



INTRODUCTION
The rich fossil record of Testudinidae (order Testudines), the clade commonly known as tortoises, extends from the Late Eocene (Vlachos and Rabi, Reference Vlachos and Rabi2018) or Late Paleocene (Branch, Reference Branch2016) to the Holocene. Compared with this extensive body fossil record, the trace fossil record is non-existent, despite tortoises forming a widespread, familiar group of extant reptiles. Roberts et al. (2008, p. 262) hinted at the potential of Pleistocene aeolianite exposures on the Cape south coast of South Africa in this regard: “They contain a rich archive of fossil mammalian trackways… as well as reptilian (testudinae).” However, no details were provided. Although absent from Australia and Antarctica, tortoises have an otherwise global distribution, and are present both on continents and islands (Ernst and Barbour, Reference Ernst and Barbour1989). Southern Africa contains the greatest diversity of extant tortoises, hosting 5 genera and 13 species of a global count of 17 genera and 58 species (Branch, Reference Branch2016).
We use the term ‘chelonian’ to denote a reptile of the order Testudines, to include sea-turtles, terrapins (to which southern African freshwater turtles are referred), and tortoises (which can be termed ‘terrestrial turtles’). The earliest known chelonian tracks are from the Early Triassic, and are older than the oldest known chelonian body fossils (Lichtig et al., Reference Lichtig, Lucas, Klein and Lovelace2017). Chelonian giantism is a recurring theme in the fossil record, which in historical times is largely confined to islands. However, tortoise giantism evolved independently in multiple continental taxa (Vlachos and Rabi, Reference Vlachos and Rabi2018), and was not limited to islands (Itescu et al., Reference Itescu, Karraker, Raia, Pritchard and Meiri2014). The largest known species, Megalochelys atlas, had a reported carapace length of ~2.1 m and estimated total length of 2.5–2.7 m. Its extinction by the Middle Pleistocene has been attributed to Homo erectus (Turtle Extinctions Working Group, Reference Rhodin, Thomson, Georgalis, Karl, Danilov, Takahashi and de la Fuente2015).
Describing a fossil giant tortoise from continental Africa, Geochelone laetoliensis, Harrison (Reference Harrison and Harrison2011) suggested that the Late Pliocene extinction of giant tortoises on the African mainland was associated with the appearance of Homo and the use of stone tool technology at ca. 2.6 Ma, and that natural selection would have favored smaller tortoise species. The recent extinction of many tortoise species has been correlated with humanity's spread (Turtle Extinctions Working Group, Reference Rhodin, Thomson, Georgalis, Karl, Danilov, Takahashi and de la Fuente2015). Today, the largest tortoise occurring on a continental mainland is the African spurred tortoise (Centrochelys sulcata) in the Sahel, which reaches lengths of 0.83 m (Branch, Reference Branch2016).
Other than occasional visits by sea-turtles for unsuccessful nesting attempts, the only large reptile that currently occurs along the Cape south coast is the leopard tortoise (Stigmochelys pardalis) (Branch, Reference Branch1994). In exceptional cases in the eastern Cape, it can reach 70 cm in length, with a mass of 40 kg (Branch, Reference Branch1994). Its range extends from the southern Cape to Ethiopia; the largest specimens are found at the northern and southern ends of the range (Branch, Reference Branch2016).
There are no historical records of other large reptiles in the southern Cape, and the Pleistocene body fossil record is devoid of reliable reports of crocodylians, varanids, or sea-turtles. Although tortoise remains are prevalent in regional paleontological or archaeological sites, evidence for large tortoises is extremely rare. However, Pleistocene ichnosites from the Cape south coast have been attributed to large crocodylians (Helm et al., Reference Helm, Cawthra, Combrink, Helm, Rust, Steer and van den Heever2020a; Helm and Lockley, Reference Helm and Lockley2021) and hatchling sea-turtles (Lockley et al., Reference Lockley, Cawthra, De Vynck, Helm, McCrea and Nel2019a), demonstrating the capacity of the trace fossil record to complement the body fossil record.
Identification of further evidence of large reptiles on the Cape south coast is therefore of interest. We report here on a trackway site east of Still Bay, attributed to a large tortoise trackmaker, and tracks from five other sites that are attributed to tortoises and terrapins (Fig. 1). The aim is to describe these ichnosites, provide new age constraints, consider trackmaker candidates, provide trackmaker length estimates, and discuss paleoenvironmental and archaeological implications.
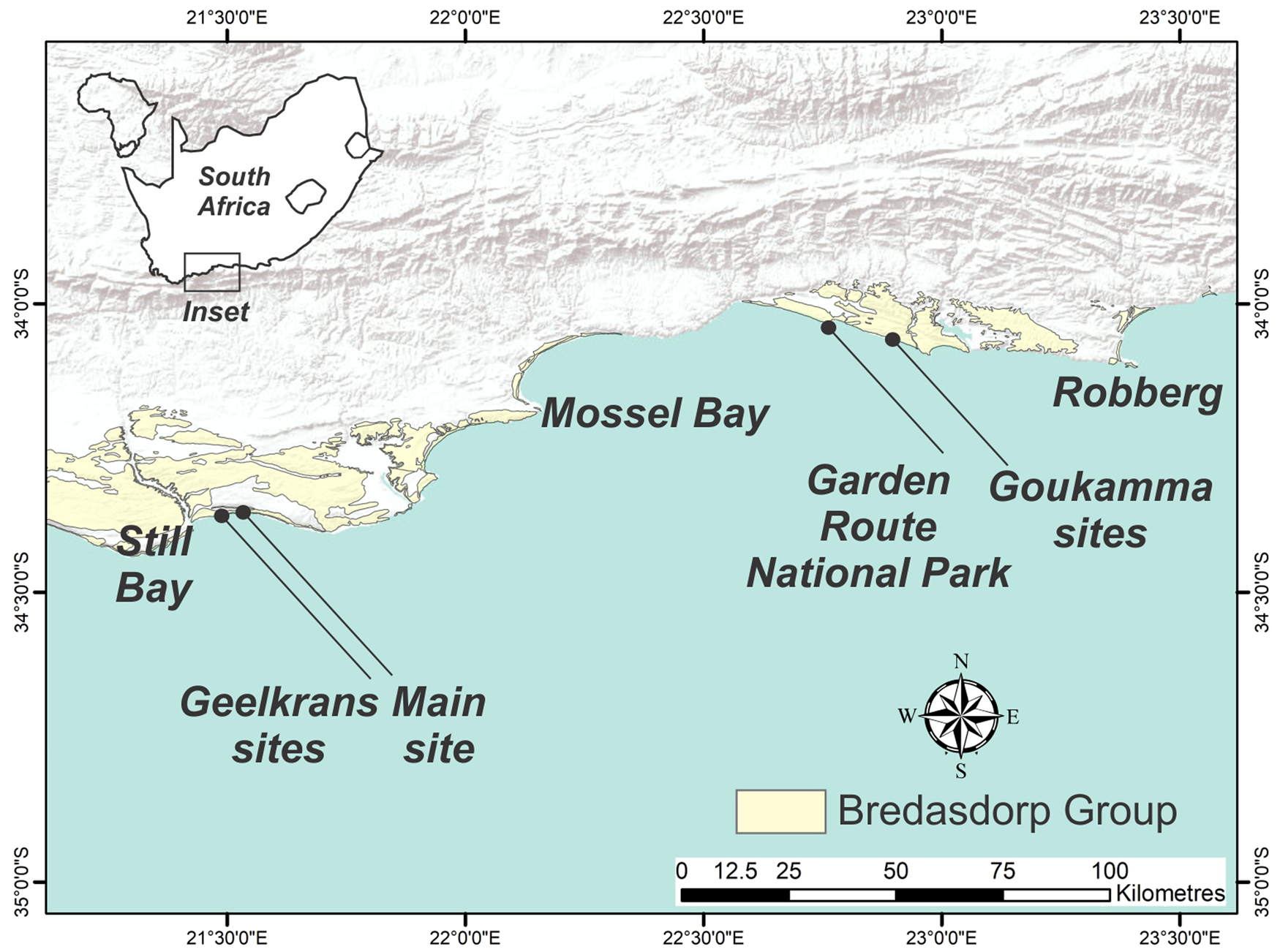
Figure 1. Locality map showing Bredasdorp Group sediments on the Cape south coast of South Africa, and chelonian ichnosites.
