



Introduction
Weed management in organic crop production is a major challenge (Baker and Mohler Reference Baker and Mohler2015; Bàrberi Reference Bàrberi2002). Organic grain crop farmers often rely heavily on mechanical weed control, which varies in effectiveness based on weather and soil moisture conditions (Bond and Grundy Reference Bond and Grundy2001). In many cases, organic farmers augment mechanical weed control practices with cultural practices that reduce weed populations and weed–crop competition (Bastiaans et al. Reference Bastiaans, Paolini and Baumann2008). Despite this integrated approach, competition from weeds is one of the main factors responsible for the yield gap between organic and conventional production (Kravchenko et al. Reference Kravchenko, Snapp and Robertson2017). For example, Cavigelli et al. (Reference Cavigelli, Teasdale and Conklin2008) reported that weed competition was the second most important factor after low nitrogen availability that prevented organic corn (Zea mays L.) yields from matching conventional corn yields in the long-term Farming Systems Project (FSP) in Maryland. In subsequent research, weeds were shown to have a greater effect on corn yield and nitrogen to have a lesser effect under dry conditions (Teasdale and Cavigelli Reference Teasdale and Cavigelli2010). In soybean [Glycine max (L.) Merr.], which forms a symbiotic relationship with bacteria that fix atmospheric nitrogen, the yield gap between organic and conventional is often almost entirely due to weed competition. For example, in the FSP, soybean yields were on average 19% lower in the organic systems compared with conventional systems, a difference that was attributed solely to weeds (Cavigelli et al. Reference Cavigelli, Teasdale and Conklin2008).
Mechanical and cultural weed management practices (e.g., soil tillage, crop rotation) and the approach that farmers use to manage nutrients can also affect weed communities and weed species diversity (Ryan et al. Reference Ryan, Smith, Mirsky, Mortensen and Seidel2010b). For example, Gruber and Claupein (Reference Gruber and Claupein2009) reported that both weed seedbank density and weed density were greater in organic plots managed with chisel-plow tillage compared with moldboard-plow tillage. Reducing tillage can also facilitate a shift in weed communities toward perennial species (Ryan et al. Reference Ryan, Mortensen, Seidel, Smith and Grantham2009a; Thomas et al. Reference Thomas, Derksen, Blackshaw, Acker, Légère, Watson and Turnbull2004), which can be more difficult to control in organic production (Gallandt Reference Gallandt, Chauhan and Mahajan2014). Secondary cultivation and other weed management practices performed after the crop has emerged can affect weed communities. For instance, rotary hoeing and tine harrowing have been shown to be more effective against shallow-rooted weeds such as giant foxtail (Setaria faberi Herrm.) and common chickweed [Stellaria media (L.) Vill.] compared with weed species capable of emerging from greater depths such as velvetleaf (Abutilon theophrasti Medik.) and wild oat (Avena fatua L.) (Gunsolus Reference Gunsolus1990). Flame weeding has been shown to be more effective against broadleaf weeds such as common lambsquarters (Chenopodium album L.), redroot pigweed (Amaranthus retroflexus L.), and A. theophrasti compared with grass weed species such as large crabgrass [Digitaria sanguinalis (L.) Scop.] and S. faberi (Taylor et al. Reference Taylor, Renner and Sprague2012). In addition to mechanical and thermal weed control practices, the form in which nutrients are supplied can affect weed growth (Blackshaw et al. Reference Blackshaw, Brandt, Janzen, Entz, Grant and Derksen2003; Davis and Liebman Reference Davis and Liebman2001; Little et al. Reference Little, Mohler, Ketterings and DiTommaso2015) and weed community composition (Menalled et al. Reference Menalled, Gross and Hammond2001). Thus, understanding the effects of different management practices on weed community structure (i.e., weed abundance and community composition) is important for developing robust organic weed management strategies.
