, dorsal and lateral views, near Ijoukak, High Atlas, sample horizon X 242; (g) MMUW 2013A-119a, large, compressed pygidium, dorsal view, Jbel Tazderout, pit B; (h) MMUW 2013A-067, incomplete cranidium, dorsal view, Bou Tiouit section, 18.0 m. Scale bars equal 5 mm.

Fig. 8. Acadoparadoxides ovatopyge Geyer & Vincent, Reference Geyer, Landing, Heldmaier, Geyer and Landing2015. Typical cranidia and pygidia. (a) MMUW 2013A-091, paratype, cranidium, dorsal view, Bou Tiouit section, 18.2 m; (b) MMUW 2013A-076a, paratype, cranidium, dorsal view, with incomplete cranidium of Kingaspidoides cf. frankenwaldensis (Wurm, Reference Wurm1925) in upper right corner, Bou Tiouit section, 19.0 m; (c, d) MMUW 2013A-081, paratype, immature cranidium, dorsal and oblique anterior views, Bou Tiouit section, 19.0 m; (e) MMUW 2013A-092, paratype, incomplete cranidium, dorsal view, Bou Tiouit section, 18.2 m; (f, j) MMUW 2013A-078a, paratype, cranidium, dorsal and oblique anterior views, Bou Tiouit section, 19.0 m; (g, k) MMUW 2013A-072, holotype, pygidium, dorsal and oblique posterior views, Bou Tiouit section, c. 18.0 m; (h, l) MMUW 2013A-099, paratype, pygidium, dorsal and oblique lateral views, Bou Tiouit section, 19.2 m; (i) MMUW 2013A-132, paratype, posterior thorax with attached pygidium, dorsal view, Jbel Tazderout, pit C; (m) MMUW 2013A-079a, paratype, pygidium, dorsal views, Bou Tiouit section, 19.0 m. Scale bars equal 5 mm.
Glabella slightly expanding in the posterior part; anterior cephalic margin more or less evenly curved or rarely slightly sub-arcuate, its dorsal face flattened throughout. Pygidium ovate in outline, posterior margin curved, occasionally slightly truncated medially; pygidial axis moderately long, of 66−72% pygidial length; pleural fields slope ventrally from axis, lateral border flat or with low convexity.
Acadoparadoxides ovatopyge has a glabella expanding anteriorly in its posterior part; relatively broad fixigenae; and a generally relatively evenly curved anterior cephalic margin. Provisionally, the cranidial width between the suture across the anterior border is nearly always slightly less than that across the palpebral lobes; this distinguishes cranidia of A. ovatopyge from those of A. pampalius, A. levisettii and A. nobilis. The pygidium is sub-oval in outline, with a generally relatively short pygidial axis and comparatively broad pleural areas. Although the pygidium may appear to be sometimes similar to that of A. pampalius and A. cf. mureroensis, the shorter rhachis, the broader pleural fields and the always distinct curvature of the posterior margin of the A. ovatopyge pygidium readily distinguishes it. This pygidium is so characteristic that it cannot be confused with those of A. nobilis or A. levisettii.
Most probably, A. ovatopyge is the species described from Spain as Hydrocephalus cf. harlani (Green, Reference Green1834) by Dies et al. (Reference Dies, Gozalo and Liñán2004), Liñán et al. (Reference Liñán, Gozalo, Dies Álvarez, Gámez Vintaned, Mayoral, Chirivella Martorell, Esteve, Zamora, Zhuravlev and Andrés2008) and Gozalo et al. (Reference Gozalo, Dies Álvarez, Gámez Vintaned, Zhuravlev, Bauluz, Subías, Chirivella Martorell, Mayoral, Gursky, Andres and Liñán2013).
An example of the pivotal role of a character that varies between the different species is the course of the anterior branch of the facial suture and the location of its junction with the anterior border furrow. This feature is not discussed in the study by Álvaro et al. (Reference Álvaro, Esteve and Zamora2018). A simple geometric analysis provided by T. Vincent (at https://sites.google.com/view/acadoparadoxides on the ‘Geometric analysis of cranidia’ page, or under goo.gl/L9zhEp) demonstrates its importance, and in combination with the accompanying pygidia in the relevant strata irrefutably proves the presence of three species at 6.0–16.5 m in the Bou Tiouit section.