GEOLOGICAL CONTEXT
Extensive exposures of Pleistocene cemented dunes (aeolianites) are found along the Cape south coast. They are concentrated in the coastline's embayments and have provided evidence for paleo-shorelines and paleo-coastal dune activity throughout the Middle and Late Pleistocene (Roberts et al., Reference Roberts, Cawthra and Musekiwa2013). Most of the exposures are mapped into the Waenhuiskrans Formation (Malan, Reference Malan1989), with broadly coeval shoreface, foreshore, and lagoonal deposits mapped into the Klein Brak Formation (Malan, Reference Malan1991). Both form part of the Bredasdorp Group. The lithology of the dunes is conducive to the preservation of a variety of sedimentary structures, paleosols (Roberts et al., Reference Roberts, Bateman, Murray-Wallace, Carr and Holmes2008), and faunal tracks and traces, which have been studied as part of the Cape south coast ichnology project. This has identified >300 Pleistocene vertebrate ichnosites, spanning 350 km of coastline between Arniston and the Robberg Peninsula (Fig. 1).
The tracksites would have been situated at the margin of the Paleo-Agulhas Plain, most of which is currently submerged. Pleistocene climate changes led to glacio-eustatic sea-level oscillations, with maximum regressions (e.g., MIS 2) exposing the entire Plain (Marean et al., Reference Marean, Cowling and Franklin2020). In contrast, during the MIS 5e sea-level highstand, at ca. 126 ka, sea level was 6–8 m higher than at present (Carr et al., Reference Carr, Bateman, Roberts, Murray-Wallace, Jacobs and Holmes2010).
Optically stimulated luminescence (OSL) dating of sites along this coastline has demonstrated that most onshore aeolianites date to MIS 5 and late MIS 6 (Roberts et al., Reference Roberts, Bateman, Murray-Wallace, Carr and Holmes2008; Carr et al., Reference Carr, Bateman, Roberts, Murray-Wallace, Jacobs and Holmes2010; Bateman et al., Reference Bateman, Carr, Dunajko, Holmes, Roberts, Mclaren, Bryant, Marker and Murray-Wallace2011; Cawthra et al., Reference Cawthra, Jacobs, Compton, Fisher, Karkanas and Marean2018). MIS 11 deposits (Roberts et al., Reference Roberts, Karkanas, Jacobs, Marean and Roberts2012) and MIS 3 deposits (Carr et al., Reference Carr, Bateman, Cawthra and Sealy2019) are less common. The abundance of tracksites results from a combination of a cohesive molding agent provided by moist sand, rapid track burial facilitated by high sedimentation rates, rapid lithification via partial solution and re-precipitation of bioclasts, and re-exposure of track-bearing surfaces through shoreline erosion (Roberts and Cole, Reference Roberts and Cole2003). The first factor may be enhanced through the binding of sand by microbial activity (Seilacher, Reference Seilacher2008).
The main tracksite described here was identified in a remote section of coastline (Fig. 1). Here cliffs as high as 50 m extend alongshore for 6 km. Sedimentary facies are dominated by moderate- to high-angle, large scale planar and wedge-planar cross-stratification, which is the result of migration of large-scale dunes (Roberts et al., Reference Roberts, Bateman, Murray-Wallace, Carr and Holmes2008). Slipface dips are close to the angle of repose of wind-blown sand, and their orientations indicate a dominant wind from the west during MIS 5e (Roberts et al., Reference Roberts, Bateman, Murray-Wallace, Carr and Holmes2008). High tides resulted in the collapse of cliff sections. Following dislodgement, loose blocks lie on unstable slopes or near the high tide mark, and may eventually slump into the sea.
This stretch of coastline is rich in ichnosites, some of which are important complements to the body fossil record. These include sea-turtle tracksites (Lockley et al., Reference Lockley, Cawthra, De Vynck, Helm, McCrea and Nel2019a), a sand-swimming golden mole site (Lockley et al., Reference Lockley, Helm, Cawthra, De Vynck and Perrin2021), a giraffe tracksite (Helm et al., Reference Helm, Cawthra, Cowling, De Vynck, Marean, McCrea and Rust2018a), and hominin traces (Helm et al., Reference Helm, Cawthra, De Vynck, Helm, Rust and Stear2019, Reference Helm, Cawthra, De Vynck, Helm, Rust and Stear2021a). Prior to our studies, only one tracksite along this coastline had been directly dated (Roberts et al., Reference Roberts, Bateman, Murray-Wallace, Carr and Holmes2008). Situated 700 m east of the main tracksite described here, it comprised elephant and bovid trackways. OSL dating produced an age range of 140 ± 8 ka to 91 ± 5 ka for the oldest to youngest dated units. The assumption has therefore been that these cliffs contain deposits dating to between MIS 6 and MIS 5b. The main tracksite described here was situated on a large, loose block in an area that had undergone significant cliff-collapse (Fig. 2).

Figure 2. (A) Arrow indicates the track-bearing block (partly covered by vegetation) on an unstable slope containing multiple large loose slabs. (B) The track-bearing block is in the foreground, facing seawards; in the distance to the east are aeolianite cliffs and the Indian Ocean. The dimensions of the track-bearing surface are ~150 × 150 cm.
TORTOISE TRACK MORPHOLOGY
Liebenberg (Reference Liebenberg2000), van den Heever et al. (Reference van den Heever, Mhlongo and Benadie2017), and Stuart and Stuart (Reference Stuart and Stuart2019) provide track descriptions of extant southern African tortoises, ranging in size from the tiny speckled padloper (Chersobius signatus) to the large leopard tortoise (Stigmochelys pardalis). Most species have five claws on the front feet and four claws on the hind feet. Tramline trackways with a wide straddle and relatively closely spaced tracks are characteristic (Fig. 3A). Drag marks are commonly made by the claws of the hind feet (van den Heever et al., Reference van den Heever, Mhlongo and Benadie2017). Scale impressions may be recorded in larger tracks under ideal substrate conditions (van den Heever et al., Reference van den Heever, Mhlongo and Benadie2017).

Figure 3. (A) A tramline trackway made by a leopard tortoise, showing a wide straddle and closely spaced tracks. (B) Tortoise tracks showing scale impressions and inward-pointing manus claw impressions (reproduced from https://imgur.com/a/jHq95). (C) Leopard tortoise manus track in wet sand; claw impressions of digits I, II, and III are deep, claw impression of IV is faint, and claw impression of V is absent. (D) Plastron of a leopard tortoise, showing longitudinal markings.
The track of the front foot (manus) faces inwards, whereas the track of the hind foot (pes) faces outwards (Fig. 3B). Pes tracks are slightly larger than manus tracks, and have an oval shape, with the long axis directed slightly outward relative to the direction of motion. Manus tracks are more rounded. The depressions made by the claws are prominent, facing inwards with manus tracks, and outwards with pes tracks (van den Heever et al., Reference van den Heever, Mhlongo and Benadie2017). In both manus and pes tracks the claws of digits I, II, and III are registered deepest; in manus tracks the digit IV claw impression is typically fainter and the digit V claw impression may be absent (Fig. 3C). Sometimes only claw impressions are registered, in what are referred to as toe-tip traces. These often occur in triplets, representing digits I, II, and III, and may occur during upslope travel and on firm surfaces. A slow walking gait results in the pes track being registered partially on the proximal portion of the manus track, producing a composite track. However, variations are possible and the pes track may appear behind the manus track, or may be superimposed on it.
Tracks registered in soft substrates (e.g., dry, unconsolidated sand) may show no morphological details. In such cases, the characteristic wide-straddle, tramline pattern with close spacing of tracks may provide the only indications of a tortoise trackmaker, and pes and manus tracks may coalesce into crater-like depressions. On firm surfaces, the plastron does not usually make contact with the substrate, but this may occur with softer substrates or when the trackmaker is tired or struggling; traces of this contact occur close to the trackway midline. Longitudinal striations oriented in the direction of motion may be detectable, indicating where the substrate was scraped by the plastron. The underside of the tortoise plastron often exhibits longitudinal grooves and ridges (Fig. 3D), which may cause the striations in such traces.
The tortoise tail is directed inferiorly and slightly posteriorly (Fig. 3B), and may be used to assist locomotion if the trackmaker is struggling. This takes the form of a dagger-like spearing of the substrate, resulting in a round depression. Rather than tail-drag impressions, which are characteristic of many reptile groups, round midline depressions may thus be seen. Displacement rims are commonly present, occurring most prominently behind tracks. Such rims are more evident during upslope travel due to gravitational effects. In summary, tortoise trackways may exhibit a variety of forms, and there is no single ideal trackway morphotype.
METHODS
Global Positioning System readings were taken at all tracksites using a hand-held device. Locality data were reposited with the African Centre for Coastal Palaeoscience at Nelson Mandela University, to be made available to researchers upon request.
Photographs at the main tracksite were taken during different lighting conditions on repeat visits. Measurements included track length and width, pace length (where feasible), inter-track length (from the posterior margin of one track to the posterior margin of the track ahead of it), interdigital distance, and external straddle (between outermost borders of the lines of the ‘tramline’, measured perpendicular to the direction of travel [sensu van den Heever et al., Reference van den Heever, Mhlongo and Benadie2017]). Results were recorded in centimeters. Positions of displacement rims were noted. Because of the unstable nature of the main tracksite, photogrammetric analysis was not feasible. However, this was performed at some of the other sites (sensu Matthews et al., Reference Matthews, Noble, Breithaupt, Falkingham, Marty and Richter2016). 3D models were generated with Agisoft Metashape Professional (v. 1.0.4) using an Olympus Tough model TG-6 camera (focal length 4.5 mm; resolution 4000 × 3000; pixel size 1.56 × 1.56 μm). The final images were rendered using CloudCompare (v.2.10-beta).