Typically, weeds are more abundant and weed communities are more diverse in organic cropping systems compared with conventional cropping systems, where synthetic herbicides are used (Hole et al. Reference Hole, Perkins, Wilson, Alexander, Grice and Evans2005; Kremen and Miles Reference Kremen and Miles2012; Menalled et al. Reference Menalled, Gross and Hammond2001; Ryan et al. Reference Ryan, Smith, Mirsky, Mortensen and Seidel2010b; Wortman et al. Reference Wortman, Lindquist, Haar and Francis2010). The agroecological consequences of increased weed species diversity are not well understood. On one hand, increased weed species diversity in organic cropping systems may help to promote ecosystem services such as pollination (Bretagnolle and Gaba Reference Bretagnolle and Gaba2015; Holzschuh et al. Reference Holzschuh, Steffan-Dewenter and Tscharntke2008), biological control of crop pests (Norris and Kogan Reference Norris and Kogan2000), and regeneration of soil health (Smith et al. Reference Smith, Ryan, Menalled, Hatfield and Sauer2011; Sturz et al. Reference Sturz, Matheson, Arsenault, Kimpinski and Christie2001). On the other hand, increased weed diversity may pose additional challenges for organic farmers, as different weeds might require different management practices for effective suppression. Previous research has reported both negative (Davis et al. Reference Davis, Renner and Gross2005) and neutral (Pollnac et al. Reference Pollnac, Maxwell and Menalled2009) relationships between weed species density and crop yield. In other research examining crop yield loss, Storkey and Neve (Reference Storkey and Neve2018) reported a strong negative relationship between weed species richness and crop yield loss in winter wheat (Triticum aestivum L.) and hypothesized that more diverse weed communities will be less competitive with crops due to phenotypic differentiation between the weeds and the crop. Hence, there remains a need to better understand how crop yields respond to both the abundance and diversity of the weed community under different forms of weed management.
We conducted a nested experiment within the long-term Cornell Organic Grain Cropping Systems Experiment (OCS) in central New York State to evaluate the impact of different organic grain management practices on weed community structure and soybean performance. We expected that different approaches to organic grain crop management would result in quantifiably distinct weed communities and that these differences in weed community structure would be associated with differences in soybean yield. Specifically, we hypothesized that (1) a cropping system with greater nutrient inputs would have greater weed abundance compared with systems with lower nutrient inputs, (2) perennial weed species would dominate the weed community in a cropping system with reduced soil tillage compared with systems with increased soil tillage and cultivation, and (3) soybean yield would be greatest in a cropping system with additional soil tillage and cultivation compared with systems with fewer weed management operations.
Materials and Methods
Long-Term Experiment
The OCS experiment was initiated in 2005 at the Cornell University Musgrave Research Farm in Aurora, NY, USA (42.73°N, 76.66°W). The soil was a moderately well-drained, calcareous Lima silt loam (fine-loamy, mixed, semiactive, mesic Oxyaquic Hapludalfs) with partial tile drainage and pH near 7.8. The experiment compared four organic cropping systems that varied in nutrient inputs, weed management, and tillage practices. The experimental design was a split-plot randomized complete block design with four replications. Cropping system (described in the following section) was the whole-plot factor and crop rotation entry point was the split-plot factor. Two crop rotation entry points were used so that two crops were represented in each system each year. Split plots measured 9.1 by 30.5 m (12.2 by 36.6 m including borders).
Cropping Systems
The four cropping systems were managed based on distinct guiding principles throughout the duration of the long-term experiment (2005 to 2015), though specific management practices varied slightly over time. The high-fertility (HF) system received multiple nutrient inputs, including legume green manures, composted chicken manure, and commercial organic fertilizers (Table 1). In contrast, fertility in the low-fertility (LF) system was supplied nearly exclusively by legume green manures. Both the HF and LF systems received intermediate levels of weed management through tillage and cultivation (Table 1). The enhanced weed management (EWM) system received supplemental weed management, mainly additional tillage and cultivation. The reduced-tillage (RT) system received less frequent and less intense tillage operations than other systems. Both the EWM and RT systems were fertilized at intermediate levels (Table 1). The entire field site was managed using organic practices and was certified by the Northeast Organic Farming Association of New York. For a detailed description of management practices used from 2005 to 2010, see Caldwell et al. (Reference Caldwell, Mohler, Ketterings and DiTommaso2014).
Table 1. Primary management differences across the four cropping system treatments in the Cornell Organic Grain Cropping Systems Experiment, Aurora, NY, USA, 2011–2015. a

a Abbreviations: C, corn; S, soybean; SP, spelt; B, winter barley; BU, buckwheat; r, annual ryegrass [Lolium perenne (L.) ssp. multiflorum (Lam.) Husnot]; c, red clover (Trifolium pretense L.); op, oat (Avena hybrida Peterm. ex Rchb.)/Austrian winter pea (Pisum sativum L.) mix.
b Crop rotation was C-S-SP/c in all systems before 2011. See Caldwell et al. (Reference Caldwell, Mohler, Ketterings and DiTommaso2014) for additional information about management during first two rotation cycles.
c Application rates per season. All systems received low-analysis starter fertilizer on corn.
d Composted poultry manure (5-5-3 N-P2O5-K2O, Kreher’s Enterprises, Clarence, NY, USA). Rates assume 14% moisture. Enhanced weed management (EWM) did not receive compost before 2011. Reduced tillage received variable compost when necessary to supplement inadequate legume nitrogen.
e Organic fertilizers applied based on phosphorus and potassium soil tests.
f Tillage practices by crop. All systems also used secondary tillage (disk, roller harrow).
g Events per season in corn and soybean.
h Two passes per year were most common except in EWM, where three passes were most common.