5. What is Acadoparadoxides mureroensis?
The problematic taxonomic status and the uncertainties about the morphology of A. mureroensis have been discussed in a number of earlier publications and particularly in Geyer & Vincent (Reference Geyer and Vincent2015). However, it is necessary to outline some additional aspects of this taxon.
Acadoparadoxides mureroensis was formally introduced by Sdzuy (Reference Sdzuy1958, p. 239, pl. 1, figs 12, 13) as Paradoxides mureroensis, a name that occurred in an unpublished manuscript by Rudolf and Emma Richter dated 1928 and was used by Lotze (Reference Lotze1929, p. 34) without an earlier illustration, diagnosis and description. Nevertheless, its later use by Sdzuy (Reference Sdzuy1958) was inadvisable. The type material, housed in the Senckenberg Museum collection, Frankfurt am Main, consists of two cranidia (holotype SMF X 1370c and paratype SMF X 1370d) and one pygidium (SMF X 1370a). All of these specimens are from what is now termed the Rambla de Valdemiedes 1 section in the Iberian Chains, Aragón Province, Spain, and were most probably collected by Franz Lotze in the 1920s. The year ‘1929’ on the collection label certainly records the year in which Lotze donated the specimens to the Senckenberg Museum.
The precise horizon from which the specimens originate is unknown, although it is given as ‘Fp. 15’ on Lotze’s original label. The stratigraphic information in Lotze (Reference Lotze1961) and Sdzuy (Reference Sdzuy1961, p. 396) indicate that ‘Fp. 15’ is ‘Aus dem Bereich A7–A15 stammend, ohne daß die genaue Schicht angegeben werden kann’ [literal translation: ‘originating from the interval A7–A15 without a possibility to refer to a precise bed’]. The reason for this bulk sample interval is uncertain. Likely, the ‘interval’ was believed to be a continuous succession bounded by faults, and a precise bed-to-bed record on the stratigraphic succession of level ‘Fp. 15’ was difficult and beyond the goals of Lotze’s pioneer work. Lotze (Reference Lotze1961) and Sdzuy (Reference Sdzuy1961) indicated that the interval A7–A15 in the Rambla de Valdemiedes 1 section includes nine different fossiliferous levels in the 17+ m succession, and subsequent studies indicate structural complications as well (e.g. Liñán & Gozalo, Reference Liñán and Gozalo1986; Gozalo et al. Reference Gozalo, Dies Álvarez, Gámez Vintaned, Zhuravlev, Bauluz, Subías, Chirivella Martorell, Mayoral, Gursky, Andres and Liñán2013).
The two cranidia of Sdzuy (Reference Sdzuy1958) are preserved in sandstone with a somewhat different lithology and obviously do not come from the same bed. The piece with the pygidium has the same lithology as cranidium SMF X 1370c, which is assigned as the holotype (refigured here in Fig. 9a, c). Both the holotype and the pygidium clearly show worn edges and scratches. They were almost certainly collected from the scree as assumed by K. Sdzuy (pers. comm. to GG, 1981). It is possible, but not at all certain, that the cranidia and the pygidium belong to the same species.

Fig. 9. Acadoparadoxides mureroensis (Sdzuy, Reference Sdzuy1958), type series from the Rambla de Valdemiedes 1 section, Iberian Chains, northern Spain. (a, c) SMF X 1370c, slightly distorted crandium, holotype, dorsal and oblique lateral views; (b, d) SMF X 1370d, slightly distorted cranidium, paratype, dorsal and oblique lateral views; (e, h) SMF X 1370a, pygidium, dorsal and posterior views; (f) SMF X 1370e, slightly distorted partial cranidium, dorsal view; (g) slightly distorted librigena, associated with SMF X 1370e, dorsal view; (i) small cranidium of Conomicmacca alta (Liñán & Gozalo, Reference Liñán and Gozalo1986) associated with SMF X 1370e, distorted, dorsal view; (j) slightly distorted partial cranidium associated with SMF X 1370e, dorsal view, with Trematobolus simplex (Vogel, Reference Vogel1962) (left), internal mould of valve. Scale bars equal 5 mm.
The holotype cranidium (SMF X 1370c) is distinctly compressed and obliquely distorted. Among other preservational problems associated with the holotype, this deformation has made the palpebral lobe nearly sub-equal in width to the ocular platform on the left-hand side, whereas it is clearly narrower (tr.) on the right-hand side. This deformation limits any utility of the available morphological characteristics. It is also worth mentioning that the specimen underwent a differential deformation so that the longitudinal axes on the right-hand side and the left-hand side are not parallel and slightly, but differently, curved (Fig. 9a). Paratype SMF X 1370d is a cranidium (preserved as part and counterpart) with considerable compression along the longitudinal axis (Fig. 9b, d).
