



Introduction
In order to fully document a well-preserved radiolarian fauna from the middle Darriwilian Table Cove Formation in Newfoundland, this paper is intended to complement earlier work of Kachovich and Aitchison (Reference Kachovich and Aitchison2020) that described spumellarian radiolarians from the same location. Archaeospicularid and entactiniid radiolarians, together with forms that remain in open nomenclature, are described herein.
Radiolarian faunas have been reported from Ordovician strata globally from North and South America, Spitsbergen, Scotland, Russia, China, and Australia (see Danelian et al., Reference Danelian, Caridroit, Noble and Aitchison2017 and references therein). Documentation of Middle Ordovician (Darriwilian) radiolarians has been presented from Newfoundland (Kachovich and Aitchison, Reference Kachovich and Aitchison2020), Argentina (Maletz et al., Reference Maletz, Albanesi and Voldman2009), Kazakhstan (Nazarov, Reference Nazarov1975; Nazarov et al., Reference Nazarov, Popov and Apollonov1977; Nazarov and Ormiston, Reference Nazarov and Ormiston1993; Pouille et al., Reference Pouille, Danelian, Pour and Popov2013, Reference Pouille, Danelian and Popov2014b), Scotland (Danelian and Clarkson, Reference Danelian and Clarkson1998; Danelian, Reference Danelian1999; Danelian and Floyd, Reference Danelian and Floyd2001; Danelian et al., Reference Danelian, Noble, Pouille and Maletz2013; Perera et al., Reference Perera, Aitchison and Nothdurft2020), and China (Li, Reference Li1995; Buckman and Aitchison, Reference Buckman and Aitchison2001; Yi et al Reference Yi, Yuan, Aitchison and Feng2018).
A lack of precise taxonomic descriptions of many taxa and stratigraphic knowledge gaps continue to hinder understanding of phylogenetic relationships among taxa within this important Phanerozoic fossil group, especially in the earliest stages of their evolution (Noble et al., Reference Noble, Aitchison, Danelian, Dumitrică, Maletz, Suzuki, Cuvelier, Caridroit and O'Dogherty2017). Many radiolarians grow outwards from an initial spicule observation, which is commonly obscured by development of later skeletal elements, such as concentric spongy, spicular, or porous shells. Important internal structural details also can be obscured or destroyed by diagenesis and/or poor preservation. This continues to impede our understanding of higher-level taxonomy. Stratigraphic and paleogeographic ranges of many of the important morphological groups remain poorly constrained because detailed reports understandably focus on isolated occurrences where taphonomic conditions have been the most favorable (Aitchison et al., Reference Aitchison, Suzuki, Caridroit, Danelian and Noble2017). Unfortunately, although radiolarians are the dominant fossils among early Paleozoic deep-water marine lithofacies such as chert, this is typically where preservation states are worse. Recent molecular investigations into both modern entactinarians and spumellarians are providing additional insight into the time of origin of these orders and their evolution.
Lower Paleozoic carbonates in Newfoundland are well known for numerous well-preserved radiolarian faunas from upper Cambrian to Lower Ordovician strata (Bergström, Reference Bergström1979; Aitchison et al., Reference Aitchison, Flood and Malpas1998; Zhang and Pratt, Reference Zhang and Pratt2000; Won and Iams, Reference Won and Iams2002, Reference Won and Iams2011, Reference Won and Iams2013, Reference Won and Iams2015; Maletz and Bruton, Reference Maletz and Bruton2005, Reference Maletz and Bruton2007, Reference Maletz and Bruton2008; Won et al., Reference Won, Iams and Reed2005; Maletz, Reference Maletz2007, Reference Maletz2011; Pouille et al., Reference Pouille, Danelian and Maletz2014a). Previous reports from the Middle Ordovician Table Cove Formation have mentioned the presence of radiolarians at Piccadilly Quarry (Bergström, Reference Bergström1979; Renz, Reference Renz1990; Zhang and Pratt, Reference Zhang and Pratt2000; Maletz and Bruton, Reference Maletz and Bruton2007), a locality well-known for other fossil occurrences. Twenty-seven archaeospicularid and entactiniid taxa, six of which belong to species newly described herein, are recognized.
Geologic setting
The Table Head Group in the Port au Port Peninsula area of Newfoundland is well studied (Stenzel et al., Reference Stenzel, Knight and James1990). Tectonically, it comprises part of an autochthonous Cambrian to Ordovician age passive margin carbonate shelf rift-drift assemblage that developed on the Laurentian margin during opening of the Iapetus Ocean (Williams and Hiscott, Reference Williams and Hiscott1987; Torsvik et al., Reference Torsvik, Smethurst, Meert, Van der Voo, McKerrow, Brasier, Sturt and Walderhaug1996; Lacombe et al., Reference Lacombe, Waldron and Williams2020). This margin was tectonically overthrust in a westerly direction by the Humber Arm Allochthon in response to ocean closure during the Taconic orogeny. The allochthon incorporates a series of nappes that includes the Cow Head Group, together with the Bay of Islands Ophiolite (Cawood and Suhr, Reference Cawood and Suhr1992) and Little Port complexes (Williams, Reference Williams1973; Jenner et al., Reference Jenner, Dunning, Malpas, Brown and Brace1991), from which numerous other Ordovician radiolarian assemblages have been described.
Locality information
Samples were collected from dark gray nodules within a 4 m thick fossiliferous, peloidal limestone section in the Table Cove Formation exposed at the Piccadilly Quarry, Port au Port Peninsula, Newfoundland (48.59155556°N, 58.92027778°W, WGS84, Figs. 1, 2). The limestones contain a fauna that includes brachiopods and hexactinellid sponge spicules (Zhang and Pratt, Reference Zhang and Pratt2000), together with a rich graptolite assemblage that correlates to the middle Darriwilian Holmograptus spinosus Biozone (Maletz and Egenhoff, Reference Maletz, Egenhoff, Guitérrez-Marco and Rábano2011). Co-occurring conodonts allow correlation to the late Dw2 Histiodella kristinae subzone (Stouge, Reference Stouge1984).
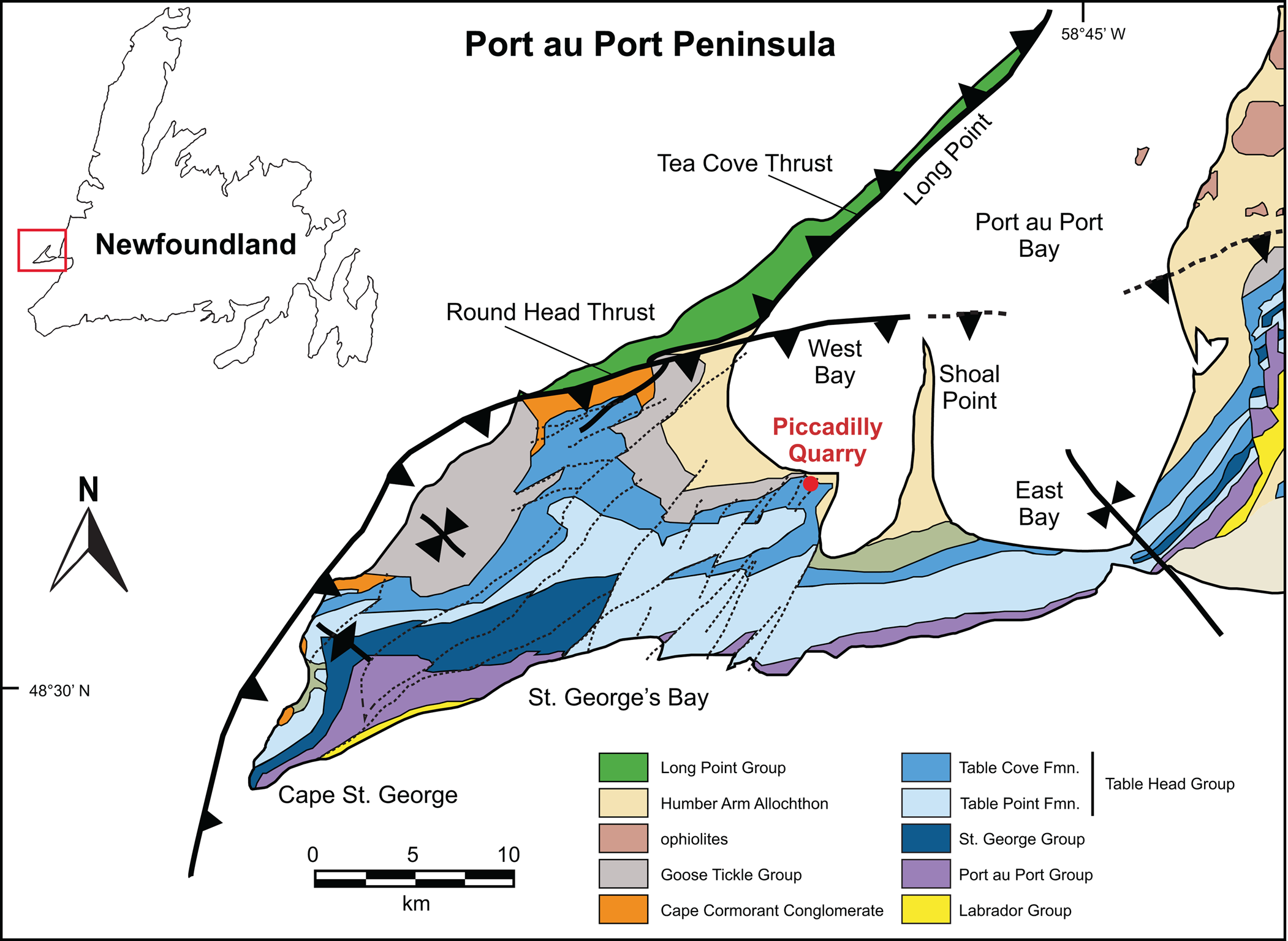
Figure 1. Locality map showing position of study area (after Cooper et al., Reference Cooper, Weissenberger, Knight, Hostad, Gillespie, Williams and Clark2001; Kachovich and Aitchison, Reference Kachovich and Aitchison2020). Inset map shows Port au Port Peninsula's location in relation to the rest of Newfoundland. Reprinted with permission from Kachovich and Aitchison (Reference Kachovich and Aitchison2020, fig. 1).

