


INTRODUCTION
Tularaemia is a zoonosis caused by the Gram-negative, pleomorphic, non-motile bacterium Francisella tularensis, capable of growing intracellularly. Because of its high infectivity and low infectious dose, F. tularensis has been classified as a potential bioterrorism agent [Reference Dennis1]. The species F. tularensis includes two clinically relevant subspecies: tularensis and holarctica which differ in virulence and geographical distribution [2]. In Europe, the subspecies most frequently detected is holarctica and various endemic regions have been identified [Reference Tärnvik, Priebe and Grunow3]. The subspecies remain phylogenetically closely related and antigenically almost similar [Reference Champion4]. Tularaemia has been reported in a broad range of animal species with varying susceptibility [2, Reference Olsufiev and Dunayeva5], while arthropods and ticks serve as vectors [2, Reference Ellis6]. Humans often acquire an infection by handling animal skins or carcasses, drinking contaminated water or eating uncooked meat from infected animals [Reference Grunow7]. Lagomorphs (hares and rabbits), various rodents (e.g. muskrats, voles, mice, lemmings, and hamsters) and insectivores (e.g. shrews and moles) are the animals most susceptible and can also serve as F. tularensis reservoirs [2, Reference Olsufiev and Dunayeva5]. Canidae and Felidae are probably resistant to the pathogen. In regions where tularaemia is endemic antibodies against F. tularensis can be detected in sera from wild animals [Reference Mörner8], and frequently outbreaks of the disease occur simultaneously in wild animals and humans [Reference Ellis6, Reference Reintjes9].
In Germany, human infections caused by F. tularensis are rare but distributed throughout the country, with some historical as well as recent ‘hot spots’ [10, 11]. Although tularaemia is a reportable disease in Germany, it can be assumed that many cases have not been recognized due to a mild course of the disease or failure to consider it as a differential diagnoses. The natural occurence of F. tularensis in Germany has not been well studied, thus, the reservoirs and transmission routes of the pathogen are largely unknown. Finally, due to the paucity of evidence-based information the risk for human tularaemia is difficult to predict.
Seroprevalence studies in various animal populations could help to estimate the occurrence of the tularaemia pathogen in nature. Two previous studies in the north of Germany revealed the absence of F. tularensis antibodies in hares [Reference Frölich12] but a seroprevalence of 3·5% in wild boars [Reference Al Dahouk13]. For the present study we used the standard methods, i.e. competitive or indirect enzyme-linked immunosorbent assay (ELISA) and Western blot (WB), for the detection of F. tularensis antibodies showing reactivity with the lipopolysaccharide (LPS) of the bacterium. The LPS of this pathogen is highly specific and represents the main target for species-specific antibodies [Reference Aronova and Pavlovich14]. In previous studies it has been shown that these antibodies did not recognize LPS of potentially cross-reacting bacteria [Reference Grunow15]. The aim of this study was to obtain more information about the prevalence of F. tularensis in the federal state of Brandenburg, a geographical region in Germany that has not been well studied and to investigate the tularaemia seroprevalence in zoo animals because this has not been investigated previously in Germany. The expected data could show the exposition of the studied animal populations to F. tularensis and could be helpful in estimating the potential risk for transmission of tularaemia from animals to humans and between animal populations. Zoo animals are of interest to study because they could have contacts with wild animals including tularaemia-transmitting species. The investigation of carnivores and omnivores could serve as indicators for a broader spectrum of wild animal species highly susceptible and sensitive to the causative agent of tularaemia, thereby simplifying the monitoring of wild life.
MATERIALS AND METHODS
Study sites
Blood from wild animals was collected in the federal state of Brandenburg which is located in the north-east of Germany (Fig. 1a, green area). Moreover, serum samples from zoo animals that had been collected between 1992 and 2007 for a serosurvey in central European zoos [Reference Probst, Speck and Hofer16] were analysed. They originated from 11 zoological gardens in Germany (Fig. 1a, red numbers): 1, Berlin Zoological Park Friedrichsfelde; 2, Berlin Zoo; 3, Dortmund Zoo; 4, Gelsenkirchen Zoo; 5, Hagenbeck Zoological Park (Hamburg); 6, Hanover Zoo; 7, Karlsruhe Zoo; 8, Krefeld Zoo; 9, Leipzig Zoo; 10, Nuremberg Zoological Park; 11, Stuttgart Wilhelma Zoological and Botanical Garden.