Leopard tortoises (two adult leopard tortoises at the Wild Rescue Wildlife Sanctuary and Nature Reserve, and 10 at FreeMe Wildlife Rehabilitation), their gaits, locomotion, and trackways were observed and analyzed. Measurements included carapace length and width, distance between manus claw impressions, distance between pes claw impressions, external straddle, and track length (including claws) and width for both manus and pes. Total body length was measured with head and neck gently protruding, without forced extension.
In the case of crocodylian trackways, total trackmaker length can be estimated either from the pes track length, according to a 12:1 ratio determined for the Nile crocodile (Crocodylus niloticus) by Thomson (Reference Thomson1972), or from interdigital distance (Helm and Lockley, Reference Helm and Lockley2021; Plint et al., Reference Plint, Helm and Lockley2022). Hitherto this has not been attempted for testitudinal trackways. In order to estimate tortoise trackmaker length, we used the ratios derived from our leopard tortoises neoichnological studies, based on the ratios of their length to external straddle, manus track length, and inter-claw distance. These results are estimates based on proxies that are variable. For example, straddle may vary slightly depending on gait, and as with crocodylians, inter-claw distance is not fixed (Sadlok and Pawełczyk, Reference Sadlok and Pawełczyk2021). The length estimates provided are merely intended to provide approximate size indications.
Samples for OSL dating of the aeolianite sediment were obtained from the region of the main tracksite, from a loose slab 1.5 km to the west (Leic20031), and from an in situ site 500 m to the east (Leic20024). Samples were obtained from near the other sites, as detailed below. The samples were shipped to the University of Leicester (U.K.), where they were fragmented under subdued red light to obtain unexposed sand-sized sediment. The samples were prepared for analysis of the coarse-grained (usually in the range 150–250 μm) quartz fraction. Equivalent doses were determined using the single aliquot regeneration method (Murray and Wintle, Reference Murray and Wintle2000, Reference Murray and Wintle2003). Details of the associated measurement conditions and quality control procedures are provided as supplementary information. Dose rates were determined using elemental concentrations obtained using inductively coupled plasma mass spectrometry (ICP-MS; for U and Th) and ICP-OES (for K). Details of the dose rates, associated dose-rate calculations, and modeling experiments concerning dose-rate changes through time due to water loss and concurrent carbonate cementation (Nathan and Mauz, Reference Nathan and Mauz2008; Mauz and Hoffman, Reference Mauz and Hoffmann2014), are provided as supplementary information.
RESULTS
Main site, east of Still Bay
The track-bearing surface measured ~150 × 150 cm. The thickness of the block, which lay in an unstable fan of slumping sandy debris amid multiple loose blocks, was >100 cm. The block, which was derived from a well-laminated aeolianite unit, formed the lowest of a sequence of three large slabs of similar thickness, stacked in layers on the debris slope, and appeared more blocky in section than did the well-bedded uppermost slab (Fig. 2A). The track-bearing surface faced south, towards the sea, at an angle of ~60° to the horizontal (Fig. 2B). The tracks were registered in epirelief. Examination of the surface in sunlight, in shadow, and at different times, allowed for full appreciation of the diversity of surface trace fossil features and sedimentary structures.
The cliffs above, from which the block originated, were largely covered by debris, and the layer of origin could not be determined, but appeared to have been a considerable height above modern sea level. However, 40 m to the east, 15 m high vertical cliffs contained paleosols, a thick, poorly bedded unit, and aeolian cross-bedded units with interdune horizons. Features most consistent with those in the track-bearing block were present about half-way up these cliffs (Fig. 2B).
A fine-grained sedimentary rock veneer was initially present on the track-bearing surface, allowing fine detail of traces to be distinguished (e.g., M1, see below), but within weeks this thin layer was eroded away in places. Regrettably, over the ensuing months, the track-bearing block slumped into the sea.
Two parallel, slightly curved (convex to the east) sets of four tracks were evident, ~69 cm apart, and multiple midline features were noted. The site was inferred to exhibit a large tramline trackway with a straddle of 69 cm (Table 1; Fig. 4 is annotated to illustrate these features under different lighting conditions). The direction of travel was from bottom to top as the surface was viewed when facing north, and tracks in the left and right sets were labelled L1–L4 and R1–R4. What we term a ‘crusted substrate phenomenon’ (implying a firm, crusted surface and softer underlying sand) was present. That is, tracks and traces were either shallow and well defined on the firm surface, or else as much as 2.5 cm deep where they had punctured the surface, resulting in poor morphological detail, but with crisp rims (some of which were slightly overhung) (Fig. 5A). The pattern in L1–L4 and R1–R4 was of alternating short and long distances between tracks.

Figure 4. Annotated tracks and traces at the main site. (A) In shade; (B) in low-angle sunlight; scale bar in cm applies to both (A) and (B).

Figure 5. (A) Tracks L1 and L2 viewed obliquely, illustrating crusted surface with crisp track margins and slight overhang; the white scale bar is 10 cm long; the white arrows indicate claw drag impressions. (B) Track R3 showing well-preserved claw-drag impressions; scale bar is in cm. (C) Track R2 showing claw drag impressions; scale bar is in cm. (D) Track L3 showing possible claw drag impressions; scale bar is in cm. (E) Striations registered by the plastron, curving to the left at M1; the large depression made by M2 is indicated by the white arrow; the white scale bar is 10 cm long.
Table 1. Measurements and comments from the main tracksite; L = length; W = width; IT = intertrack length; ID = interdigital length. All readings are in cm.

Many tracks exhibited posterior, posteromedial, or posterolateral displacement rims. The tracks showed variation in length and width (Table 1). In one case, the track (R3) had not punctured the crusted surface and had not penetrated into the underlying, softer sand. Instead, well-preserved, parallel claw-drag impressions were evident (Fig. 5B). The most accurate interdigital distance could be measured from this track, despite the medial claw impression (digit I) being partially occluded by infill.
R2 appeared to exhibit at least four long claw-drag impressions; those of I and II were well preserved. These tapered distally to sharply angled anterior ends (Figs. 4B, 5C). L3 contained three less well-defined impressions (Fig. 5D).
R1 was the longest track (Fig. 4), but on closer inspection, although it was truncated by the edge of the surface posteriorly and laterally, it was noted to be a composite track: a larger proximal depression (~13 cm long) was separated by a shallow ridge from a smaller (~11 cm long) distal depression. The medial margin was more prominent proximally and distally, and hence slightly narrower in between, consistent with a composite track.
Three sets of faint longitudinal features were noted proximal to L1, aligned in the direction of the trackway, and ~2 cm long. The distance between the central set and left set was 4 cm, and between the central set and the right set was 8 cm. Their true length may have been greater, because their anterior ends were truncated by the posterior margin of the displacement rim (Fig. 5A).
In addition, four features (or clusters of features) were identified close to the midline, and were labelled M1, M2, M3, and M4. At the proximal end of the trackway, and truncated by the proximal margin of the surface, was an area 25 cm in length and 13 cm in width (M1), containing as many as 13 parallel, narrowly spaced, slightly curved (convex to the east), shallow striations at regular intervals (Fig. 5E). The alignment and curvature of these striations was similar to that of the trackway. Within weeks of identification, these striations were almost undetectable. M2 consisted of a round depression, ~4 cm in diameter, within the area of M1 striations (Fig. 5E). Proximal to it were three round depressions, ~1 cm in diameter and ~ 3 cm apart from each other, aligned in a linear pattern in the direction of motion (Fig. 4B). M3 comprised a large depression containing as many as 10 striations (ridges and grooves) aligned in the direction of motion (Fig. 4). M4 comprised three depressions (M4a, M4b, and M4c) in a linear pattern, described here from proximal to distal (Fig. 4B). M4a had a symmetrical appearance, a maximum length of 4 cm, and width of 6 cm. M4b and M4c were small, round depressions, ~1.5 cm in diameter.
Other tracksites
We identified other tracksites with features suggesting tortoise (and possibly terrapin) trackmakers of various sizes. Because tortoise tracks are hitherto undescribed in the fossil record, we present six examples. Tracksites were concentrated in three zones (Fig. 1): (1) 7 km west of the main tracksite in the Geelkrans Nature Reserve (five sites, two of which are presented here); (2) 115 km east of the main tracksite, in the Garden Route National Park (three sites, two of which are presented here); and (3) 125 km east of the main tracksite, in the Goukamma Nature Reserve (three sites, two of which are presented here). In all cases tracks were evident in epirelief.