Figure 2. Stratigraphic context of the Table Cove Formation at the Piccadilly Quarry. (1) Lithostratigraphic log (after Maletz and Egenhoff, Reference Maletz, Egenhoff, Guitérrez-Marco and Rábano2011; Kachovich and Aitchison, Reference Kachovich and Aitchison2020) showing the sampled levels (PD01, -03, -05, -07, -09, -11, -12, and -13). Generalized stratigraphy after Knight (Reference Knight1991) and James et al. (Reference James, Botsford and Williams1987); (2) field photo, facing southwest, of the exposure of gently dipping beds of rhythmically bedded limestone. Reprinted with permission from Kachovich and Aitchison (Reference Kachovich and Aitchison2020, fig. 3).
Radiolarian biostratigraphy
Ordovician stratigraphy used herein follows North American and Newfoundland zonal schemes for conodonts and graptolites (Morris and Kay, Reference Morris and Kay1966; Williams et al., Reference Williams, Boyce and James1987; Maletz, Reference Maletz2009; Loydell, Reference Loydell2012). Globally, twelve radiolarian assemblages are presently recognized in the Ordovician (Aitchison et al., Reference Aitchison, Suzuki, Caridroit, Danelian and Noble2017). Middle Ordovician radiolarian biostratigraphy follows the work of Nazarov and Popov (Reference Nazarov and Popov1980), Nazarov (Reference Nazarov1988), Danelian and Popov (Reference Danelian and Popov2003), Jones and Noble (Reference Jones and Noble2006), MacDonald (Reference MacDonald2006), Pouille et al. (Reference Pouille, Danelian, Pour and Popov2013), and Aitchison et al. (Reference Aitchison, Suzuki, Caridroit, Danelian and Noble2017), and stratigraphic correlation for the radiolarians described herein is discussed elsewhere (Kachovich and Aitchison, Reference Kachovich and Aitchison2020).
Materials and methods
The materials and methods used in this investigation, including those for μ-CT imagery and analysis, are presented in detail by Kachovich et al (Reference Kachovich, Sheng and Aitchison2019) and Kachovich and Aitchison (Reference Kachovich and Aitchison2020). Examples of micro-CT models of skeletons of various ‘primitive’ entactinarians, with a six-rayed internal spicule with a median bar (MB) (colored in green) and а spherical sphere with irregular and three-dimensional meshwork (gray), are presented in Figure 3.
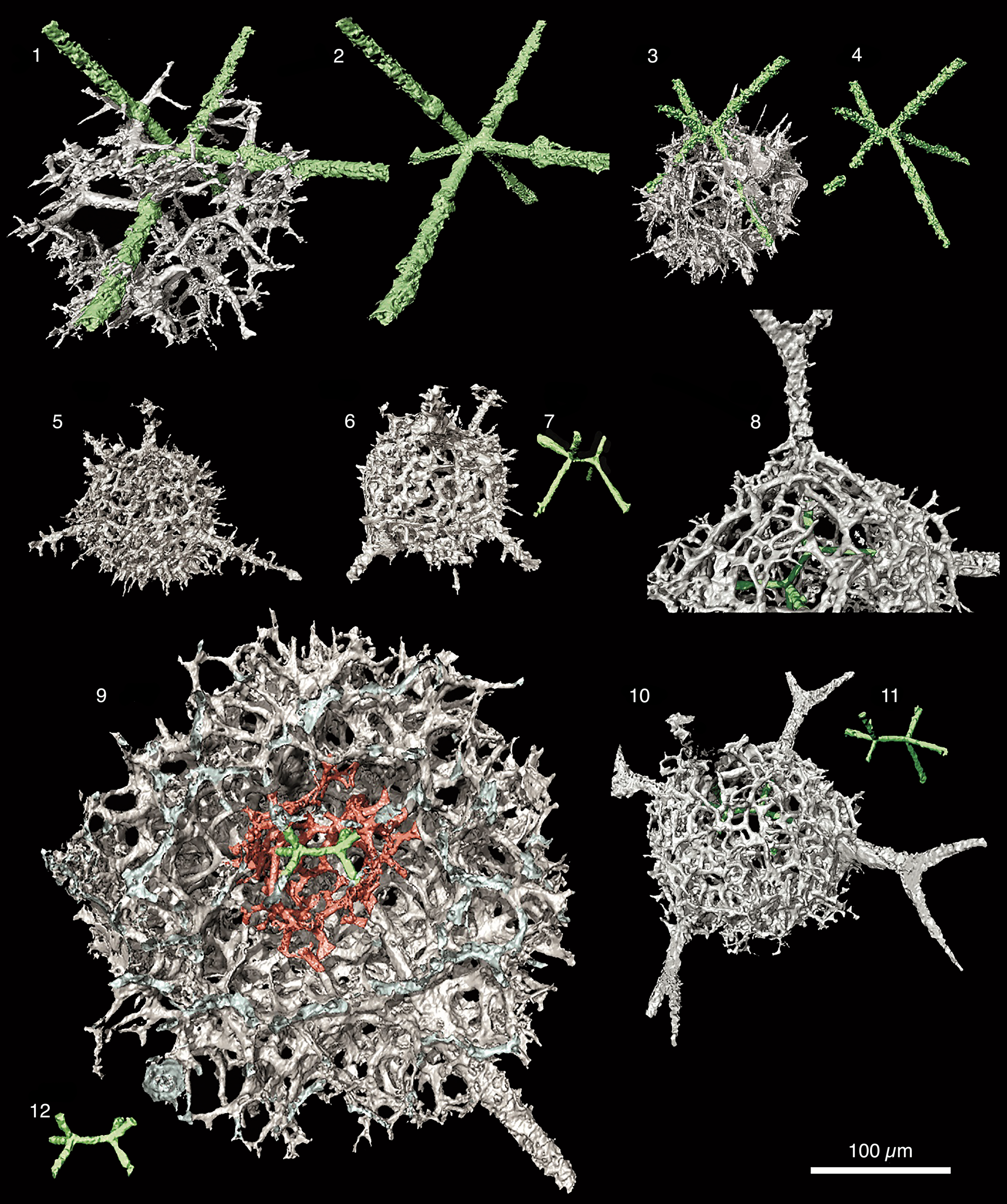
Figure 3. Micro-CT models of skeletons of various ‘primitive’ entactinarians, with a six-rayed internal spicule with a median bar (MB) (colored in green) and а spherical sphere with irregular and three-dimensional meshwork (gray). MB varies in length, thickness, and position within the skeleton between different families. Scale bar = 100 μm. (1) Ramuspiculum laxum n. sp. (UQSEES_M1S03 holotype from PD09), (2) MB = 6.6 μm; (3) Sphaeroentactinia sp. aff. S. integrata (Maletz and Bruton) (UQSEES_M2S07 from PD13), (4) MB = 11.5 μm; (5, 6) Varispiculum ectospiculatum Won and Iams (UQSEES_M3S12 from PD13), (7) MB = 16.8 μm; (8, 10) Entactiniid gen. and sp. indet. A (UQSEES_M2S05 from PD05), (11) MB = 27.8 μm; (9) Spongentactinia armillata (Nazarov) (UQSEES_M7S20 from PD13), (12) MB = 33.8. μm.
Repositories and institutional abbreviations
Specimens examined in this study are curated at the School of Earth and Environmental Science, University of Queensland (UQSEES), Brisbane, Australia. Other repositories for holotypic material referred to in the text include the Geological Institute, Moscow (GIN), and Geological Survey of Canada (GSC).
Systematic paleontology
Taxonomic review
Although a rigid framework for classification of early polycystine radiolarians is needed, this is currently difficult to develop because of imperfections in the fossil record and relatively few studies that have been made on internal skeletal elements. In this study, specimens from well-established families, such as the Archeoentactiniidae, Echidninidae, Palaeospiculidae, Entactiniidae, and Aspiculidae, were observed using μ-CT scans, SEM, and TLM images, and possible relationships among families and genera are discussed.
Family- and genus-level systematics follow Noble et al. (Reference Noble, Aitchison, Danelian, Dumitrică, Maletz, Suzuki, Cuvelier, Caridroit and O'Dogherty2017). The family Aspiculidae and its genus Nyfrieslandia, as well as the genus Westernbrookia within the family Echidninidae, are emended. The characteristics of the above-mentioned families and genera are re-evaluated in the light of μ-CT investigations.
Phylum Radiozoa Cavalier-Smith, Reference Cavalier-Smith1987
Class Polycystina Ehrenberg, Reference Ehrenberg1838, sensu Riedel, Reference Riedel1967
Order Archaeospicularia Dumitrică, Caridroit, and De Wever, Reference Dumitrică, Caridroit and De Wever2000
Family Archeoentactiniidae Won and Below, Reference Won and Below1999
Genus Pararcheoentactinia Won and Iams, Reference Won and Iams2002
Type species
Pararcheoentactinia reedae Won and Iams, Reference Won and Iams2002.
Pararcheoentactinia reedae Won and Iams, Reference Won and Iams2002
Figure 4.5
- Reference Won and Iams2002
Pararcheoentactinia reedae Won and Iams, p. 13, figs. 8–10.
- Reference Won, Iams and Reed2005
Pararcheoentactinia reedae; Won et al., p. 438, fig. 5.14–5.16.
- Reference Won, Iams and Reed2007
Pararcheoentactinia reedae; Won et al., p. 504, fig. 4.3, 4.4.
- Reference Pouille, Danelian and Maletz2014
Pararcheoentactinia reedae; Pouille et al., Reference Pouille, Danelian and Maletz2014a, pl. 1, fig. 15, pl. 2, figs.16–25.
- Reference Won and Iams2015
Pararcheoentactinia spp. cf. P. reedae; Won and Iams, p. 12, pl. 1, figs. 10a, b.

Figure 4. Scanning electron micrographs of radiolarians extracted from limestones from the Piccadilly Quarry, Newfoundland Canada. Scale bar = 100 μm: (1, 2) Svalbardospiculum multifurcatum Won, Iams, and Reed; PD02; (3, 4) Sphaeroentactinia sp.; PD05; (5) Pararcheoentactinia reedae Won and Iams; PD09; (6–10) Sphaeroentactinia sp. aff. S. integrata (Maletz and Bruton); PD13; (11, 12) Svalbardospiculum sp. aff. S. hexaradiatum (Won and Iams); PD13; (13–16) Entactiniid gen. and sp. indet. C; (13) PD02; (14–16) PD13.
Holotype
Specimen (GSC-122014 GP6-44), Green Point Formation, Cow Head Group, Canada (Won and Iams, Reference Won and Iams2002, fig. 9.13).
Occurrence
Upper Cambrian, Franconian–Lower Ordovician, upper Floian, Cow Head Group, Newfoundland, Canada.
Materials
Single specimen (sample PD09), Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Specimen fits within range of variation among Pararcheoentactinia reedae Group reported by Won and Iams (Reference Won and Iams2002). This discovery extends the known range of P. reedae to the mid-Darriwilian.
Genus Sphaeroentactinia Maletz and Bruton, Reference Maletz and Bruton2007
Type species
Sphaeroentactinia hexaspinosa Maletz and Bruton, Reference Maletz and Bruton2007.
Remarks
Taxa within the genus Sphaeroentactinia can be distinguished by the varying development of the initial spicule and the degree of integration of spinules into the skeletal wall.
Sphaeroentactinia sp. aff. S. integrata Maletz and Bruton, Reference Maletz and Bruton2007
Figures 3.3, 3.4, 4.6–4.10, 5; Supplemental Data file 1
- aff. Reference Maletz and Bruton2007
Sphaeroentactinia integrata Maletz and Bruton, p. 265, figs. 6D, 14A–I.

Figure 5. Micro-CT model UQSEES_M2S07 of Sphaeroentactinia sp. aff. S. integrata (Maletz and Bruton) illustrating the relationship between the distinct bar-centered initial spicule (green) and the outer skeleton (gray). The 3D meshwork originates from irregularly spaced positions on the basal rays (red) and appears to have no taxonomic value. Labels A and B are arbitrary and attached to spines in order to allow viewers to better follow specimen rotation. (1, 2) Same μ-CT model viewed at different angles at the same magnification; (3) zoom view of initial spicule in center of model.
Description
The initial spicule is eccentrically integrated into the sphere and consists of a MB (9–17 μm long) and three rays with a tetrahedral distribution at each end. A hollow, spherical skeleton (dimeter [Ø] = 137–153 μm) with a disorderly interwoven meshwork of thin, straight bars formed from a bar-centered spicule with six-rays. The terminal primary rays branch trichotomously to form a sphere in connection with the growing mesh of the apophyses starting from the primary rays. The spherical skeleton diameter is ~95–115 μm. The three apical rays are free from the outer sphere, where the three long basal rays (up to 330 μm long) pass through and form a part of the skeletal wall. The MB on the initial spicule is positioned ectopically on the skeletal wall. The outer sphere is constructed from compactly interwoven thin spinules that originate on the basal rays to form a polygonal or angular meshwork. The surface is very rough because of the protruding parts of the bars. The inner surface of the outer sphere is smooth.
Materials
Fourteen specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Sphaeroentactinia sp. aff. S. integrata is distinguished from other taxa within Sphaeroentactinia by the integration of most of the primary rays in the sphere. Furthermore, the sphere is developed from a connection of primary rays and apophyses, and not exclusively from apophyses, as in S. hexaspinosa. In the SEM images, the microbar is sometimes difficult to observe due to the orientation of the specimen. Commonly the six spines are slightly asymmetrical, suggesting a bar-centered spicule with a short median bar. Maletz and Bruton (Reference Maletz and Bruton2007) described and illustrated the species in some detail, based on material mounted on SEM stubs. Material investigated using μ-CT displays (Fig. 4) the irregular relationship between the distinct bar-centered initial spicule and the meshwork of the outer sphere. The irregularly positioned three-dimensional meshwork appears to have no taxonomic value. Sphaeroentactinia integrata differs from S. hexaspinosa with integration of distinct rays of the spicule into the spherical skeleton.
Sphaeroentactinia robusta Won and Iams, Reference Won and Iams2015
Figure 6.3, 6.6, 6.7, 6.9
- Reference Maletz2007
Archaeoentactinid sp. indet. Maletz, p. 80, fig. 3I.
- Reference Won and Iams2015
Sphaeroentactinia robusta Won and Iams, p. 21, pl. 5, figs. 2, 3.