Several changes were made to OCS management practices in 2011 to address emerging agronomic challenges. From 2005 to 2010, all four cropping systems had the same 3-yr rotation of corn–soybean–winter spelt/red clover (Triticum spelta L./Trifolium pratense L.). During this time, weed abundance increased in the HF and RT systems due to high nutrient inputs (HF) and inadequate mechanical control (RT) (Caldwell et al. Reference Caldwell, Mohler, Ketterings and DiTommaso2014). Consequently, the crop sequence in the HF and RT was changed in 2011 to a 6-yr rotation of corn–soybean–winter spelt–winter barley/buckwheat (Hordeum vulgare L./Fagopyrum esculentum Moench)–soybean–winter spelt/red clover to vary the timing of tillage in these systems and disrupt weed life cycles. The LF and EWM systems maintained the original crop rotation (Table 1). Also in 2011, an experimental ridge tillage practice in the RT system was replaced by a combination of chisel-plow and deep zone tillage (Table 1) to provide better mechanical weed control. Starting in 2011, we applied compost to spelt in the EWM system and increased spelt seeding rates (to match the EWM rate) in all systems as an additional cultural weed control practice (Table 1).
Nested Experiment
In 2014 and 2015, a nested experiment was conducted in the soybean phase of each crop rotation entry point. Soybean followed buckwheat (HF, RT) or corn (LF, EWM) in both years of the nested experiment. Soybean ‘Dares’ (relative maturity 0.8, food grade, imperfect yellow hilum; Butterworks Farm, Westfield, VT, USA) was inoculated with N-Dure™ (Bradyrhizobium japonicum, INTX Microbials, Kentland, IN, USA) and planted in 76-cm rows at 642,000 seeds ha−1 (2014) or 715,000 seeds ha−1 (2015) using a four-row planter (John Deere, Moline, IL, USA). Soybean seeding rates were higher than conventional rates in order to hasten crop canopy closure and suppress weeds.
Weed Infestation Treatments
Two weed infestation treatments, ambient weed (AW) and weed-free (WF), were implemented in subplots (3.0 by 6.1 m) that were randomly located in each soybean main plot. AW subplots received standard soybean management practices for the systems in which they were located (Table 2). WF subplots were hand weeded every 1 to 2 wk from June 27 to August 7 (2014) and July 2 to August 6 (2015) in addition to receiving standard weed management practices.
Table 2. Dates of management practices in the nested experiment (2014–2015). a
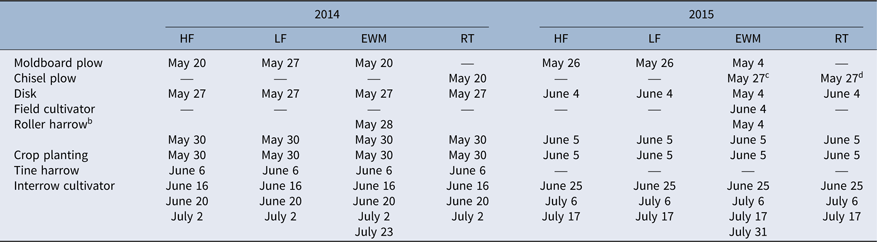
a Systems and years: EWM, enhanced weed management; HF, high fertility; LF, low fertility; RT, reduced tillage. 2014, entry point A; 2015, entry point B.
b With or without roller-harrow tines engaged depending on residue.
c Chisel plow was used in EWM in 2015 to disrupt rhizomatous perennial weeds.
d Chisel plow was used two times in RT in 2015.
Sampling
Biomass and Density
Soybean and weed aboveground biomass in each subplot were sampled in two 0.25-m2 quadrats (76 by 33 cm) on August 18 (2014) and August 14 to 15 (2015). All stems rooted in the quadrat were clipped at soil level, separated by species, counted, oven-dried at 40 C for at least 2 wk, and weighed. Data from the two quadrats in each subplot were averaged before analyses.
Soybean Yield
Soybeans were harvested on October 6 (2014) and October 7 to 8 (2015) from the inner two rows of each subplot using a plot combine that measured grain mass and moisture content (Almaco, Nevada, IA, USA). Soybean grain yields were standardized before analyses (g m−2 at 13% moisture), accounting for the area where soybean plants were removed during previous sampling (i.e., biomass quadrats).
