



INTRODUCTION
Until relatively recently, skeletons dating to the Paleoindian and Archaic periods were completely unknown in the Maya area. A handful of early skeletons found elsewhere in Mexico have generated great interest due to the fact that Mesoamerica forms a geographic bottleneck through which Paleoindian groups would have passed on their way south (González et al. Reference González, Jiménez, Hedges, Huddart, Ohman, Turner and Pompa y Padilla2003; Ríos-Muñoz Reference Ríos-Muñoz2013). However, in recent years, a trove of new data from the region's earliest inhabitants has emerged (Figure 1). Following a recent initiative by Mexico's Instituto Nacional de Antropología e Historia (INAH) to carry out archaeological surveys of caves and cenotes in the Yucatan (Atlas Arqueológico Subacuático para el Registro, Estudio y Protección de los Cenotes en la Península de Yucatán), numerous submerged sites containing evidence of human activity dating to the Late Pleistocene to Early Holocene have been documented (Figure 2). These contexts contain stone tools and various species of extinct pre-Holocene megafauna (some with evidence of human processing), and several submerged caves around Tulum, Quintana Roo have thus far produced 10 Paleoindian skeletons (Table 1). These skeletons vary in preservation, with the better-preserved among them being the focus of more extensive study.

Figure 1. Map of sites in the Maya region with early skeletons. Map created by Wrobel based on map by Sémhur, https://commons.wikimedia.org/wiki/File:Maya_civilization_location_map-blank.svg.
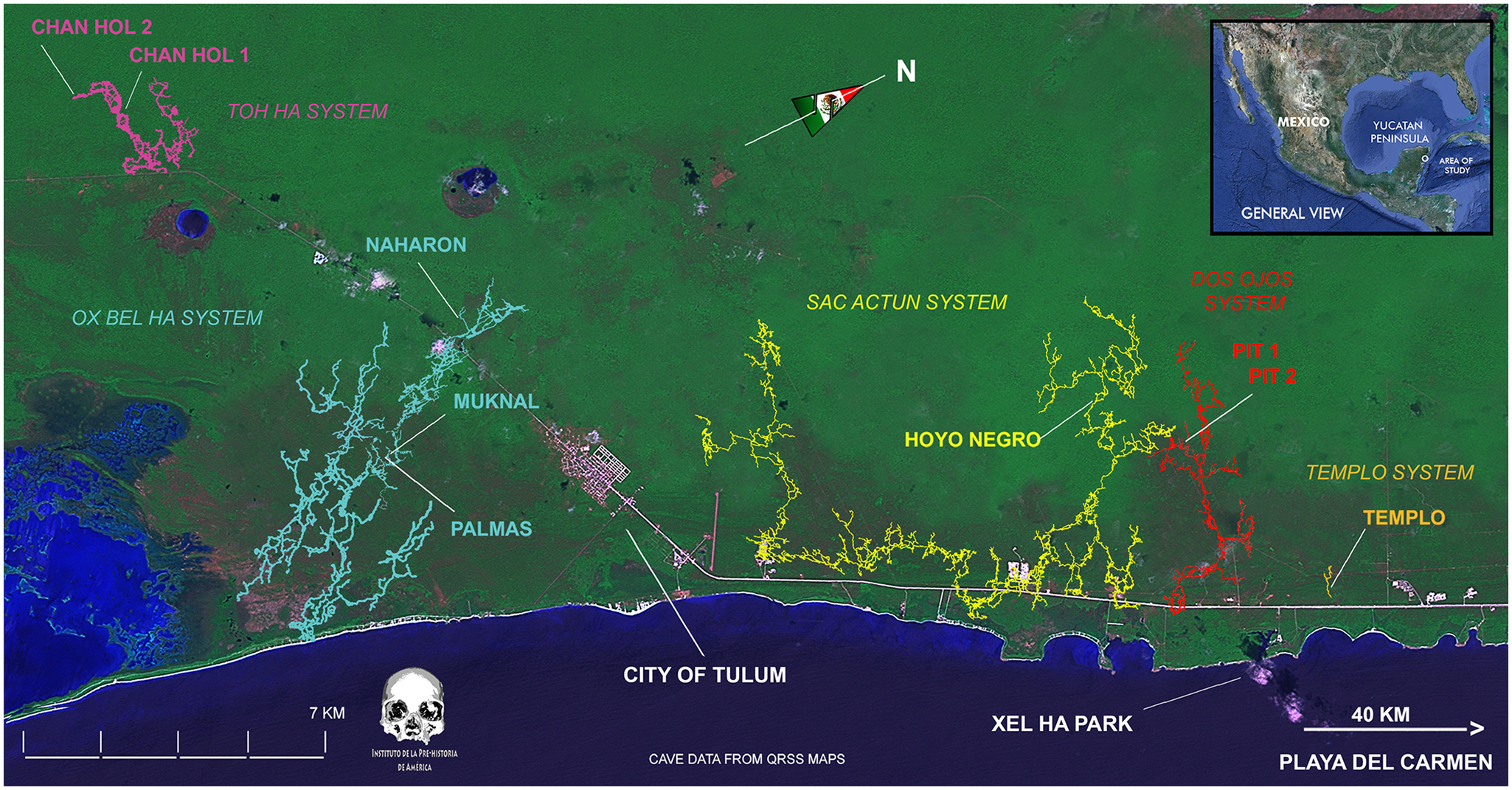
Figure 2. Coast of the Mexican state of Quintana Roo, with location of cenotes and caves containing sites with human skeletons and associated Pleistocene fauna. Map by Jerónimo Avilés Olguín; reproduced from Hubbe et al. (Reference Hubbe, Mata, Herrera, Benavente Sanvicente, González, Sandoval, Olguín, Núñez and Von Cramon-Taubadel2020:Figure 1) with permission.
Table 1. List of preceramic human skeletons from the Maya area.
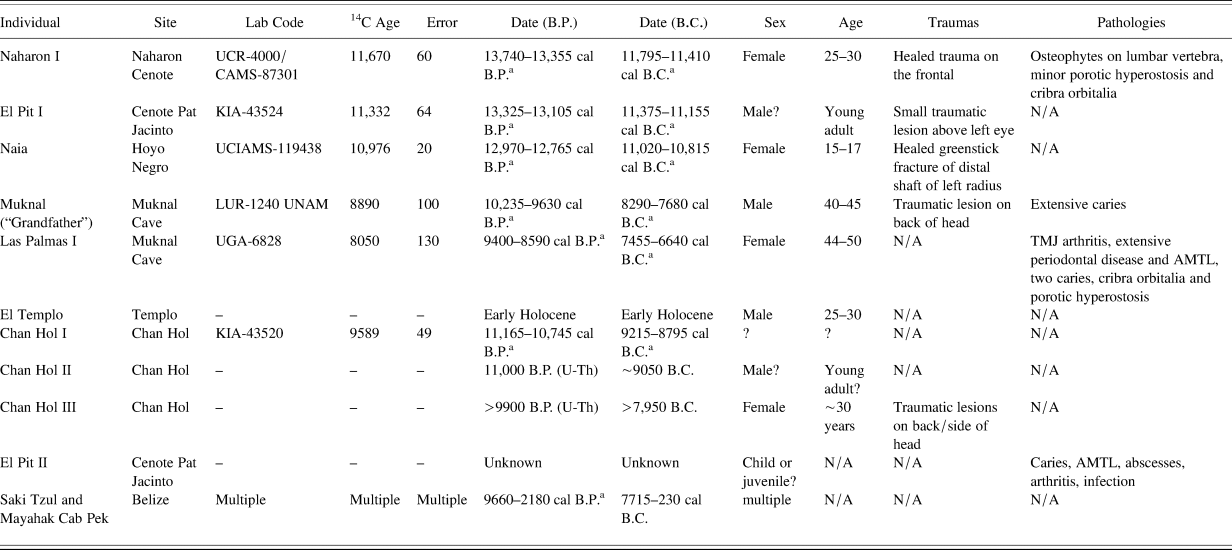
Note: ATML = antemortem tooth loss; TMJ = temporomandibular joint.