Figure 6. Scanning electron micrographs of radiolarians extracted from limestones from the Piccadilly Quarry, Newfoundland, Canada. Scale bar = 100 μm. (1) Protoentactinia sp.; PD05; (2, 4, 5) Ramuspiculum laxum n. sp.; (2) PD13; (4, 5) PD02; (3, 6, 7, 9) Sphaeroentactinia robusta Won; (3) PD13; (6, 7, 9) PD05; (8, 13) Varispiculum ectospiculatum Won and Iams; PD13; (10, 11) Ramuspiculum sp.; PD05; (12) Echidnina sp. aff. E. conexa Won, Iams, and Reed; PD02.
Holotype
Specimen (GSC 131223), Cow Head Group, Canada (Won and Iams, Reference Won and Iams2015, pl. 5, fig. 3).
Occurrence
Ordovician, Upper Floian to Dapingian, Cow Head Group, Newfoundland, Canada.
Description
Well-developed, bar-centered spicule, which repeatedly branches to form a loose sub-globular meshwork. Initial spicule is prominent and thicker in comparison to the delicate, loose meshwork. The main rays may be branched up to four or five times, but this is difficult to determine due to the delicate nature of the meshwork.
Materials
Four specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
The relative rarity of Sphaeroentactinia robusta may be related to the likelihood of preservation of this form, given the delicate nature of the meshwork. This discovery extends the known range of S. robusta to the mid-Darriwilian.
Sphaeroentactinia sp.
Figure 4.3, 4.4
Description
Small sub-globular skeleton (Ø ~75 μm) constructed from delicately branched rays and apophyses from a robust initial spicule. The sphere has a hollow interior. The initial spicule is ectopically placed into the sphere, where the visible external primary spines attain a length of 100 μm with a pointed tip. The MB is ~10–15 μm long and 5–8 μm wide. Secondary external spines may be formed. These are much smaller than the primary rays and, thus, easily differentiated.
Materials
Two specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Only two broken specimens were observed from sample PD05, thus the true nature of this Sphaeroentactinia variant cannot be properly assessed.
Family Echidninidae Kozur, Mostler, and Repetski, Reference Kozur, Mostler and Repetski1996
Genus Echidnina Bengtson, Reference Bengtson1986 sensu Won and Iams, Reference Won and Iams2002
Type species
Echidnina runnegari Bengtson, Reference Bengtson1986.
Echidnina sp. cf. E. conexa Won, Iams, and Reed, Reference Won, Iams and Reed2005
Figure 6.12
- cf. Reference Won and Iams2002
Echidnina sp. Won and Iams, p. 26, figs. 12.15, 12.16.
- cf. Reference Won, Iams and Reed2005
Echidnina conexa Won et al., p. 447, figs. 3.14–3.16a, b.
- cf. Reference Won, Iams and Reed2005
Echidnina semiconexa Won et al., p. 447, figs. 3.7–3.12.
- cf. Reference Won, Iams and Reed2005
Echidnina severedeformis Won et al., p. 449, figs. 4.7, 4.8a, b.
- cf. Reference Pouille, Danelian and Maletz2014a
Echidnina conexa Pouille et al., pl. 5, figs. 1–29.
Description
Skeleton made of interlocked spicules. Numerous more or less fused six-rayed, point-centered spicules, packed densely together in such a way as to form a hollow sphere with a latticed meshwork. Rays pass tangentially to the inner surface and, when fused, display a smooth inner structure. The free ends of the rays, directed outwardly, are short.
Materials
One specimen, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Diameter of the outer sphere of the specimen described (140 μm) is noticeably less than that of the holotype of E. conexa. This discovery extends the known range of Echidnina to the mid-Darriwilian.
Genus Varispiculum Won and Iams, Reference Won and Iams2015
Type species
Varispiculum ectospiculatum Won and Iams, Reference Won and Iams2015.
Remarks
The genus Varispiculum lacks the skeletal meshwork of a protoentactiniid and has minor spicules. It differs from all other genera of the family Echidninidae in the presence of spinules along the rays that form а part of the skeletal meshwork. Differences between species can be found in the development of the primary spicule and the way the spicule is integrated into the skeletal wall.
Varispiculum ectospiculatum Won and Iams, Reference Won and Iams2015
Figures 3.5–3.7, 6.8, 6.13; Supplemental Data file 2
- Reference Won and Iams2015
Varispiculum ectospiculatum Won and Iams, p. 17, pl. 4, figs. 12, 13, pl. 5, figs. 8–14.
Holotype
Specimen (GSC 131232), Cow Head Group, Canada (Won and Iams, Reference Won and Iams2015, pl. 5, fig. 9a, b).
Occurrence
Lower to Middle Ordovician, Соw Head Group, Newfoundland, Canada.
Description
Small sphere (Ø = 100 μm) with irregular and three-dimensional meshwork. A prominent three-rayed, bar-centered spicule that is positioned ectopically inside the skeleton. Spinules common and densely developed along the rays, some of them connected to skeletal meshwork. The distal parts of the rays commonly protrude as outer spines, along which radially and equidistantly or sub-equidistantly distributed spinules (thorns) are densely developed and contribute to form the skeletal meshwork.
Materials
Two specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Relatively uncommon among the samples examined. This discovery extends the known range of Varispiculum ectospiculatum to the mid-Darriwilian.
Genus Westernbrookia Won, Iams, and Reed, Reference Won, Iams and Reed2007
Type species
Westernbrookia diversa Won, Iams, and Reed, Reference Won, Iams and Reed2007.
Remarks
Emended from Won et al. (Reference Won, Iams and Reed2007) to include forms with up to 11 radial rays that arise from a polygonal spicular system and a single ray that is connected to the pylome rim and develops into a short main spine.
Details of the delicate internal structure of this taxon were only revealed during observation with μ-CT. Our material appears to be a form that may be intermediate between Westernbrookia and the Upper Ordovician taxon Kalimnasphaera maculosa Webby and Blom, Reference Webby and Blom1986 (Webby and Blom, Reference Webby and Blom1986; Noble and Webby, Reference Noble and Webby2009). The ‘‘double-walled latticed medullary shell’’ described by Noble and Webby (Reference Noble and Webby2009) from the Katian, K. maculosa, was not observed. However, there are possible relics of a second internal structure that might have been too delicate to be preserved. As noted by Pouille et al. (Reference Pouille, Danelian, Pour and Popov2013), geologically older forms of Kalimnasphaera are likely to only preserve the second “medullary shell” because these older forms are much thinner and other spheres may be easily dissolved by taphonomic processes. Unlike the Upper Ordovician Kalimnasphaera taxa (and other Paleozoic families), the spherical “inner shell” (here termed “microsphere”) forms as intergrowths between the initial tetrapetaloid structure with a short MB and four wide pores. The spines (most readily observed using a μ-CT scan, such as that in Supplemental Data file 3 where the microsphere is colored red) are joined by arches in the proximal part to form four wide pores (the fourth poorly developed) with the MB in the center. This tetrapetaloid structure is a characteristically seen in younger radiolarians, such as Hollandosphaera hexagonium (Hollande and Enjumet, Reference Hollande and Enjumet1960), which is common in Mesozoic–Cenozoic assemblages (Dumitrică, Reference Dumitrică2013).
The most noteworthy feature observed in the middle Darriwilian genus Westernbrookia is the distinct spicular system with a MB and a primitive tetrapetaloid structure. Heteropolar inner spicular systems are also common among radiolarians with a pylome in the families Pylentonemidae, Proventocitidae, and Popofskyellidae. The precise development of the spicular systems in these families has not been described in detail because of the difficulties (even with μCT technology) in studying such a very thin skeletal element that is rarely well preserved.
Westernbrookia polygonata new species
Figures 7.3, 7.6, 7.9, 8, 9.9, 9.11, 9.12; Supplemental Data file 3

Figure 7. Compilation of μ-CT models, SEM images, and line drawings illustrating the family Aspiculidae from sample PD13. (1, 4, 7) Nyfrieslandia sp. aff. N. complicata; (2, 5, 8) Nyfrieslandia ramosissima n. sp.; (3, 6, 9) Westernbrookia polygonata n. sp.; (10, 11) Aspiculum irregulare n. sp.; (12) line drawing of Upper Ordovician Kalimnasphaera maculosa after Webby and Blom (Reference Webby and Blom1986).

Figure 8. Compilation of line drawings, μ-CT models, and SEM image of the holotype Westernbrookia polygonata n. sp. (UQSEES_M4S13), illustrating the heteropolar microsphere in relation to its skeleton; median bar (MB); (1–3) shown at the same scale and (6–8) shown at the same scale. (1) Schematic representation of the relationship of skeletal features; note the eleventh ray (s-py), in the basal position, does not develop into a main spine on the outer sphere; (2) details showing the true relationship between features; note the large pylomate opening (py); (3) detailed diagram of the microsphere; (4) apical view of a μ-CT segmentation of the microsphere showing the relationship between the MB and five apical rays (rb = radial bar, LG = longitudinal gate, TG = transverse gate); (5) planar extraction of the apical and basal hemispheres; (6) basal view of the μ-CT model of Westernbrookia polygonata n. sp. with enlargement of internal detail; (7) showing the alignment of the pylome, large pore on the microsphere aligned with the pylome and the MB on the apical side of the microsphere; (8) a microsphere comparable to Westernbrookia polygonata n. sp. that was reported by Maletz and Bruton (Reference Maletz and Bruton2007) from the Lower to Middle Ordovician (upper Floian to lower Dapingian) of the Valhallfonna Formation, Spitsbergen.

Figure 9. Radiolarians extracted from limestones from the Piccadilly Quarry, Newfoundland, Canada. Images are scanning electron micrographs, except (3, 4), which are transmitted-light images. Scale bar = 100 μm. (2, 4) Aspiculum sp. A; PD13; (3, 7, 8) Aspiculum sp. B; PD13; (1, 10, 13) Aspiculum irregulare n. sp.; PD13, (10 holotype UQSEES_2015PD13_01); (5, 6) Westernbrookia sp.; PD13; (9, 11, 12) Westernbrookia polygonata n. sp.; PD13; (14–17) Spongentactinia nazarovi n. sp.; (14) PD09; (15, 17) PD13; (16) PD05, holotype UQSEES_2015PD05_01.
Holotype
UQSEES_M4S13, Figure 8, Supplemental Data file 3; sample PD13, Middle Ordovician, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Diagnosis
Spherical to polygonal, pylomate outer sphere (outer sphere Ø = 180–198 μm) with a distinct polarized spicular system. A large irregular pylome opening (Ø = 74–95 μm), where the rim of the pylome opening is fringed with a small, unornamented ridge (5–7 μm thick). The outer sphere is thick and regularly latticed, with rounded to angular-oval pores (Ø = 8–26 μm, average Ø = 20 μm), and bears short and thin by-spines at vertices. Incomplete, loose apophyses on the main spine.
Occurrence
Middle Ordovician, Table Cove Formation, Newfoundland, Canada.
Description
The initial spicule is incorporated into the apical portion of the microsphere wall. It is hardly discernible, but gives rise to a heteropolar microsphere. The inner spicular system (Ø = 56 μm) is constructed of thin pore-bars that form irregular or polygonal pore frames. Pores on the microsphere are angular to oval, with irregular size and arrangement (31 pores ranging from 3–49 μm; average 31 μm). The microsphere is integrated with an ectopically placed, bar-centered inner spicular system. There is strict geometry between the positions of the MB and the pylome, where the MB is apical and the pylome is basal (Fig. 8.2). When the inner spicular system is viewed in a nominally apical position, four proximal hexagonal rings around a thick MB (28 μm long, 15 μm wide) are observed. In the basal half, pores become more polygonal with less-regular shape and arrangement.
The microsphere contains numerous polygonal, rarely hexagonal framed pores that bear 11 long, thin radial bars (rb). Radial bars originate in the wall of the microsphere rather than in the initial spicule from the bars or triple junction of the microsphere lattice. One spine is located around the pylome on the outer sphere in the antapical position and develops into a short outer spine. An additional spicular system is potentially preserved as broken nodes within the microsphere.
Eleven radial bars connect the microsphere to the outer sphere and continue as outer spines. The rod-like outer spines (up to 180 μm long) are arranged concentrically around, or radiate from, a central point in spherical symmetry, but have a monaxial heteropolar symmetry. Outer spine breadth decreases gradually distally, where one outer spine can be observed bearing poorly developed apophyses. Numerous conical by-spines (up to 35 μm long) emerge from the pore-bar junction and are occasionally pyramidal due to a short prolongation of the cortical spongy meshwork along them.
Etymology
From the polygonal shape of the innermost sphere.
Materials
Four specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
The discovery of a primitive tetrahedral initial spicule in lower Paleozoic forms is significant, and many species with a tetrahedral initial spicule previously have been reported from the Triassic (Dumitrică, Reference Dumitrică2013). The apparent long absence of this kind of spicule structure in the fossil record is curious and may be a result of convergent evolution. Currently, a comparison is difficult because it is unclear whether or not the microsphere of Triassic forms is homologous to the microsphere in lower Paleozoic species.
Westernbrookia polygonata n. sp. can be compared with late Tremadocian taxa assigned to the genus Westernbrookia reported from Canada. Westernbrookia polygonata n. sp. is most comparable to W. cancella Won, Iams, and Reed, Reference Won, Iams and Reed2007 (Won et al., Reference Won, Iams and Reed2007, p. 521, fig. 12.7–12.10). Westernbrookia cancella has a thick and delicate spongy layer (outer sphere Ø = 210 μm comparable to 180–198 μm in W. polygonata n. sp.), more than ten outer spines (11 in W. polygonata n. sp.), where one of the spines is associated with the pylome, which is also observed in W. polygonata n. sp. Westernbrookia cancella can be distinguished from W. polygonata n. sp. by having a more delicate internal spicular system. Furthermore, W. cancella has smaller pores on the outer sphere (~9 μm vs. 35 μm in W. polygonata n. sp.), a smaller pylome (Ø = 74–95 μm vs. ~50 μm in W. cancella), and shorter rod-like outer spines (up to 180 μm long vs. 110 μm in W. cancella).
The inner system is most closely comparable to that observed in W. diversa Won, Iams, and Reed, Reference Won, Iams and Reed2007 (Won et al., Reference Won, Iams and Reed2007, p. 519, figs. 15.1–15.12). However, it differs from W. diversa by its smaller spicular system (Ø = 56 μm vs. 90–115 μm in W. diversa), large, well-developed pylome (Ø = 74–95 μm vs. ~33 μm in W. diversa), greater number of outer spines (11 main spines vs. ≤ 8 outer spines in W. diversa). Westernbrookia polygonata n. sp. has a similar number of outer spines to W. ovata (>12), but differs in the smaller dimensions of its outer and inner sphere (outer sphere Ø = 180–198 μm, inner sphere Ø = 56 μm in W. polygonata n. sp. vs. ~270 μm and ~75 μm in W. ovata).
The inner system also resembles that of the Lower–Middle Ordovician (upper Floian–lower Dapingian) unidentified microsphere reported by Maletz and Bruton (Reference Maletz and Bruton2007, fig. 2.A) from the Valhallfonna Formation, Spitsbergen. The microspheres share a similar construction of thin pore-bars that form irregular or polygonal pore frames, rarely hexagonal framed pores, and are of a similar size (Ø = 52 μm in Maletz and Bruton, Reference Maletz and Bruton2007 vs. 56 μm in W. polygonata n. sp.). The two microspheres bear 11 long, thin radial bars, where one spine is located around the pylome. Furthermore, the microsphere in the form described by Maletz and Bruton (Reference Maletz and Bruton2007) also appears to be heteropolar, where a thin MB can be observed in the background of the SEM image. They differ as the rays from the Spitsbergen material taper and do not form an outer sphere.
Because our observations are based on only four specimens, we cannot gauge the true range of variability of this taxon, and it is difficult to know exactly what the diagnostic characters are. The taxon W. diversa is reported to have an internal structure that differs among specimens (Won et al., Reference Won, Iams and Reed2007, p. 519, figs. 15.1–15.12). From the four specimens observed, the inner spicular system appears to be stable.
Westernbrookia sp.
Figure 9.5, 9.6
Description
Well-developed outer (Ø = 145–155 μm) and inner spheres (Ø = 75 μm) are constructed by thin pore-bars that form irregular or polygonal pore frames. Pores on the inner sphere are angular to oval, with irregular size and arrangement. The outer sphere is thick and regularly latticed, with rounded to angular-oval pores (Ø ~5 μm), and bearing short and thin by-spines at vertices.
The inner sphere contains numerous polygonal, rarely hexagonal framed pores that bear numerous long, thin radial bars. Radial bars connect the microsphere to the outer sphere and continue as outer spines. The rod-like outer spines (up to 200 μm long) decrease gradually distally, where one outer spine was observed to bear poorly developed apophyses at various levels. An incomplete, loose additional sphere develops from the main spines.
Materials
Three specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Westernbrookia sp. has characteristics intermediate between the inaniguttids and the aspiculids.
Family Palaeospiculidae Won in Won and Below, Reference Won and Below1999
Remarks
Although the spicular system is generally cubic, in cases where all elements are well preserved, the skeleton has a spherical shape because of the density of the spinules (apophyses). Palaeospiculids that do not have a skeletal wall made up of fused spinules are readily identifiable. However, they are rare in the material studied and comprise <1% of the radiolarians counted. In contrast, palaeospiculids with a well-developed outer sphere constructed from fused spinules are abundant, comprising 19% of the radiolarians encountered. Won et al. (Reference Won, Iams and Reed2007) noted an evolutionary trend in the palaeospiculids—from an isolated spicular system (middle Cambrian) to party fused (middle Tremadocian) then completely fused (late Tremadocian) spheres. The presence of palaeospiculids, with or without spheres, indicates that archaic forms persist alongside their descendants in the middle Darriwilian.
Genus Neopalaeospiculum Won, Iams, and Reed, Reference Won, Iams and Reed2007
Type species
Neopalaeospiculum laxum Won, Iams, and Reed, Reference Won, Iams and Reed2007.
Remarks
The taxa from Newfoundland described herein have a six-rayed spicular system that appears to be derived from the genus Svalbardospiculum. Neopalaeospiculum differs from Svalbardospiculum in having a spherical skeletal wall made up of completely fused meshwork from third-order spinules. Previous researchers have emphasized the angular meshes as the most critical criterion for establishing different species of Polyentactinia, placing less emphasis on other characteristics. For genus-level classification, this has, over time, led to a heterogeneous taxon.
Holotype
UQSEES_M1S04, Figure 10.4, Supplemental Data file 4; sample PD09, Middle Ordovician, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.