Data Analysis
Weed Abundance and Community Composition
Weed species richness and evenness were calculated in each AW subplot. Richness was determined as the number of weed species present per 0.5 m−2. Evenness describes the relative allocation of weed biomass or weed density among species and was calculated using the following equation:
 $${\rm{Evenness = - }}\sum {\left[ {{{\rm{P}}_{\rm{i}}} \times {\rm{ln}}\left( {{{\rm{P}}_{\rm{i}}}} \right)} \right]/\ln \left( {\rm{S}} \right)} $$
$${\rm{Evenness = - }}\sum {\left[ {{{\rm{P}}_{\rm{i}}} \times {\rm{ln}}\left( {{{\rm{P}}_{\rm{i}}}} \right)} \right]/\ln \left( {\rm{S}} \right)} $$
where P i is the proportion of each weed species’ biomass or density relative to the subplot total, and S is species richness of the subplot. Evenness ranges from near 0 (one species highly dominant) to 1 (all species equally abundant).
Weed density, biomass, richness, and evenness were analyzed using mixed-model ANOVA. Fixed effects were year, system, and their interaction. Two random effects (block and system within block) were included in the model. Residuals were visually checked for homogeneity of variance. Weed density and biomass were ln(x + 1) transformed to correct heteroscedasticity; back-transformed least-squares means are reported. Least-squares means were grouped at significant (P < 0.05) factor levels using the Tukey method. All ANOVAs were performed in R v. 3.2.0 (R Foundation for Statistical Computing, Vienna, Austria; R Development Core Team 2018).
Effects of cropping system on weed community composition under AW conditions were analyzed using multivariate techniques in PC-ORD v. 6.08 (MjM Software, Gleneden Beach, OR, USA). We compared weed communities in the AW treatment between cropping systems using permutation-based multivariate analysis of variance (PERMANOVA) with Bray-Curtis distance measures. Weed species occurring in fewer than two AW subplots were omitted. A two-way factorial design (year by system) was first used to test for overall effects. Statistical significance was estimated using a randomization procedure (5,000 runs). Then, pairwise comparisons at significant factor levels were tested manually using a one-way factorial design. Where necessary due to missing data, we used a repeated stratified random sampling procedure and reported P-values after 500 iterations (Peck Reference Peck2010).
To evaluate weed community composition and identify dominant species (i.e., those making up 95% of the total biomass), individual weed species were ranked within each system and year. Association of individual weed species with particular cropping systems was tested using indicator species analysis (Dufrêne and Legendre Reference Dufrêne and Legendre1997). Indicator values for each species were calculated by multiplying relative abundance by relative frequency within a cropping system. Indicator values range from 0 (not detected) to 100 (exclusive association). Significance of indicator values was estimated using a Monte Carlo procedure (5,000 runs) and considered significant when P < 0.1. Data from each year were analyzed separately.
Soybean Performance
Soybean biomass and yield were analyzed using mixed-model ANOVA. WF and AW treatment subplots were analyzed in a single model. Fixed effects were year, system, treatment, and their interactions. Three random effects (block, system within block, and entry point within system within block) were included in the model. The relationship between soybean biomass and soybean yield was tested using correlation analysis. We also used correlation analysis to test for relationships between (1) weed species richness (measured in the AW treatment subplot) and soybean biomass (measured in AW and WF), (2) weed species richness (measured in the AW treatment subplot) and soybean yield (AW and WF), and (3) weed species richness (measured in the AW treatment subplot) and percent biomass loss and percent yield loss in the AW relative to the WF treatment, calculated as (WF − AW)/WF.
Results and Discussion
Monthly mean temperatures during both soybean-growing periods of the experiment (May to October in 2014 and 2015) were within 4 C of the historical average (Figure 1). Precipitation in 2014 was similar to the historical average. In 2015, heavy spring rainfall (Figure 1) and poor drainage at the field site caused ponding in some plots in early June, which resulted in patchy soybean germination and slow early-season growth in all systems.

Figure 1. Monthly average temperature (lines) and total precipitation (bars) at the Musgrave Research Farm in Aurora, NY, USA (42.73°N, 76.66°W), 2014–2015. Actual conditions (solid, black) are shown beside 30-yr historical averages (dotted, white). Data were accessed through the Northeast Regional Climate Center database (http://climod.nrcc.cornell.edu).