The Senckenberg Museum collection also houses partial cranidia labelled as ‘Paradoxides mureroensis’, which according to the label are from the same locality and stratum (‘Fp. 15’) and were certainly collected by Lotze along with the type material. One of these cranidia (Fig. 9f) is associated with a nicely preserved thoracic segment not mentioned by Sdzuy (Reference Sdzuy1958, Reference Sdzuy1961). The same slab has a librigena (SMF X 1370e; figured by Sdzuy Reference Sdzuy1961, pl. 16, fig. 12; refigured in Fig. 9g), which represents the best preserved free cheek known from A. mureroensis if it actually belongs to the same species.
A few slightly better preserved cranidia were figured by Sdzuy (Reference Sdzuy1961, pl. 16, figs 7, 10, 11), but all are distinctly dorsoventrally flattened and mostly from the collective sample horizon ‘Fp. 15’. An exception is specimen L3242 from the Münster University collection (Sdzuy, Reference Sdzuy1961, pl. 16, fig. 7), which is listed as originating from ‘Fp. 7’ (as well as cranidium L 3239, Sdzuy, Reference Sdzuy1961, pl. 16, fig. 3). These specimens came from level A8, which is well above the base of the occurrences of A. mureroensis in level A7 according to Sdzuy (Reference Sdzuy1961, pp. 32, 33, fig. 10). Level ‘Fp. 7’ (= level A8) also yields specimens of Hamatolenus lotzei figured in Sdzuy (Reference Sdzuy1961, pl. 11), including its holotype. In the Bou Tiouit section, this species has its acme around the occurrence of A. ovatopyge, but the cranidia in Sdzuy (Reference Sdzuy1961, pl. 16, figs 3, 7) appear to be distinctly different from those of A. ovatopyge. However, these two cranidia almost certainly can be assigned neither to A. mureroensis (if compared with the holotype cranidium), nor to A. ovatopyge, A. nobilis or A. levisettii from Morocco.
The pygidium from the type series (SMF X 1370a; Fig. 9e, h) is generally taken as typical for the species, whereas the cranidia have not received much attention. However, pygidium SMF X 1370a is sheared at an angle of 24° relative to the length axis and suffered oblique rotational distortion, which is particularly visible on the left-hand side. By contrast, the pleural area on the right-hand side experienced a slight oblique distortion. This makes a confident reconstruction of the specimen’s precise original size difficult. Sdzuy’s (Reference Sdzuy1958, pl. 1, fig. 13) original figure is retouched and does not show the precise morphology. The blackening of the matrix surrounding the specimen is not accurately done, and therefore seems to show small differences when compared with the unretouched margin (compare with Sdzuy Reference Sdzuy1961, pl. 17, fig. 5).
Additional pygidia assigned to A. mureroensis were figured by Sdzuy (Reference Sdzuy1961, pl. 17, figs 1–4). These differ considerably in outline, although they show a pronounced distortion, and it is fairly certain that they do not belong to a single species. All come from ‘Fp. 15’ so that the precise stratigraphic occurrence is unknown.
The type series thus includes a morphological variety of sclerites that show tectonic deformation/distortion. However, morphological differences can be recognized between the specimens that apparently indicate that two different species were united under A. mureroensis by Sdzuy (Reference Sdzuy1958). Alternatively, it cannot be ruled out that all specimens of the type series belong to the same species, but it is impossible to prove this, and morphometric analysis does not permit an answer to this question.
At any rate, it is puzzling why the reconstruction of A. mureroensis in Álvaro et al. (Reference Álvaro, Esteve and Zamora2018, fig. 4) shows distinct differences from the sclerites in the type series in both the cranidium and pygidium. The topotype pygidium selected in Álvaro et al. (Reference Álvaro, Esteve and Zamora2018, fig. 15) that served as the template for their reconstruction differs so much from the paratype refigured herein that it is obvious that not just a single species assignable to A. mureroensis occurs in the Rambla de Valdemiedes 1 section. Although the precise morphology of A. mureroensis is unknown to date, careful bed-to-bed sampling in the type section at Murero may be able to reveal the characters and morphological plasticity of the species so that the species does not need to be restricted to the holotype.