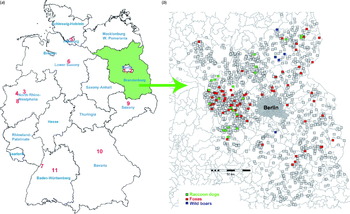
Fig. 1. (a) Sampling sites of captive animals in Germany (sample collection 1, red numbers indicate the sampling sites) and wild animals (serum collection 2, green area). (b) Regional map of Brandenburg depicting sampling locations of wild animals. Coloured boxes indicate individuals that tested positive, grey boxes those that tested negative. Each box represents one individual; green, raccoon dogs; red, foxes; blue, wild boars.
Data and sample collection
A total of 1122 serum samples from 1122 animals, comprising mainly ungulates, were collected in the zoos and zoological parks (serum collection 1). They represented 73 species of Bovidae, 20 species of Cervidae, 14 species of Equidae, nine species of Camelidae, two species of Caviidae and one species each of Giraffidae, Suidae and Rhinocerotidae, respectively. Berlin Zoo provided serum samples taken from a broad range of animal species, comprising Elephantidae, primates, Rhinocerotidae, Ursidae, Felidae, birds, Hippopotamidae and Tapiridae, representing a total of 46 different species. All sera from zoo animals were summarized in two groups – ungulates and other species (Table 1). Blood samples were collected during immobilization for routine clinical treatment (worming, vaccination) or transport. Only clinically healthy animals were tested. The sample set was assembled from archival blood collected by zoo veterinarians between 1992 and 2007. After centrifugation for 10 min at 3000 rpm, serum samples were stored at −20 °C until testing.
Table 1. Results of ELISA and Western blot for F. tularensis antibodies in different serum samples of zoo and wild animals
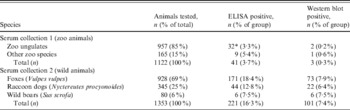
* Competitive ELISA.
A total of 1353 serum samples (serum collection 2) from 928 wild foxes (Vulpes vulpes) (69% of total number), 345 wild raccoon dogs (Nyctereutes procyonoides) (25%) and 80 wild boars (Sus scrofa) (6%) were investigated and data were summarized by these three species groups (Table 1). Foxes and raccoon dogs were shot in Brandenburg between 2006 and 2009 for the monitoring of rabies within the framework of an official state monitoring programme and transported to the regional analytical authority. There, necropsies were conducted by a professional pathologist and blood was taken directly from the heart or from visceral cavities. Prior to shipment to the laboratory, samples were frozen at –80 °C for 1 week to eliminate possible contamination with Echinococcus multilocularis eggs. Sera from wild boars were collected from 2005 to 2008 during classical swine fever monitoring, also within the framework of a state investigative programme. Serum or EDTA blood were taken by the hunters, sent to the laboratory and stored at −20 °C until investigation. A vaccine against tularemia is not available and not applied in Germany for the animal population. Therefore, the presence of anti-F. tularensis LPS antibodies should be the result of a natural infection.
Serological tests
Bacterial preparation, standard LPS ELISA and WB
Preparation and inactivation of bacteria, LPS ELISA and WB were essentially done as described previously with minor modifications [Reference Schmitt17, Reference Jenzora18]. For detection of antibodies against F. tularensis in animal sera (except zoo ungulates, see below), 50 μl/well of protein G–horseradish peroxidase (HRP) conjugate (Millipore, USA) were used in a 1:500 dilution for ELISA and a 1:1000 dilution for WB. All ELISA-positive sera were confirmed by WB.