Weed Abundance in AW Conditions
Analysis of weed density data showed a year effect and a system effect, but no year by system interaction (Table 3). Weed density was 56% greater in 2015 compared with 2014. The wet conditions in early June 2015 prevented tine harrowing and delayed the first interrow cultivation until 3 wk after planting (Table 2). Thus, once interrow cultivation was finally possible, weeds were larger than the ideal size for control, and the operation was less effective, a problem that has been discussed in previous literature (Gunsolus Reference Gunsolus1990). Previous research has shown that propane flame weeding can be more effective than rotary hoeing under wet conditions, and thus could be a potential solution to the problems encountered in 2015, when early-season control practices were compromised (Taylor et al. Reference Taylor, Renner and Sprague2012).
Table 3. Results of ANOVA on weed density, biomass, species richness, and species evenness under ambient weed conditions.a

a Systems and years: EWM, enhanced weed management; HF, high fertility; LF, low fertility; RT, reduced tillage. 2014, entry point A; 2015, entry point B.
b For density and biomass, ANOVA was performed on ln(x + 1) transformed values, and back-transformed least-squares means are reported.
c Similar letters within a column indicate no significant difference (P > 0.05).
d Similar letters within a year (uppercase in 2014 and lowercase in 2015) indicate no significant difference between systems.
When pooled over years, mean weed density ranged from 28 to 82 stems m−2 and was lower in the EWM system compared with the LF and RT systems (Table 3). Results indicate that the additional cultivation operations in EWM were effective at reducing weed density. Weed densities in our experiment were greater than weed densities found in organic soybean in Iowa (Delate and Cambardella Reference Delate and Cambardella2004) but lower than weed densities found in organic soybean in Pennsylvania (Ryan et al Reference Ryan, Mortensen, Bastiaans, Teasdale, Mirsky, Curran, Seidel, Wilson and Hepperly2010a).
Average weed biomass ranged from 12 to 69 g m−2 across systems and years (Table 3). Weed biomass in all systems was low compared with that reported in previous years in the OCS (Caldwell et al. Reference Caldwell, Mohler, Ketterings and DiTommaso2014) and in other long-term organic grain experiments (Davis et al. Reference Davis, Renner and Gross2005; Menalled et al. Reference Menalled, Gross and Hammond2001; Ryan et al. Reference Ryan, Smith, Mortensen, Teasdale, Curran, Seidel and Shumway2009b). Contrary to our first hypothesis, no differences in weed biomass between cropping systems were observed (Table 3). In previous years in the OCS, weed biomass varied by cropping system. For example, in 2008, weed biomass was greater in the RT system than the other systems, and in 2009, weed biomass was greater in the HF system than the other systems (Caldwell et al. Reference Caldwell, Mohler, Ketterings and DiTommaso2014). During our experiment in 2014 to 2015, weed density and biomass were not different in the HF system compared with the other systems, despite a history of greater poultry manure compost inputs in the HF system. In contrast, Caldwell et al. (Reference Caldwell, Mohler, Ketterings and DiTommaso2014) reported almost double the weed biomass in the HF compared with the LF system during the first 6 yr of the experiment. In other research, weed abundance has been shown to increase with nutrient additions, especially when nutrients are supplied in excess of crop demand (Blackshaw et al. Reference Blackshaw, Brandt, Janzen, Entz, Grant and Derksen2003; Davis and Liebman Reference Davis and Liebman2001). Lower than expected weed biomass in the HF and RT systems was likely due to the crop rotation changes in 2011 (i.e., replacing corn with winter barley and buckwheat in the crop rotation of those systems).
Weed Community Composition in AW Conditions
Analysis of weed species richness data showed a year by system interaction (Table 3). In both years, weed species richness was greater in RT than in EWM (Table 3). This result suggests that tillage and cultivation are strong weed community assembly filters. Differences between systems appeared to be intensified by greater spring rainfall resulting in less-effective mechanical weed control in 2015. For example, in 2014, average weed species richness in HF and LF was not different than in RT or EWM, whereas in 2015, weed species richness was lower in HF and LF than in RT and was greater in LF than in EWM (Table 3). Our results are congruent with previous research in conventional production showing that tillage has a large effect on weed species richness and that reduced tillage promotes greater weed species richness (Sosnoskie et al. Reference Sosnoskie, Herms and Cardina2006). In contrast to weed species richness, weed species evenness was not affected by cropping system, meaning that weed communities in all cropping systems were similarly homogenous (Table 3).
PERMANOVA on weed density data indicated that weed community structure differed at the year by cropping system interaction level (Table 4). Weed community composition in RT differed from LF and EWM in both years, and LF differed from EWM in 2015. PERMANOVA on weed biomass data indicated that weed communities differed by year and by system, but not at the interaction level (Table 4). The weed community in RT differed from LF and EWM across years, and HF differed from EWM across years. Differences in weed community structure between years might have been due to efficacy of in-season weed management based on weather conditions, legacy effects of crop rotation entry point, or spatial heterogeneity of the field site.