The Geelkrans sites
At the western site, >20 tracks were present on loose aeolianite slabs. Two possible trackways were present, with a probable tramline pattern (straddle ~18 cm), possible ‘crusted substrate phenomenon,’ and a pattern of probable pes-manus pairs. Track length = 5 cm; track width = 4 cm. One trackway is shown in Figure 6A.

Figure 6. (A) A portion of one of the trackways at the western Geelkrans site; scale bar is in cm. (B) 3D photogrammetry image of trackway at the eastern Geelkrans site; horizontal and vertical scales are in meters.
A short distance to the east, a loose slab exhibited tracks with a straddle of ~18 cm, with four closely spaced tracks in one ‘line’ of a tramline, and one track in the other ‘line’ of a tramline. Track length = 3 cm; width = 3 cm (Fig. 6B).
The Garden Route National Park (GRNP) sites
At the western site, a loose slab contained teardrop-shaped depressions (~1.5 cm long and 0.8 cm wide, with faint displacement rims) in groups of three (Fig. 7A). The distance between the depressions was ~2 cm. The depressions were on a surface inferred to be from a lagoonal environment (Helm et al., Reference Helm, Cawthra, Combrink, Helm, Rust, Steer and van den Heever2020a).
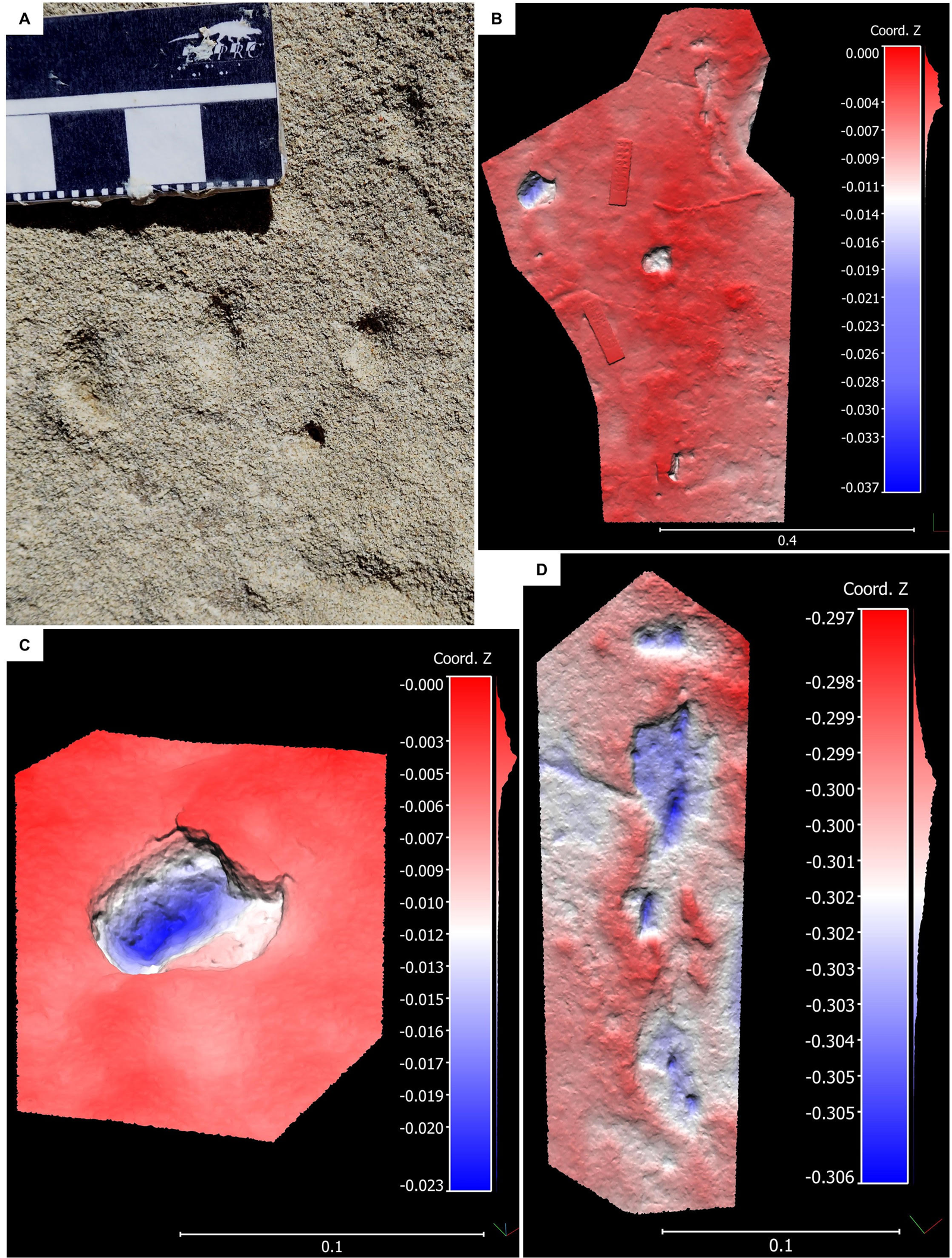
Figure 7. (A) An image of the ‘toe-tip traces’ at the western GRNP site; scale bar in cm and mm. (B) 3D photogrammetry image from the eastern GRNP site, illustrating the crusted substrate phenomenon with missing tracks. (C) 3D photogrammetry tilt view of the left track in (B) illustrating an example of a crust overhang. (D) 3D photogrammetry of the distal track features in (B), see text for interpretation details. Horizontal and vertical scale in the photogrammetry images are in meters.
To the east, a truncation surface on a large fallen block contained several tracks (Fig. 7B), showing the ‘crusted substrate phenomenon,’ with areas where expected tracks were missing. A crust remnant overhung two tracks (Fig. 7C). The largest track (Fig. 7B, 7C) was 6 cm wide; the smallest track (Fig. 7B, 7D) was 4.5 cm wide with forward-pointing digit impressions. Approximately 3 cm ahead of the anterior margin of this track were two small circular depressions (~1.3 cm apart) within a shallow depressed area, and small depressions of a similar size were present ~10 cm behind its posterior margin. These distal features were associated with two narrow, longitudinal, linear depressions (Figure 7D). The surface has been associated with a lagoonal environment (Helm et al., Reference Helm, Cawthra, Combrink, Helm, Rust, Steer and van den Heever2020a).
The Goukamma sites
The Goukamma 1 site comprised a loose slab below low coastal cliffs, which was frequently deeply covered by beach sand. Both Waenhuiskrans Formation and Klein Brak Formation outcrops occur in this area. The slab appeared to have been in this locality for some time, and its surface had been subjected to wind and water erosion. The surface contained linear ripples, one of which was flat-topped. The slab displayed two oval depressed features on its surface, interpreted here as putative tracks (Fig. 8).

Figure 8. (A) Large depressions on the surface at the Goukamma 1 site; scale bar is in cm. (B) Depressions on the surface of the Goukamma 2 site, typically in sets of three; scale bar is in cm.
When viewed facing north, the larger depression was proximal to the smaller, distal depression. The proximal depression measured 33 cm in maximum length and 20 cm in width. Two small oval depressions, ~3 cm long, were situated just ahead of it. The minimum distance between the proximal and distal depressions was 20 cm. The distal depression was smaller, rounder, with maximum length of 22 cm, and width of 15 cm. Just anterior to it and towards the right were three depressions, ~2 cm in diameter and 5 cm apart. The longitudinal axis of the proximal depression was directed more to the left, and the longitudinal axis of the distal depression was directed more to the right.
Situated 400 m east of the Goukamma 1 site, the Goukamma 2 site consisted of a loose slab, a surface of which contained small depressions, ~1–2 cm in length and width. These typically occurred in sets of three (Fig. 8B). The distance between impressions in each set was ~2.5 cm.
OSL dating results
The luminescence dating results are provided in Table 2 (see also supplementary information). The ages span the typical range of aeolianite along this coastline, with the initial age estimates ranging from 139 ± 10 ka to 70 ± 4 ka (MIS 6 to MIS 4). The results of the Rcarb dose rate modeling suggest that including the effects of carbonate cementation (and concomitant pore water loss) in the dose rate calculations tends to reduce the resulting OSL ages, although given the age uncertainties, the various scenarios for timing of cementation do not strongly affect the resulting ages. For Leic20024 and Leic20003, the age differences resulting from use of the dose rate modeling are small, but the difference is more significant for the older samples. Given the likely need for a proximal sediment source to form the extensive MIS 5e coastal dune systems on this coastline, the modeled MIS 5e ages are perhaps more geomorphologically plausible than the MIS 6 ages obtained via conventional dose-rate calculations (130–140 ka). We therefore suspect that the modeled ages for Leic21008, Leic20031, and Leic20027 are more plausible (i.e., more likely MIS 5e or very late MIS 6). Additional OSL ages of 76 ± 5 ka and 73 ± 5 ka have been obtained recently for locales, respectively, 0.1 km and 0.5 km west of the Goukamma 1 and 2 sites (Helm et al., Reference Helm, Carr, Cawthra, De Vynck, Dixon, Stear, Stuart, Stuart and Venter2022).
