


Introduction
Tobacco (Nicotiana tabacum L.) remains an economically important crop in the state of Kentucky, with almost US$400 million in sales in 2017 (NASS 2017). Weeds are a significant challenge for tobacco producers. Weed management in tobacco is primarily accomplished with a combination of PRE soil-residual herbicides, cultivation, and hand removal (Pearce et al. Reference Price, van Santen and Sarunaite2017). There are relatively few POST herbicides for tobacco, and they provide limited control of broadleaf weeds. Tobacco is grown on 1-m row spacing, in part to accommodate cultivation equipment and also to maximize the intact leaf yield of large plants. However, this wide row spacing lengthens the time until canopy closure, thereby reducing the effectiveness of crop–weed competition for weed control. The use of strip tillage or no-till, combined with cover cropping that leaves a mulch layer between the crop rows, has proven to be promising for reducing weed emergence and suppressing weed growth in high-value vegetable crops (Brainard et al. Reference Brainard, Peachey, Haramoto, Luna and Rangarajan2013; Haramoto and Brainard Reference Haramoto and Brainard2017; Luna and Staben Reference Luna and Staben2002) and likely can be applied to tobacco production as well. However, growers may face trade-offs in terms of managing the cover crop optimally for weed suppression or providing optimum conditions for planting and successful tobacco establishment.
Winter cover crops, typically wheat (Triticum aestivum L.), are common in Kentucky tobacco production, which has traditionally relied on extensive tillage and high rates of nitrogen fertilizer (Pearce et al. Reference Pearce, Bailey and Walker2017). These cover crops are used primarily to scavenge excess nutrients (Kaspar et al. Reference Kaspar, Jaynes, Parkin and Moorman2007) and prevent soil erosion (Blanco-Canqui et al. Reference Blanco-Canqui, Shaver, Lindquist, Shapiro, Elmore, Francis and Hergert2015). The main goal of the cover crop—whether suppressing weeds, scavenging nutrients, serving as a forage source, or reducing soil erosion while minimizing planting issues—determines the species used and typically informs cover crop management decisions like termination time and method. Multispecies mixtures with small grains, legumes, and brassicas are often promoted over cover crop monocultures by agencies like the Natural Resource Conservation Service (NRCS n.d.), necessitating the need for research into how management by cover crop mixture composition combinations influence weed management and crop yield in production systems.
The impact of cover crops on weed management within the tobacco crop will likely depend on species selection and termination time and method. Mulches formed by surface cover crop residues may suppress weeds through reducing the amount of light that reaches the soil surface (Teasdale and Mohler Reference Teasdale and Mohler1993), harboring seedling diseases (Conklin et al. Reference Conklin, Erich, Liebman, Lambert, Gallandt and Halteman2002), and providing a physical barrier to seedling emergence (Teasdale et al. Reference Teasdale, Rice, Cai and Mangum2012). Nitrogen immobilization by small grain residues may suppress growth of emerged weeds, leading to smaller weeds in these systems (Wells et al. Reference Wells, Reberg-Horton, Smith and Grossman2013). Weed suppression by cover crop mixtures (i.e., those including a small grain and legume) has been shown to be more influenced by biomass production than by the mixture composition (Finney et al. Reference Finney, White and Kaye2016), though others have noted that weed suppression by legume cover crops does not last as long as that provided by small grain residues alone (Cornelius and Bradley Reference Cornelius and Bradley2017; Norsworthy et al. Reference Norsworthy, McClelland, Griffith, Bangarwa and Still2010; Palhano et al. Reference Palhano, Norsworthy and Barber2018). The nitrogen contribution of cover crop mixtures may be less important to regulating weed emergence and growth in systems like tobacco that receive high rates of nitrogen fertilizer that is typically broadcast over the entire field width, even in reduced-tillage systems like strip tillage or no-till (Pearce et al. Reference Pearce, Bailey and Walker2017). Because more biomass is typically linked to better weed suppression (Webster et al. Reference Webster, Scully, Grey and Culpepper2013), cover crops that are terminated later in the season likely result in more residue to suppress weeds. However, terminating earlier in the season results in improved planting conditions and higher yields (Pearce et al. Reference Pearce, Ditsch, Zeleznik and Turner2002). Thus, managing cover crops to optimize weed management may be at odds with what is best for tobacco production.
On the one hand, cover crop mulches may suppress weed emergence through physical, chemical, and biological means. On the other, dense cover crop residues may also interfere with the efficacy of soil-residual herbicides—a risky strategy in a tobacco cropping system that relies heavily on residual herbicide use. If rainfall occurs soon after application, soil herbicides may be washed from the cover crop residues into the soil, with the net effect dependent on the quality and quantity of cover crop residue present and the timing and amount of rainfall received (Ghadiri et al. Reference Ghadiri, Shea and Wicks1984; Reddy et al. Reference Reddy, Locke, Wagner, Zablotowicz, Gaston and Smeda1995). However, dry periods after application can lead to prolonged binding, with subsequent release dependent on the type of cover crop residue and compound applied (Anderson Reference Anderson1988; Ayeni et al. Reference Ayeni, Majek and Hammerstedt1998; Carbonari et al. Reference Carbonari, GLGC, MLB, JRM and Velini2016). In Kentucky, sulfentrazone, typically in combination with carfentrazone-ethyl, is a commonly used soil-residual herbicide in tobacco production. When sulfentrazone was applied to sugarcane (Saccharum spp.) residue, 64% to 77% of the herbicide was released to the soil with 20,000 kg residue ha−1 and 5,000 kg residue ha−1, respectively, if 20 mm of simulated rain occurred within 24 h of application (Carbonari et al. Reference Carbonari, GLGC, MLB, JRM and Velini2016). To our knowledge, lower amounts of surface plant residues have not been studied with sulfentrazone. Between 43% and 52% of acetanilide herbicides reached the soil surface when applied over 1,120 kg ha−1 of wheat residue, and ultimately, the amount reaching the soil surface after irrigation depended on the amount of residue and the compound (Banks and Robinson Reference Banks and Robinson1986). For metolachlor, only 30% reached the soil surface under 2,240 kg ha−1 of wheat residue after 12.5 mm of irrigation, whereas more than 50% of acetochlor and alachlor were released (Banks and Robinson Reference Banks and Robinson1986).