Figure 10. (1–4, 6, 7) Compilation of μ-CT models, line drawings, and SEM image illustrating the complex spicular system and skeletal development of Neopalaeospiculum sp. and Neopalaeospiculum piccadilliensis n. sp. (1–3) μ-CT model of Neopalaeospiculum sp. (M1S04 from PD13) with digitally segmented spicular system at different orientations; (4) μ-CT model of Neopalaeospiculum piccadilliensis n. sp. (UQSEES_M1S04: holotype from PD09) with digitally segmented spicular system at different orientations; (6) diagram of Neopalaeospiculum sp. showing the relationship of the spicular system and main spines (ms); (7) diagram of Neopalaeospiculum piccadilliensis n. sp. showing the relationships among the initial spicule, inner spicular system, main spines, and outer sphere. (5) SEM image of the Lower Ordovician Palaeospiculum neofurcatum re-illustrated from Won et al., Reference Won, Iams and Reed2005 (fig. 10.18) with the inner spicular system highlighted. (1–5) are shown at the same scale.
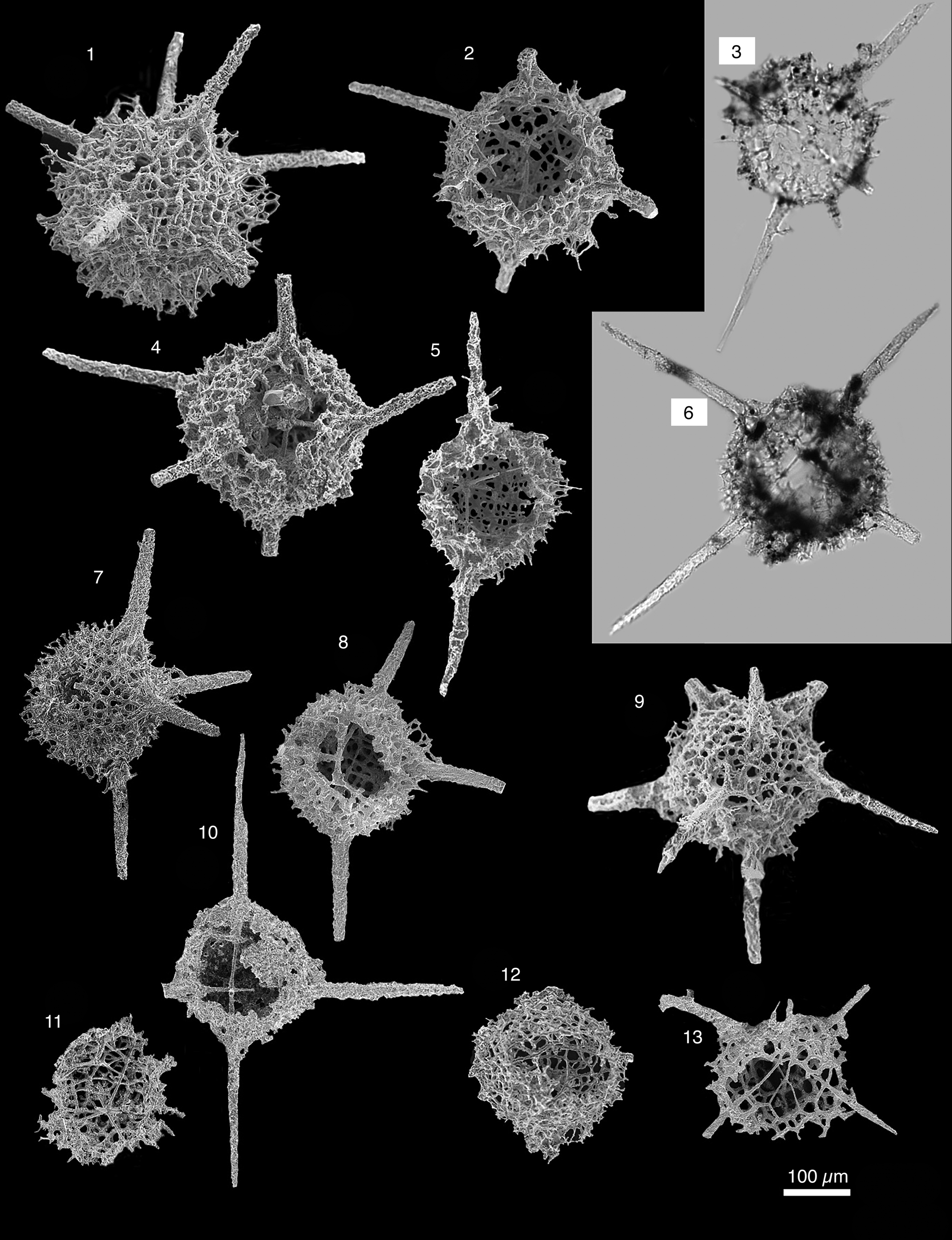
Figure 11. Radiolarians extracted from limestones from the Piccadilly Quarry, Newfoundland, Canada. Images are scanning electron micrographs, except (3, 6), which are transmitted-light images. Scale bar = 100 μm: (1, 2, 4, 6, 9) Neopalaeospiculum piccadilliensis n. sp.; (1) PD13; (2, 4, 9) PD09; (6) PD11; (3, 5, 7, 8, 10–13) Neopalaeospiculum sp.; (3, 5) PD13; (7, 8) PD09; (10–13) PD05.
Diagnosis
A single spherical outer sphere (Ø = 200–280 μm) displaying a latticed meshwork of interwoven bars bearing seven thick outer spines, which originate from the rays of an eccentrically positioned, point-centered spicule. The delicate internal framework consists of a point-centered initial spicule with six rays and two sets of whorls on each ray.
Occurrence
Middle Ordovician, Table Cove Formation, Newfoundland, Canada.
Description
The initial spicule is isometric where four first-order spinules arise from the rays at an equal distance (63 μm from point center to first set of whorls). The first-order spinules join with the adjacent spinules from another ray to form a cubic structure, where the initial spicule passes through the middle of the cube (Fig. 10.7). Two sets of whorls are rotated by 45°, whereas the other four sets of whorls produce spines parallel to the initial spicule. The second set of whorls on the rays occur at different lengths—15 μm, 21 μm, 38 μm, 39 μm, 46 μm, and 50 μm. This causes the initial spicule and cubic spicular system of the first set of whorls to be eccentrically positioned. From this second set of whorls on the main rays, a three-dimensional, compact meshwork of fused branches constructs the outer sphere. The corners of the inner cubic spicular system are also connected to the outer sphere to produce additional support. Some short spinules branch outward, freely on the meshwork. These short branches are partly fused and cover the surface. At the second set of whorls, the main rays immediately thicken to produce six of the seven main spines (Ø ~7 μm on the internal spicular system to ~28 μm on the main outer spines). Main spines (up to 260 μm long) maintain their thickness until they taper rapidly, distally. The seventh large main spine arises from the junction of the two first-order spinules that are rotated 45° to the initial spicule. The seventh main spine is generally the same width and length as the six main spines from the initial spicule, but can be distinguished because it is tangential to the other main spines. Main spines are rarely curved.
Etymology
Named for the Piccadilly Quarry locality from which it is described.
Materials
Eighty-eight specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Neopalaeospiculum piccadilliensis n. sp. is relatively common, comprising 10% of radiolarians counted. Neopalaeospiculum piccadilliensis n. sp. generally can be distinguished from Neopalaeospiculum sp. by its larger size (Ø = 200–280 μm vs. 180–195 μm) and the number of spines (seven vs. five).
Neopalaeospiculum piccadilliensis n. sp. has a six-rayed spicular system comparable to that of Svalbardospiculum sp. aff. S. hexaradiatum (Won and Iams, Reference Won and Iams2002) (Fig. 4.11, 4.12), with a point-centered spicule and two sets of whorls on each ray. The difference is that the skeleton develops for Svalbardospiculum sp. aff. S. hexaradiatum on the first set of whorls and not the second. Furthermore, the second set of whorls is short and terminal after the first-order. The inner spicular system also has an affinity to that of Neopalaeospiculum laxum Won, Iams, and Reed, Reference Won, Iams and Reed2007 (Won et al., Reference Won, Iams and Reed2007, fig. 5.4, 5.6) and Palaeospiculum neofurcatum Won, Iams, and Reed, Reference Won, Iams and Reed2007 (Won et al., Reference Won, Iams and Reed2005, fig. 10.17–10.19). Neopalaeospiculum laxum and P. neofurcatum commonly have five (rarely six) first-order spinules off two of the first sets of whorls. The material in this study further differs from P. neofurcatum in having a fused and well-developed outer skeleton.
Neopalaeospiculum sp.
Figures 10.1–10.3, 10.6, 11.3, 11.5, 11.7, 11.8, 11.10–11.13; Supplemental Data file 5
Description
A single spherical outer sphere (Ø = 200–280 μm) constructed from fused bars, bearing five outer spines. The skeleton is formed of a loose meshwork of thick straight and curved bars interwoven in a disorderly manner or fused. The five main spines originate from the rays of an eccentrically positioned spicule. A set of first-order spinules develop on four of the rays at an equal distance (70 μm from initial spicule). Main spines (up to 210 μm long) maintain their thickness until they taper rapidly, distally. One of the main spines is commonly curved.
Materials
One hundred specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
The initial spicule is developed from a six-rayed point spicule where one of the rays is reduced. Four of the rays are repositioned (at 149°), compensating for the reduction of the sixth spine.
Genus Ramuspiculum Won and Iams, Reference Won and Iams2002
Type species
Ramuspiculum multiramosum Won and Iams, Reference Won and Iams2002.
Remarks
The material examined in this study is tentatively assigned to the genus Ramuspiculum because of the presence of a very short microbar (6.6 μm long) and continuation of the primary rays, where the first-order whorls develop. Because the main rays are preserved at each whorl, the main skeletal framework of Ramuspiculum laxum n. sp. resembles Svalbardospiculum. However, the genus Svalbardospiculum preserves the hexactine skeleton (four spinules) with each whorl. Thus, taxa with a reduction in the spinules at the first level (three spinules; Fig. 12.3) are assigned to the genus Ramuspiculum. The microbar is noteworthy, and it joins two sets of three rays. If the specimen is orientated to view one set of the three rays, the initial spicular structure closely resembles the hexactine skeleton, and it is difficult to distinguish from taxa assigned to Svalbardospiculum. A slight off-set of the rays can be recognized rarely at this angle.