Table 2. Summarized OSL data, including measured equivalent doses (from the central age model [CAM] weighted mean) and ages obtained with conventional dose rate estimations (using 3 ± 3% water contents). The two right-hand columns provide age estimates obtained using the RCarb model for a reduction in water content and the formation of 20 ± 5% carbonate cement, either rapidly after burial (first 33% of burial time) or for the duration of burial (~90% of burial time). Full details of the equivalent dose measurements and dose rates are provided in the supplementary file.

DISCUSSION
Consideration of trackmaker candidates
Tramline trackways contain left and right lines of tracks that are clearly and widely separate, yet form part of a double-line unit (Stuart and Stuart, Reference Stuart and Stuart2019) (i.e., they exhibit a wide straddle). They may be made by a variety of extant trackmakers: crocodiles, lizards, tortoises, terrapins, sea-turtles, frogs, and many types of invertebrates (Stuart and Stuart, Reference Stuart and Stuart2019). In the examples described herein, the straddle size excludes all but the largest contenders. Furthermore, the presence of forward-pointing claw impressions excludes sea-turtles, which have a different locomotion pattern (Lockley et al., Reference Lockley, Xing and Xu2019b). The suite of possible trackmakers is thus limited to crocodylians, large varanids, large tortoises, and terrapins. First, however, we consider two cases that are not related to tramline trackways.
Bovids
Could the depressions at the main site represent two independent, parallel trackways? In such a case, the longer the trackway exposure, the greater the likelihood of convergence or divergence of the individual trackways, which would exclude a tramline trackway. There is no evidence of such phenomena at the main site, and the straddle remains constant along the trackway length, even as it curves to the left. However, a longer exposure of tracks would allow this to be determined with greater confidence.
This is relevant because R3 might be construed to exhibit bovid track morphology, with two-digit impressions separated by a ridge (representing the interdigital sulcus) and tapering sharply anteriorly. Furthermore, a long bovid trackway on the Goukamma coastline contains not only well-preserved tracks with obvious bovid morphology, but also some tracks that are substantially larger and wider, with indistinct morphology (Helm et al., Reference Helm, McCrea, Cawthra, Thesen and Mwankunda2018b). It could therefore be argued that there are two parallel artiodactyl trackways and that, with the exception of R3, the tracks exhibit poor morphology.
However, the outward curvature of both well-impressed claw impressions is different from the orientation of bovid digit impressions. Moreover, our interpretation is that a third depression is present to the left of the two obvious depressions of R3, but is largely infilled with sediment, although its anterior tip can be seen and measured. Tridactyl morphology excludes a bovid origin. Moreover, the midline features (M1–M4) cannot be readily explained if two independent trackways are postulated, but are consistent with plastron and tail traces in a tramline trackway. For these reasons, we are confident in the interpretation of a tramline trackway.
Proboscideans
At the Goukamma 1 site (Fig. 8A), where only one manus-pes set of tracks is arguably present, it is impossible to determine the presence or absence of a tramline trackway. Overall track size and shape are not inconsistent with proboscidean tracks, and numerous elephant tracksites have been documented along the Cape south coast (Helm et al., Reference Helm, Lockley, Moolman, Cawthra, De Vynck, Dixon, Stear and Thesen2021b). However, the indentations situated ahead of the tracks, inferred to represent digit traces, are not consistent with those of proboscideans.
Crocodylians
Crocodylian trackway morphology has been well documented by Farlow et al. (Reference Farlow, Robinson, Kumagai, Paladino, Falkingham, Elsey and Martin2017) from long trackways of the American crocodile (Crocodylus acutus). Southern African tracking manuals consistently depict tracks of the Nile crocodile (Crocodylus niloticus) (Liebenberg, Reference Liebenberg2000; van den Heever et al., Reference van den Heever, Mhlongo and Benadie2017; Stuart and Stuart, Reference Stuart and Stuart2019). Crocodylian pes tracks are substantially longer than they are wide. Both manus and pes tracks exhibit long digit impressions that taper distally, with claw impressions registered ahead of the digit tips.
In a typical gait pattern, the pes is placed behind the manus track, resulting in a short-long sequence. Typically, the short segment of this sequence appears opposite the long segment in the contralateral line. Long belly-drag impressions may fill most of the space between the lines of the tramline, upon which is often superimposed a sinuous tail drag impression (Stuart and Stuart, Reference Stuart and Stuart2019, p 219). None of these features is present at the main tracksite.
The only track with relative dimensions similar to that of a crocodylian pes is R1, which upon closer inspection is a composite pes-manus track. In particular, the midline features present at the main tracksite are not compatible with a crocodylian trackmaker, hence a crocodylian origin can be excluded.
Varanids
The two largest extant lizards in southern Africa are the water monitor (Varanus niloticus) and rock monitor (Varanus albigularis). There are no historical, archaeological, or paleontological records of either species from the Cape south coast. Differences between varanid tracks and crocodylian tracks include digit shape, digit orientation, and claw position. Monitor lizards may attain a total length of 220 cm (Keates, Reference Keates2016). Van den Heever et al. (Reference van den Heever, Mhlongo and Benadie2017) noted that a 100 cm long monitor lizard would register a manus track 6 cm long and a pes track 8 cm long.
However, a long tail drag impression is usually present (Liebenberg, Reference Liebenberg2000; van den Heever et al., Reference van den Heever, Mhlongo and Benadie2017; Stuart and Stuart, Reference Stuart and Stuart2019), which is inconsistent with the midline features noted at the main site, and is absent from all the other sites. Track dimensions and footfall patterns are not consistent with those of monitors, which can thus be excluded.
Terrapins
Terrapin tracks often just show toe-tip traces (claw impressions) in a characteristic pattern, often in threes (Stuart and Stuart, Reference Stuart and Stuart2019, p. 211, fig. K). Such traces may also occur in shallow sub-aqueous environments as a result of bottom-walking, and if the water is deeper, swim traces may be registered (Lockley et al., Reference Lockley, Cart, Martin, Prunty, Houck, Hups, Lim, Kim, Gierlinski, Lockley and Lucas2014, Reference Lockley, McCrea, Buckley and King2018).
Trackways typically have tramline features, with a wide straddle. Drag marks made by the claws of the hind feet are less common than in tortoise trackways (van den Heever et al., Reference van den Heever, Mhlongo and Benadie2017). When complete terrapin tracks are registered, they may resemble tortoise tracks, but the alignment of the claw impressions differs, being orientated more in the direction of travel in both manus and pes tracks. In a normal walking gait, the pes track is found behind the manus track and does not overlap it, unlike the typical tortoise trackway pattern (van den Heever et al., Reference van den Heever, Mhlongo and Benadie2017).
Although terrapins are freshwater turtles and tortoises are terrestrial turtles, caution is required in basing track identification solely on environment. Tortoises access water at times, and their tracks may therefore occur in moist substrates. The leopard tortoise is fond of water, and often floats in lakes (Branch, Reference Branch2016). Its tracks can thus be found on sub-aqueous surfaces. Terrapins bask on shores, and on occasion can be found on dry substrates a short distance from water or during migration after rains (Branch, Reference Branch2016).
The largest extant terrapin species in southern Africa is the Nile soft-shelled terrapin, Trionyx triunguis, which reaches a maximum length of 120 cm and enters the subcontinent in the northwest (Branch, Reference Branch2016). In addition to having soft shells and often attaining large sizes, members of this family have only three claws on both the manus and pes.
Of the numerous indigenous terrapin species in southern Africa, a species of helmeted terrapin (Pelomedusidae, Pelomedusa galeata) is the only species that occurs currently on the Cape south coast (Petzold et al., Reference Petzold, Vargas-Ramírez, Kehlmaier, Vamberger, Branch, Du Preez and Hofmeyr2014). The maximum reported length of 32.5 cm (Branch, Reference Branch2016) probably refers to this species. Single specimens of undifferentiated Pelomedusidae are known from the archaeological record, from Gamtoos AK1 (Hendey and Singer, Reference Hendey and Singer1965), and Klipdrift Shelter on the Cape south coast (Reynard and Henshilwood, Reference Reynard and Henshilwood2017). Two specimens identified as marsh terrapins were reported from Peers Cave (Peterhans and Singer, Reference Peterhans and Singer2006).
