



No CrossRef data available.
Article contents
Evaluation of indigenous and exotic soybean accessions for yield, resistance to frog-eye leaf spot and yellow mosaic virus diseases
Published online by Cambridge University Press: 15 December 2023
Abstract
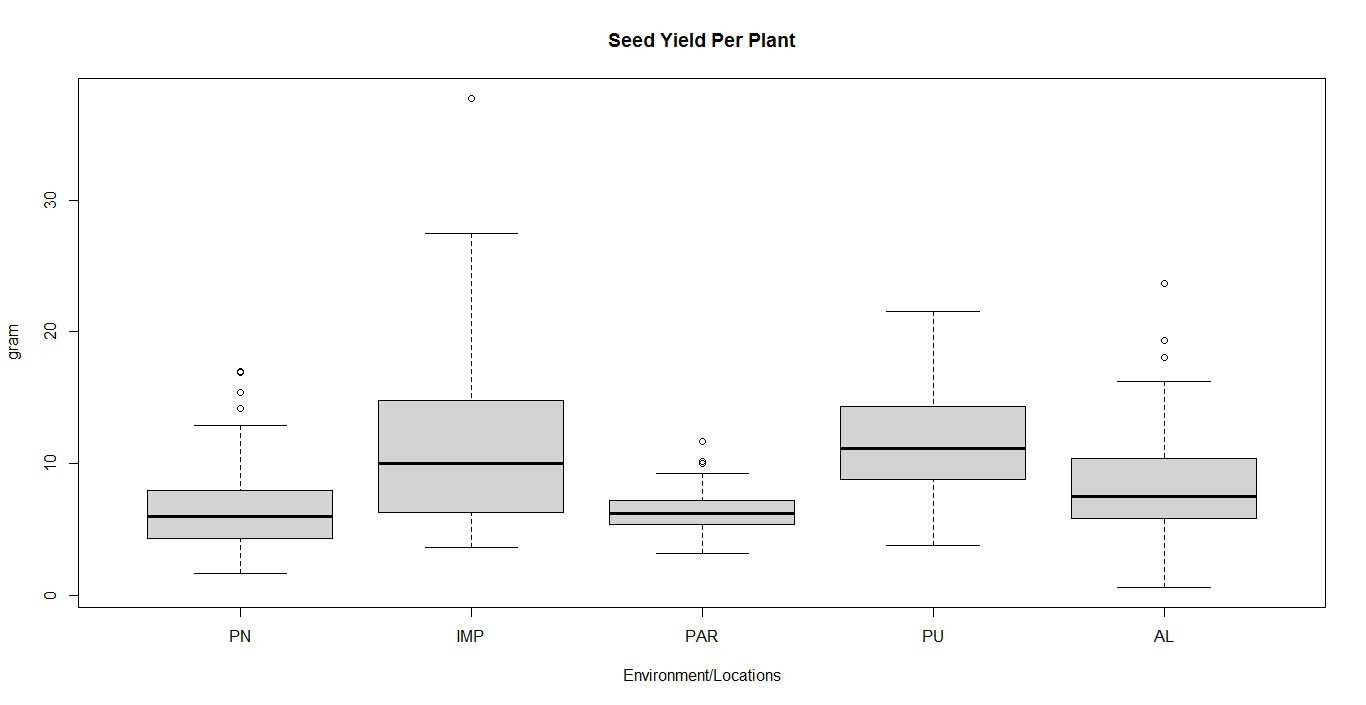
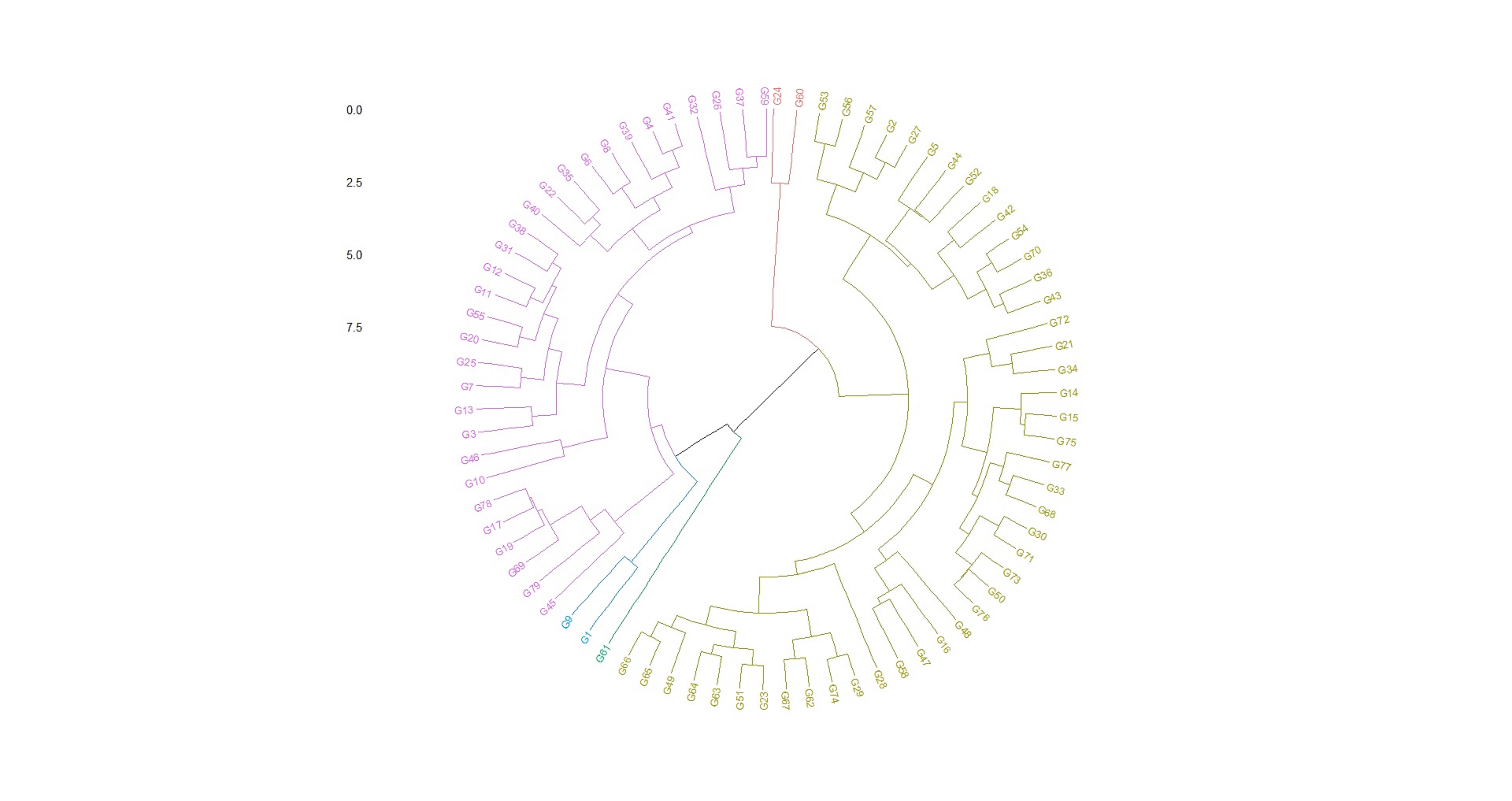
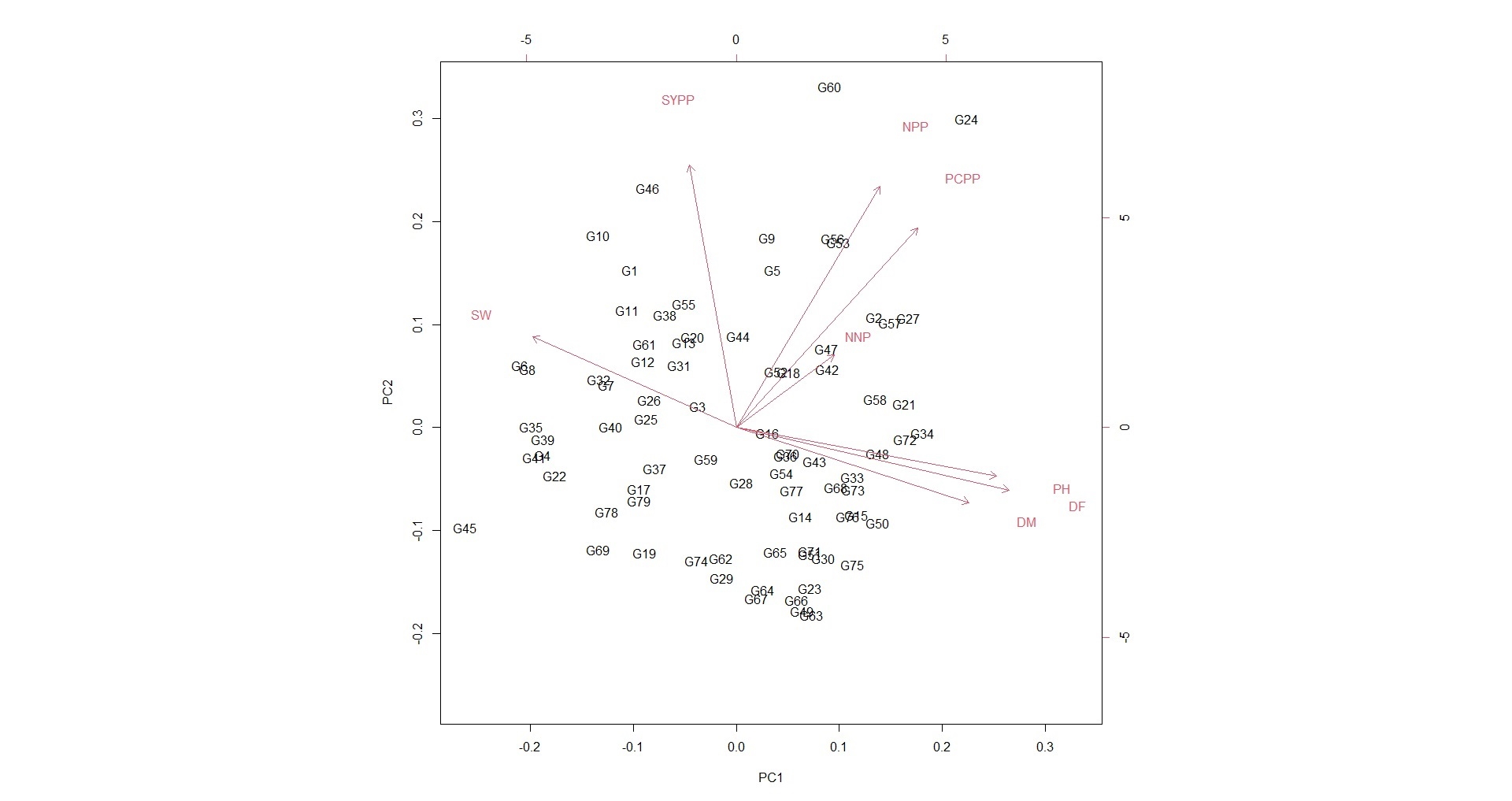
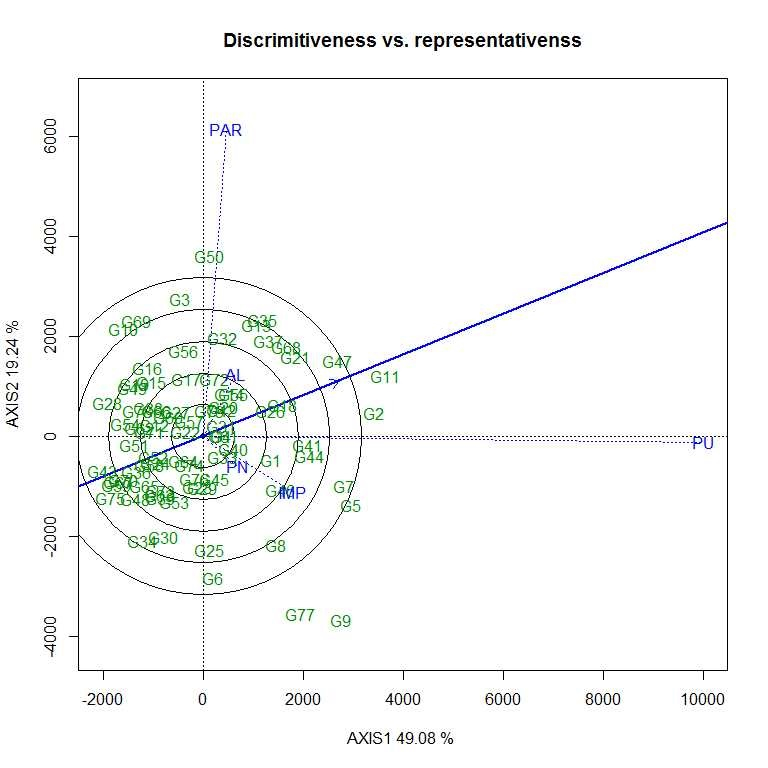
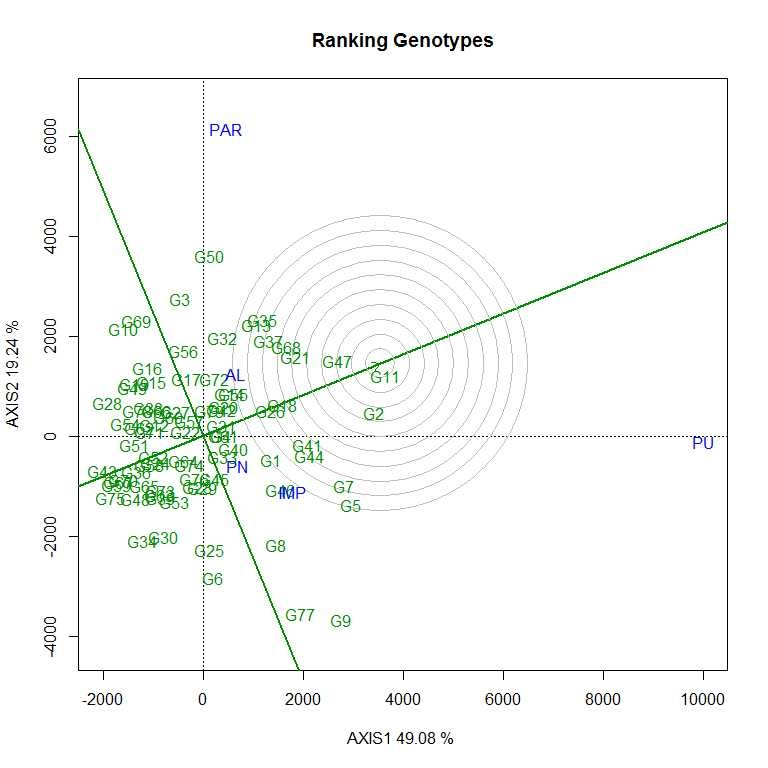
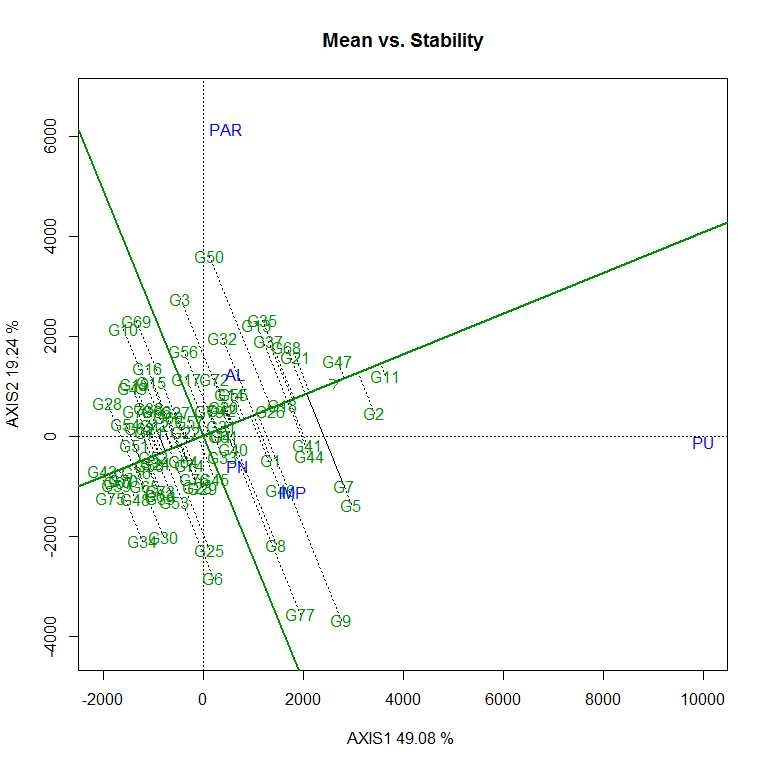
Soybean is a major source of vegetable oil and protein worldwide. Globally, India is among the top five producers where soybean