In addition to cover crop residue amount, residue quality (i.e., legumes vs. small grain) can also impact the interception of soil-residual herbicides. Sorption of chlorimuron-ethyl (measured by Kf, or the Freundlich coefficient—the distribution of the compound between the sorbed and solution phase) was almost 2.5 times greater on hairy vetch (Vicia villosa Roth.) residue compared with cereal rye (Secale cereale L.) residue, and less of the compound was found in soil under hairy vetch compared with cereal rye (Reddy et al. Reference Reddy, Locke, Wagner, Zablotowicz, Gaston and Smeda1995). Thus, weed management in high cover crop residue systems is complicated by the types of herbicides used and the ability of the producer to time these applications around weather forecasts.
Removing aboveground residue, as in a forage system, may help alleviate concerns about residual herbicide interception, in addition to alleviating difficulty transplanting into residue that can result in reduced tillage systems; on the other hand, no cover crop residue remains to suppress weed emergence once the soil-residual herbicides have degraded and are no longer effective. Kentucky has many cow/calf operations that could potentially use cover crops as an additional forage source in the spring, providing an additional economic benefit and incentive for growers to adopt cover crops (Schomberg et al. Reference Schomberg, Fisher, Reeves, Endale, Raper, Jayaratne, Gamble and Jenkins2014; Sulc and Franzluebbers Reference Sulc and Franzluebbers2014). Using cover crops for forage also influences species selection, or the decision to plant cover crop mixtures, and the amount of residue remaining in the system. Thus, this management practice represents another set of trade-offs that should be examined.
The objectives of this study were to examine the impact of cover crop composition and termination method on weed control and yield in no-till tobacco. Four site-years of on-farm trials specifically compared: (1) cover crop composition (mixture containing cereal rye and forbs compared with a wheat monoculture); (2) termination treatment imposed on the cover crops—early chemical termination (“early terminated”), late chemical termination (“late terminated”), and a biomass removal to simulate forage production (“removal”); and (3) the presence or absence of soil-residual herbicides. We hypothesized that we would see few impacts of including legumes in a cover crop mixture relative to a small grain cover crop monoculture on weed density and biomass and on yield, as tobacco receives high amounts of inorganic nitrogen fertilizer. Our second hypothesis was that, in the absence of the soil-residual herbicide, the late-terminated treatment cover crops, regardless of species, would be the most effective in suppressing weed emergence due to larger amounts of residue present; we also hypothesized that we would potentially observe reduced tobacco yields in the late-termination treatment if poor soil conditions persisted after transplanting. Finally, we hypothesized that the treatments that left the most cover crop residue on the soil surface would have the greatest potential to intercept and bind the soil-residual herbicide, especially if dry conditions occurred after application. Specifically, if an activating rainfall did not occur soon after application, we predicted that the late-terminated treatment would have greater herbicide interception than the early-terminated treatment, and that both would intercept more than the removal treatment. This would lead to reduced efficacy of the residual herbicide, particularly early in the season.
Materials and Methods
Plot Establishment
This experiment was conducted on three commercial tobacco farms in three counties in Kentucky during 2016 and 2017. Four site-years were evaluated—Garrard County in 2016 (G16) and 2017 (G17; two different fields on the same farm were evaluated), Franklin County in 2016 (F16), and Scott County in 2017 (S17). Table 1 provides details for each site including soil type, GPS coordinates of the farms, and previous crop grown in each field used. At each site-year, a large strip (at least 10 m-wide by 100-m long) of a cover crop mixture was established with a no-till drill in the fall before each experiment. With one exception (F16), a monoculture wheat cover crop was also sown. Composition of the mixture used and seeding rates at each site-year are provided in Table 1. Mixtures included cereal rye, hairy vetch, and crimson clover (Trifolium incarnatum L.) at all site-years; G16 also included canola (Brassica napus L.). The mixture composition at the sites, and the decision not to include the wheat monoculture at F16, was driven by the growers’ interest in the experimental treatment factors.
Table 1 GPS coordinates, soil type, and previous crop in all fields used in the 4 site-years of this experiment.

a Seeding rates of the individual components of the cover crop mixture and wheat monoculture.
b Fine-loamy, mixed, semiactive, mesic Typic Hapludult.
c Canola, did not winterkill.
d Oilseed radish (Raphanus sativus L.), winterkilled.
e Fine-silty, mixed, active, mesic Oxyaquic Fragiudalf.
f Fine, mixed, active, mesic Typic Hapludalf.
g Soybean [Glycine max (L.) Merr.].
h The wheat monoculture cover crop was not sown at F16.
i Fine-silty, mixed, active, mesic Ultic Hapludalf.
j Fine-silty, mixed, active, mesic Fluventic Hapludoll.
k No brassica cover crop was included in the mix examined at S17.
A strip block design was used to examine three termination method treatments: early kill, chemically terminated with all above- and belowground biomass left in situ (“early terminated”); late kill, chemically terminated with all above- and belowground biomass left in situ (“late terminated”); and late kill, with all aboveground biomass cut and removed to simulate a forage cutting (“removal”). The large cover crop strip (mixture or monoculture wheat) was divided into four blocks for replication based on field position and variability. Within each of these blocks, each termination treatment (i.e., main plot treatment) was replicated once. Main plots were 6.4-m wide by 9.1-m long.