a Original 2σ dates were recalibrated with OxCal v. 4.4.2 (Bronk Ramsey Reference Bronk Ramsey2020) using the IntCal20 calibration curve (Reimer et al. Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey, Butzin, Cheng, Edwards, Friedrich, Grootes, Guilderson, Hajdas, Heaton, Hogg, Hughen, Kromer, Manning, Muscheler, Palmer, Pearson, van der Plicht, Reimer, Richards, Scott, Southon, Turney, Wacker, Adolphi, Büntgen, Capano, Fahrni, Fogtmann-Schulz, Friedrich, Köhler, Kudsk, Miyake, Olsen, Reinig, Sakamoto, Sookdeo and Talamo2020).
Another recent project in southern Belize has documented a large number of burials from rock shelters that span the Paleoindian, Archaic, and Maya Formative periods. Detailed data from these individuals are forthcoming, and because of the relatively large number of skeletons, their broad time range, and the excellent preservation, these have the potential to reveal unique and insightful glimpses of some of the region's early human inhabitants and to provide an understanding of the transition from hunting and gathering to agriculture.
AN EXPLANATION OF THE RADIOCARBON DATES PRESENTED IN THIS ARTICLE
Although some of the skeletons discussed in this review are among the best-preserved early skeletons in the New World, submerged contexts, contamination, and a lack of collagen has made direct dating of human skeletal remains complicated and controversial (Chatters et al. Reference Chatters, Allen and Jones2014; González González et al. Reference González González, Terrazas, Stinnesbeck, Benavente, Avilés, Rojas, Padilla, Velásquez, Acevez, Frey, Graf, Ketron and Waters2014; Stinnesbeck et al. Reference Stinnesbeck, Becker, Hering, Frey, González, Fohlmeister, Stinnesbeck, Frank, Mata, Benavente, Avilés, Aceves, Zell and Deininger2017; Stinnesbeck et al. Reference Stinnesbeck, Stinnesbeck, Mata, Olguín, Sanvicente, Zell, Frey, Lindauer, Sandoval, Morlet, Nuñez and González2018; Taylor Reference Taylor2009). Radiocarbon dates on bone apatite and residual proteins from several laboratories have established absolute ages between 8591 and 13,721 cal b.p. (Arroyo et al. Reference Arroyo, Luna, Chatters, Rissolo, Chávez, Nava and Barba2015, modified from González and Huddart Reference González and Huddart2008). However, dating human skeletal remains from submerged contexts remains problematic, since the submerged bone is susceptible to contamination from fossil carbon (Stinnesbeck et al. Reference Stinnesbeck, Becker, Hering, Frey, González, Fohlmeister, Stinnesbeck, Frank, Mata, Benavente, Avilés, Aceves, Zell and Deininger2017:2). Furthermore, as Devièse et al. (Reference Devièse, Stafford, Waters, Wathen, Comeskey, Becerra-Valdivia and Higham2018:171) have pointed out, more accurate radiocarbon dating using pretreatment methods, such as XAD chromatography to deal with contamination, is needed to build robust chronologies for the peopling of the Americas.
Given that this overview presents the published 14C dates in the literature of the region, we consider all published early dates from Yucatan in relation to a range of data derived from geological context and associated materials. For instance, in some cases, it is clear from the presence of articulations and drip formations on the bones that bodies had decayed prior to inundation of the caves caused by rising sea levels during the last deglaciation, between 13,000 and 7,600 years ago (Collins et al. Reference Collins, Reinhardt, Rissolo, Chatters, Nava Blank and Luna Erreguerena2015). Uranium-thorium dating of calcite formations on bones also provides a minimum age. However, solely using these methods to establish the age of human occupation is problematic, since they provide only a terminus post quem for when the cave formation was initiated, which could have occurred much later than the human occupation being assessed. Finally, radiocarbon dating is often possible on associated materials, such as charcoal and animal bones (also subject to the same contamination as human remains), as well as carbon residue left on the ceilings from torches. Regardless, we discuss the dates presented in Table 1 and whether they show temporal correspondence with the archaeological contexts (or other published radiocarbon data) in which they were recovered.