is a major oilseed grown under diverse agro-climatic conditions by small and marginal farmers. The present study aims to identify soybean varieties with higher yield levels, resistance to pestdiseases and adaptability to climatic fluctuations. One hundred and twenty-five (125) indigenous and exotic soybean germplasm accessions and five checks were evaluated and characterized for eight agro-morphological traits at five testing locations and also screened for frog-eye leaf spot (FLS) and yellow mosaic virus (YMV) diseases under hot-spot locations during the rainy season. A wide range of variability was observed among accessions for days to 50% flowering (39–59), plant height (41–111 cm), number of nodes/plant (10–30), pod clusters/plant (14–39), number of pods/plant (40–102), days to maturity (96–115), grain yield/plant (4.89–16.54 g) and 100-seed weight (6.02–13.72 g). Among various traits, 100-seed weight (0.45), number of pods/plant (0.60) and number of pod clusters/plant (0.38) were found to be major yield-contributing traits as they exhibited highly significant correlation with grain yield/plant. Principal components PCI and PCII with eigen value >1 accounted for 42.66 and 27.08% of the total variation, respectively. Accessions G24 (EC 393222) from Taiwan and G40 (IMP-1) from the USA belonging to cluster IV were found promising for multiple yield traits and JS 20–38 from cluster III for earliness as per cluster analysis. GGE biplot average environment coordination (AEC) view revealed that the accessions viz., G11 (EC 333872), G2 (EC 251506) and G47 (TNAU-S-55) were the best performing stable genotypes in terms of grain yield/plant across locations. Twelve accessions had a high level of resistance against both FLS and YMV diseases under natural hot-spot conditions which can be utilized as promising donors in the soybean breeding programme.