The cover crops in the early-termination plots were terminated with paraquat (Gramoxone SL 2.0®, 1.12 kg ai ha−1, Syngenta Crop Protection, Greensboro, NC) plus 1% v-v crop oil concentrate (COC) in mid-April (approximately 6 wk before tobacco planting at most site-years; Table 2); the same rate of paraquat plus COC was applied to the late-termination plots in mid-May (approximately 1 mo before tobacco planting at most site-years; Table 2). All applications were made with an ATV-mounted sprayer using a spray volume of 234 L ha−1. In the removal plots, forage was cut with a sickle bar mower, and all aboveground biomass was raked and removed from the plot. All plots also received an additional application of paraquat (0.66 kgha−1) approximately 3 wk before tobacco planting (Table 2); sulfentrazone (354 g ai ha−1) plus carfentrazone-ethyl (39 g ai ha−1; Spartan Charge©, FMC Corporation, Philadelphia, PA) was also applied at this time for residual weed control. While this last application of paraquat was made to the entire main plot, a subplot (1.5-m long by whole plot width) was left unsprayed with the residual herbicides in each main plot to assess the impacts of the cover crop residues alone on suppressing weeds. Hereafter, subplots without the residual herbicides will be referred to as “unsprayed,” and subplots with the sulfentrazone and carfentrazone-ethyl application are referred to as “sprayed.”
Table 2 Timing of relevant field operations at the 4 site-years.
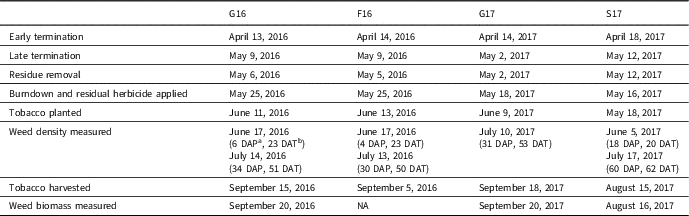
a DAP, days after planting tobacco.
b DAT, days after residual herbicide treatment.
All fields were fertilized with phosphorus and potassium according to soil test results; 280 kg N ha−1 were also broadcast applied as urea (treated with a urease inhibitor) across the entire field in each site-year before planting. At all sites, except S17, tobacco was planted in early to mid-June; S17 was planted on May 18, 2017 (Table 2). A modified no-till transplanter was used at all sites. This unit has row cleaners followed by a subsoiling shank with coulters and cutting disks followed by the planting units. Imidacloprid (0.17 kg ai ha−1) was applied in the transplant water for management of aphids (Myzus spp.) and tobacco flea beetle (Epitrix hirtipennis F.E. Melsheimer, 1847).
Data Collection and Analysis
Cover Crop Sampling
Before all termination treatments, cover crop biomass was sampled from two 0.25-m−2 quadrats in each subplot. Samples were separated into component cover crop species and weeds, then dried at 60 C until a consistent weight was achieved, and weighed.
Weed Density and Biomass Sampling
Weed density was measured in mid-June at 3 site-years (G16, F16, and S17). Heavy precipitation events at the G17 site during this period precluded fieldwork, including counting weeds. In 2016, this count occurred 23 d after the residual herbicide was applied and within 1 wk of planting; at S17, this count was 20 d after the herbicide was applied and 18 d after planting (Table 2). Weed density was assessed in mid-July at all sites—approximately 30 to 34 d after planting at G16, F16, and G17, and 60 d after planting at S17. Weed biomass at harvest was also collected at all site-years except F16.
Weeds were counted in sprayed and unsprayed subplots in all main plots. Due to low weed density, the early-season density evaluation at each site-year was typically performed over a larger area (i.e., 2 to 6 m−2, depending on the density at each site-year). For the midseason density evaluation, two 0.25-m−2 quadrats were counted in each subplot. Quadrats were placed within the second or third tobacco row to avoid the edge rows; quadrats were also placed within the center of the subplots. A tobacco stalk was placed in the corner of the quadrat, and all quadrats were aligned in the same manner so a consistent area of the tobacco row and area between the rows was sampled. All weeds within the quadrat were identified to species and counted. Weed biomass at harvest was measured by clipping all aboveground biomass in two 0.25-m−2 quadrats in all subplots. Samples were dried at 60 C until a consistent weight was achieved, then weighed.
Tobacco was harvested from the center two rows of the sprayed subplots within each main plot following the conventional practice of stalk cutting and spearing the plants onto wooden sticks for curing. A maximum of 30 stalks were harvested from the center of each plot (approximately 16 m2); the number of harvested stalks was recorded if fewer than 30 (i.e., there was reduced stand due to losses at transplanting). All sticks were tagged with plot number and transported to a curing barn where they were hung for 10 to 12 wk. After curing, the leaves were removed from the stalks by hand and weighed to estimate yield of cured leaf.
Data Analysis
Before analysis, all cover crop biomass and weed density and biomass data were averaged over subsamples within an experimental unit. All data were analyzed with SAS software using PROC MIXED (v. 9.4, SAS Institute, Cary, NC). Data were either log or square-root transformed as necessary to meet normality assumptions. Grouping was used to account for heterogeneous variances if necessary, and Akaike information criterion and Bayesian information criterion values were compared to select the best-fit model. Before data analysis, we selected α=0.1 as our significance level due to the variable nature of our sites and weed density in general, in addition to our desire to capture potential treatment effects that may warrant a more detailed examination in future studies. A combination of single degree of freedom contrasts and effects slicing was used to separate significant interactions and determine differences within significant main effects.
Cover Crop Biomass
To examine potential differences in total cover crop biomass production between the mixture and wheat monoculture at site-years with both plantings (all except F16), data were subjected to a two-way ANOVA with site-year by composition as the main plot factor and termination treatment as a fixed subplot factor. Block (replicate) was treated as a random factor. Total biomass production by the cover crop mixture and the proportion of total biomass comprised of legumes (hairy vetch and/or crimson clover) were analyzed using a two-way ANOVA and biomass data from all 4 site-years. Site-year was the main plot factor, with termination treatment as a fixed subplot factor and block as a random factor. The effect of termination treatment on canola biomass and proportion of total biomass composed of canola was also conducted using a one-way ANOVA for the G16 site-year.