Radiocarbon dating has been central to archaeology since its development by Willard Libby (Arnold and Libby Reference Arnold and Libby1949; Libby et al. Reference Libby, Anderson and Arnold1949). There have been four “radiocarbon revolutions” since that time (Bayliss Reference Bayliss2009): first, Libby's development of the method; second, the development of a calibration curve using tree-ring and coral records (Pearson Reference Pearson1987; Reimer et al. Reference Reimer, Baillie, Bard, Bayliss, Beck, Bertrand, Blackwell, Buck, Burr, Cutler, Damon, Edwards, Fairbanks, Friedrich, Guilderson, Hogg, Hughen, Kromer, McCormac, Manning, Bronk Ramsey, Reimer, Remmele, Southon, Stuiver, Talamo, Taylor, van der Plicht and Weyhenmeyer2004; Suess Reference Suess1965; Stuiver et al. Reference Stuiver, Reimer, Bard, Beck, Burr, Hughen, Kromer, McCormac, van der Plicht and Spurk1998); third, the development of AMS 14C dating (Taylor Reference Taylor1995); and fourth, the development of Bayesian chronological modelling (Bronk Ramsey Reference Bronk Ramsey1994, Reference Bronk Ramsey2020; Stuiver et al. Reference Stuiver, Reimer and Reimer2021). Radiocarbon dating is based on the half-life of 14C of 5,730 years, meaning that in that time half of the 14C present at the time of an organism's death would be lost through radioactive decay (Bronk Ramsey Reference Bronk Ramsey2008:250). Samples selected for dating are pre-treated based on material type (e.g., Acid-Base-Acid for charcoal samples), after which 14C/12C and 13C/12C ratios are measured using conventional decay counting methods or accelerator mass spectrometry (AMS). Given that the amount of 14C in the atmosphere has not been constant over time, experts have utilized tree-rings and coral records to create calibration curves going back 55,000 years for terrestrial records in the northern (Reimer et al. Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey, Butzin, Cheng, Edwards, Friedrich, Grootes, Guilderson, Hajdas, Heaton, Hogg, Hughen, Kromer, Manning, Muscheler, Palmer, Pearson, van der Plicht, Reimer, Richards, Scott, Southon, Turney, Wacker, Adolphi, Büntgen, Capano, Fahrni, Fogtmann-Schulz, Friedrich, Köhler, Kudsk, Miyake, Olsen, Reinig, Sakamoto, Sookdeo and Talamo2020) and southern (Hogg et al. Reference Hogg, Heaton, Hua, Palmer, Turney, Southon, Bayliss, Blackwell, Boswijk, Ramsey, Pearson, Petchey, Reimer, Reimer and Wacker2020) hemispheres. This calibration stage often employs calibration software, such as Calib (Stuiver et al. Reference Stuiver, Reimer and Reimer2021) or OxCal (Bronk Ramsey Reference Bronk Ramsey1994, Reference Bronk Ramsey2020), with simple user interfaces to allow ease of use for even the most inexperienced user. Finally, the calibrated date is compared to the archaeological context to ensure that the sample accurately dates the event of interest.