The desire to rely on an easily recognizable and stratigraphically distinct paradoxidid species in the Iberian successions for biostratigraphic correlations appears to have prompted subsequent authors to assign virtually all specimens from this lowermost part of the middle Cambrian in the Iberian stratigraphic concept (e.g. Gozalo et al. Reference Gozalo, Liñán, Dies Álvarez, Gámez Vintaned, Mayoral, Linnemann, Nance, Kraft and Zulauf2007, Reference Gozalo, Liñán, Gámez Vintaned, Dies Álvarez, Chirivella Martorell, Zamora, Esteve, Mayoral, Rábano, Gozalo and García-Bellido2008, Reference Gozalo, Dies Álvarez, Gámez Vintaned, Zhuravlev, Bauluz, Subías, Chirivella Martorell, Mayoral, Gursky, Andres and Liñán2013) to A. mureroensis, although distinct differences can be seen in the relatively few specimens figured. It is also noteworthy that A. mureroensis is mentioned in at least 38 different publications on the Cambrian in Spain, but only figured in about ten publications. In these publications, the specimens are often considerably distorted, although regarded as being typical of the species (e.g. Gozalo et al. Reference Gozalo, Liñán, Dies Álvarez, Gámez Vintaned, Mayoral, Linnemann, Nance, Kraft and Zulauf2007, fig. 1a, b), and the pygidia of this ‘species’ differ in their morphology so distinctly that they have been described as different ‘morphotypes’ explained by ‘sexual dimorphism’ (e.g. Gozalo et al. Reference Gozalo, Liñán and Díes2003). This assumption of a highly plastic morphology has allowed assignment of distinct sclerites to the same species.
It is the conclusion of this report that severe tectonic distortion of the topotype series of A. mureroensis and the likelihood that Iberian reports of this species mean that no reliable taxonomic diagnosis, description and illustration of this purportedly key West Gondwanan trilobite are credible. For this reason, this purported species is regarded as a nomen dubium. This report’s description of A. cf. mureroensis from Morocco is of a taxon known from sclerites that have a resemblance to the deformed material from the Iberian Chains and are possibly identical with the holotype of A. mureroensis.
The difficulties in the correct identification of A. mureroensis contrasts with the willingness to identify this species in almost every succession. Prior to rigorous description and illustration of material from Morocco, A. mureroensis was noted to occur in the sections in the Atlas ranges (e.g. Gozalo et al. Reference Gozalo, Liñán and Álvaro1993; Sdzuy, Reference Sdzuy, Gámez Vintaned and Liñán1995; Sdzuy et al. Reference Sdzuy, Liñán and Gozalo1999). Subsequently, these reports of A. mureroensis occurrences from Morocco were shown to be erroneous and many of the specimens referred to A. nobilis (see Geyer, Reference Geyer1998). In addition, a large number of additional records of A. mureroensis were also erroneous or are based on assumptions without careful morphological analysis. These A. mureroensis identifications include such trilobites as Acadoparadoxides deani Geyer & Vincent, Reference Geyer and Vincent2015 from Turkey (e.g. Dean et al. Reference Dean, Martin, Monod, Bozdoğan, Gül, Özgül and Turgut1993, Dean & Özgül, Reference Dean and Özgül1994) or A. briareus Geyer, Reference Geyer1993 from younger strata of the Jbel Wawrmast Formation in the Tarhoucht area, A. eopinus (Solov’ev, Reference Solov’ev1969), A. anabaraspissimus (Solov’ev, Reference Solov’ev1969), Eccaparadoxides? rozanovi (Egorova in Egorova et al. Reference Egorova, Shabanov, Rozanov, Savitskiy, Chernysheva and Shishkin1976) and ‘E.’ cultus (Egorova in Egorova et al. Reference Egorova, Shabanov, Rozanov, Savitskiy, Chernysheva and Shishkin1976) from the Siberian Platform, and specimens of A. harlani (Green, Reference Green1834) from Avalonian Massachusetts (e.g. Gozalo et al. Reference Gozalo, Dies Álvarez, Gámez Vintaned, Zhuravlev, Bauluz, Subías, Chirivella Martorell, Mayoral, Gursky, Andres and Liñán2013) to A. mureroensis. Moreover, it appears as sound in this context that typical Moroccan Atlas species occurring in the Assemame quarry such as Latoucheia epichara Geyer, Reference Geyer1990a, and Protolenus (Hupeolenus) dimarginatus Geyer, Reference Geyer1990a were mistaken as the Iberian species Hamatolenus (H.) ibericus Sdzuy, Reference Sdzuy1961 and Alueva undulata Sdzuy, Reference Sdzuy1961 in Zamora et al. (Reference Zamora, Álvaro, Clausen, Esteve, Devaere, Clausen and Álvaro2014, fig. 35F and 35L, respectively).