Competitive ELISA
The competitive ELISA was based on methods described previously and was used for the investigation of the zoo ungulates because of the unknown protein G binding capacity of the broad range of animal sera [Reference Greiser-Wilke, Soine and Moennig19]. Briefly, flat-bottomed 96-well polystyrene plates (PolySorp, NUNC, Germany) were coated by passive absorption with 50 μl/well LPS of F. tularensis (Micromun, Germany) at a concentration of 0·5 μg/ml in carbonate/bicarbonate buffer (pH 9·0) for 1 h at 37 °C. Wells were washed three times with phosphate-buffered saline complemented with 0·05% Tween 20 (PBS-T), and 50 μl serum diluted 1:10 were added in duplicate wells and incubated for 45 min at 37 °C. Fifty μl of a highly specific monoclonal F. tularensis anti-LPS antibody, not cross-reacting with other bacteria [Reference Greiser-Wilke, Soine and Moennig19, Reference Hofstetter20], conjugated to HRP (Seramun, Germany), were directly added without washing in an end-point concentration of 1 μg/ml per well and incubated at 37 °C for 30 min. After incubation wells were washed five times with 300 μl PBS-T. The wells were filled with 200 μl o-phenylendiamine (OPD; Sigma Aldrich, Germany) as substrate; reaction was stopped after 10 min by adding 50 μl of 2·5 m sulphuric acid. Optical density (OD) values were then recorded at 492 nm with a Sunrise Plate Reader (Tecan Instruments, Germany) interfaced with a computer. Results were expressed as percentage inhibition and were derived from the mean OD values for each sample by the following formula:
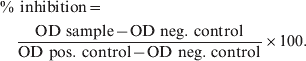 $$\eqalign{\tab\percnt {\rm \ inhibition}\equals\cr\tab\quad {{{\rm OD\ sample} \minus {\rm OD\ neg.}{\rm\ control}} \over {{\rm OD\ pos}{\rm.\ control} \minus {\rm OD\ neg.}{\rm\ control}}} \times 100.{\rm \ }$$
$$\eqalign{\tab\percnt {\rm \ inhibition}\equals\cr\tab\quad {{{\rm OD\ sample} \minus {\rm OD\ neg.}{\rm\ control}} \over {{\rm OD\ pos}{\rm.\ control} \minus {\rm OD\ neg.}{\rm\ control}}} \times 100.{\rm \ }$$Samples were considered as positive if they inhibited more than 45% [mean +3 standard deviation (s.d.)]. Only samples showing inhibition in competitive ELISA greater than 45% were confirmed by WB analyses. Samples with clear positive WB signal (LPS ladder) were stated as positive for F. tularensis antibodies.
ELISA cut-off levels
ELISA results below the mean blank from at least three independent experiments +1 s.d. calculated from the negative control sera were estimated as ‘negative’. Results above the mean blank +3 s.d. were assumed to be ‘positive’, whereas all results between these two values were defined as ‘borderline’. False-positive results in ELISA were avoided by confirming borderline and positive results in WB showing the presence of antibodies against the specific LPS ladder. The seroprevalence was calculated for individual animal groups by percentage of WB positively confirmed sera in relation to all sera of the serum collections of zoo or wild animals, respectively.
RESULTS
We tested a total of 1122 serum samples from captive animals of German zoos (collection 1, Fig. 1a, red numbers) and 1353 serum samples from wild animals from the area of the federal state of Brandenburg (serum collection 2, Fig. 1a, green area). The results are presented in Table 1. In collection 1, a total of 41 (3·7%) samples tested seropositive by competitive and/or indirect ELISA and three (0·3%) of these sera were confirmed positive by WB, including one hippopotamus (Hippopotamus amphibius), one wisent (Bison bonasus) and one African wild ass (Equus africanus).
In collection 2 we detected 101 (7·4%) animals showing positive signals for the LPS of F. tularensis confirmed by WB (Table 1). The localization of the sampling sites and the serological results for all animals from collection 2 are shown in Figure 1b. Figure 2a shows a confirmatory WB result of ELISA-positive raccoon dogs and foxes. The typical LPS ladder is only present in positive samples, whereas negative samples show no signal.