Keywords
- Type
- Research Article
- Information
- Copyright
- Copyright © The Author(s), 2023. Published by Cambridge University Press on behalf of National Institute of Agricultural Botany
References
Agarwal, DK, Billore, SD, Sharma, AN, Dupare, BU and Srivastava, SK (2013) Soybean: introduction, improvement and utilization in India-problems and prospects. Agricultural Research 2, 293–300.CrossRefGoogle Scholar
Bhartiya, A and Aditya, JP (2016) Genetic variability, character association and path analysis for yield and component traits in black seeded soybean lines under rainfed condition of Uttarakhand hills of India. Legume Research 39, 31–34.Google Scholar
Bhartiya, A, Aditya, JP, Kumari, V, Kishore, N, Purwar, JP, Agrawal, A and Kant, L (2017) GGE biplot & AMMI analysis of yield stability in multi-environment trial of soybean [Glycine max (L.) Merrill] genotypes under rainfed condition of north western Himalayan hills. The Journal of Animal and Plant Sciences 27, 227–238.Google Scholar
Bhartiya, A, Aditya, JP, Kumari, V, Kishore, N, Purwar, JP, Agrawal, A, Kant, L and Pattanayak, A (2018) Stability analysis of soybean [Glycine max (L.) Merrill] genotypes under multi-environments rainfed condition of North Western Himalayan hills. Indian Journal of Genetics and Plant Breeding 78, 342–347.Google Scholar
Chander, S, Oliveira, ALG, Gedil, M, Shah, T, Otusanya, GO, Asiedu, R and Chigeza, G (2021) Genetic diversity and population structure of soybean lines adapted to sub-Saharan Africa using single nucleotide polymorphism (SNP) markers. Agronomy 11, 604.CrossRefGoogle Scholar
DES (2020) Directorate of Economics & Statistics, Ministry of Agriculture & Farmers Welfare, Government of India. Database. Available at https://eands.dacnet.nic.in/APY_96_To_06.htm (accessed 31 March 2023).Google Scholar
FAOSTAT (2020) Food and Agriculture Organization of the United Nations (FAO). FAOSTAT Database. Available at http://faostat.fao.org/site/291/default.aspx (accessed 31 March 2023).Google Scholar
Fu, Y-B (2015) Understanding crop genetic diversity under modern plant breeding. Theoretical and Applied Genetics 128, 2131–2142.CrossRefGoogle ScholarPubMed