Figure 12. (1–5) Ramuspiculum laxum n. sp. (UQSEES_M1S03 holotype from PD09), compilation of images of μ-CT model and SEM images; the microbar is difficult to detect depending on the view angle; all images to the same scale; A = apical ray, B = basal ray. (1, 2) Initial spicules that develop into the main spines are digitally segmented (in green) from the outer meshwork for individual observation; (1) μ-CT model orientated and (4) (re-oriented around a vertical axis) to illustrate how the false impression of the existence of point-centered spicule arises, when instead there is a microbar; (3) close-up of the two apical rays to demonstrate the outer sphere construction of whorls, by first- and second-order spinules; (5) SEM image of Ramuspiculum laxum n. sp. from PD09. (6) SEM image of Svalbardospiculum sp. aff. S. hexaradiatum (Won and Iams) from PD02.
Ramuspiculum laxum new species
Figures 3.1, 3.2, 6.2, 6.4, 6.5, 12.1–12.5; Supplemental Data file 6 μ-CT video file M1S03
- aff. Reference Pouille, Danelian and Popov2014
Polyentactinia sp. cf. P. spinulenta Pouille and Danelian in Pouille et al., 2014 (Pouille et al., Reference Pouille, Danelian and Popov2014b)
Holotype
UQSEES_M1S03, Figures 3.1, 3.2, 12.1–12.5; Supplemental Data file 6; sample PD09, Middle Ordovician, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Diagnosis
A large, but poorly developed, spicular sphere (Ø = 220–300 μm) formed from a complex structure of numerous branching spinules. An eccentrically placed, bar-centered, six-rayed spicule is positioned just within the skeletal wall. From the six rays, several generations of branching spinules (apophyses) can be observed.
Occurrence
Middle Ordovician, Table Cove Formation, Piccadilly, Newfoundland, Canada.
Description
The branching occurs at distinct intervals, mostly 84 μm on the basal rays and 63 μm on the apical rays, where whorls of rays are produced. Second- and third-order branching spinules are more irregularly placed. Three spinules (perpendicular to the ray at 120° on the same plane) of the first-order arise on all of the rays, where the main rays are preserved and continue outside the outer sphere as main spines. First-order spinules are prominently developed and are of equal diameter on the basal rays (Ø = 11.3 μm). The first-order spinules that branch from the apical rays are thinner (Ø = 7.5 μm) and terminate. The surface of the rays between the central point and first spinules is commonly smooth. Microbar clearly shown in μ-CT mode (Figs. 3.1, 3.2, 12).
Etymology
From the Latin laxus, meaning loose.
Materials
Five specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Our material has affinity to the upper Darriwilian form Polyentactinia sp. cf. P. spinulenta Pouille and Danelian described from the Shundy Formation, Kazakhstan in Pouille et al. (Reference Pouille, Danelian and Popov2014b, pl. 2, figs. 9, 10). They noted the presence of thick main spines, a small MB (~8 μm) and the development of whorled rays (three rays on the first-order). Material described in Pouille et al. (Reference Pouille, Danelian and Popov2014b) differs by having a slightly larger outer sphere (Ø = 220–300 μm vs. 330–365 μm for Polyentactinia sp. cf. P. spinulenta) and a thicker outer sphere made from a partly or completely fused meshwork. It is possible that the material reported in Pouille et al. (Reference Pouille, Danelian and Popov2014b) evolved from Ramuspiculum sp. (as described herein).
Ramuspiculum sp.
Figure 6.10, 6.11
Description
Small globular skeleton (Ø = 170–205 μm) developed from repeated branching of the initial spicule. Ectopically placed, bar-centered spicule with three rays at each end. The microbar is very short (~5 μm long) and can be distinguished by the slightly offset main rays.
Materials
Two specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Ramuspiculum sp. differs from Ramuspiculum laxum n. sp. in its smaller size and less well-defined spicular structure.
Genus Svalbardospiculum Maletz and Bruton, Reference Maletz and Bruton2007
Type species
Svalbardospiculum arenigium Maletz and Bruton, Reference Maletz and Bruton2007.
Remarks
The main difference between Svalbardospiculum and Palaeospiculum is in the development of higher generations of whorled rays on the elongated primary rays in the genus Svalbardospiculum (Maletz and Bruton, Reference Maletz and Bruton2007). Previously, Svalbardospiculum had been recovered only from the Lower Ordovician (lower Tremadocian) in Newfoundland (Won and Iams, Reference Won and Iams2002; Won et al., Reference Won, Iams and Reed2005; Pouille et al., Reference Pouille, Danelian and Maletz2014a), and the Lower–Middle Ordovician (upper Floian–lower Dapingian) of Spitsbergen (Maletz and Bruton, Reference Maletz and Bruton2007). However, we can confirm the presence of this genus in the Middle Ordovician (middle Darriwilian).
Svalbardospiculum multifurcatum (Won, Iams, and Reed, Reference Won, Iams and Reed2005)
Figure 4.1, 4.2
- Reference Won, Iams and Reed2005
Palaeospiculum multifurcatum Won et al., p. 451, fig. 11.1–11.14.
- Reference Pouille, Danelian and Maletz2014a
Paleospiculum multifurcatum Pouille et al., pl. 4, fig. 1.
Holotype
Specimen (GSC-125580 GP26A-40), Cow Head Group, Green Point, Canada (Won et al., Reference Won, Iams and Reed2005 p. 451, fig. 11.11–11.14).
Occurrence
Lower to Middle Ordovician, Cow Head Group, Newfoundland, Canada.
Description
A small spicular sphere (Ø = 110 μm) constructed from a hexactine skeleton. The point-centered spicule is eccentrically placed and has six rays and spinules on all of the rays. Four spinules of the first-order arise on all of the rays (~47 μm on the basal rays and ~30 μm on the apical rays). Each ray can be divided into apical and basal rays, with more spinules present on the basal rays than on the apical rays. Second-order spinules may or may not develop.
Materials
Five specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Maletz and Bruton (Reference Maletz and Bruton2007) reassigned Palaeospiculum multifurcatum and P. hexaradiatum, described by Won et al. (Reference Won, Iams and Reed2005), to their new genus Svalbardospiculum based on the observation of several generations of whorled rays. Multiple generations of whorled rays are easily recognized. The material in this study is identical to the lowermost Tremadocian form Palaeospiculum multifurcatum (misspelled as Paleospiculum) from the Green Point section, Newfoundland, illustrated by Pouille et al. (Reference Pouille, Danelian and Maletz2014a, pl.4, fig. 1), but we follow the reassignment of Maletz and Bruton (Reference Maletz and Bruton2007). This discovery extends the known range of Svalbardospiculum multifurcatum to the mid-Darriwilian.
Svalbardospiculum sp. aff. S. hexaradiatum (Won and Iams, Reference Won and Iams2002)
Figures 4.11, 4.12, 12.6
- Reference Won and Iams2002
aff. Palaeospiculum hexaradiatum Won and Iams, p. 28, fig. 3.9–3.11.
Description
Polar, point-centered, six-rayed spicule with multiple generations of hexactine spinules. The main rays that project from the initial spicule can reach lengths of up to 210 μm from the center. Radially arranged first- and second-order spinules on each ray intermeshing (never fused) to form a roughly spherical skeleton (Ø = 220–250 μm). Branching occurs at distinct distances, mostly ~63 μm on the basal rays and 58 μm on the apical rays, where whorls of rays are produced. A second set of first-order spinules, slightly variable in length (62–73 μm from first set), arise on the basal rays, but terminate and do not develop into a second sphere. The basal rays are equal in length and are slightly longer than the apical rays. The number of spinules is variable on each ray, and spinules on the apical rays are generally less well developed than on the basal rays. The surface of the rays between the central point and first spinules is commonly smooth.
Materials
Five specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Specimens in this study show affinity with the skeletal structure found in the upper Cambrian Svalbardospiculum hexaradiatum (Won and Iams, Reference Won and Iams2002, figs. 3.9–3.11). Our material differs from S. hexaradiatum by its larger size (outer sphere Ø = 220–250 μm vs. 200 μm in S. hexaradiatum) and longer spines (up to 210 μm long from the initial spicule vs. 165 μm in S. hexaradiatum).
In samples from the upper Cambrian, Won and Iams (Reference Won and Iams2002) observed an additional set of first-order spinules (found distally on the rays), which was commonly imperfectly preserved or absent because of its weak development. In the samples from this study, this second set of first-order spinules is well developed, but they are generally thin and short (up to 30 μm long) and terminal compared to the first set of whorls.
Family Protoentactiniidae Kozur, Mostler, and Repetski, Reference Kozur, Mostler and Repetski1996
Genus Protoentactinia Kozur, Mostler, and Repetski, Reference Kozur, Mostler and Repetski1996
Type species
Protoentactinia gracilispinosa Kozur, Mostler, and Repetski, Reference Kozur, Mostler and Repetski1996.
Protoentactinia sp.
Figure 6.1
Description
Six-rayed, bar-centered (MB = 74 μm long) spicule. The loose spongy skeleton formed by repeated branching of the initial spicule develops a loose sub-globular shape (Ø = 280 μm). Repeated branching occurs at a consistent length (~90 μm), and the width of the branch decreases in diameter at each level. The first-order spinules branch into four spinules that taper and commonly terminate (60 μm long).
Materials
Two specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Only two specimens were identified, making assessment of the 3D aspect of the skeleton and assignment of the specimen to species level challenging.
Order Entactinaria Kozur and Mostler, Reference Kozur and Mostler1982
Remarks
De Wever et al. (Reference De Wever, Dumitrică, Caulet and Caridroit2001) elevated the Suborder Entactinaria Kozur and Mostler to Order status with the definition: Polycystina having as initial skeleton a spicule with a variable number of spines arising from the two ends of a MB or from a center. Spines equal or unequal, disposed in variable patterns, and usually separated into an apical and a basal group. A well-developed outer sphere is usually present, latticed or spongy, or a combination of both; when multiple, it is usually concentrically disposed, never along an axis. MB or spicule center located inside the innermost sphere, or quite eccentric in its wall.
Family Entactiniidae Riedel, Reference Riedel1967, emend. Won, Reference Won1997
Description
The spicule is directly connected to the outer sphere or to a microsphere (sometimes described as a macrosphere; Dumitrică, Reference Dumitrică2013), when this exists. The outer sphere is constructed from apophyses to form a three-dimensionally spherical skeleton. The absence or presence of the inner sphere is always constant at the genus and species levels. The ends of the radial beams are connected to each other by apophyses, which form the internal or basal layer of the spongy outer sphere.
Remarks
The main outer spines are normally the continuations of the rays of an internal spicule, and their number and orientation are commonly constant at the species level, but variable at the genus level. Although entactiniids are one of the best-known groups, their detailed systematic description is far from complete.
Genus Spongentactinia Nazarov, Reference Nazarov1975
Type species
Spongentactinia fungosa Nazarov, Reference Nazarov1975.
Remarks
The material consists of latticed microspheres that include a six-rayed median, bar-centered spicule. A densely spongy outer sphere originates from branches at various levels along each ray. A thick spongy layer is developed on top of a weakly developed, latticed basal layer, and is separated from the microsphere.
Spongentactinia armillata (Nazarov in Nazarov and Popov, Reference Nazarov and Popov1980)
Figures 3.9, 3.12, 13, 14.8, 14.12, 14.14, 14.15; Supplemental Data file 7
- aff. Reference Fortey and Holdsworth1971
Entactinia Fortey and Holdsworth, pl. 10, fig. 4.
- Reference Nazarov and Popov1980
Haplentactinia armillata Nazarov in Nazarov and Popov, p. 55, pl. 16, figs. 9, 10.
- Reference Nazarov1988
Haplentactinia armillata; Nazarov, pl. 9, fig. 12.
- Reference Aitchison1998
Haplentactinia armillata; Aitchison, p. 79, pl. 2, fig. U.
- Reference Perera, Aitchison and Nothdurft2020
Haplentactinia armillata; Perera et al., p. 3, fig. 4Gg, j.
Holotype
Haplentactinia armillata Nazarov, Specimen No. GIN 4333/58 in the Geological Institute, Moscow. Lower–Middle(?) Ordovician, Arenigian–Llanvirnian(?), Kogashsky Horizon, Kysylzharskaya Suite, Central Kazakhstan (Nazarov and Popov, Reference Nazarov and Popov1980 p. 55, pl. 16, figs. 9, 10).
Occurrence
Ordovician, upper Dapingian, Akzhal Mountains, Kazakhstan (Nazarov and Popov, Reference Nazarov and Popov1980); Lower Ordovician, Spitsbergen (Fortey and Holdsworth, Reference Fortey and Holdsworth1971); Lower Ordovician, Ballantrae Complex, Scotland (Aitchison, Reference Aitchison1998); Middle Ordovician, Southern Uplands terrane, Hawkwood Burn, Scotland (Perera et al., Reference Perera, Aitchison and Nothdurft2020).
Description
Two distinct layers including an inner latticed layer with polygonal pores, containing a six-rayed median, bar-centered spicule and a thickly developed spongy layer. The outer surface (Ø = 310–390 μm) is loosely spherical without definite outward termination of the lateral branches. A doubled-layered outer sphere constructed of irregularly arranged fused bars that form a poorly developed basal layer. This meshwork is ~100 μm thick and is developed from three to four apophyses on the main rays. Six long spines (up to 260 μm long) originate from the six-rays of the bar-centered spicule. Between the second sphere and the periphery of the spongy meshwork, the spines are either relatively narrow or increase gradually in breadth distally (4–7 μm). Beyond the spongy meshwork, the spines increase greatly in breadth (10–37 μm) and retain this breadth until they taper to a point, distally. Spines can be straight or slightly curved, and commonly branch distally to produce a prominent bifurcated spine. This bifurcation is not always present, but when it is, it is only developed on one, or rarely two, of the six spines.
The MB is eccentrically positioned within a sack-like or ovoid heteropolar microsphere (Fig. 13). The initial spicule consists of a MB, two apical rays, and four basal rays. Rays are interconnected by bars and arches to the microsphere, where one set of basal rays are connected twice to the microsphere. The length of the MB is 33.8 μm. The size of the microsphere is 89.2 μm wide, 130 μm long, and 88.6 μm deep. However, due to the irregular shape of the microsphere it cannot always be accurately measured on broken specimens.
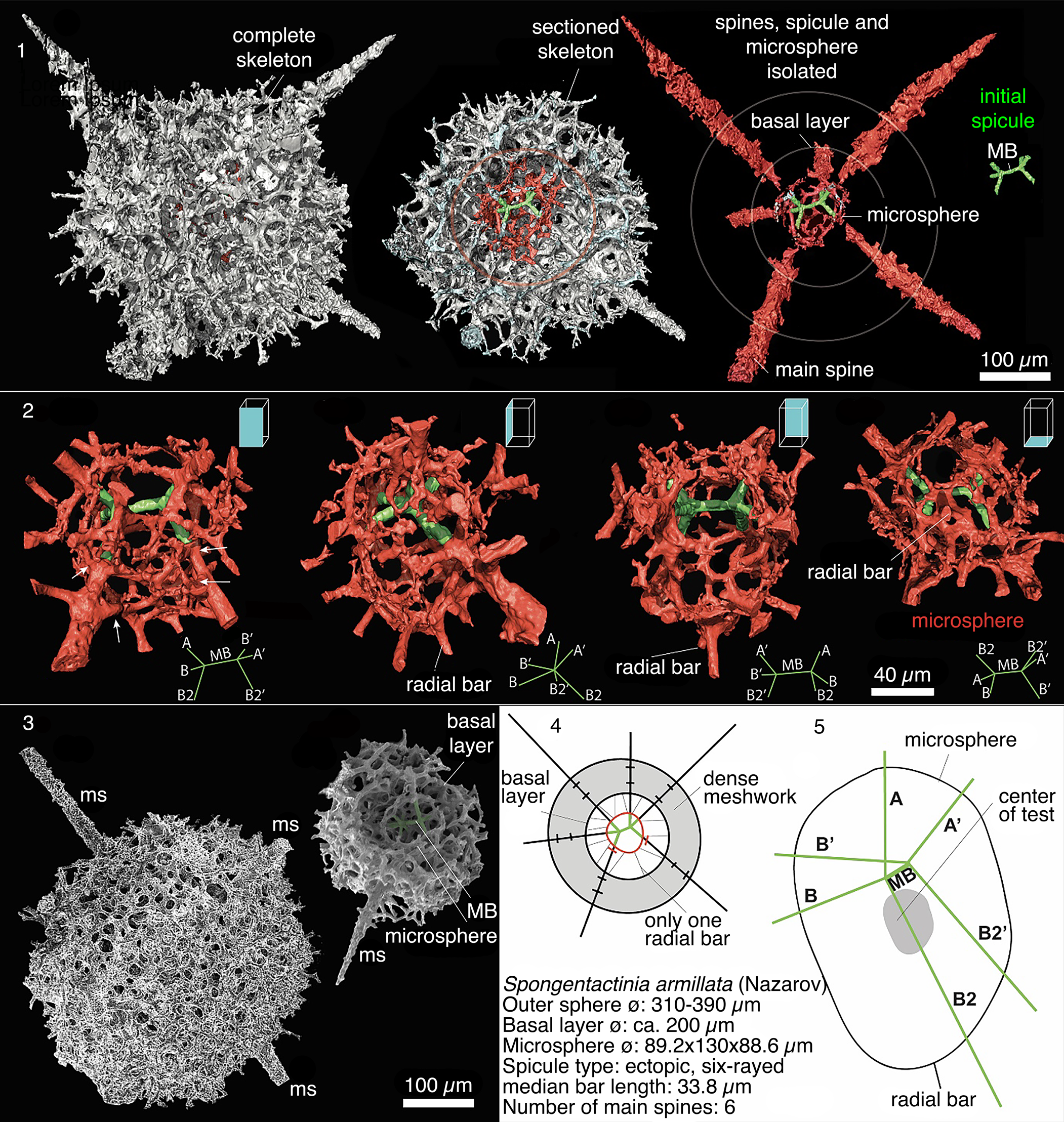
Figure 13. Compilation of μ-CT models (M3S09 from sample PD13), SEM images, and schematic drawings of Spongentactinia armillata (Nazarov). (1) Outer meshwork, microsphere, initial spicule, and main spines (ms) digitally segmented for individual observation; each segment to the same scale; (2) microsphere, radial bars, and initial spicule at four different orientations; A, B, B’, B2, B2’ are arbitrary labels for spines to allow viewers to better follow specimen rotation; (3) SEM images; MB = median bar; (left) Spongentactinia armillata (Nazarov) (from sample PD13); (right) Spongentactinia sp. (from sample PD05); (4) Diagram showing configuration of spheres, spines, and initial spicule; (5) schematic diagram of the microsphere.
The microsphere is complexly interwoven and is separated from the outer meshwork by a 60–75 μm gap. Microsphere is connected to the outer sphere by nineteen rays. The rays are all forked such that the total number of rays that join the outer sphere is 38. These rays anastomose to form a basal layer. The rays arising from the microsphere are randomly directed projections, except one bar that is radially orientated towards the center of the microsphere located at the opposite position to the initial spicule. Only the prominent rays from initial spicule continue outside the outer cortical layer into the main spines.
Materials
Twenty-three specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Twenty-three specimens identified as Spongentactinia armillata were examined in this study using both SEM and TLM, but internal features could only be observed using μ-CT scanning (M3S09 from sample PD13). Spongentactinia armillata has only been documented once from the Middle Ordovician (Nazarov and Popov, Reference Nazarov and Popov1980). Previously, the skeletal development has not been studied in any detail. Nazarov and Popov (Reference Nazarov and Popov1980) originally placed Spongentactinia armillata with Haplentactinia because the thick spongy layer obstructed observation of a microsphere. Observations with the μ-CT reveal important internal features that indicate assignment of this taxon to the genus Spongentactinia is more appropriate than to Haplentactinia. The basal layer is poorly developed as a result of the numerous rays originating on the microsphere to support the large test. Since the rays from the initial spicule project to the outer sphere, the orientation of the main spines can confidently be used for identification this taxon.
Spongentactinia nazarovi new species
Figures 9.14–9.17, 14.7, 14.10, 14.13
Holotype
Spongentactinia nazarovi UQSEES_2015PD05_01, Figure 9.16; from sample PD05, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Diagnosis
Two distinct layers, an inner latticed layer with polygonal pores, containing a six- to eight-rayed, bar-centered spicule and a thickly developed spongy layer.
Occurrence
Middle Ordovician, Table Cove Formation, Newfoundland, Canada.
Description
The outer sphere surface (Ø = 200–285 μm) consists of irregularly arranged and fused bars to form densely packed meshwork. Doubled-layered outer sphere is roughly spherical, but varies in thickness from 10–55 μm. The outer sphere originates from three apophyses on the main rays, where the most-proximal level develops into a well-defined latticed basal layer.
Five to six main spines (up to 180 μm long; e.g., Figs. 9.14, 14.7, 14.13) originate from eccentrically positioned spicule (colored light green on Fig. 9.16, 9.17) within a spherical-ellipse microsphere. The thickest part of the meshwork in the outer sphere correlates with the opposite position of the eccentrical spicule. The microsphere is separated from the outer meshwork by a 28–46 μm gap where only a few bars connect the two parts. The length of the MB (~30 μm long) and size of the microsphere (Ø ~80–90 μm) is constant. Three rays arise from one end of the MB, where all three develop into a main spine. Five spines arise from the other end, where only two or three spines develop into main spines. No bifurcation was observed on the distal end of the outer spine.
Etymology
Named in honor of Boris Nazarov for his significant contributions to Ordovician radiolarian studies.
Materials
Thirty-one specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
The apophyses arising from the radial beams seem to play a role in forming the outer sphere. They are better developed in Spongentactinia armillata than in Spongentactinia nazarovi n. sp., which has a MB and microsphere of comparable dimensions. The number of rays arising from the microsphere of Spongentactinia nazarovi n. sp. are reduced, and they are weaker and commonly not connected to the outer sphere. The irregular shape of the microsphere seen in Spongentactinia armillata could not be confirmed in Spongentactinia nazarovi n. sp. This may be an artefact of SEM imaging of sectioned radiolarians, which provides only a unilateral view.
The skeletal structure of Spongentactinia nazarovi n. sp. closely resembles that of S. armillata, but is clearly distinguished by its smaller size (outer sphere surface Ø = 200–285 vs. Ø = 310–390 μm in S. armillata; length of the main spines (up to 180 μm vs. 260 μm long in S. armillata). This form is similar to Spongentactinia armillata (Nazarov), but also differs in several respects: the size of the gap between microsphere and outer meshwork (~50 vs. ~60–75 μm gap in S. armillata); reduction of rays arising from the microsphere; and the number of rays on the initial spicule (eight vs. six in S. armillata). It is unclear whether the thickness of the spongy skeletal wall is related to ontogeny or some ecologic factor because S. armillata and Spongentactinia nazarovi n. sp. possess many internal differences.
Entactiniids gen. and sp. indet.
Remarks
Numerous other entactiniid specimens occur in samples from the Table Cove Formation. Their outer appearance is similar to that of entactiniids that have a thick spongy skeleton. The skeleton is mostly filled by three-dimensional meshes, where numerous spines originate from a complex inner polyhedron. Due to the difficulties observing the internal structure, it is not possible to assign them to any particular genus with certainty, therefore they are placed in open nomenclature herein.
Description
Forms characterized by large, slightly polygonal, lattice skeletons (Ø = 300–350 μm). Pores commonly larger (Ø = 10–28 μm) with more angular outline, framed by four to six lattice bars. Occasionally some of the pores become rounded and the skeletal structure appears slightly spongy. A three-dimensional meshwork of interwoven bars that loosely constructed three to four concentric layers. Layers formed by distal branching of beams arising from previous layer, of 11, curved at an angle to the layers, branched, or linked laterally. Numerous external, rodlike by-spines arise from incomplete interlayer beams (up to 100 μm long). Latticed internal polyhedron at the center of the skeleton, bearing six to eight rod-like beams. The distal parts of these beams commonly protrude as massive, rodlike outer spines. Main spines may exceed double the length of outer sphere diameter (up to 240 μm long). Commonly one spine is longer than the other and may possess a large, well-developed bifurcation.
Materials
Sixty specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Entactiniid gen. and sp. indet. A differs from B by its robust skeleton and its well-developed bifurcated main spine. Furthermore, the outer sphere is generally slightly smaller than that of B (Ø = 300–350 μm for A and 300–440 μm for B). Importantly, the radial rays do not originate from a central point or spicule, as seen in Spongentactinia armillata. Entactiniid gen. and sp. indet. A mimics the bifurcated spine of Spongentactinia armillata, but the internal framework upon which genus level assignment is based is radically different. It needs to be stressed that external morphological features, such as the angular mesh and large bifurcated spine, are not the most critical criteria for genus- or species-level identification. See remarks for S. armillata for further species comparison.
Entactiniid gen. and sp. indet. B
Figures 14.3–14.6, 14.11, 15.2, 15.3
Description
Numerous radial spines that are the basis of a multilayered, sub-spherical skeleton (outer sphere Ø = 300–440 μm). The robustness of the spines is variable; where the rays of the spicule and the radial components (radial rays) of the tissue are thicker toward the outside and protrude out of the skeleton surface, they appear as outer spines. Analysis with the μ-CT reveals that the inner structure has no MB and no R-frame structure. Spines originate from a complex internal polyhedron (Fig. 15.2, 15.3). Meshwork is very loose and produces a quadrangular or triangular pore pattern. Beams arise radially at many levels at regular and short distances from the inner structure. The beams join one another and with those of other rays, and some of the beams can become radial rays.
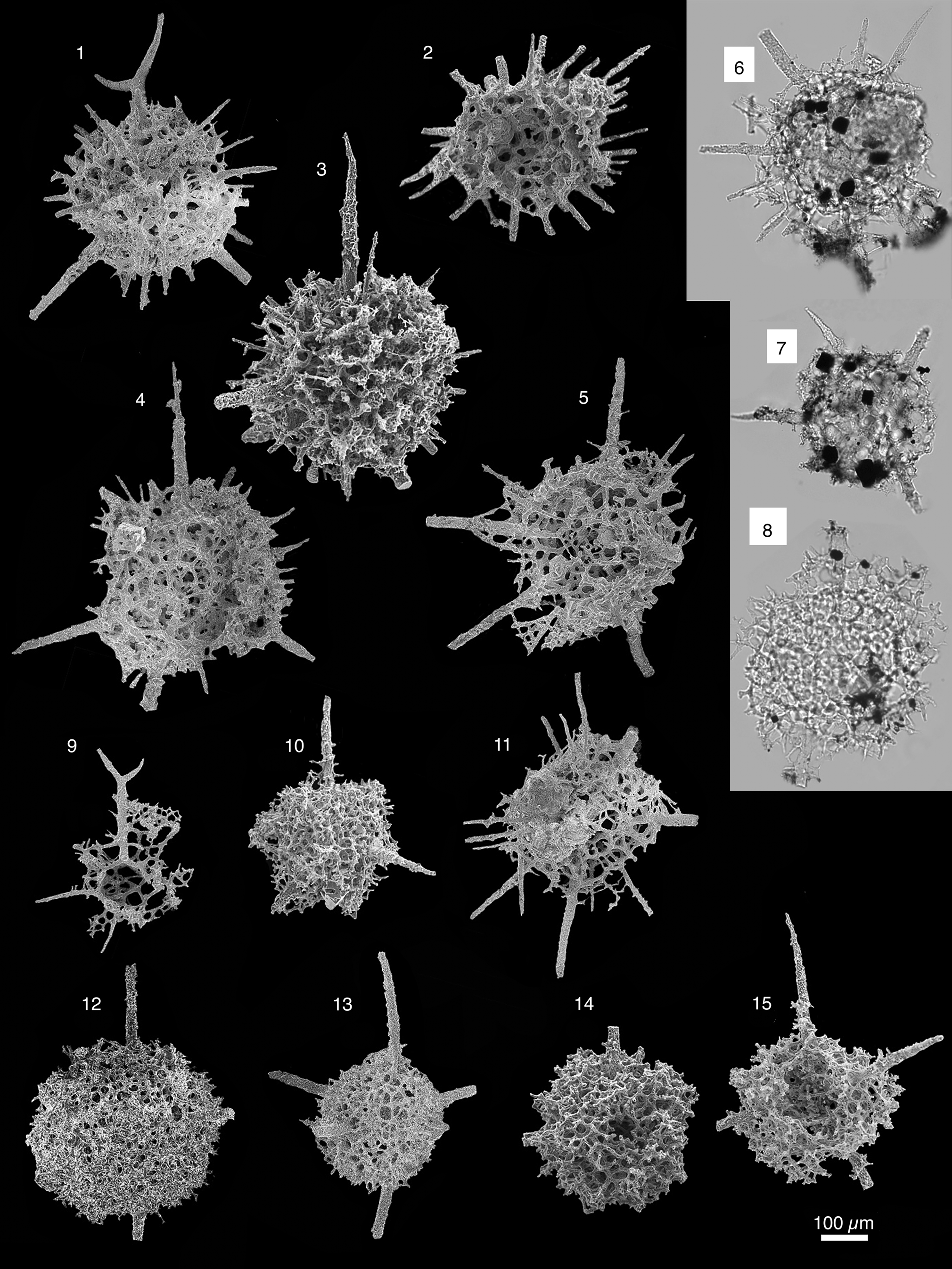
Figure 14. Radiolarians extracted from limestones from the Piccadilly Quarry, Newfoundland, Canada. Images are scanning electron micrographs, except (6–8), which are transmitted-light images. Scale bar = 100 μm. (3–6, 11) Entactiniid gen. and sp. indet. B; (11) PD09; (3–6) PD13. (1, 2, 9) Entactiniid gen. and sp. indet. A; PD13. (7, 10, 13) Spongentactinia nazarovi n. sp.; (7, 13) PD09; (10) PD13. (8, 12, 14, 15) Spongentactinia armillata (Nazarov) PD13.