Fig. 2. Western blot for confirmation of ELISA samples tested positive. (a) Examples of positive samples of raccoon dogs and foxes from serum collection 2 showing the typical lipopolysaccharide (LPS) ladder. (b) Western blot with F. tularensis LPS extract showing sera of the hippopotamus that tested positive from serum collection 1 (zoo animals), depicting the seroconversion of the animal. Note the typical LPS ladder only in positive samples from 2004 and 2006. (+), Positive control; (–) negative control; raccoon dogs and foxes = serum collection 2; sera from hippopotamus (serum collection 1); 1, from 2001; 2, from 2003; 3, from 2004; 4, from 2006.
As regards the seropositive hippopotamus, we were able to analyse consecutive serum samples throughout the years 2001, 2003, 2004 and 2006. In 2001 and 2003 they were negative; however, in 2004 and 2006 they were positive indicating a seroconversion during this period (Fig. 2b).
DISCUSSION
Historically, tularaemia in humans was spread throughout Germany with a relatively low incidence reported [10]. Some data indicating higher prevalence could be observed in the north-east and south-west of Germany. Over the last 10 years, from 2001 until November 2011, 119 human cases of tularaemia with 1–31 cases per year have been reported, most of them from the western parts of Germany [11]. During the last years, a tendency for an increase in the number of reported human cases has become apparent, but the real prevalence of tularaemia in Germany remains unknown. A cross-sectional serological study in Germany revealed that these numbers might be underestimated (seroprevalence 0·2%). A seroprevalence study investigating hunters (seroprevalence 1·7%) showed an elevated risk of exposure to the tularaemia pathogen which was also underlined by a tularaemia outbreak in a group of hunters in 2006 [Reference Schmitt17, Reference Jenzora18, Reference Hofstetter20]. In addition, outbreaks or single cases in wild animals, e.g. hares, and semi-free-living animals (marmosets and cynomolgus monkeys) have been described in Germany more recently [Reference Müller21–Reference Mätz-Rensing23].
Zoo animals could be at risk for F. tularensis infections because an oversupply of food can lead to a dense rodent population which could carry and transmit the pathogen. Usually, rodent control programmes reduce this risk, but tularaemia has been described in zoo animals previously [Reference Hoelzle24–Reference Preiksaitis29]. Therefore, it was one aim of our study to conduct the seroprevalence study for tularaemia in different German zoos thereby obtaining further hints for possible sources of infection. Overall, we did not find any striking data on tularaemia seroprevalence using spot samples from zoo animals throughout Germany. The reasons for these negative results might be that we did not target the appropriate animal population (for this study we could only use pre-existing sera) or that zoos do not represent areas with an elevated risk of acquiring tularaemia. Interestingly, the single zoo animals which tested positive comprised a hippopotamus (H. amphibius), a wisent (B. bonasus) and an African wild ass (E. africanus). For ungulates, susceptibility to F. tularensis has been described before [Reference Gordon, McLaughlin and Nutiuthai30, Reference Hoberg31], but these are the first cases reported in a hippopotamus or an equid. Additionally, we were able to show for the hippopotamus that the infection occurred on the premises of Berlin Zoo between the years 2003 and 2004, since the animal was tested negative in 2001 and 2003 and the indirect ELISA and WB for LPS antibodies of F. tularensis were both positive in 2004 and 2006. It can only be speculated on the source and mode of transmission of the tularemia agent in this case. Animals and humans could be infected by contaminated food or water. The source of F. tularensis could be carcasses or excrement of infected rodents which cannot be excluded in this case. This clearly shows the need for continuous rodent control programmes and also the investigation of suspicious deaths of rodents in confined environments.