Ghimire, KH, Joshi, BK, Gurung, R and Sthapit, BR (2018) Nepalese foxtail millet [Setaria italica (L.) P. Beauv.] genetic diversity revealed by morphological markers. Genetic Resources and Crop Evolution 65, 1147–1157.CrossRefGoogle Scholar
Gireesh, C, Husain, SM, Shivakumar, M, Satpute, GK, Kumawat, G, Arya, M, Agarwal, DK and Bhatia, VS (2015) Integrating principal component score strategy with power core method for development of core collection in Indian soybean germplasm. Plant Genetic Resources: Characterization and Utilization 15, 230–238.CrossRefGoogle Scholar
Hou, Z, Liu, B and Kong, F (2022) Regulation of flowering and maturation in soybean. In Hon-Lam, M and Li, MW (eds), Advances in Botanical Research. London: Academic Press, pp. 43–45.Google Scholar
IBPGR (1984) International Board for Plant Genetic Resources, Descriptors of Soybean. Rome: IBPGR Secretariat, IBPGR/84/183.Google Scholar
Islam, Md R, Fujita, D, Watanabe, S and Zheng, S (2019) Variation in photosensitivity of flowering in the world soybean mini-core collections (GmWMC). Plant Production Science 22, 220–226.CrossRefGoogle Scholar
Islam, MS, Muhyidiyn, I, Islam, MR, Hasan, MK, Hafeez, ASMG, Hosen, MM, Saneoka, H, Ueda, A, Liu, L, Naz, M, Barutçular, C, Lone, J, Raza, MA, Chowdhury, MK, Sabagh, AE and Erman, M (2022) Soybean and sustainable agriculture for food security. In Ohyama, T, Takahashi, Y, Ohtake, N, Sato, T and Tanabata, S (eds), Soybean - Recent Advances in Research and Applications. London: Intech Open. http://dx.doi.org/10.5772/intechopen.104129Google Scholar
Joshi, D, Singh, PK, Adhikari, S and Rani, S (2018) Screening of soybean germplasm for important disease prevalent in North India. International Journal of Chemical Studies 6, 2731–2733.Google Scholar
Kofsky, J, Zhang, H and Song, BH (2018) The untapped genetic reservoir: the past, current, and future applications of the wild soybean (Glycine soja). Frontiers in Plant Science 9, 949.CrossRefGoogle ScholarPubMed