Figure 15. Micro-CT models of Entactiniid gen. and sp. indet. A and Entactiniid gen. and sp. indet. B. In both models the inner skeleton is obstructed by the growth of a large calcite crystal (false colored purple to aid identification). (1) Entactiniid gen. and sp. indet. A (M2S05); (2) Entactiniid gen. and sp. indet. B (M2S06); (3) Entactiniid gen. and sp. indet. B, (M2S08) digitally dissected in various orientations.
Materials
Fifty-eight specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Entactiniid gen. and sp. indet. B is distinguished by its unusual pore pattern and partly angular silhouette. See remarks above for the comparison between Entactiniid gen. and sp. indet. A and B.
Entactiniid gen. and sp. indet. C
Figure 4.13–4.16
Description
Entactiniids with an open-latticed outer shell. Six-rayed spicule is visible inside many of the specimens. Up to six short robust main spines, which commonly exhibit bifurcated or even trifurcated terminations.
Materials
Forty-four specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
This form is characterized by the bifurcated or trifurcated terminations of main spines.
Order Incertae Sedis
Family Aspiculidae Won, Iams, and Reed, Reference Won, Iams and Reed2005
- Reference Won, Iams and Reed2005
Aspiculumidae Won, Iams, and Reed, p. 438 (nom. correct. Maletz, Reference Maletz2011, p. 130).
Remarks
Emended from Won, Iams, and Reed (Reference Won, Iams and Reed2005) to incorporate skeletons consisting of two encapsulated, polygonal spheres. Inner sphere constructed from spicular system of fused bars. An additional irregular outer sphere and pylome can develop in some specimens. Radial main spines.
Currently, the variability of the inner spicular system among aspiculid genera is difficult to understand and compare with other Paleozoic radiolarians. Observations in this study indicate a high degree of variability in the dimensions and geometry of the outer sphere and inner spicular system (Fig. 7). However, all genera appear to be united by the mode of construction of their skeletal features. That is, a construction of pore-bars with rounded cross-sections and slightly polygonal outlines, radial spines, and a three-dimensional mesh between the outer spines. Because the genus Kalimnasphaera shares similar features, we consider it may belong with the aspiculids.
Genus Aspiculum Won, Iams, and Reed, Reference Won, Iams and Reed2005
Type species
Aspiculum eccentricum Won, Iams, and Reed, Reference Won, Iams and Reed2005.
Aspiculum irregulare new species
Figures 7.10, 7.11, 9.1, 9.10, 9.13
Holotype
Aspiculum irregulare, UQSEES_2015PD13_01, Figure 9.10; sample PD13, Middle Ordovician, middle Darriwilian, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Diagnosis
A small but distinctive sphere (Ø = 107–146 μm) that is rather coarse and irregular in outline. The outer sphere has polygonal pores, which is formed from straight bars. Up to 23 thick, rod-like main spines, where a number of them extend into the outer sphere, indicating the presence of an inner sphere.
Occurrence
Middle Ordovician, Table Cove Formation, Newfoundland, Canada.
Description
Spines are evenly dispersed over the skeleton and variably developed (up to 167 μm long; average 100 μm long). Outer spines produce apophyses at various distances. These apophyses connect to adjacent spines through a highly irregular, thick, but loose three-dimensional mesh.
Etymology
From the Latin irregularis, for its irregular, non-spherical inner sphere.
Materials
Three specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
The nature of the inner structure is not preserved, however the irregular sphere and spiraliform additional sphere are distinct from other aspiculids.
Aspiculum sp. A
Figure 9.2, 9.4
Description
The outermost part of the skeleton (Ø = 116–200 μm) is a two-dimensional spherical surface composed of a delicate angular and irregular meshwork. The innermost part is eccentrically placed. Poorly developed basal layer. Because radial elements protrude through the outer wall, the outermost layer is locally indistinct.
Materials
Two specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Easily distinguished by its loose labyrinthine skeleton. An internal spicule was not observed in the material in this study due to obstruction by the outer sphere. Similar to Polyentactinia polygonia Foreman, Reference Foreman1963 where the skeleton is formed from very loose and angular long bars. Polyentactinia polygonia differs in having a bar-centered spicule with six or eight rays, from which arise spinules that branch several times and anastomose.
Aspiculum sp. B
Figure 9.3, 9.7, 9.8
Description
Delicate three-dimensionally interwoven skeleton constructed of thin bars (Ø ~210 μm). Poorly developed basal layer. Spines do not meet at a central point.
Materials
Three specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Difficult to assess the complex nature of the inner structure.
Genus Nyfrieslandia Maletz and Bruton, Reference Maletz and Bruton2007
Type species
Nyfrieslandia sphaeroidea Maletz and Bruton, Reference Maletz and Bruton2007.
Remarks
Emended to include a spicular system that constructs a polygonal inner sphere, where numerous radial spines originate and project as main spines on the outer sphere. The outer sphere is well developed, but highly variable in both size and shape. An additional irregular outer sphere is developed from irregularly placed apophyses on the main spines.
Limited information on the nature of the innermost sphere is available due to a high degree of infilling in the μ-CT scan. However, the inner sphere is produced largely from a spicular system of fused bars. This inner sphere can be either polygonal or spherical, based on how many pores are present. The outer sphere can closely resemble the outer spheres of inaniguttids. However, the genus Nyfrieslandia generally can be distinguished by the polygonal outline of the large hollow inner sphere (Ø ~90–100 μm for Nyfrieslandia; generally, <60 μm in the inaniguttids).
Maletz and Bruton (Reference Maletz and Bruton2007) suggested potential assignment to the order Spumellaria. However, the material in this study suggests a spicular system analogous to that in the order Entactinaria has developed. Furthermore, Nyfrieslandia has a similar mode of skeletal construction to that of Kalimnasphaera, which also has an uncertain high-level taxonomic assignment. We suggest these two groups may have a close relationship.
Nyfrieslandia sp. aff. N. complicata (Maletz and Bruton, Reference Maletz and Bruton2007)
Figures 7.1, 7.4, 7.7, 16, 17.4, 17.5, 17.7, 17.8, 17.16 Supplemental Data file 8
- aff. Reference Maletz and Bruton2007
Nyfrieslandia complicata Maletz and Bruton, p. 281, figs. 26A–O.
- aff. Reference Won and Iams2013
Nyfrieslandia crassispinosa Won and Iams, fig. 6K.
Description
Well-developed outer sphere (Ø = 183–200 μm) and an extremely thin and hollow inner sphere (Ø = 90–100 μm), formed from rounded bars. Both spheres are developed from interconnected round bars that develop a coarse mesh with irregular pores. The inner sphere is polygonal and formed from straight bars. Numerous thin, radiating spines that originate on the inner sphere are variable in length (60–110 μm). Spines are evenly dispersed at ~42° to neighboring spines. The numbers and positions of by-spines are highly variable in the material in this study, but generally ~22–39 spines per specimen are present. One specimen has a thinner outer sphere and spines, where the spines have a maximum length of only 99 μm (Fig. 17.5). The outer spines are variably developed and are considerably reduced in a few specimens. Outer spines produce apophyses at various distances. These apophyses connect to adjacent spines through a highly irregular, loose, three-dimensional mesh. No complete additional outer sphere was observed (Figs. 16, 17.16).