The 14C dates presented in Table 1 and throughout this review have been recalibrated from previously published dates following the recent Intcal20 Northern Hemisphere calibration curve (Reimer et al. Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey, Butzin, Cheng, Edwards, Friedrich, Grootes, Guilderson, Hajdas, Heaton, Hogg, Hughen, Kromer, Manning, Muscheler, Palmer, Pearson, van der Plicht, Reimer, Richards, Scott, Southon, Turney, Wacker, Adolphi, Büntgen, Capano, Fahrni, Fogtmann-Schulz, Friedrich, Köhler, Kudsk, Miyake, Olsen, Reinig, Sakamoto, Sookdeo and Talamo2020), which features improvements based on statistically integrated evidence from a variety of independent sources (e.g., tree-rings, lacustrine and marine sediments, speleothems, and coral), as well as extension of the atmospheric record back to 13,900 cal b.p. based on tree-ring records (Reimer et al. Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey, Butzin, Cheng, Edwards, Friedrich, Grootes, Guilderson, Hajdas, Heaton, Hogg, Hughen, Kromer, Manning, Muscheler, Palmer, Pearson, van der Plicht, Reimer, Richards, Scott, Southon, Turney, Wacker, Adolphi, Büntgen, Capano, Fahrni, Fogtmann-Schulz, Friedrich, Köhler, Kudsk, Miyake, Olsen, Reinig, Sakamoto, Sookdeo and Talamo2020:726). Significant improvements were made in several areas of the calibration curve, including for the Late Glacial period around 12,500 cal b.p. and the Hallstatt Plateau around 2600 cal b.p. (Reimer et al. Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey, Butzin, Cheng, Edwards, Friedrich, Grootes, Guilderson, Hajdas, Heaton, Hogg, Hughen, Kromer, Manning, Muscheler, Palmer, Pearson, van der Plicht, Reimer, Richards, Scott, Southon, Turney, Wacker, Adolphi, Büntgen, Capano, Fahrni, Fogtmann-Schulz, Friedrich, Köhler, Kudsk, Miyake, Olsen, Reinig, Sakamoto, Sookdeo and Talamo2020:744–745).
THE EARLIEST SKELETONS
Naia
Perhaps the most famous of the early skeletons found in the Maya area is “Naia” (HN5/48), a female adolescent (15–17 years old) found in the submerged pit of Hoyo Negro. Dating based on multiple AMS 14C measurements of bone apatite and enamel bioapatite from independent labs place her between 12,970 and 12,765 cal b.p. (UCIAMS-119438:10,976 ± 20, from Chatters et al. Reference Chatters, Allen and Jones2014, recalibrated using OxCal v. 4.4.2 [Bronk Ramsey et al. Reference Bronk Ramsey2020] with the IntCal20 calibration curve [Reimer et al. Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey, Butzin, Cheng, Edwards, Friedrich, Grootes, Guilderson, Hajdas, Heaton, Hogg, Hughen, Kromer, Manning, Muscheler, Palmer, Pearson, van der Plicht, Reimer, Richards, Scott, Southon, Turney, Wacker, Adolphi, Büntgen, Capano, Fahrni, Fogtmann-Schulz, Friedrich, Köhler, Kudsk, Miyake, Olsen, Reinig, Sakamoto, Sookdeo and Talamo2020]), making this exceptionally well-preserved skeleton one of the oldest in the Americas (Stinnesbeck et al. Reference Stinnesbeck, Becker, Hering, Frey, González, Fohlmeister, Stinnesbeck, Frank, Mata, Benavente, Avilés, Aceves, Zell and Deininger2017). XAD purification of the dentine protein of human and gomphothere molars was attempted at two labs (Stafford Research Lab and The Pennsylvania State University), although both failed to produce a pseudomorph (Chatters et al. Reference Chatters, Allen and Jones2014:Supplemental Materials). Although bioapatite dates are less than ideal due to contamination from dissolved organic carbon, they were used as a maximum age for the skeleton and were consistent with U-Th ages of florets on animal remains (ranging between 18.8 ± 0.3 ka and 11.9 ± 0.3 ka; Chatters et al. Reference Chatters, Allen and Jones2014:752), producing an initial 230Th/232Th atomic ratio of 32 ± 16 ppm (Chatters et al. Reference Chatters, Allen and Jones2014:Supplemental Materials:14). Extensive in situ documentation of the site and human remains, including photogrammetric modeling (Rissolo et al. Reference Rissolo, Blank, Petrovic, Chávez, Jaskolski, Luna and Chatters2015), has facilitated detailed analyses and interpretation of the context. Chatters and colleagues (Reference Chatters, Allen and Jones2014:S15) argue that her placement in the cave was likely accidental and followed a fall into the deep chamber from an upper passage. Noting that the bones, scattered over an area measuring 5 m × 1.5 m, formed clusters of partially articulated elements, they argue that her body began decomposing while floating in shallow floodwaters, which later retreated, depositing disconnected body segments on the cave floor.