Weed Density and Biomass
Due to variability in field management, planting dates, and weather, each site-year was analyzed separately for the weed density and biomass data. The cover crop composition (i.e., mixture or wheat monoculture) was treated as location, with blocks nested within location. Cover crop termination treatments were treated as fixed main plot factors, with the herbicide treatment as a fixed subplot factor. Replications of the cover crop treatments within the species locations were treated as random factors; the herbicide subplot factor was only replicated once within each main plot. Because the sampling size varied depending on sampling date, all weed density and biomass data were converted to number per square meter for presentation; untransformed data are also presented in all cases.
Tobacco Plant Density and Yield
The tobacco crop was assessed by analyzing two variables—number of plants in the harvest area and total marketable yield. Both were subjected to a two-way ANOVA, with separate analyses for each site-year. Cover crop composition was treated as fixed location, with the termination treatments as fixed treatments within location and replicate as a random factor.
Results and Discussion
Cover Crop Biomass and Composition
Cover Crop Biomass—Composition Effects
Results of an initial two-factor ANOVA of the effects of site-year and composition on cover crop biomass (analysis results not shown) showed that cover crop biomass was generally greater in the mixtures than in the monoculture (Table 3; site-year by species interaction P=0.003). At the S17 and G17 site-years, the mixture produced 23% to 29% more biomass than the wheat monoculture cover crop (Table 3; P=0.0021 and P=0.013 for effects slicing, respectively, for the 2 site-years). Biomass was similar between the mixture and the wheat at G16 (P=0.11). The cover crop mixture contained cereal rye as the small grain component, making direct comparisons to the monoculture wheat cover crop difficult. Cereal rye does produce more biomass than wheat in Kentucky in the spring (ERH, unpublished data). However, greater biomass production by the mixture at these 2 site-years could illustrate the “overyielding” potential of small grain/legume mixtures (Hayden et al. Reference Hayden, Ngouajio and Brainard2014; Sainju et al. Reference Sainju, Whitehead and Singh2005) due to functional complementarity.
Table 3 Cover crop biomass (mean with SE in parentheses) produced at the 4 site-years, with two-way ANOVA results of site-year and termination treatment analysis.Footnote a

a Means within the same column with the same lowercase letter (site-year effects) or uppercase letter (termination treatment effects) are not significantly different at α=0.1 (main effects separated using single degree of freedom contrasts).
b The wheat monoculture was not planted at F16.
Cover Crop Biomass—Termination Treatment and Site-Year Effects
The early-terminated cover crops produced less biomass than the late-terminated ones. For the cover crop mixture, the removal treatment was also associated with greater biomass compared with the late termination (averaged over site-years). It is unclear why this occurred, as the removal treatments were sampled before biomass removal, and this operation occurred either on the same day (both site-years in 2017; Table 2) or before (both site-years in 2016; Table 2) the late-termination treatment. Wheat cover crop biomass was greater at G16 compared with G17 and S17 (Table 3). Mixture cover crop biomass was also higher at G16 than at these two sites, with intermediate mixture biomass production at F16 (Table 3).
Cover Crop Mixture Composition
Cover crop mixture composition also differed between site-years and the termination treatments (Table 3). The greatest percentage of cover crop biomass composed of legumes was found at F16 (38.1%), while the lowest was found at S17 (13.7%). The F16 site-year had more legume biomass (proportionally) compared with G16 and S17, while S17 had proportionally less legume biomass than F16 and G17 (Table 3). Termination treatment also influenced the percentage of legume biomass, with early-terminated mixtures having, on average, 37% less legume biomass than the late-terminated mixture and mixture with residue removed (Table 3). Although we did not measure forage quality in this trial, differences in legume biomass would likely influence this. Canola was included in the mixture planted at G16; termination treatment did not affect canola biomass (P=0.599) or the proportion of canola in the total cover crop biomass at this site-year (P=0.184). Canola was approximately 50% of the dry cover crop biomass at G16 (unpublished data).
Early-Season Weed Density
The same species dominated each site-year at this sampling time—smooth pigweed (Amaranthus hybridus L.), prickly sida (Sida spinosa L.), carpetweed (Mollugo verticillata L.), and annual grass seedlings that were not identified to species. (During the midseason weed density evaluation, dominant annual grasses were identified as large crabgrass [Digitaria sanguinalis (L.) Scop.] and goosegrass [Eleusine indica (L.) Gaertn.].) With few exceptions (i.e., between two and four subplots per site-year), these species comprised greater than 50% of the total weed density at this time.
We only detected cover crop composition impacts on early-season weed density at 1 site-year (Table 4). At G16, weed density in the unsprayed subplots following the wheat monoculture cover crop was 2.1 weeds m−2, while weed density following the mixture was only 1.2 weeds m−2. Weed density was similar following the two cover crop types in the sprayed subplots. Detection of cover crop composition effects on early-season weed density at only 1 site-year, and a small effect at that site-year, provides support for our first hypothesis that soil nitrogen release from the inclusion of legumes in a cover crop mixture would have limited impacts on weed emergence in our high applied nitrogen cropping system. Interestingly, this effect of cover crop species composition on early-season weed density in the unsprayed subplots where only the cover crop residues were affecting weeds was noted only at G16. This site-year had similar amounts of wheat monoculture and cover crop mixture biomass (Table 3). It was also the only site-year to include canola, which made up a considerable portion (approximately 50%) of the dry biomass.