Table 4. Results of PERMANOVA on weed community using weed density and weed biomass under ambient weed conditions.a

a Systems and years: EWM, enhanced weed management; HF, high fertility; LF, low fertility; RT, reduced tillage. 2014, entry point A; 2015, entry point B.
To elucidate weed community differences identified by PERMANOVA, we ranked the weed species present in each cropping system and year based on their abundance. The number of dominant weed species (i.e., those making up 95% total biomass) ranged from two to eight and was lowest in EWM in both years, reflecting lower weed species diversity compared with other systems (Table 5). Two summer annual species, common ragweed (Ambrosia artemisiifolia L.) and S. faberi, were present in seven out of eight cropping system by year combinations, and in some cases were highly abundant. Winter barley and buckwheat were planted in the HF and RT systems to reduce weed problems; however, they also became volunteer weeds. For example, in the HF and RT systems in 2014, buckwheat accounted for more than 40% of total weed biomass, but only 3% of weed density.
Table 5. Rank abundance of dominant weed species (i.e., species accounting for 95% of total weed biomass) in each cropping system under ambient weed conditions, with percentages and absolute values of weed density (D, stems m−2) and biomass (B, g m−2) presented. a

a Systems and years: EWM, enhanced weed management; HF, high fertility; LF, low fertility; RT, reduced tillage. 2014, entry point A; 2015, entry point B.
b Asterisks (*) denote perennial species.
Perennial weeds were abundant in both years; however, their relative abundance and the occurrence of specific species among systems was not consistent across years. Differences in weed species occurrence between years can be partially attributed to soybeans being grown in different areas (i.e., crop rotation entry point A in 2014 and crop rotation entry point B in 2015). In 2014, perennial weeds comprised 33%, 62%, 11%, and 15% of total weed biomass in the HF, LF, EWM, and RT systems, respectively, and in 2015 they made up 3%, 31%, 84%, and 39% of the weed biomass in these systems (Table 5). Perennial sowthistle (Sonchus arvensis L.) was the second most abundant species in the HF system in both years, accounting for 19% and 15% of total weed biomass in 2014 and 2015, respectively. This species was also highly abundant in the LF system in 2014 (56% of total weed biomass). Hedge bindweed [Calystegia sepium (L.) R. Br.] accounted for 6% and 31% of total biomass in the LF system and 11% and 84% of total biomass in the EWM system in 2014 and 2015, respectively. Canada thistle [Cirsium arvense (L.) Scop.] accounted for 15% and 20% of total weed biomass in the RT system in 2014 and 2015, respectively. Five out of eight dominant weed species in the RT system in 2015 were perennial, providing partial support for our second hypothesis that perennial weed species would dominate the weed community in a cropping system with reduced soil tillage. Our results are congruent with Gruber and Claupein (Reference Gruber and Claupein2009), who found C. arvense density, biomass, and seedbank abundance increased with chisel plowing compared with when more intense primary tillage methods were used. However, other perennial species such as Sonchus arvensis and C. sepium were more abundant in the HF, LF, and EWM systems compared with the RT system (Table 5). The relatively high abundance of perennial weed species in the HF, LF, and EWM systems, despite more intensive tillage, was likely due to the tillage and cultivation practices failing to reduce carbohydrate reserves in roots (Sonchus arvensis) and rhizomes (C. sepium). In a meta-analysis of perennial weed management in organic cropping systems, Orloff et al. (Reference Orloff, Mangold, Miller and Menalled2018) report that although mechanical weed control was the most commonly researched method, its efficacy was variable, and it did not outperform other methods of control. Differences in sprouting activity and readiness of root and rhizome buds in relation to tillage and cultivation is another potential reason why different perennial species were dominant in different cropping systems (Brandsæter et al. Reference Brandsæter, Fogelfors, Fykse, Graglia, Jensen, Melander, Salonen and Vanhala2010).
Indicator species analysis showed that 11 weed species were associated with cropping systems at the P < 0.1 level (Table 6). One species was associated with the EWM system, three were associated with the HF system, and three were associated with the LF system, while eight species were associated with the RT system (counting years separately). This indicates that management practices used in the different cropping systems acted as weed community filters. Twice as many species were observed to be significant indicators of a given system in 2015 compared with 2014. As discussed previously, heavy precipitation in June 2015 reduced the efficacy of mechanical weed management practices and likely contributed to the difference in indicator species across years. Two species, C. album and buckwheat, were consistently associated with the RT system in both years. Two other species were inconsistent indicators across years. Specifically, C. sepium was associated with the EWM system in 2014 and the LF system in 2015, and S. faberi was associated with the RT system in 2014 and the LF system in 2015. In addition to C. sepium, other perennial indicator species included dandelion (Taraxacum officinale F. H. Wigg.), which was associated with the HF system in 2014, and C. arvense, which was associated with the RT system in 2015. The large number of species associated with the RT system is consistent with our observation that this system had both high species richness and a distinct weed community relative to the other systems.