Figure 16. Series of images for μ-CT model UQSEES_M5S16 of Nyfrieslandia sp. aff. N. complicata Maletz and Bruton; outer shell is colored yellow; inner sphere (red) is obstructed by the growth of a large calcite crystal (false colored purple to aid identification). (1, 3) Scale bar = 100 μm; (2) inner shell with outer shell digitally removed for clarity, scale bar = 50 μm; (3) inner sphere rotated 180° relative to (2).
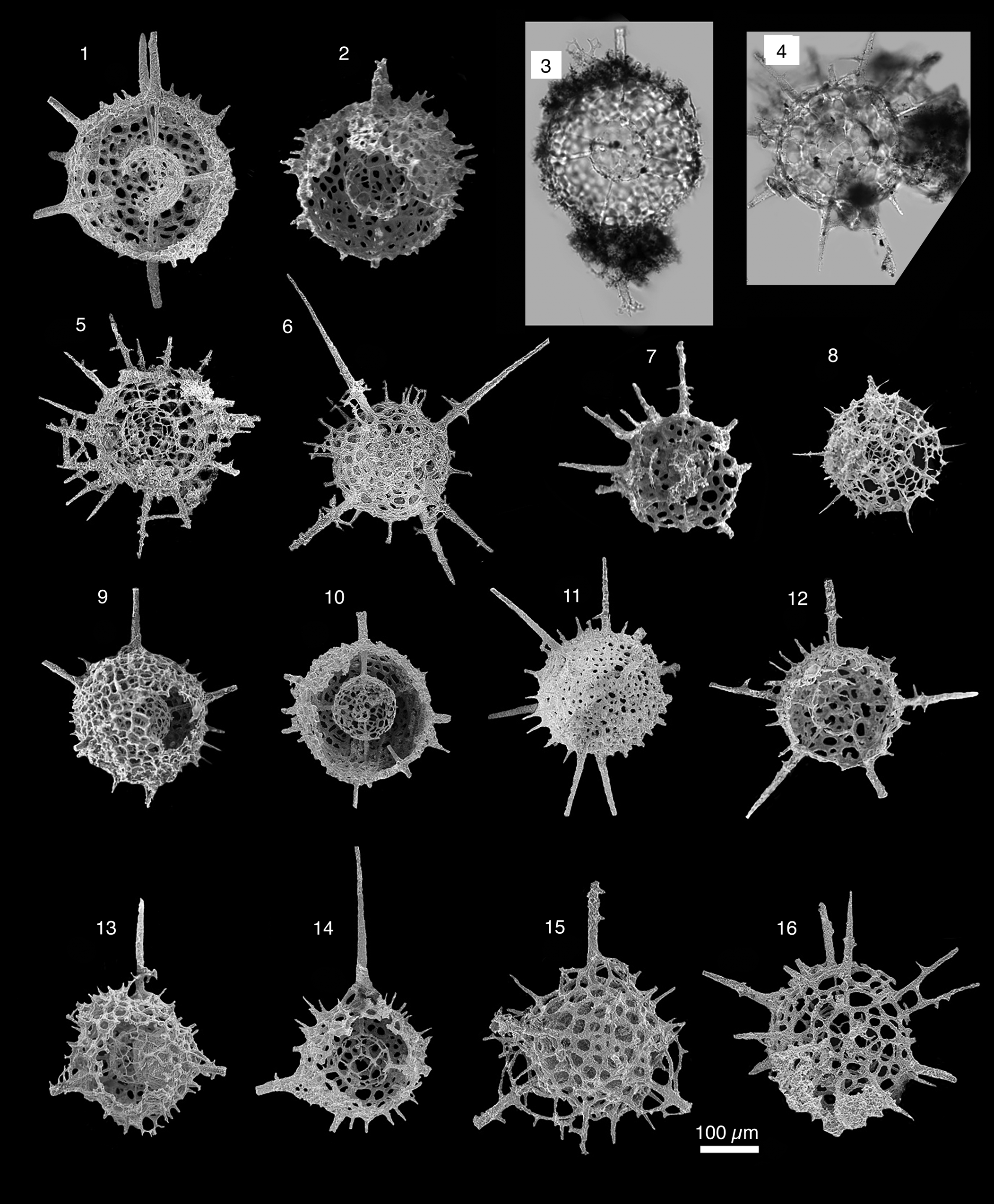
Figure 17. Radiolarians extracted from limestones from the Piccadilly Quarry, Newfoundland, Canada. Images are scanning electron micrographs, except (3, 4), which are transmitted-light images. Scale bar = 100 μm. (1–3) Nyfrieslandia sp. B.; PD13; (4, 5, 7, 8) Nyfrieslandia sp. aff. N. complicata (Maletz and Bruton); PD13; (6) Nyfrieslandia ramosissima n. sp.; PD09; (9–11) Nyfrieslandia sp. A.; PD13; (12–15) Nyfrieslandia ramosissima n. sp. (12–14) PD13, (15) PD05; (16) Nyfrieslandia sp. aff. N. complicata (Maletz and Bruton); PD05.
Materials
Eighteen specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
This form is uncommon, but is easily recognized with its abundance of distinct, long outer spines that exhibit irregularly placed apophyses. Nyfrieslandia sp. aff. N. complicata appears to be highly variable in its development.
Material in this study has affinity with the Lower Ordovician Nyfrieslandia complicata Maletz and Bruton, Reference Maletz and Bruton2007 (Maletz and Bruton, Reference Maletz and Bruton2007, figs 26A–O; Won and Iams, Reference Won and Iams2013, fig. 6K) and N. crassispinosa Won and Iams, Reference Won and Iams2013 (Won and Iams, Reference Won and Iams2013, fig. 6I–M), based on comparable dimensions (outer and inner spheres Ø = 90–120 μm and 50 μm for Nyfrieslandia crassispinosa; ~190 μm and 120–140 μm for Nyfrieslandia sphaeroidea; 160–190 μm and 90–110 μm for Nyfrieslandia complicata; vs. 190–200 μm and 90–100 μm herein). Our material differs from Nyfrieslandia complicata in possessing longer outer spines (~30–40 μm, Maletz and Bruton, Reference Maletz and Bruton2007 vs. 56–99 μm herein); a thinner inner sphere and a slightly larger gap between the outer and inner sphere (35–40 μm, Maletz and Bruton, Reference Maletz and Bruton2007 vs. 42–52 μm herein).
Holotype
Nyfrieslandia ramosissima n. sp., UQSEES_M03S09, Figure 18, Supplemental Data file 9, sample PD05, Middle Ordovician, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.

