




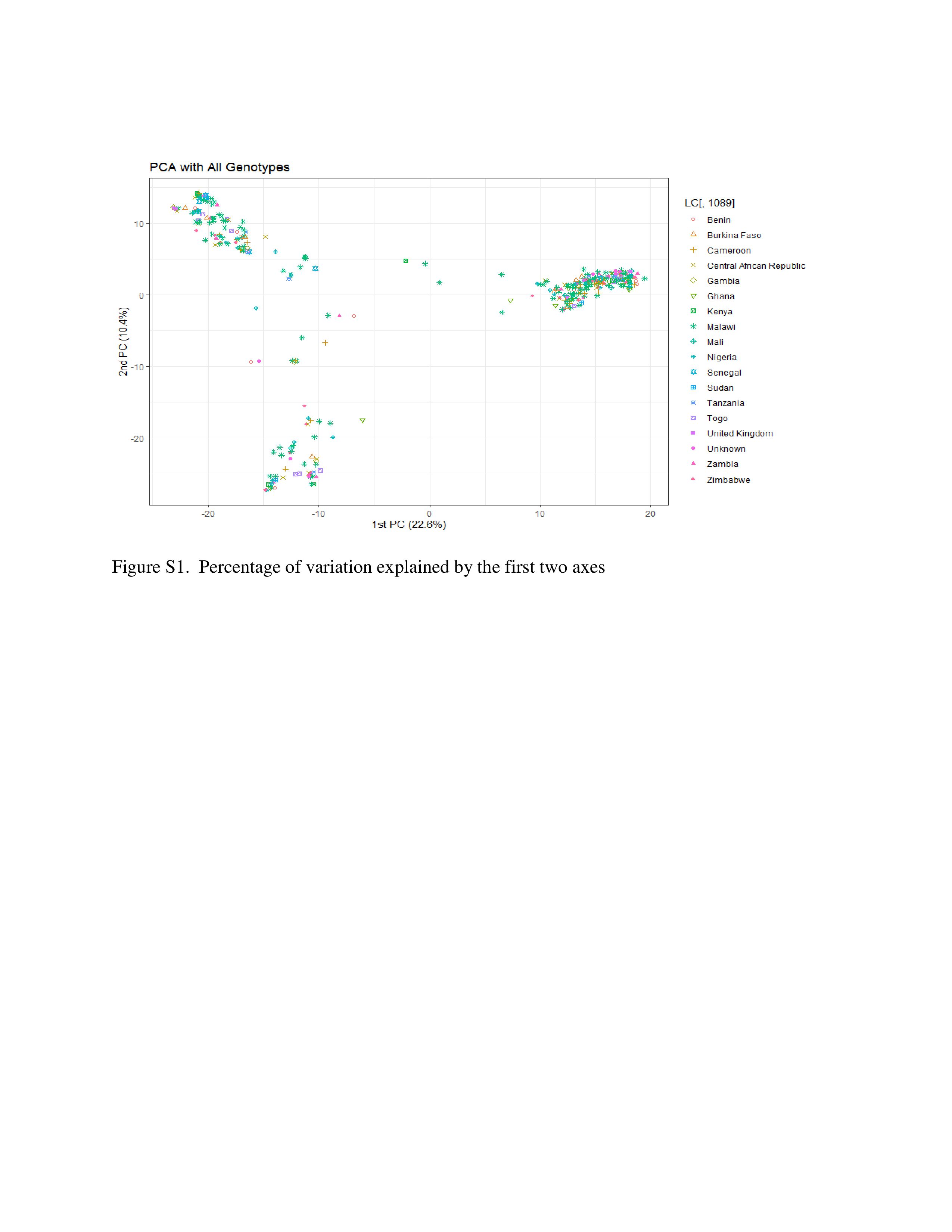
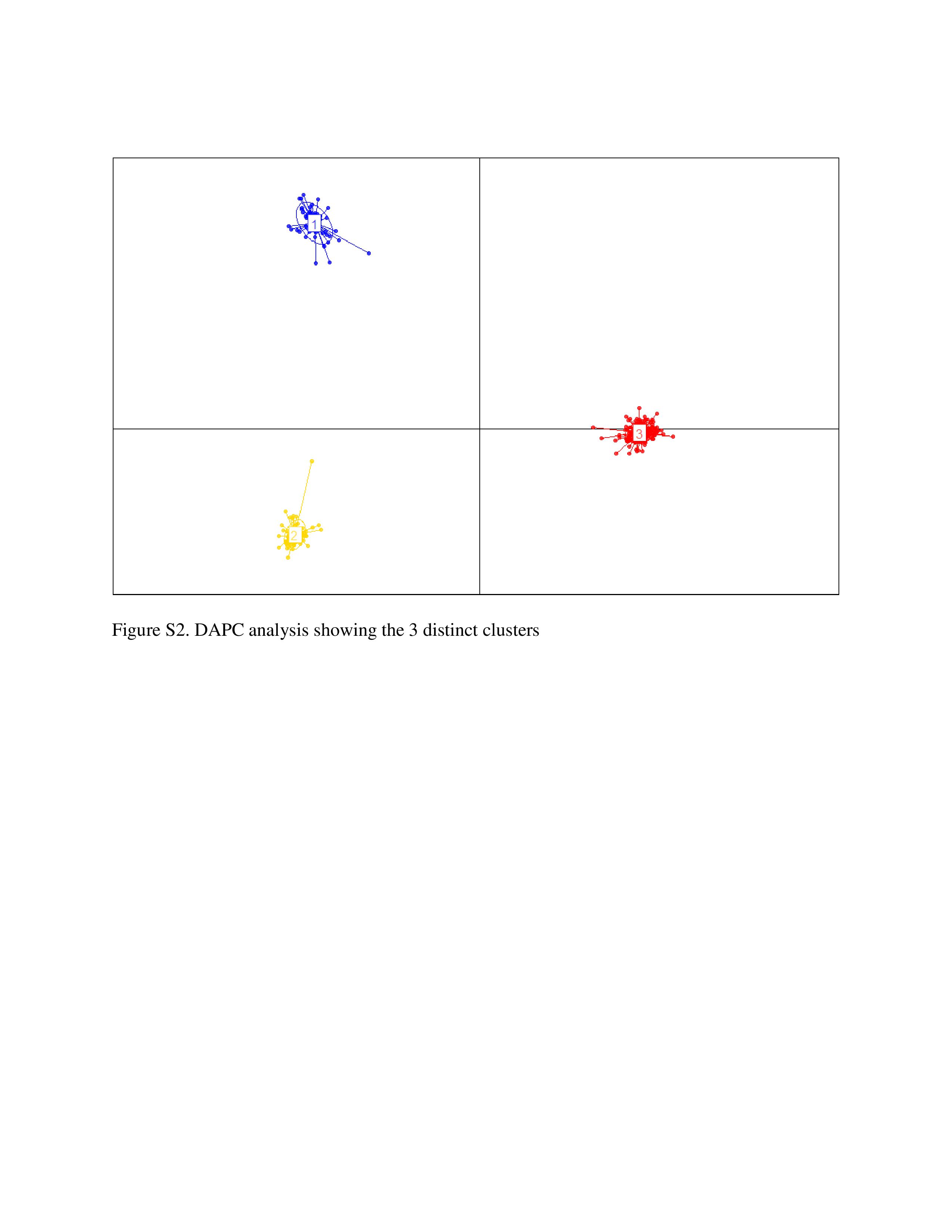


Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
El Bilali, Hamid
Kiebre, Zakaria
Nanema, Romaric Kiswendsida
Dan Guimbo, Iro
Rokka, Veli-Matti
Gonnella, Maria
Tietiambou, Sheirita Reine Fanta
Dambo, Lawali
Nanema, Jacques
Grazioli, Francesca
and
Acasto, Filippo
2024.
Mapping Research on Bambara Groundnut (Vigna subterranea (L.) Verdc.) in Africa: Bibliometric, Geographical, and Topical Perspectives.
Agriculture,
Vol. 14,
Issue. 9,
p.
1541.