Table 4 Results (P-values) of a three-way ANOVA for early-season weed density (4–6 d after planting at G16 and F16, and 18 d after planting at S17) and for significant interactions.Footnote a

a Main plot factors were cover crop composition (C; mixture or wheat monoculture) and termination treatment (T; early or late termination, or cover crop residue removal), with + or – soil-residual herbicides (H) as the subplot factor. Mean response for the significant C*H interaction at G16 is discussed in the text; mean response for the T main effect and T*H interactions are shown in Figure 1.
b At F16, only cover crop mixture was examined, so no comparison between cover crop species composition was made.
c Early-season weed density was not assessed at the G17 site-year.
Termination treatment affected early-season weed density at all site-years, either as a main effect or through an interaction with the herbicide application (Table 4). At both G16 and S17, termination treatment affected early-season weed density only within the unsprayed subplots, though in different ways. In unsprayed subplots at S17, the late termination resulted in lower weed density compared with the early-termination and removal treatments (Figure 1). At G16, however, the late-termination treatment resulted in higher weed density compared with the other two termination treatments (Figure 1). Similarly, at F16, early weed density was higher in the late-termination treatment than in the early-termination and cover crop residue removal treatments across both sprayed and unsprayed subplots (Figure 1; main effect of termination treatment). (Wheat monoculture was not planted at F16, so a two-way ANOVA was conducted with the main plot factor of termination treatment and subplot factor of residual herbicide; this ANOVA indicated a significant termination treatment effect, but no interaction with the herbicide [Table 4].) Within the sprayed subplots at both G16 and S17, no termination treatment effects on early-season weed density were detected.

Figure 1 Early-season weed density (mean ± SE), measured in early to mid-June from 4 to 18 d after tobacco planting, across termination treatments. Weed density was not measured at this time at G17. Within each site-year, bars with the same letter are not significantly different at α=0.1. At G16 and S17, termination treatment only affected early-season weed density in subplots without the soil-residual herbicide (“unsprayed”). At F16, the main effect of termination treatment was significant, regardless of the herbicide applied.
Results observed at the S17 site-year provided some support for our second hypothesis that weeds would be suppressed by the thicker mulch layer provided by the late-termination treatment in the absence of the soil-residual herbicide. However, we did not observe this at the G16 and F16 site-years. Many studies have noted that, in the absence of soil-residual herbicides, increased amounts of cover crop residue generally correlate with lower weed density (Price et al. Reference Price, van Santen and Sarunaite2016; Teasdale et al. Reference Teasdale, Beste and Potts1991; Webster et al. Reference Webster, Scully, Grey and Culpepper2013). Our findings are not this consistent, as the G16 and F16 site-years generally had more cover crop biomass than S17 (Table 3).
Previous research has demonstrated that thinner cover crop mulches (produced from approximately 2,700 kg ha−1 oat (Avena sativa L.) cover crop biomass, less than in the current study; Table 3) had variable impacts on weed emergence depending on moisture conditions (Haramoto and Brainard Reference Haramoto and Brainard2017). Powell amaranth (Amaranthus powellii S. Watson) emergence was typically suppressed by surface cover crop residues in wetter conditions, but sometimes increased in drier conditions. In the current trial, S17 received more precipitation in May and early June compared with the two 2016 site-years (unpublished data), suggesting that greater weed density following the late-termination treatment in 2016 may have resulted from increased soil moisture under the cover crop residue. This effect was observed regardless of the herbicide treatment (Table 4), so it does not appear to be due to reduced efficacy of the soil-residual herbicides. The mixture at S17 did have the lowest percentage of legumes and was composed primarily of cereal rye, which perhaps explains why there was no impact of the cover crop species at this site-year.
It is also possible that the different responses to cover crop termination time, particularly between F16 and S17, which had similar weed density (Figure 1), were due to differences in the weed community. Annual grasses (later identified as D. sanguinalis and E. indica) were predominant at both site-years, comprising 41% and 59% of the total weed density at F16 and S17, respectively, at the early-season evaluation. Authors have noted variable results for annual grass suppression with cover crop residues in no-till systems. Digitaria sanguinalis emergence was either similar to or reduced following cereal rye residue compared with no residue (Brainard et al. Reference Brainard, Bryant, Noyes, Haramoto and Szendrei2016). Eleusine indica density was reduced following cereal rye and hairy vetch, relative to no residue, though impacts of these residues on density of this species and of D. sanguinalis were not consistent (Teasdale et al. Reference Teasdale, Beste and Potts1991). These studies highlight that multiple factors may regulate the influence of cover crop residues on weed emergence and density, perhaps including the regulation of soil moisture, as noted earlier.
We did not observe termination treatment effects in the subplots sprayed with residual herbicides at any site-year. This suggests that delaying termination from the early timing to the late timing did not result in significant interception of the soil-residual herbicides. Given the lack of impact between the late-termination time (i.e., thick cover crop residue layer) and the removal treatment (no cover crop residue), it also suggests that leaving the cover crop residue in the field did not contribute to herbicide interception. However, ample rainfall (unpublished data) soon after application in all years likely mitigated this potential and limited our ability to test our third hypothesis. Each site-year received at least 10 mm of precipitation within 1 to 5 d of the herbicide application. G16 received 84.8 mm of precipitation between the application and this first density evaluation, F16 received 66.3 mm, and S17 received 80.8 mm. (G17 also received 84.6 mm of precipitation between the herbicide application and planting, including almost 24 mm within 2 d of application.) Repeating this study in drier conditions would be necessary to gain a better understanding of the effect of cover crop residue on residual herbicide efficacy. The current results, however, demonstrate the application of previous work on herbicide degradation and release kinetics (Banks and Robinson Reference Banks and Robinson1986; Carbonari et al. Reference Carbonari, GLGC, MLB, JRM and Velini2016; Ghadiri et al. Reference Ghadiri, Shea and Wicks1984; Reddy et al. Reference Reddy, Locke, Wagner, Zablotowicz, Gaston and Smeda1995)—if there is ample rainfall after application, soil-residual herbicides applied to cover crop residues can still be effective in reducing weed density (Figure 1).