Figure 18. Compilation of μ-CT models, SEM images and schematic drawings of Nyfrieslandia ramosissima n. sp. (holotype: UQSEES_M03S09 from PD05). (1–3) Spheres are digitally segmented for individual observation; each segment to the same scale; (1) complete specimen showing the growth of an additional outer sphere; (2) incomplete inner sphere preserving six radial bars (rb); (3) outer sphere; (3, 5) μ-CT models at different orientations; (4) SEM image of an outer sphere demonstrating the randomly distributed apophyses branching from the main radial bars; (6) diagram showing configuration of spheres, main spines (ms), long by-spine (bs); (7) close up of the relationship of the growth of an additional outer sphere; (8) close up of the randomly orientated apophyses arising from an outer spine that is connected to the outer sphere.
Diagnosis
Skeleton consisting of three spheres. The outer sphere is distinctive and consists of a loose 15–25 μm labyrinth. Six radial main spines (up to 250 μm long) are slightly curved convex towards a long by-spine.
Occurrence
Middle Ordovician, Table Cove Formation, Newfoundland, Canada.
Description
Skeleton consisting of three spheres: outer sphere Ø = 260–295 μm; well-defined median sphere 183–195 μm; inner sphere 100 μm. Median and inner spheres are typically thin and appear to be constructed of thin bandages producing irregular, subcircular pores (Ø = 10–25 μm).
The long by-spine originates from the pore-frame junctions of the median sphere. It is one third of the diameter of the main spine. Randomly distributed apophyses branch from the main radial bars between the median sphere and outer sphere to develop arches. These arches are commonly not in line with median and outer sphere growth. Numerous short by-spines originate from the pore-frame junctions of the median sphere to connect with the outer sphere. No pylome was observed.
Etymology
From the Latin, rāmus, meaning branched, for its common development of an additional sphere via randomly distributed apophyses branching from the main radial bars.
Materials
Sixteen specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Although it has an externally similar appearance, Nyfrieslandia ramosissima n. sp. differs from the inaniguttid Triplocuccus aksuranesis Pouille and Danelian in Pouille et al., Reference Pouille, Danelian, Pour and Popov2013 (see Kachovich and Aitchison, Reference Kachovich and Aitchison2020, fig. 8) in development of a basal layer and does not have a well-developed median sphere. Nyfrieslandia ramosissima n. sp. has randomly distributed apophyses branching from the main radial bars between the median and outer spheres. The development of the outer and the additional outer sphere is homologous to the Late Ordovician Kalimnasphaera maculosa Webby and Blom, Reference Webby and Blom1986 μ-CT scanned by Asatryan et al. (Reference Asatryan, Aitchison and Webby2017).
Nyfrieslandia sp. A
Figure 17.9–17.11
Description
Well-developed outer sphere (Ø = 210–231 μm) and an extremely thin and hollow inner sphere (Ø = 90–100 μm) formed from rounded bars. Numerous, irregularly shaped pores (outer sphere ~16 pores across the diameter; inner sphere ~11 pores across the diameter) are developed on the outer and inner sphere. Nine to eleven radiating spines originate on the inner sphere and extend as outer spines (up to 147 μm long). Numerous, short (~30 μm) by-spines develop on the outer sphere. Outer spines rarely produce apophyses.
Materials
Three specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Nyfrieslandia sp. A is distinguished from all other taxa in the genus Nyfrieslandia by its numerous pores on its outer and inner sphere (number of pores across the diameter of the inner sphere ~11 pores vs. 5 pores in Nyfrieslandia ramosissima n. sp., 9 pores in Nyfrieslandia sp. aff. N. complicata and Nyfrieslandia sp. B). Due to the development of numerous pores on each sphere, the polygonal shape characteristic of the aspiculids is rounded and commonly difficult to distinguish.
Nyfrieslandia sp. B
Figure 17.1–17.3
Description
Large well-developed outer sphere (Ø = 290–310 μm) an extremely thin and hollow inner sphere (Ø = 105–119 μm) formed from rounded bars. The outer sphere wall is thick with oval to angular shaped pores (Ø ~11 μm). Nine to ten main spines originate from the inner sphere and extend as short outer spines (up to 118 μm long).
Materials
Three specimens, Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada.
Remarks
Nyfrieslandia sp. B lacks the irregular meshwork created by apophyses connected to the main outer spines. The larger outer sphere also differs from other taxa in the genus Nyfrieslandia, but is united by the size of the inner sphere, which is constructed by bars.
Discussion
Faunal composition
The diversity of radiolarians freed by acetic acid from their host samples, PD01–PD13, is discussed by Kachovich and Aitchison (Reference Kachovich and Aitchison2020) and is updated to reflect taxonomic assignments presented herein (Table 1). Approximately half of all the radiolarians recovered were spumellarians (Kachovich and Aitchison, Reference Kachovich and Aitchison2020), with the balance being archaeospiculids, enactinarians, or incertae sedis. Among the remaining forms described herein, the majority are attributable to the Entactiniidae (43%) and Palaeospiculidae (41%), with ~9% belonging to the Aspiculidae.
Table 1. List of archaeospicularians, entactinarians, and other radiolarians of uncertain affinity from the Table Cove Formation, Piccadilly Quarry, Newfoundland, Canada, with numbers of specimens recovered indicating the relative abundance of different taxa and faunal diversity.

Spherical entactinarians
The spherical skeleton in entactinarians is produced through multiple branching of apophyses on the initial spicule. The fusing of these spinules, to create a defined spherical wall, is a heterogenetic feature that has developed many times in the evolution of entactinarians (e.g., Proventocitiidae, Protoentactiniidae, and Echidniniidae). As such, this feature should only be considered a genus-level feature in early Paleozoic radiolarian taxonomy.
The exact biostratigraphic distribution of groups (e.g., the palaeospiculids) is not yet well documented and their scarcity in some assemblages could simply be due to their rare occurrence. The complex spicular system of Neopalaeospiculum was analyzed with the μ-CT, which revealed the development of multiple generations of whorled rays on the elongated primary rays, indicating that it has affinities to the genus Svalbardospiculum. Previously, Won et al. (Reference Won, Iams and Reed2007) compared the spicular system of Neopalaeospiculum to that of Palaeospiculum. Externally however, Neopalaeospiculum, as reported herein, resembles the upper Darriwilian form Polyentactinia sp. cf. P. spinulenta Pouille and Danelian from the Shundy Formation, Kazakhstan (see Pouille et al., Reference Pouille, Danelian and Popov2014b, pl. 2, figs. 9, 10). In both groups, a similar dimension and orientation of the six spines that are projections of the initial spicule, as well as the size and construction of the outer shell by fused spinules, can be observed. However, the initial spicule of Neopalaeospiculum emanates from a point-centered spicule and not from a short bar-centered spicule (microbar ~8 μm long). More noticeable is the difference in the first-order of spinules (3 vs. 4–6 spinules on each whorl of Neopalaeospiculum). Even though the external appearances of the two groups are indistinguishable, they are likely derived from two different linages: Neopalaeospiculum from the genus Svalbardospiculum and forms such as those reported by Pouille et al. (Reference Pouille, Danelian and Popov2014b) may be from Ramuspiculum.
The majority of the entactinarians described herein are of a primitive type, having initial spicules that are not differentiated into an apical and a basal group and do not form a heteropolar “microsphere.” The six-rayed, ectopically positioned spicule is a stable feature and was already a common feature by the middle Darriwilian (Fig. 3). The arrangement of the six rays and the position and length of the median bar (MB) is rarely variable at the species level and occasionally at the genus level. Won (Reference Won1997) reported similar limits to variation in internal skeletal structure of Devonian entactiniids at both levels. All entactinarians within our material show a close association between main spines and the initial spicule. In radiolarians in which the inner structure is not known, the presence of eccentrically placed spines may indicate the presence of a primary spicule.
The origin of the bar-centered radiolarian spicule can be traced back to the upper Cambrian (Kozur et al., Reference Kozur, Mostler and Repetski1996). However, phylogenetic relationships are uncertain because no intermediate forms that show connections to the point-centered taxa exist. Comparing our material to Ramuspiculum from the Lower to Middle Ordovician (upper Floian to lower Dapingian) of Spitsbergen (Maletz and Bruton, Reference Maletz and Bruton2007), we observe the development of the point-centered spicule with six-rays into a six-rayed spicule with a short microbar. The microbar develops from an additional whorl of rays before starting to branch at the end points of the rays. The bar-centered spicule in the palaeospiculids appears to have developed separately from the median-bar (MB) that is already well developed in other families. However, the frequently bifurcated spine development of the skeleton closely resembles the construction of the skeleton in other entactinarian families.
The taxonomy of most groups discussed herein inevitably raises some questions that cannot be answered given the current state of knowledge. Some of these radiolarians contain spicular elements (e.g., tetrapoidal spicular system; Dumitrică, Reference Dumitrică2013) similar to early Mesozoic taxa, but are currently unknown in the late Paleozoic. The variability of the inner spicular system within the aspiculids is difficult to understand and to compare with other Paleozoic radiolarians.
Acknowledgments
We thank D. Wang and C. Evans at the University of Queensland Julius Kruttschnitt Mineral Research Center (Brisbane, Australia) for their technical assistance and advice. Financial support towards investigation of early Paleozoic radiolarian evolution was provided by the Australian Research Council (ARC DP1501013325 to Aitchison). We also thank reviewers T. Danelian and O. Obut, as well as Associate Editor P. Noble and J. Kastigar in the Journal of Paleontology Editorial Office for their helpful suggestions and edits that have improved earlier versions of this manuscript.
Data Availability Statement
Also available from UQ eSpace data repository. DOI: http://dx.doi.org/10.14264/uql.2019.296.




















 Open access
Open access