Naia's skeleton presents several clues about her life and death. Even considering her young age, dental wear is quite light when compared to the dentitions of other early skeletons in the Americas (Cucina et al. Reference Cucina, Herrera and Chatters2019). Naia presents periodontal disease, which is not particularly uncommon in hunter-gatherers (Littleton Reference Littleton2018), but also extensive dental caries, which is more typical of later populations in the region that had high-carbohydrate maize-dominated diets (Cucina and Tiesler Reference Cucina and Tiesler2003). Instead, it seems likely she must have had extensive access to fruit and/or honey (Chatters et al. Reference Chatters, Allen and Jones2014). A healed greenstick fracture of the distal shaft of her left radius probably resulted from a fall during childhood, while perimortem breaks evident on her pelvis were likely sustained as a result of her fall into the chamber.
Naranjal Subsystem
Naharon I (or “Naharon Eve”), a 25- to 30-year-old female, was represented by approximately 80 percent of a mostly disarticulated skeleton (González González et al. Reference González González, Rojas, Acevez, Avilés, Analco, del Río, Luna, Velázquez, Stinnesbeck, Terrazas, Benavente, Leshikar-Denton and Erreguerena2008a, Reference González González, Rojas, Terrazas, Benavente, Stinnesbeck, Aviles, de los Ríos and Aceves2008b). However, partial articulations of the vertebrae (González González et al. Reference González González, Rojas, Acevez, Avilés, Analco, del Río, Luna, Velázquez, Stinnesbeck, Terrazas, Benavente, Leshikar-Denton and Erreguerena2008a:132) suggest that decay occurred in situ (González González et al. Reference González González, Terrazas, Stinnesbeck, Benavente, Avilés, Rojas, Padilla, Velásquez, Acevez, Frey, Graf, Ketron and Waters2014:329). She shows signs of pathologies in the form of osteophytic changes to a lumbar vertebra, indicating the onset of arthritis, minor porotic hyperostosis, and cribra orbitalia, signifying metabolic disturbances, and healed trauma on the frontal (González González et al. Reference González González, Rojas, Acevez, Avilés, Analco, del Río, Luna, Velázquez, Stinnesbeck, Terrazas, Benavente, Leshikar-Denton and Erreguerena2008a:132). Additionally, a congenital defect is present in the form of completely fused second and third vertebrae, an indicator of Klippel-Feil syndrome. There is no indication of whether the individual suffered from disfigurement or reduced mobility, which affect some individuals with Klippel-Feil. A femur sample provided a radiocarbon date of 13,740–13,355 cal b.p. (UCR-4000/CAMS-87301:11,670 ± 60 [González González et al. Reference González González, Rojas, Terrazas, Benavente, Stinnesbeck, Aviles, de los Ríos and Aceves2008b:13] recalibrated using OxCal v. 4.4.2 [Bronk Ramsey et al. Reference Bronk Ramsey2020] using the IntCal20 calibration curve [Reimer et al. Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey, Butzin, Cheng, Edwards, Friedrich, Grootes, Guilderson, Hajdas, Heaton, Hogg, Hughen, Kromer, Manning, Muscheler, Palmer, Pearson, van der Plicht, Reimer, Richards, Scott, Southon, Turney, Wacker, Adolphi, Büntgen, Capano, Fahrni, Fogtmann-Schulz, Friedrich, Köhler, Kudsk, Miyake, Olsen, Reinig, Sakamoto, Sookdeo and Talamo2020]) derived from its amino acid fraction. However, the authors did not report the purification methods used in radiocarbon dating the femur sample; therefore, we are unable to assess the reliability of the results. Furthermore, attempts to replicate the result in other labs were unsuccessful (González González et al. Reference González González, Rojas, Terrazas, Benavente, Stinnesbeck, Aviles, de los Ríos and Aceves2008b:13).