Giantism is not recorded in Pelomedusidae, but is well documented in Podocnemididae (formerly classified within Pelomedusidae): a fossil terrapin from Venezuela exceeded 2.5 m in length (Branch, Reference Branch1994). Representatives of Podocnemididae are reported from the Miocene and Pliocene of central Africa: the last known continental African representative was Turkanemys pattersoni from Lothagam in Kenya (Wood, Reference Wood, Leakey and Harris2003). These large terrapin species were highly aquatic. Despite the above-mentioned caveat that terrapins may be found a short distance from water, the possibility of the tracks at the main site being made by a giant terrapin on a dryland dune surface seems remote. A tortoise origin appears far more likely. The same cannot be said of the putative Goukamma 1 tracks. If conditions were favorable on the Cape south coast during MIS 5e for crocodiles (Helm et al., Reference Helm, Cawthra, Combrink, Helm, Rust, Steer and van den Heever2020a; Helm and Lockley, Reference Helm and Lockley2021) and breeding sea-turtles (Lockley et al., Reference Lockley, Cawthra, De Vynck, Helm, McCrea and Nel2019a), it is conceivable that giant terrapins were present. If the Goukamma 1 putative tracks were made by a chelonian, then the suggestion of three inward-facing claw impressions favors a giant tortoise, but the distance between putative pes and manus tracks is more consistent with a terrapin.
In summary, we confidently attribute the main trackway east of Still Bay to a giant tortoise trackmaker, but take a more cautious approach with the Goukamma 1 site, where the tracks remain putative, and both giant tortoise and giant terrapin trackmakers are plausible. As discussed below, terrapins and tortoises are plausible trackmakers at the western GRNP site and Goukamma 2 site, while the features at the eastern GRNP site suggest a terrapin trackmaker.
Interpretation
Main site, east of Still Bay
A substantial cliff-collapse event must have occurred to account for the presence of multiple large loose blocks slumping down an unstable slope (Fig. 2). This probably occurred not long before the track-bearing surface was identified. The brief preservation of the surface veneer allowed for interpretation of ichnological details, which subsequently became undetectable. Consequently, although track morphological detail was sometimes lacking where the crusted surface had been penetrated, preservation of the surface features was of high quality, and included crisp track margins, crust overhang features, and preservation of claw-drag features where the crusted surface had not been penetrated (Figs. 4, 5).
The inference of a large tramline trackway was bolstered by the parallel nature of the lines of the tramline, both curving slightly to the left. R1 and L1 were both considerably longer than wide, and a composite track (proximal pes track, distal manus track) was inferred in R1. Although this level of detail was not discernible in L1, it was also probably a composite track. The footfall pattern could not be determined: possibly the shorter inter-track distance of L2-L3 was associated with pes-manus pairs, but possibly a direct register led to pes tracks being superimposed on manus tracks in L2 and L3. L4 was essentially uninterpretable, and R2 and R3, while showing claw-impression detail, were not identifiable to manus/pes level.
The digit/claw impressions evident in R3 and R2 indicated a trackmaker of substantial size, as did the straddle of 69 cm. It was inferred that the trackmaker in these areas was scraping its claws on the crusted surface, which it did not puncture, resulting in shallow linear impressions.
The short striations proximal to L1 were interpreted as claw-drag impressions that probably were made during protraction, as is commonly evident in tortoise trackways. The distances between these traces suggested that not all claws registered drag marks, and an inter-claw distance of ~4 cm.
The midline features and clusters were of diagnostic significance. M1, containing longitudinally orientated, parallel striations that followed the curve of the tramline trackway, clearly represented the dragging of a midline feature, interpreted as a plastron drag that did not puncture the surface crust.
The single larger circular depression at M2 was registered after passage of the plastron, with M1 striations evident proximal and distal to it. We interpreted this as a probable deep tortoise tail impression, where the tail speared and penetrated the crust. The three smaller circular depressions proximal to it (in a linear pattern in the orientation of the trackway) were probably associated with the trackway. The most parsimonious interpretation is that they represent tail-tip impressions. The possibility of randomly situated erosional features cannot be excluded, but the rarity of such features on the surface, other than close to the midline, makes this unlikely.
The large, midline M3 depression, with striations in the form of grooves and ridges, was interpreted as a plastron trace. However, here the plastron punctured the crust and gouged a deep depression. M1 and M3 thus presented plastron traces that differed widely in morphology, reflecting whether the substrate was scraped (M1) or punctured (M3). M4a was interpreted as a small scuff mark, probably registered by the plastron. M4b and M4c, situated close to the midline, were interpreted as tail-tip traces. Displacement rims resulted from forward propulsion of the manus and pes, pushing sediment posteriorly, or representing the gravitational effect of upslope travel.
The overall impression is of a trackmaker that was struggling on a crusted substrate that had a variable capacity to support its weight. In addition to the expected features of a tortoise trackway (wide straddle, closely spaced tracks), this led to the registration of claw drag marks, plastron traces, and tail-tip traces.
Other tracksites
The combination of findings at the Geelkrans sites (Fig. 6) suggested tortoise trackmakers, which are more probable on aeolianite surfaces than terrapins. The western GRNP site (Fig. 7A) exhibited impressions in sets of three, interpreted as ‘toe-tip traces’ made by chelonian claws. This pattern occurs more commonly with terrapin traces than tortoise traces.
The eastern GRNP site showed the ‘crusted-substrate phenomenon,’ with ‘missing tracks,’ and a crust overhang visible in two tracks. It is not possible to infer the presence of a tramline trackway with confidence (Fig. 7B), although the track on the left might be part of the left line and the remaining tracks might be part of the right line. Because the left track is slightly larger than the others, it is possible that more than one trackmaker was present. The orientation of claw impressions in the right line is in the orientation of the tracks, suggesting a terrapin trackmaker, but the longitudinal grooves, interpreted as claw drag marks, are more consistent with a tortoise. The claw traces in the left track are directed slightly outwards. Distally, the probable pattern is of a pes track with manus claw impressions lying 3 cm ahead of it. The distance between these features is more consistent with a terrapin trackmaker. A terrapin appears to be a more probable trackmaker candidate.
The surface of the Goukamma 1 slab (Fig. 8A) was too small to determine whether the ripple marks were formed by wind or sub-aqueously. The distal depression, with three indentations anterior to it, appropriately positioned for claws I, II, and III, resembled a chelonian manus track. The ratio of the putative track dimensions to the claw impression size and inter-claw distance matched those of tortoise tracks (van den Heever et al., Reference van den Heever, Mhlongo and Benadie2017). Examination of the scale bar, however, implies a truly gigantic trackmaker. The proximal oval depression and the orientation of its long axis were consistent with the form of a chelonian pes track, slightly larger than anticipated. The small depressions ahead of it were appropriately positioned for claw impressions. A left pes-manus pair was suggested, with manus claw impressions directed slightly inwards. This would explain the absence of the right line of a tramline, which would be situated beyond the margin of the surface. The distance between the two putative tracks, which was large for a tortoise, is more consistent with a terrapin trackway pattern. However, leopard tortoises enter bodies of water (Branch, Reference Branch2016) and their footfall patterns on sub-aqueous surfaces might be different from those on dryland substrates.
These speculations are tempered by the advice of Sarjeant (Reference Sarjeant, Gillette and Lockley1989, p. 369), whose “Ten Paleoichnological Commandments” remain fundamental to ichnology. The relevant ‘commandment’ reads: “a trackway—a series of successive footprints of both, or of all four, feet—is the best possible basis for the definition of a footprint ichnospecies. A set of prints—impressions of all four feet, or at least of a manus and a pes—is the next best basis.” The problem is that although a putative manus-pes pair is present, the manus track, with its probable claw impressions, is more convincing than the more amorphous pes track. Length estimates for the putative trackmaker are presented below. We take a cautious approach, acknowledging possibilities but avoiding firm conclusions.
The Goukamma 2 site (Fig. 8B) resembles the western GRNP site in exhibiting probable chelonian toe-tip traces. Again, these are commonly registered by terrapins, but are plausible for both terrapins and tortoises.
Estimation of trackmaker length
Three ratios were determined (Table 3), and used to estimate trackmaker length: (1) the mean (n = 11) total body length/trackway straddle is 1.53, (2) the total body length/manus track length (n = 1) is 11.8 (total length 71 cm, manus track length 6.0 cm), and (3) the mean (n = 8) total body length/distance between manus claw impressions is 29.8.
Table 3. Data from measurement of leopard tortoises and their tracks; DBMCI = distance between manus claw impressions. All length readings are in cm.

Using the straddle of 69 cm measured at the main tracksite, and the total length/straddle value of 1.53, a total trackmaker length estimate of 106 cm is obtained. If ratios for the individual tortoises assessed are used, an estimated length range of 95–119 cm is obtained. These estimates assume that the relationship between total length and straddle is isometric, but this cannot be known with certainty given the absence of massive present-day leopard tortoises.