Midseason Weed Density
At this evaluation, more differences in the weed community composition were apparent. F16 was still dominated by annual grasses and broadleaves, with D. sanguinalis being the dominant grass and A. hybridus, M. verticillata, and hairy galinsoga (Galinsoga quadriradiata Cav.) being the dominant broadleaf weeds (comprising >60% of the total weed density in all but three subplots). G16 and G17 were dominated by D. sanguinalis, S. spinosa, and M. verticillata; these species represented >60% of the total weed density in all but one subplot at both site-years. The community at S17 was more diverse; annual grasses (E. indica and D. sanguinalis) continued to comprise approximately 40% of the total weed density, but annual fleabane [Erigeron annuus (L.) Pers.] and dandelion (Taraxacum officinale F. H. Wigg.) were more dominant broadleaves. Amaranthus hybridus and M. verticillata were also present to a lesser extent.
Density measurements conducted in mid-July were approximately 30 d after planting (DAP) for all sites except S17 (Table 2). This site was planted earlier, so counts were performed at 60 DAP. Species composition (i.e., cover crop mixture or wheat monoculture) influenced midseason weed density at only 1 site-year (Table 5). Termination treatment affected midseason weed density at all site-years, interacting with the soil-residual herbicide at 1 site-year (Table 5).
Table 5 Results (P-values) of a three-way ANOVA for midseason weed density and for slicing interactions.Footnote a

a Main plot factors were cover crop composition (C; mixture or wheat monoculture) and termination treatment (T; early or late termination, or cover crop residue removal), with + or – soil-residual herbicides (H) as the subplot factor. Mean response for the C*H interaction is shown in Figure 2A, while the mean response for the T*H interaction and the T main effects are shown in Figure 2B.
b At F16, only cover crop mixture was examined, so no comparison between cover crop species composition is available.
As with early-season weed density, cover crop species composition only affected midseason weed density in 1 site-year—G16. In contrast to earlier in the season, however, differences were detected in the sprayed subplots. Midseason weed density was greater following the mixture than following the wheat (Figure 2A), but was similar following both the mixture and the wheat in unsprayed subplots. Although this site-year was associated with higher cover crop mixture biomass than most of the other site-years (Table 3), we expected that the large amounts of inorganic nitrogen fertilizer (i.e., 280 kg N ha−1) would have overwhelmed any nitrogen contributions from increased legume biomass. Why we observed differences in weed density between the cover crop types, and only in the sprayed subplots, remains unclear.

Figure 2 Midseason weed density (mean ± SE) measured in mid-July, from 30 to 60 d after tobacco planting at G16 (A, effect of the herbicide*cover crop composition) and at multiple site-years (B, effect of termination treatment). In A, bars within the same subplot (i.e., sprayed or unsprayed) with the same uppercase letter are not significantly different at α=0.1 according to single degree of freedom contrasts. Also in A, bars with the same cover crop composition (i.e., mixture or wheat monoculture) with the same lowercase letter are not significantly different at α=0.1 according to single degree of freedom contrasts. In B, bars with the same uppercase letter within each site-year or site-year*herbicide combination are not significantly different at α=0.1. Additionally, for S17, bars within the same termination treatment across residual herbicide treatments with the same lowercase letter are not significantly different at α=0.1.
Prevalence of both M. verticillata and annual grasses changed between the early- and midseason density evaluations at G16. Mollugo verticillata was present in 20% of the subplots early in the season, increasing to 67% of subplots by midseason. While it represented only 10% of the total weed density early in the season, it was 20% of the total weed density by midseason. Annual grasses were noted in 48% of subplots at the early-season evaluation, and this increased to 88% of subplots by the midseason evaluation. Annual grasses represented 28% of total weed density early in the season, increasing to 48% by midseason. In a system without soil-residual herbicides, M. verticillata density was reduced by cover crop residues in no-till sweet corn (Zea mays L.) in 3 of 4 yr (Teasdale et al. Reference Teasdale, Beste and Potts1991). However, density was typically lower following leguminous hairy vetch than following cereal rye; this reduction was significant in one of four years, and M. verticillata density was never lower following cereal rye compared with hairy vetch (Teasdale et al. Reference Teasdale, Beste and Potts1991). We are not aware of any other research suggesting why its emergence may be suppressed by different types of residue. As noted previously, impacts of cover crop residues on annual grass density can also be variable (e.g., Brainard et al. Reference Brainard, Bryant, Noyes, Haramoto and Szendrei2016; Teasdale et al. Reference Teasdale, Beste and Potts1991). Digitaria sanguinalis and E. indica densities were often numerically (though not statistically) lower following cereal rye residue compared with hairy vetch residue where soil-residual herbicides were not used (Teasdale et al. Reference Teasdale, Beste and Potts1991), a finding in contrast to ours. Given the differential response of weed density to these cover crop compositions at this site-year, and that this site-year was the only one to include canola (which was up to 50% of the cover crop biomass), this mixture warrants further study for weed management.
Termination treatment also impacted midseason weed density at the other site-years, either as a main effect (G16, F16, and G17), or interacting with the herbicide (S17; Table 5). At S17, midseason weed density was similar following the early- and late-termination treatments, but only in the unsprayed subplots, as termination treatment did not affect weed density in sprayed subplots (Table 5). At G16 and F16, the late-terminated treatment resulted in lower midseason weed density than both the early-terminated and removal treatments (Figure 2B)—the opposite of what was observed earlier in the season. At G17, however, midseason weed density was highest following the late-termination treatment. This was likely due to poor tobacco establishment in the late-termination treatment at this site-year (Table 6). Compared with the early-terminated and removal treatments, the number of harvestable tobacco plants in the late-terminated treatment was reduced by approximately 40% (Table 6). Lack of crop plants led to a reduced competition environment, likely allowing for more weeds to emerge. This stand reduction, which occurred only at this site-year, was likely due to the transplanter not being adjusted properly and highlights the difficulty that growers can face when transplanting into intact cover crop residues in no-till situations.