6. Summary
The stratigraphic sequence of the (provisional) Cambrian Stage 4 – Wuliuan boundary beds in the Jbel Ougnate region, eastern Anti-Atlas of Morocco, is interpreted to accurately record the phylogenetic development of trilobite species in this interval, particularly of ellipsocephaline and protolenine species and at least six species of Acadoparadoxides, the older of which were described in detail by Geyer & Vincent (Reference Geyer and Vincent2015). The assumption published by Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) that A. pampalius Geyer & Vincent, Reference Geyer and Vincent2015, A. levisettii Geyer & Vincent, Reference Geyer and Vincent2015, A. cf. mureroensis (Sdzuy, Reference Sdzuy1958), A. ovatopyge Geyer & Vincent, Reference Geyer and Vincent2015 and A. nobilis Geyer, Reference Geyer1998 are all identical with A. mureroensis (Sdzuy, Reference Sdzuy1958) described from the Iberian Chains, northern Spain, is shown herein to be incorrect. The study of Álvaro et al. (Reference Álvaro, Esteve and Zamora2018) is based entirely on a morphometric analysis, ignores stratigraphic occurrences, individual deformation-related compaction and the original relief, and underestimates some of the earlier described differences between these species. Their suggested synonymy is based on the inability to detect differences by morphometrics. The conclusion is inappropriate because differences cannot be detected solely by the applied morphometric techniques, and other features such as distinctive character associations and the stratigraphic association of sclerites are distinctive of different paradoxidine species. Emphasis of the diagnostic characters and differences between earlier named species of Acadoparadoxides indicate that their differences are often subtle but coherent with the stratigraphic differences.
The emphasis in this and earlier reports on Acadoparadoxides species focuses on distinguishing relatively subtle but consistent morphologic differences between the sclerites of similar species (Geyer, Reference Geyer1998; Geyer & Landing, Reference Geyer and Landing2001; Geyer & Vincent, Reference Geyer and Vincent2015). A comparable focus is now emphasized in neontologic work in which consistent differences in minor morphologic features and the presence of unique features correspond to genomic differences that define distinctive taxa (see review in Westrop et al. Reference Westrop, Landing and Dengler2018). This approach does not necessarily lead to ‘oversplitting’ of existing taxa, but does allow the evaluation of palaeospecies and other taxa that are geographically widespread, purportedly morphologically ‘plastic’ and seem to be particularly ‘useful’ for interregional and interhabitat biostratigraphic correlations.
This report also emphasizes and details the currently unresolved problems in the diagnosis, description, formal illustration and, consequently, identification of the purportedly geographically widespread, early paradoxidine species A. mureroensis. This species is regarded as a nomen dubium known from tectonically distorted material from Iberia, which may include the sclerites of more than one species in its topotype series. As a consequence, if a number of Acadoparadoxides species should ultimately prove to be synonymous, A. mureroensis should not be regarded as a senior synonym. Even the identification of A. mureroensis from some of the Iberian sections, such as the Porma and Valdoré sections in the Cantabrian Mountains, should be re-evaluated. Indeed, any well-preserved Acadoparadoxides material from the lower–middle Cambrian boundary interval of West Gondwana should be compared with what we conclude as the better defined species from Morocco.
Author ORCIDs
Gerd Geyer, 0000-0003-3904-0432; Jakub Nowicki, 0000-0002-3936-5547
Acknowledgements
The contribution of GG to this article was supported by the German Research Foundation (DFG) in the framework of research grant GE 549/22-1. We are most grateful to T. Vincent (Aberdeen) for additional field data and his extensive geometric studies on the material as well as his thoughtful considerations which considerably enhanced the article. The authors are furthermore indebted to F. A. Sundberg (Phoenix, AZ) and E. B. Naimark (Moscow) as well as an anonymous reviewer for criticism and considerations on the initial manuscript.



 Open access
Open access