Our ratio for total length/distance between manus claw impressions is 29.8. Using the inter-claw distance of 5 cm measured at the main tracksite (although a manus origin for the claw impressions is not certain), a total length estimate of 149 cm is obtained. Because the inter-claw distance may vary, and because the measured distance might relate to either manus or pes, we regard the estimate derived from the straddle as more reliable. The inferred inter-claw distance of 4 cm derived from the traces proximal to L1 (which probably represent pes claw drag impressions) yields a length estimate of 119 cm.
Our ratio for total length/manus track length is 11.8. Based on this, the trackmaker length estimate for the western Geelkrans site is 50–60 cm, and for the eastern site is 30–40 cm. The estimates for the western and eastern GRNP sites and the Goukamma 2 site traces are based on inter-claw distances of 2 cm, 1.3 cm, and 2.5 cm, respectively. These yield length estimates of 57 cm, 39 cm, and 71 cm. While these are based on tortoise track measurements, the tracks may have been made by terrapins.
The Goukamma 1 site yields prodigious length estimates. Because a chelonian origin cannot be conclusively inferred, these estimates require caution. Measurements of the putative tracks include manus track length and inter-claw distance. Based on manus size of 22 cm and the ratio of 11.8, a length estimate of 260 cm is obtained. Based on the putative inter-claw distance of 10 cm, a length estimate of 298 cm is obtained. An alternative involves noting that the inter-claw distance at the Goukamma 1 site is double that recorded at the main site, where a total length estimate based on a straddle of 109 cm was obtained. Assuming that such measurements are scalable, doubling this yields a length estimate of 218 cm. While these estimates suggest a truly giant trackmaker, they are consistent with the largest recorded tortoises or terrapins.
‘Record’ reported total body lengths for the leopard tortoise represent exceptional cases from large sample sizes. It is likely that the tracks at the main tracksite were made by a tortoise from somewhere within the regional size range at the time. A Pleistocene tortoise of exceptional size, and its tracks, could have been substantially larger.
Large leopard tortoises or extinct species?
Was the tortoise trackmaker at the main site a large chronosubspecies of the leopard tortoise, or could it represent an extinct species, unknown in the body fossil record? For some southern African Pleistocene mammals and birds, body size was larger than it is today. Klein (Reference Klein1986) reported that mammalian carnivore size varied significantly during the Pleistocene, being larger during glacial phases. Tyrberg (Reference Tyrberg2007) reported that continental Late Pleistocene bird species were often larger than their extant descendants. This is borne out by evidence from the Cape south coast of larger-than-expected Pleistocene avian tracks (Helm et al., Reference Helm, Lockley, Cawthra, De Vynck, Helm and Thesen2020b). Furthermore, Slavenko et al. (Reference Slavenko, Tallowin, Itescu, Raia and Meiri2016) noted with regard to late Quaternary reptile extinctions that extinct reptiles, including “turtles,” were larger than their extant counterparts.
The estimated size of the trackmaker at the main site is ~50% larger than extant leopard tortoises. Maximum size of extant leopard tortoises varies geographically, being largest at the northern (Sudan) and southern (southeastern Cape) ends of the species’ range: Fritz et al. (Reference Fritz, Daniels, Hofmeyr, González, Barrio-Amorós, Široký, Hundsdörfer and Stuckas2010) reported a maximum carapace size in male leopard tortoises in Tanzania that was <50% of that of large male tortoises of this species in South Africa. This difference exceeds the estimated 50% size difference between the trackmaker at the main site and the largest extant leopard tortoises, which arguably supports the notion of a giant Cape south coast leopard tortoise trackmaker.
If the putative tracks at the Goukamma 1 site indeed have a chelonian origin (giant tortoise or giant terrapin), the prodigious estimated size makes it more likely that they were made by an extinct species. Even the total body length estimate of 39 cm for the trackmaker at the eastern GRNP site, which may have been a terrapin, is larger than the maximum reported length of 32.5 cm for the only terrapin currently found in the southern Cape, Pelomedusa galeata (Branch, Reference Branch2016).
The relative absence of large tortoises in the archaeological record
Tortoise remains were prevalent at many southern Cape Middle Stone Age (MSA) archaeological sites (Reynard and Henshilwood, Reference Reynard and Henshilwood2017). Those from MSA sites are generally larger than those in Later Stone Age (LSA) sites, which is attributed to MSA human populations being less dense, with less intensive predation on tortoises (Steele and Klein, Reference Steele and Klein2013). Nonetheless, the majority of these tortoises were of relatively small size.
Many studies at these sites were performed without regard to detailed tortoise taxonomy. For example, tortoise remains were fairly common at Boomplaas Cave, ~75 km inland from the modern coastline, but tortoise remains were not identified to species level (Faith, Reference Faith2011). Likewise, at the Gamtoos (AK1) site in the eastern Cape, an undetermined genus of tortoise was reported (Hendey and Singer, Reference Hendey and Singer1965).
Thompson and Henshilwood (Reference Thompson and Henshilwood2014), analyzing MIS 5a tortoise remains at the Blombos Cave (30 km west of the main tracksite), found that tortoises were the dominant taxon, with an almost exclusive occurrence of the angulate tortoise (Chersina angulata): 3190 specimens of this species were identified, with just one from the leopard tortoise. The angulate tortoise is a medium-sized tortoise, with a maximum body length of 35 cm (Hofmeyr, Reference Hofmeyr, Rhodin, Pritchard, van Dijk, Saumure, Buhlmann, Iverson and Mittermeier2009).
Although considerable numbers of tortoises have been reported for other archaeological sites in the western Cape (Klein and Cruz-Uribe, Reference Klein, Cruz-Uribe, Parkington and Hall1987, Reference Klein and Cruz-Uribe2000; Klein et al., Reference Klein, Avery, Cruz-Uribe, Halkett, Hart, Milo and Volman1999, Reference Klein, Avery, Cruz-Uribe, Halkett, Parkington, Steele, Volman and Yates2004; Steele and Klein, Reference Steele and Klein2013) or historic sites (Cruz-Uribe and Schrire, Reference Cruz-Uribe and Schrire1991), these exclusively relate to the angulate tortoise. Tortoises were commonly encountered in MIS 4 faunal remains from Klipdrift Shelter, 80 km west of the main tracksite, and the presence of angulate tortoises was noted. At De Hangen in the Cederberg, in a minimum number of individuals of >300, angulate tortoises dominated, although other species could not be excluded (Parkington and Poggenpoel, Reference Parkington and Poggenpoel1971). Klein et al. (Reference Klein, Avery, Cruz-Uribe, Halkett, Parkington, Steele, Volman and Yates2004, p. 5710) noted that the angulate tortoise was “by far the most common species in the historic environment,” and that it “dominates all other regional MSA and LSA samples.” Branch (Reference Branch1994) noted that the leopard tortoise was historically absent from the south-western Cape.
Two exceptions to the archaeological domination of angulate tortoises have been identified, but they represent a smaller species. Peterhans and Singer (Reference Peterhans and Singer2006) identified a single parrot-beaked tortoise (Homopus areolatus) specimen on the Cape Peninsula, along with 11 specimens of the angulate tortoise and two of the marsh terrapin. At Pinnacle Point Cave 13B, 50 km east of the main tracksite, tortoise remains were found in layers ranging in age from MIS 6 to MIS 5b (Thompson, Reference Thompson2010). Both angulate tortoises and parrot-beaked tortoises were identified, the former dominating. Homopus areolatus has a maximum length of 16 cm (Branch, Reference Branch2016). Thompson (Reference Thompson2010) drew attention to the presence of the leopard tortoise in the region today, but identified no remains of this species at Pinnacle Point Cave 13B.
There is thus a mismatch between the tortoises encountered in the body fossil record obtained via archaeological sites, and the ichnosites attributed to large tortoise trackmakers herein. In addition to the findings at the main tracksite and Goukamma 1 site, those from the other sites are in the 30–70 cm range, which is larger than the tracks of angulate tortoises and terrapins that occur in the region today and more consistent with those of leopard tortoises.
A bias is acknowledged in the ichnology record, whereby larger, heavier trackmakers made larger, deeper tracks, which are more likely to be recognized and interpreted. If, as seems plausible, tortoises preferred walking on firm, crusted surfaces, it is possible that smaller, lighter species seldom punctured the crust, and their tracks are thus absent from regional paleosurfaces. Nevertheless, the near-total absence of leopard tortoise remains from the Cape south coast warrants explanation.
The single example of leopard tortoise remains at Blombos Cave (Thompson and Henshilwood, Reference Thompson and Henshilwood2014) indicates that the species was present in the southern Cape during the Late Pleistocene. One explanation is that the rarity of large tortoise remains reflects their extreme regional rarity. Another is that they were present, but seldom preyed upon by humans. This seems unlikely, given the penchant for humans to butcher large tortoises for their meat, contributing to their extinction at diverse global locations. The wide range of ages assigned to the MSA and LSA archaeological sites refutes the possibility that different periods are represented by the trace fossil and body fossil records. Perhaps the shells of giant tortoises are less readily preserved, fragment more easily, and are thus not recognized, although their bones should be preserved, as well as the bones of smaller tortoises.