Table 6 Mean (SE) number of tobacco plants and yield at the 4 site-years, with two-way ANOVA results (P-values) of cover crop composition and termination treatment.Footnote a

a Tobacco was only harvested from the + soil-residual herbicide subplots. Within the same column, means followed by the same letter are not significantly different at α=0.1.
The soil-residual herbicide applied in May continued to be effective in reducing midseason weed density at all site-years (Table 5). The herbicide reduced weed density by 80% at F16 (from 285 weeds m−2 in unsprayed subplots to 54 weeds m−2 in sprayed subplots) and by 78% at G17 (from 144 weeds m−2 in unsprayed subplots to 32 weeds m−2 in sprayed subplots). At some site-years, the herbicide efficacy was influenced by other factors (e.g., G16, interaction with species; S17, interaction with termination treatment). At G16, the residual herbicide reduced weed density by 65% following the mixture and by 82% following the wheat (Figure 2A); while it was effective in reducing density following both types of cover crops, its efficacy at this time was reduced somewhat following the mixture. This effect was not observed earlier in the season (Table 4) or in any of the other site-years (Table 5). It is possible that more nitrogen was present in the soil at G16 due to the larger amount of cover crop mixture biomass (Table 3). It is also possible that the canola, which was only present at G16, also played a role in mediating herbicide efficacy. However, lack of impact on the early-season weed density seems to refute this possibility.
S17 was the only site-year to show differences in the herbicide efficacy between termination treatments (Table 5). Weed density was 47% to 58% lower in the removal and early-termination treatments, respectively, with the soil-residual herbicide compared with the unsprayed subplots (Figure 2B, lowercase letters). Density was not affected, however, by the herbicide application in the late-termination treatment (Figure 2B, lowercase letters). There is not evidence that the herbicide was less effective in the late-termination treatment, as density was similar across all termination treatments in the sprayed subplots (Figure 2B, uppercase letters). This, then, suggests that, without soil-residual herbicides, the residue in this treatment was still providing weed suppression to a similar degree as the cover crop residue plus herbicides.
Weed Biomass at Harvest
Weed biomass measured at harvest followed trends in weed density from earlier in the season at some site-years, but not others. Cover crop species composition influenced weed biomass at harvest only at G16, and the termination treatment influenced weed biomass at the other 2 site-years at which it was measured (Table 7).
Table 7 Results (P-values) of a three-way ANOVA on weed biomass at harvest and for slicing interactions.Footnote a

a Main plot factors were cover crop composition (mixture or wheat) and termination treatment (early or late termination, or cover crop residue removal), with + or – soil-residual herbicides as the subplot factor.
b Weed biomass was not measured at F16.
As with midseason weed density at G16, cover crop species composition influenced final weed biomass in unsprayed subplots (Figure 3A) but not in sprayed subplots. Final weed biomass in the unsprayed subplots following wheat, however, was much different than midseason weed density. Although there were more weeds midseason following the wheat monoculture in the unsprayed subplots compared with the sprayed subplots (Figure 2A, lowercase letters), final weed biomass was similar (Figure 3A, lowercase letters). Above- and belowground residue from the wheat cover crop may have been immobilizing nitrogen, leading to decreased growth of emerged weeds and decreasing biomass (i.e., Wells et al. Reference Wells, Reberg-Horton, Smith and Grossman2013). That this composition effect was observed at G16 but not the other site-years may have been due to the larger amounts of wheat cover crop biomass produced at this site-year compared with the others (Table 3). Lack of impact on the tobacco yield (Table 6) somewhat refutes nitrogen immobilization as a potential explanation, though the more competitive tobacco plants may have been better able to use available soil nitrogen. Allelopathy from wheat residues (Wu et al. Reference Wu, Pratley, Lemerle and Haig2001) could also be responsible for reducing weed biomass at this site-year with more cover crop biomass. Cereal rye, a component of the cover crop mixture, is also noted to be allelopathic (Weston and Duke Reference Weston and Duke2003). However, cereal rye comprised only about 40% of the mixture biomass at G16 (unpublished data).

Figure 3 Weed biomass (mean +/- SE) measured at harvest at G16 (A, effect of herbicide*cover crop species composition) and at multiple site-years (B, effect of termination treatment, with and without soil residual herbicide if significant). In A, bars with the same letter (upper or lower case) are not significantly different at α=0.1 according to single degree of freedom contrasts. In B, bars with the same capital letter within each site-year are not significantly different at α=0.1. Note that the ANOVA for S17 required grouping to account for unequal variances. Within S17 in B, bars from the same termination treatment with the same lowercase letter are also not significantly different at α=0.1 according to single degree of freedom contrasts.
Termination treatment influences on final weed biomass at G17 and S17 were similar to their impacts on midseason weed density. At G17, the late-termination treatment resulted in greater weed biomass at harvest (Figure 3B). This showed a similar trend to the midseason weed density and is likely explained by the poor tobacco stand in this treatment. At S17, final weed biomass was affected by termination treatment in both sprayed and unsprayed subplots. Trends, however, were similar to the midseason weed density. In the unsprayed subplots, biomass was lowest in the late termination and similar in the other two; these two termination treatments had similar impacts on midseason weed density without the residual herbicide (Figure 2B). Weed biomass at harvest was low in the subplots with residual herbicides—less than 40 g m−2 in all treatments. The early-terminated treatment had lower biomass than the other two treatments. This may be explained by higher yields in the early-terminated treatment (see below).