Kumawat, G, Singh, G, Gireesh, C, Shivakumar, M, Arya, M, Agarwal, DK and Husain, SM (2015) Molecular characterization and genetic diversity analysis of soybean (Glycine max (L.) Merr.) germplasm accessions in India. Physiology and Molecular Biology of Plants 21, 101–107.CrossRefGoogle ScholarPubMed
Kuswantoro, (2017) Genetic variability and heritability of acid-adaptive soybean promising lines. Biodiversitas: Journal of Biological Diversity 18, 378–382.CrossRefGoogle Scholar
Lal, SK, Rana, VKS, Sapra, RL and Singh, KP (2005) Screening and utilization of soybean germplasm for breeding resistances against Mungbean Yellow Mosaic Virus. Soybean Genetics Newsletter 32, 8083.Google Scholar
Liu, L, Song, W, Wang, L, Sun, X, Qi, Y, Wu, T, Sun, S, Jiang, B, Wu, C, Hou, W, Ni, Z and Han, T (2020) Allele combinations of maturity genes E1-E4 affect adaptation of soybean to diverse geographic regions and farming systems in China. PLoS ONE 15, e0235397.CrossRefGoogle ScholarPubMed
McDonald, SC, Buck, J and Li, Z (2022) Automated, image-based disease measurement for phenotyping resistance to soybean frogeye leaf spot. Plant Methods 18, 103. https://doi.org/10.1186/s13007-022-00934-7CrossRefGoogle ScholarPubMed
Mishra, KK, Bhartiya, A and Kant, L (2021) Identification of resistance sources against frogeye leaf spot (FLS) disease of soybean caused by Cercospora sojina under hot spot conditions of Uttarakhand hills. Indian Phytopathology 74, 1117–1121.CrossRefGoogle Scholar
Naveesh, YB, Prameela, HA, Basavaraj, S and Rangaswamy, KT (2020) Screening of soybean genotypes to soybean yellow mosaic virus disease. International Journal of Current Microbiology and Applied Sciences 9, 2070–2076.CrossRefGoogle Scholar
Oliveira, MF, Nelson, RL, Geraldi, IO, Cruz, CD and de Toledo, JFF (2010) Establishing a soybean germplasm core collection. Field Crops Research 119, 277–289.CrossRefGoogle Scholar
Pandey, S and Bhatore, A (2018) Genetic diversity analysis for quantitative traits in indigenous germplasm of lentil in Madhya Pradesh. Journal of Pharmacognosy and Phytochemistry 7, 279–283.Google Scholar
Shannon, C and Weaver, W (1949) The Mathematical Theory of Communication. Chicago: The University of Illinois, pp. 3–24.Google Scholar