The study area for wild animals encompassed the federal state of Brandenburg, surrounding the German capital Berlin and located in the north-east of Germany. Sixteen human cases of tularaemia were reported from this region between the years 2001 and 2011 [11] but almost nothing is known about the natural occurrence of F. tularensis in this area. In Mecklenburg-Western Pomerania, a northern neighbouring federal state, retrospectively a relatively high seroprevalence was found in hares (10·7%) and a lower one in wild boars (0·09%) in sera from the years 1976–1989 [Reference Dedek32, Reference Dedek33] as well as more recently in boars (3·1% in 1995/1996) [Reference Al Dahouk13]. To the south-west of Brandenburg in Thuringia, F. tularensis has recently been isolated from a hare and in the western neighbouring state, Saxony-Anhalt, we reported a tick-transmitted human tularaemia [Reference Müller21, Reference Lübbert34]. Therefore, the second aim of our study was to learn more about the activity of the tularaemia pathogen in Berlin and Brandenburg by a serological study of wild animals, also considering the above-mentioned positive results from a hippopotamus in Berlin Zoo. There are no ongoing rodent control programmes which could contribute to monitoring zoonoses in the natural environment. However, a state control programme was conducted from 2006 to 2009 to monitor rabies in wild animals in the region, including predators and omnivores. We took advantage of this programme to investigate specimens from predators and omnivores for the presence of antibodies against F. tularensis. We assumed that these animals might have a higher risk of becoming infected when consuming rodents. Rodents are highly susceptible to F. tularensis and often the source of infection in other mammals. In addition, wild animals are regularly infested with ticks, a known vector for F. tularensis [Reference Gyuranecz35–Reference Hopla38]. Thus, 1353 serum samples of wild omnivores and predators from Brandenburg were investigated. Among foxes, raccoon dogs and wild boars we were able to show a seroprevalence of F. tularensis of 7·5% on average, which indicates that tularaemia might be endemic in wild animals in Brandenburg. There appears to be a tendency that more positive samples could be found in the West of Berlin than in other regions of Brandenburg. This may coincide with two human cases reported from this region in 2011. In the same year, 17 human cases were reported from throughout Germany [11]. In addition, this is also consistent with a very recent observation in 2012 that F. tularensis could be confirmed as the cause of death of a beaver from this region [Landeslabor Berlin-Brandenburg, Frankfurt (Oder), Germany,unpublished data].
Our results also revealed that seroprevalence in these animals might serve as an indicator for enzootic tularaemia activity. These animals clearly develop a strong antibody titre against F. tularensis LPS after exposure to this pathogen. Clinical signs of tularaemia have not been observed in any of the examined animals. This is not surprising as an exposure to the pathogen need not be necessarily linked to a persisting infection or clinical disease. This is particularly true for the present study because we investigated animal species with a low sensitivity of contracting tularaemia [2, Reference Olsufiev and Dunayeva5, Reference Magnarelli, Levy and Koski39]. In previous studies foxes were found to be seropositive for tularaemia with a high proportion, and carnivores together with wild boars are recognized as valuable sentinels in the detection of pathogens in wildlife populations and could serve as biological indicators [Reference McCue and O'Farrell40, Reference Hofer41]. All three sentinel groups studied here have the potential to be used as indicators for the presence of the tularemia pathogen; however, we preferred foxes because the relative high number of sera was easy to obtain from within the framework of other investigations.
In addition to serological screening, organs like lung and liver from foxes and raccoon dogs were investigated by PCR and microbiological culturing in a parallel study, and F. tularensis spp. holarctica was isolated from the liver of one fox [Reference Nattermann42]. This isolation underlined the results of the serological screening and documented the presence of infectious F. tularensis in the study region.
In summary, it can be concluded that our approach allowed detection of the presence of the tularaemia pathogen in a region of Germany in which the occurrence of F. tularensis had not been suspected previously. This could contribute to explaining the incidence of single human cases in this area. However, the actual epidemiological situation and the determination of risk factors for human infections require further investigation. Additional isolates of F. tularensis would underline the presence of the infectious pathogen; further attempts are in progress. Long-term surveillance of wildlife could help to identify areas with a higher risk of infection and also identify the natural reservoirs of F. tularensis in Germany. Although an invasion of wild animals into zoos can not be excluded, the transmission of tularaemia seems to be a rare event. The investigation of more various and highly susceptible animal species could provide further information on the exposure of zoo animals to the tularaemia pathogen.
ACKNOWLEDGEMENTS
The authors thank H. Ranisch for excellent technical assistance. This work was partly supported by a grant from the Federal Ministry of Education and Research (project no. 13N9601).
DECLARATION OF INTEREST
None.