The residual herbicide applied in May continued to influence weed biomass at harvest. At G16, the herbicide reduced biomass by 75% following the mixture (Figure 3A, lowercase letters on black bars) but did not influence biomass following wheat (Figure 3A, lowercase letters on gray bars). At G17, herbicide also resulted in 75% less weed biomass across both cover crop species and all termination treatments (from 154.4 g m−2 in unsprayed subplots to 37.6 g m−2 in sprayed subplots). At S17, the herbicide reduced weed density in the sprayed subplots relative to unsprayed subplots by 97% in early terminated, by 81% in late terminated, and by 92% in removal (Figure 3B). Thus, by the end of the season, only the wheat at G16 showed evidence that it was still suppressing weeds in the absence of the soil-residual herbicides (Figure 3A). Midseason density was high in this treatment (Figure 2A), but these weeds did not accumulate much biomass by the end of the season. Because we did not collect yield separately in the sprayed and unsprayed subplots, we cannot conclude decisively whether these weeds impacted yield. However, it is not uncommon for small grain cover crop residues to reduce nutrient availability, resulting in smaller weeds (Wells et al. Reference Wells, Reberg-Horton, Smith and Grossman2013).
Tobacco Stand and Yield
Cover crop composition did not influence the number of tobacco plants or yield at any site-year, but as noted earlier, tobacco stand was influenced by cover crop termination time (Table 6). At G16, fewer plants were found in the early-termination treatment compared with the removal treatment, with an intermediate stand in the late-terminated treatment (Table 6). These differences, however, were small, and yield at this site-year was not influenced by the termination treatment. At G17, the reduced stand in the late-termination treatment likely led to reduced yield (Table 6). Finally, at S17, stand was not affected by treatment, but yield in the early-termination treatment was higher than in the other two treatments (Table 6). Waterlogged soils at this site caused some plant damage, which may have reduced yields in these treatments (RP, personal observation). It is possible that the increased root biomass from late-terminated cover crops was holding more soil moisture, which exacerbated the impact of flooding; tobacco roots are sensitive to anaerobic conditions. This site-year demonstrates the importance of crop–weed competition—final weed biomass was lower (Figure 3B) and tobacco yield higher (Table 6) following the early-termination treatment in the sprayed subplots (yield was only collected from the sprayed subplots).
Overall, the results give the most support for our hypothesis that the late-termination treatment would lead to the most weed suppression. This effect was consistently observed at one of the site-years (S17). This site-year had the shortest time between the residual herbicide application and planting, and tobacco plants were the oldest at all density evaluations and thus likely exerting the most competitive influence. At 2 site-years, the late-termination treatment resulted in higher early-season weed density, perhaps due to increased soil moisture during a dry period, but lower midseason weed density. Impacts on weeds were also mediated by poor crop establishment at 1 site-year (G17)—the late-termination treatment resulted in reduced tobacco establishment; the resulting competition-free environment led to more weeds emerging and higher densities in the late-terminated treatment. By harvest, impacts on weed density mostly continued to manifest in impacts on weed biomass. There was no interaction with the soil-residual herbicide application at these site-years by the midseason density evaluation, which suggests that there was no immobilization and subsequent release of the herbicide by the cover crop residues. It is likely that the higher amount of residue in the late-terminated treatment continued to suppress weed emergence through this midseason evaluation time.
Cover crop species composition only influenced weeds at 1 of 3 site-years (G16) for which we could compare a cover crop mixture to a monoculture wheat cover crop. In the absence of soil-residual herbicides, the mixture seemed to suppress early weed emergence better than the wheat monoculture cover crop. By midseason, weed density was similar following both cover crop types without herbicides, but was again lower following the mixture where herbicides were applied (Figure 2A). By harvest, however, weed biomass was much lower following wheat than following the mixture in the absence of soil-residual herbicides (Figure 3A). This site-year consistently had higher cover crop biomass than the other site-years (Table 3) and also had canola in the cover crop mixture. Canola could have led to differences in the residue composition that we could not study at other site-years. Given, however, that these differences were only observed at 1 site-year, our results show minimal additional weed management benefit of more complex, and more expensive, cover crop mixtures. It is important to note as well that our cover crop mixture included cereal rye as a small grain, which was compared with a monoculture wheat cover crop. While we are not aware of any studies comparing performance of mixtures containing wheat with mixtures containing cereal rye, it is possible that there are key differences between them that should limit the applicability of direct comparisons of our cereal rye–containing mixture with the monoculture wheat. In addition, while some legume-produced nitrogen will be available soon after termination (Jackson Reference Jackson2000), higher amounts of nitrogen release from legume biomass may take years to accrue, particularly in no-till (Villamil et al. Reference Villamil, Bollero, Darmody, Simmons and Bullock2006). Thus, the short-term nature of our study (conducted over the course of 1 yr at each site) limits our ability to capture these longer-term benefits. On the other hand, our results demonstrate that growers wishing to adopt mixtures for forage production to receive additional revenue or to capture these longer-term benefits can receive similar weed management benefits in a tobacco production system that uses soil-residual herbicides. For growers seeking to better utilize cover crops in reduced-tillage tobacco production, our results suggest that, over the short term, weed management benefits will be similar between a monoculture wheat cover crop and a cereal rye/forb mixture. Our results highlight the importance of transplanter adjustment when planting into cover crop residues, as poor crop stands can lead to increased weed pressure. With adequate tobacco stands, however, later cover crop termination and soil-residual herbicides provided the most consistent weed control over all site-years.
Acknowledgments
Altria provided funding for this project. No conflicts of interests have been declared. We thank Troy Rankin, Justin Wilmot, and David Talley for use of their fields for this experiment. Jon Anderson with Kentucky Department of Fish and Wildlife planted the cover crops and the tobacco. Thank you also to Matthew Allen, Jack Zeleznik, Tori Stanton, Austin Sherman, Vinicius dos Santos Cunha, Michael Hurak, Haley Taylor, Grace Ragain, Nolan Stephenson, and Fabian Leon for field assistance.