Table 6. Results from indicator species analysis using weed biomass and weed density under ambient weed conditions. a b

a Systems and years: EWM, enhanced weed management; HF, high fertility; LF, low fertility; RT, reduced tillage. 2014, entry point A; 2015, entry point B.
b Significance of indicator values (IV) was assessed with a Monte Carlo procedure and species with a significant (P < 0.1) IV are reported.
c Asterisks (*) denote perennial species.
Results from this research are congruent with previous research on the effect of management practices on weed community assembly. Ryan et al. (Reference Ryan, Smith, Mirsky, Mortensen and Seidel2010b) analyzed the soil weed seedbank in a long-term cropping systems experiment that compared a standard conventional management system to two organic management systems that differed in nutrient input (manure vs. legume). Indicator species analysis showed that T. officinale was associated with the manure-based organic cropping system that had higher nutrient inputs, whereas S. faberi was associated with the legume-based organic cropping system that had lower nutrient inputs (Ryan et al. Reference Ryan, Smith, Mirsky, Mortensen and Seidel2010b). Interestingly, C. album and C. arvense, which were both associated with the RT system in this research, were both associated with the conventional system, which used reduced tillage relative to the legume-based and manure-based organic cropping systems (Ryan et al. Reference Ryan, Smith, Mirsky, Mortensen and Seidel2010b).
Soybean Performance
Average soybean population, measured in August after weed management practices occurred, did not differ by cropping system (P > 0.05) and was 473,800 plants ha−1 in 2014 and 517,900 plants ha−1 in 2015, representing 74% and 72% of seeding rates, respectively. We observed a year by system interaction effect on soybean biomass, but no differences based on weed management treatment (Table 7). Soybean biomass was not affected by cropping system in 2014. In 2015, soybean biomass was lower in LF than in RT, whereas soybean biomass in HF and EWM did not differ from LF or RT.
Table 7. Results of ANOVA on soybean biomass and yield under ambient weed and weed-free conditions. a

a Systems and years: EWM, enhanced weed management; HF, high fertility; LF, low fertility; RT, reduced tillage. 2014, entry point A; 2015, entry point B.
b For each effect, similar letters within a column indicate no significant difference (P > 0.05).
c For the year × system interaction effect, similar letters within a year (uppercase in 2014 and lowercase in 2015) indicate no significant difference between systems.
We observed effects of year, system, and weed management treatment on soybean yield, but no interactions between these factors (Table 7). Although the results from the analysis of soybean yield varied slightly from the soybean biomass results, soybean yield and soybean biomass were correlated (r = 0.70, P < 0.001). Compared with historical soybean yields in the OCS trial (2005 to 2010), yields in our experiment were average to above average in 2014, but below average in 2015 (Caldwell et al. Reference Caldwell, Mohler, Ketterings and DiTommaso2014). Lower yields in 2015 appeared to be caused by high precipitation in June 2015 and poor drainage at the field site limiting soybean germination and early growth. Low precipitation in August 2015 may have limited later growth and pod fill (Figure 1). Across both years, EWM had lower soybean yields than HF and RT (P < 0.05), whereas LF yields were intermediate. Thus, we did not find support for our third hypothesis that soybean yield would be greatest in a cropping system with additional soil tillage and cultivation compared with systems with fewer weed management operations. Soybeans in the EWM system lagged behind the HF and RT systems in vegetative growth stage during both growing seasons (Ball Reference Ball2017).
Low soybean yields in the EWM system were unexpected, considering that during the first 6 yr of the OCS experiment (2005 to 2010), soybean yields in EWM were among the highest (Caldwell et al. Reference Caldwell, Mohler, Ketterings and DiTommaso2014). There are several possible explanations for low yields in EWM. Extra interrow cultivation in late July (Table 2) could have damaged soybean roots and depleted soil moisture during late-summer pod fill. Early false-seedbed tillage and other extra tillage in EWM in previous years might have damaged soil structure and depleted soil organic matter. Furthermore, the LF and EWM systems have a history of lower nutrient inputs compared with the HF and RT systems, and soil nutrient depletion may also have limited yield potential.