A final possibility, which in our view is the most likely, is that large tortoises were heavy to carry, and that they were instead butchered where they were found. This would be a manifestation of the ‘schlepp effect,’ whereby the likelihood of a carcass not being transported is related to size and distance from camp, leading to the under-representation of larger animals in the archaeological record (Perkins and Daly, Reference Perkins and Daly1968). This is consistent with predictions derived from ‘Optimal Foraging Theory,’ whereby living organisms opt for the best cost-to-benefit ratio, and the energy expended during transportation would ‘eat’ into the total calories available from the prey (Hawkes et al., Reference Hawkes, Hill and O'Connell1982). In the southern African context, Parkington and Poggenpoel (Reference Parkington and Poggenpoel1971, p. 21) noted: “small animals tend to be brought to the home base whole and will thus contribute bones to the kitchen refuse, whereas larger animals tend to be cut up on the spot and may never be traced at the home base.” Thackeray (Reference Thackeray1979) suggested that large animals were butchered and prepared before returning to camp, resulting in the absence of their bones in the archaeological record. In a possible analogy, a giraffe (Giraffa camelopardalis) tracksite was reported 500 m to the east of the main tracksite, but no giraffe bones have been recorded from the region. It was suggested that giraffe either were not hunted or scavenged, or that their bony remains were not regularly transported to the shelters that form the majority of archaeological and paleontological sites (Helm et al., Reference Helm, Cawthra, Cowling, De Vynck, Marean, McCrea and Rust2018a).
Ethnography provides another means to consider these possibilities. Marshall (Reference Marshall1999) spent years with the San in the Kalahari Desert in the 1950s. Her reporting documented, inter alia, their relationship with tortoises, including the use of tortoise shells as containers by healers. The shells of young tortoises of three species were used for this purpose, including Stigmochelys pardalis babcocki (a subspecies of the leopard tortoise). Tortoises were regarded as “smaller game.” Branch (Reference Branch2016) commented that the San used larger tortoise shells as vessels, and that there was a taboo on eating tortoise meat except by the very young and old, to prevent exploitation of an easily collected food source. Two additional studies of San groups in the Kalahari shed light on the role of tortoises in the diet. Silberbauer (Reference Silberbauer and Bicchieri1972) reported that one band killed an estimated 440 tortoises in one year for food, but did not specify the species. Lee (Reference Lee and Bicchieri1972) noted that the leopard tortoise, easily collected and “weighing up to 8 pounds,” was a great favorite and could feed a family of four. Rock art also demonstrates a relationship between hominins and tortoises, but does not allow for species identification or size estimates. No examples have been attributed to MSA artists.
The role of substrate and crust
A ‘crusted substrate phenomenon’ was noted at a number of sites, implying firm sedimentary veneers or crusted surfaces that overlay softer sediments. On such surfaces, no readily discernible track may be registered, and it is possible that faint trackways go unrecognized in the absence of ideal, low-angle lighting. This may be particularly true of small, light-weight animals such as rodents, birds, and invertebrates. The main tracksite provides an example at M1 of the kind of detail that may be preserved.
However, as is evident at this site, where the crusted surfaces is neither thick nor firm, the trackmaker may break through the crust and register a deep, amorphous track; evidence of the crust may be preserved as a distinctive ‘overhang’ at the track margin. The result is a trackway that seems to be missing individual tracks (e.g., the eastern Geelkrans site and eastern GRNP site.) Quadrupedal reptiles with a wide straddle, such as tortoises, might be adept at using crusted surfaces to facilitate locomotion, because their weight is distributed over a relatively large area, rendering them less likely to penetrate the crust. The ‘crusted substrate phenomenon’ is readily observed in snow (examples are presented in Fig. 9).

Figure 9. (A) bounding gait pattern (direction towards the viewer) of a hare in snow on a crusted surface; manus tracks did not penetrate the crust, whereas the pes tracks, despite being larger, exerted more pressure and created deep cavities that have coalesced. (B) Galloping gait pattern (direction away from the viewer) of a medium-sized canid. The first two sets of four tracks were made on a firm, crusted surface; in the third set, the manus tracks did not penetrate the crust but the pes tracks did, and in the following sets all the tracks penetrated the crust.
The chemical nature of these crusts on the Cape south coast has not yet been established, and may be a subject for further research. They seem related to an initially moistened surface (e.g., following a rain shower), which has partially dried out. A crystalline salt layer that might further bind the crust (and potentially create a veneer that enhances preservation of track detail) is a possibility. The development of a microbial or biofilm layer may enhance this process, as suggested by Seilacher (Reference Seilacher2008), who used the term ‘bioglue.’ Helm et al. (Reference Helm, Anderson, Buckley, Cawthra and De Vynck2017) discussed contemporary biofilm growth on Pleistocene aeolianite exposures on the same coastline.
Dating and paleoenvironment
The OSL ages from west and east of the main site (Leic21031, Leic20024; 131 ± 10 ka and 104 ± 8 ka, respectively) place the site within a range from late MIS 6 to MIS 5c, which is within the range established for the region by Roberts et al. (Reference Roberts, Bateman, Murray-Wallace, Carr and Holmes2008) of 140 ± 8 ka to 91 ± 5 ka. The OSL ages from east of the Geelkrans sites of 126 ± 9 ka (Leic21008) and from near the GRNP sites of 125 ± 7 ka (Leic20027) are both most likely from MIS 5e. Results from near the Goukamma 1 and 2 sites are clearly younger (from MIS 5a or MIS 4). The temporal range thus established for these ichnosites spans a variety of conditions and global temperatures—from the MIS 6 glacial (when the coastline would have been kilometers to the south) through the MIS 5e interglacial, when sea levels reached 6–8 meters above current levels (Carr et al., Reference Carr, Bateman, Roberts, Murray-Wallace, Jacobs and Holmes2010), through MIS 5a to the MIS 4 glacial phase.
Vlachos and Rabi (Reference Vlachos and Rabi2018) found no obvious correlation between body size and climate, but acknowledged that larger sample sizes might be required. It was speculated that the effect might be variable: cooler temperatures might shorten periods of annual growth and lead to smaller size, but might lead to increased size though gigantothermy (Paladino et al., Reference Paladino, O'Connor and Spotila1990), in which body temperature is maintained through large size, low metabolic rate, and the insulating effect of peripheral tissues. Angielczyk et al. (Reference Angielczyk, Burroughs and Feldman2015) demonstrated that extant tortoises do not follow Bergmann's Rule (whereby organisms at higher latitudes are expected to be larger than those at lower latitudes, in order to conserve heat). It therefore seems prudent to refrain from confident inferences on Pleistocene paleoenvironment from the perspective of very large tortoises or terrapins.
CONCLUSIONS
The addition of tortoise tracks and traces fills a gap in the global ichnology record. Ichnological evidence of large tortoises on the Cape south coast was not anticipated. The interception of the main track-bearing block during its journey between cliff-collapse and the sea was fortuitous. While an inferred tortoise trackmaker length of more than a meter at this site does not place it in the length range of truly giant tortoises (as described in the body fossil record and currently from islands), it is considerably larger than the largest extant southern Africa tortoises. This implies either a large chronosubspecies of the leopard tortoise or an extinct species. In contrast, the size of the putative chelonian Goukamma 1 trackmaker would indeed make it a giant, but the evidence is insufficient to reach this conclusion.
The inferred age range of the tracksites described suggests the presence of tortoises (and possibly terrapins) in the region over a period of ca. 65 kyr, spanning MIS 6 through MIS 4. This is not surprising, given the prevalence of so many extant tortoise species in southern Africa. The relative absence of large tortoises from the body fossil record and archaeological record is intriguing. The schlepp effect provides the likeliest explanation for the mismatch between these records and the ichnological record.
Tortoise tracks and traces exhibit many forms, with features that vary substantially depending on the nature of the substrate. The variety of ichnological features associated with crusted surfaces serves as a reminder of the need for caution in trackway interpretation. A diligent search for additional chelonian tracks and traces on the Cape south coast and elsewhere is indicated.
Acknowledgments
We thank Gill Simpson, Brenda Piek, Richard Nhyanhi, Karla Grobler, and Charlize du Preez (Wild Rescue NPC), and Michaela White and Wade Whitehead (FreeMe Wildlife) for their assistance with tortoise biometrics. We appreciate support from SANParks, CapeNature, Given Banda, Renee Rust, Francis Thackeray, and Peter Todd. Thank you to Editor Derek Booth and Associate Editor Tyler Faith for their insightful suggestions. Reviewer Hendrik Klein and an anonymous reviewer provided thorough, greatly appreciated ideas for improvement of the original manuscript.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/qua.2022.50















 Open access
Open access