Sharma, P, Dupare, BU and Patel, RM (2016) Soybean improvement through research in India and socio-economic changes. Legume Research 39, 935–945.Google Scholar
Singh, RJ and Hymowitz, T (1999) Soybean genetic resources and crop improvement. Genome 42, 605–616.CrossRefGoogle Scholar
Terzic, D, Popovic, V, Tatic, M, Vasileva, V, Đekic, V, Ugrenovic, V, Popovic, S and Avdic, P (2018) Soybean area, yield and production in world. XXII Eco-Conference, Ecological Movement of Novi Sad, Ministry of Education, Science and Technology Development of the Republic of Serbia.Google Scholar
Vennampally, N, Bhartiya, A, Singh, CP, Devi, HN, Deshmukh, MP, Verghese, P, Singh, K, Mehtre, SP, Kumari, V, Maranna, S, Kumawat, G, Ratnaparkhe, MB, Satpute, GK, Rajesh, V, Chandra, S, Ramteke, R, Khandekar, N and Gupta, S (2021) WAASB-based stability analysis and simultaneous selection for grain yield and early maturity in soybean. Agronomy Journal 4, 3089–3099.Google Scholar
Yan, W (2002) Singular value partitioning in biplot analysis of multi-environment trial data. Agronomy Journal 94, 990–996.Google Scholar
Yang, W and Weaver, DB (2001) Resistance to frogeye leaf spot in maturity groups VI and VII of soybean germplasm. Crop Science 41, 549–552.CrossRefGoogle Scholar

Bhartiya et al. supplementary material 1
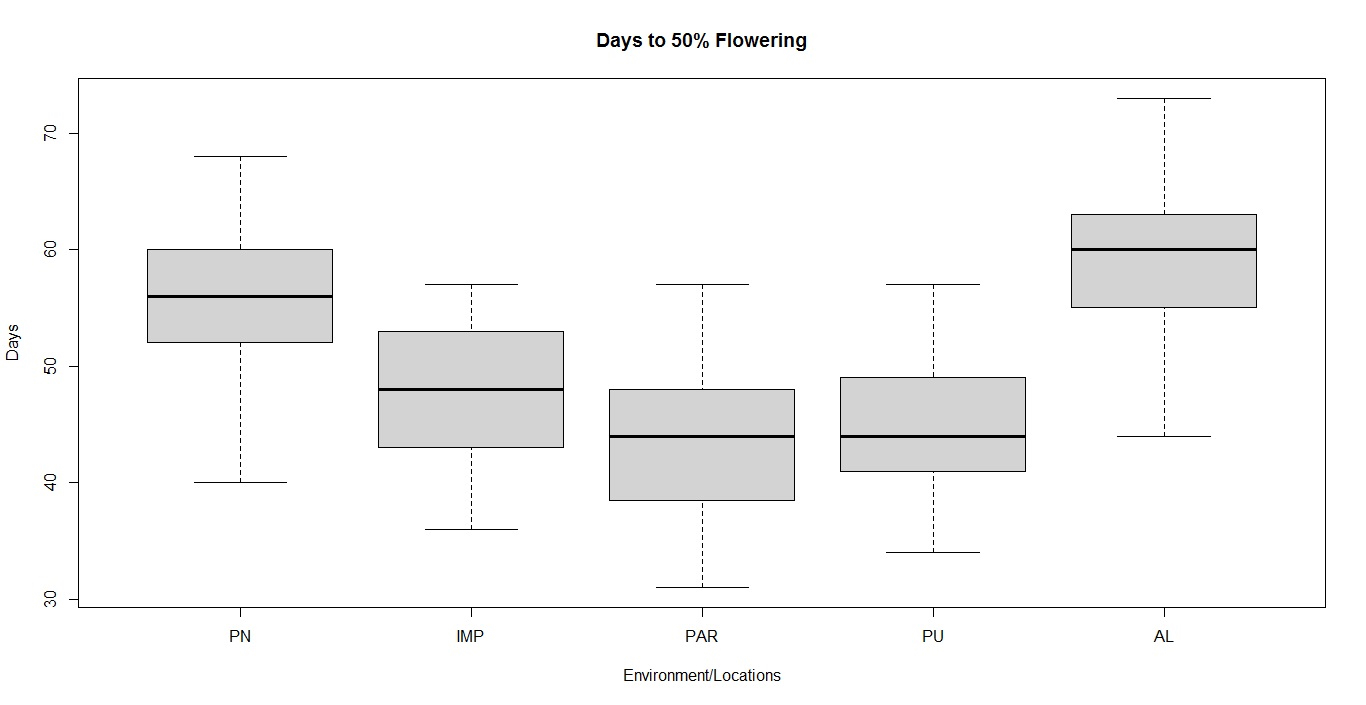
File
103.7 KB
Bhartiya et al. supplementary material 2
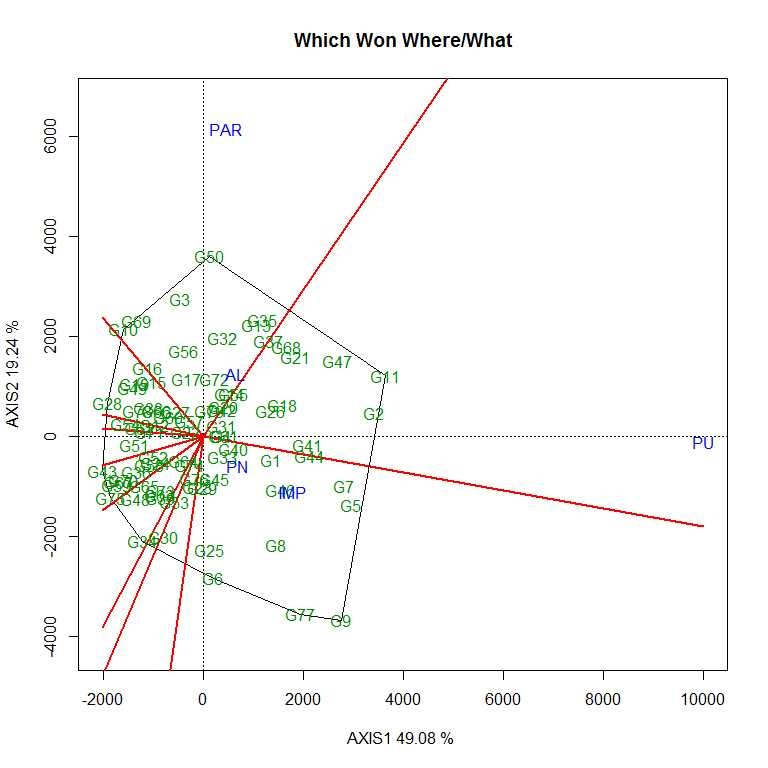
File
200.8 KB
Bhartiya et al. supplementary material 3
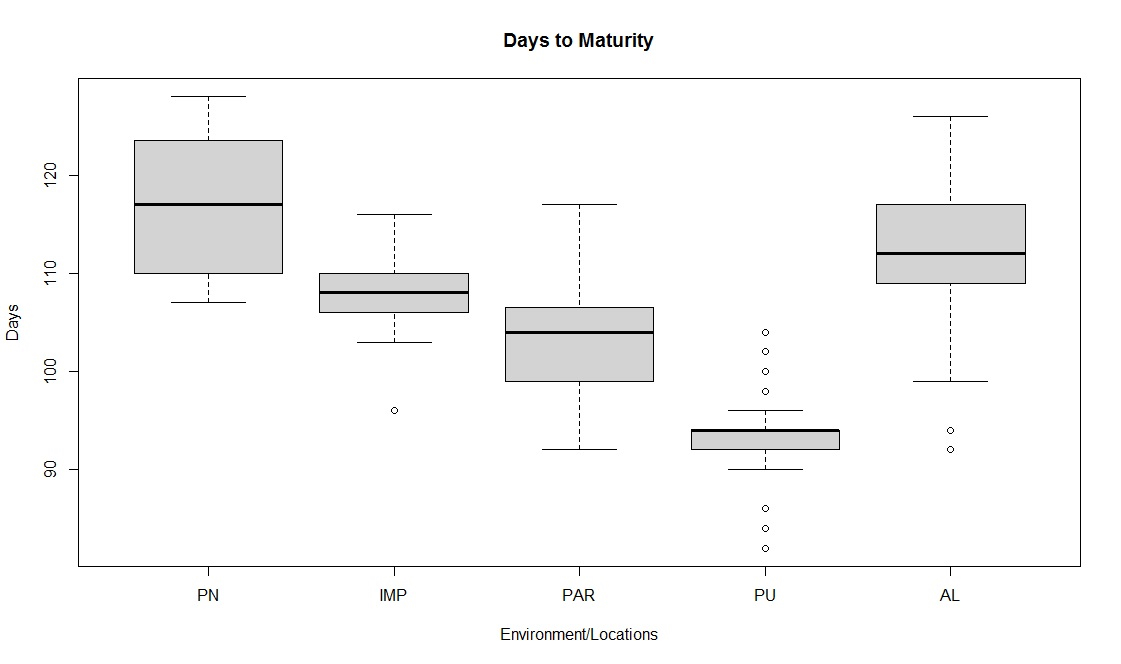
File
86.3 KB
Bhartiya et al. supplementary material 4
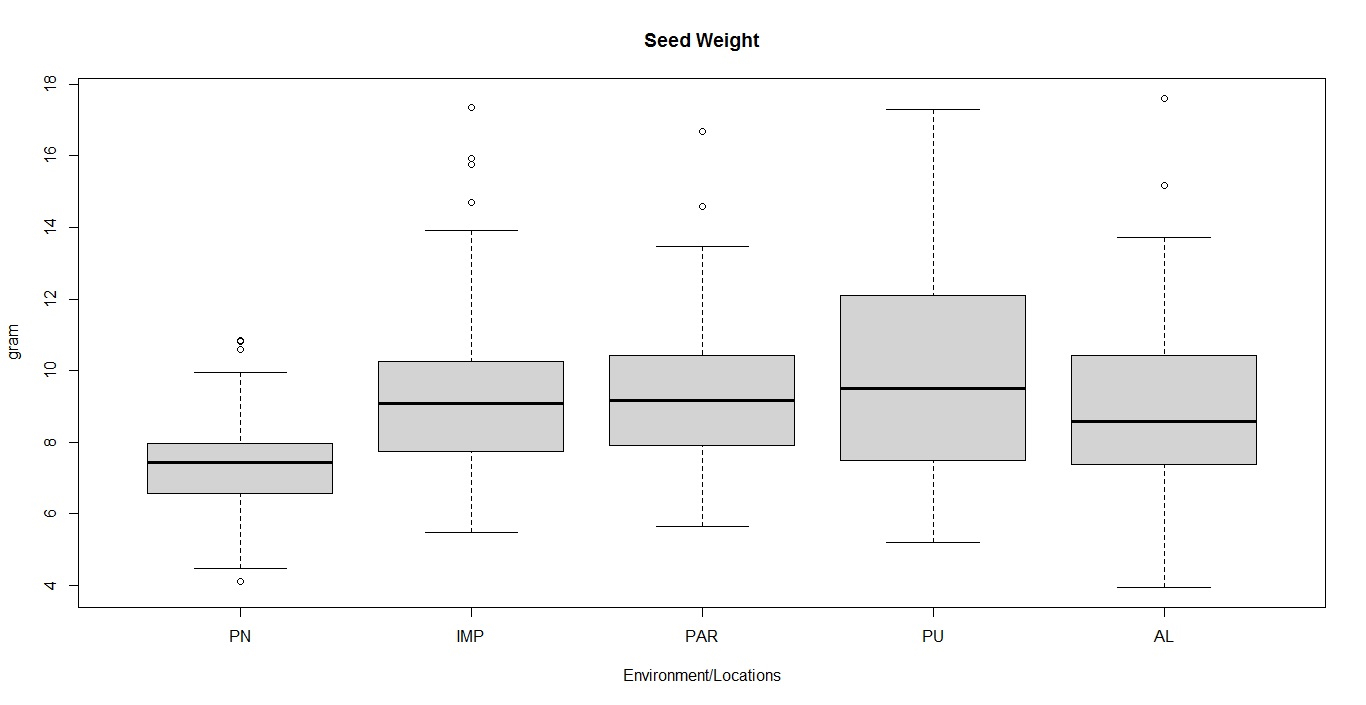
File
106 KB
Bhartiya et al. supplementary material 5
File
98.3 KB
Bhartiya et al. supplementary material 6
File
550.1 KB
Bhartiya et al. supplementary material 7
File
212.1 KB
Bhartiya et al. supplementary material 8
File
200.4 KB
Bhartiya et al. supplementary material 9
File
197.8 KB
Bhartiya et al. supplementary material 10
File
189.3 KB
Bhartiya et al. supplementary material 11

File
21.7 KB
Bhartiya et al. supplementary material 12
File
12.9 KB