Soybean biomass did not differ between the AW and WF treatments (Table 7). However, soybean grain yield was 10% greater in the WF treatment compared with the AW treatment (Table 7). The lack of a treatment effect on soybean biomass could be due to the fact that biomass samples were collected earlier than grain yields, which may have been before the onset of weed competition effects. The RT system in our experiment had the highest weed density and species richness, greatest number of indicator weed species, and a weed community that was distinct from both the LF and EWM systems. Despite this, soybean yields in the RT system were among the highest in our experiment, suggesting that high weed abundance and diversity is not necessarily associated with low yield.
Our WF treatment allowed us to examine the soybean production potential of the soil in each cropping system in the absence of weed competition. No relationship was found between weed species richness in AW subplots and soybean biomass in either the AW or WF treatments in either year (Table 8). However, we found a positive relationship between soybean yield measured in WF subplots and weed species richness measured in adjacent AW subplots in both years (Table 8). This suggests that management factors that promoted weed community diversity, such as reduced tillage, may have also increased yield potential of the soil. It is also plausible that diverse weed inputs in years past may have contributed positively to soil quality and yield potential. However, the relationship between weed species richness and soybean yield measured in the AW subplots was not significant, suggesting that weed competition prevented the higher yield potential from being realized. Our results echo those of Ryan et al. (Reference Ryan, Mortensen, Bastiaans, Teasdale, Mirsky, Curran, Seidel, Wilson and Hepperly2010a), who found higher yield potential, but similar actual yields, in long-term organic versus conventional systems. We also tested for relationships between weed species richness in the AW subplots and percent soybean biomass and yield loss using data from both the AW and WF subplots (sensu Storkey and Neve Reference Storkey and Neve2018); however, these relationships were not significant (Table 8).
Table 8. Pearson correlations between weed species richness (species 0.5 m−2) in the ambient weed (AW) treatment, soybean biomass and yield (g m−2) in AW and weed-free (WF) treatments, and percent biomass and yield loss in the AW relative to the WF treatment.

* P < 0.1.
* * P < 0.01.
Results from our research suggest that crop and soil management that led to higher weed species richness in the past might have increased the soybean yield potential relative to management practices that led to lower weed species richness. Greater weed species diversity could increase the diversity of plant residue inputs to the soil and thus increase soil resource pool diversity and overall capacity of the soil to support plant growth (Smith et al. Reference Smith, Mortensen and Ryan2010, Reference Smith, Ryan, Menalled, Hatfield and Sauer2011). Alternatively, past management practices that reduced weed species richness might have also decreased the yield potential of the soil. In other words, the more frequent interrow cultivation in the EWM system that resulted in lower weed populations and weed community diversity might have degraded soils and lowered the capacity of the soil to support soybean growth. It is also possible that shorter-term effects such as root pruning, soil moisture depletion, and soil compaction from late summer cultivation might account for these results.
The purpose of this research was to evaluate weed communities and their effects on soybean performance in a long-term organic cropping systems experiment. In our 2-yr nested experiment, we observed differences in weed density, weed species richness, and weed community composition among cropping systems primarily related to differences in tillage intensity, rather than nutrient inputs, across the four cropping systems that were tested. Overall, soybean yields were 10% greater in the WF subplots compared with the AW subplots. Although hand weeding used in the WF subplots may not be practical in large-scale organic grain crop production, our results suggest that investing in improved practices that reduce weed competition might be profitable given price premiums for organic crops. Counterintuitively, soybean yields were lowest in the EWM system, where additional tillage and cultivation resulted in relatively low weed abundance and diversity. This suggests that intensive mechanical weed control, though effective in reducing weed abundance and diversity, might also reduce soybean yield potential. Future research should aim to increase crop yield in organic production systems by gaining insights into how weed management practices influence not only weed density and weed community structure, but also soil organic matter, soil nutrient reserves, soil moisture, soil hardness, crop rooting depth, and other factors that influence crop yield potential.
Acknowledgments
The authors thank Sandra Wayman for helpful comments on earlier drafts of this article, and Ann Bybee-Finley, Laura Campos, Tania Carvalho, Jason Chari, Andrew Coon, Ellie Crowell, Kiera Crowley, Alex Eacker, Lauren Gabuzzi, Julian Garcia, Lauren Hill, Ashley Jernigan, Jeff Liebert, Chris Pelzer, Kirby Peters, Sandra Wayman, Sarah Wharton, and Laurel Wolfe for help in the field and lab work. This material is based upon work that was supported by the National Institute of Food and Agriculture, U.S. Department of Agriculture Controlling Weedy and Invasive Plants Program award number 2013-67014-21318 and Hatch project 1010566. No conflicts of interest have been declared.