Las Palmas I (or “Señora de Las Palmas”) is a mostly articulated skeleton placed flexed on the left side, and thought to have been originally wrapped and interred in an upright, seated position (González González et al. Reference González González, Rojas, Acevez, Avilés, Analco, del Río, Luna, Velázquez, Stinnesbeck, Terrazas, Benavente, Leshikar-Denton and Erreguerena2008a:133). The individual, whose well-preserved skeleton is 90 percent complete, is estimated to be a woman aged between 44 and 50 years. She was the subject of a forensic reconstruction by Mexico's INAH, which briefly generated some misguided narratives in the popular press, based on noted facial similarities to modern peoples from southeastern Asia and Indonesia (rather than northeast Asia, where most evidence for the origin of New World populations points). AMS analysis of nearby charcoal deposits from a hearth produced calibrated ages (2σ) of 10,210–9905 cal b.p. (INAH-2123, 8941 ± 39 [González González et al. Reference González González, Rojas, Terrazas, Benavente, Stinnesbeck, Aviles, de los Ríos and Aceves2008b:Table 1]) and 8595–8425 cal b.p. (INAH-2118, 7740 ± 39 [González González et al. Reference González González, Rojas, Terrazas, Benavente, Stinnesbeck, Aviles, de los Ríos and Aceves2008b:Table 1]), and a direct date on the bone produced a calibrated age (2σ) of 9400–8590 cal b.p. (UGA-6828: 8050 ± 130 [González González et al. Reference González González, Rojas, Terrazas, Benavente, Stinnesbeck, Aviles, de los Ríos and Aceves2008b:Table 1] recalibrated using OxCal v. 4.4.2 [Bronk Ramsey et al. Reference Bronk Ramsey2020] using the IntCal20 calibration curve [Reimer et al. Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey, Butzin, Cheng, Edwards, Friedrich, Grootes, Guilderson, Hajdas, Heaton, Hogg, Hughen, Kromer, Manning, Muscheler, Palmer, Pearson, van der Plicht, Reimer, Richards, Scott, Southon, Turney, Wacker, Adolphi, Büntgen, Capano, Fahrni, Fogtmann-Schulz, Friedrich, Köhler, Kudsk, Miyake, Olsen, Reinig, Sakamoto, Sookdeo and Talamo2020]). Because the authors did not report the purification methods used in radiocarbon-dating the bone sample, we are unable to assess the reliability of the results. However, given that the results are relatively comparable with the charcoal dates, and U-Th dating yielded an age between 10,000 and 12,000 cal b.p. (González González et al. Reference González González, Rojas, Terrazas, Benavente, Stinnesbeck, Aviles, de los Ríos and Aceves2008b:13) that provides a terminus post quem for the bone ages, it is likely the dates are relatively accurate. No initial Th amounts are reported.
Approximately 60 meters from the Las Palmas site, Muknal is a partial skeleton of a male aged 40–45 years old who was placed in a small chamber at the end of a narrow, 35-m long tunnel (González González et al. Reference González González, Terrazas, Stinnesbeck, Benavente, Avilés, Rojas, Padilla, Velásquez, Acevez, Frey, Graf, Ketron and Waters2014; Stinnesbeck et al. Reference Stinnesbeck, Stinnesbeck, Mata, Olguín, Sanvicente, Zell, Frey, Lindauer, Sandoval, Morlet, Nuñez and González2018). Pathologies include arthritis of the temporomandibular joints, extensive evidence of periodontal disease and antemortem tooth loss, an injury to the back of the head, and dental attrition and two caries in the remaining teeth (González González et al. Reference González González, Terrazas, Stinnesbeck, Benavente, Avilés, Rojas, Padilla, Velásquez, Acevez, Frey, Graf, Ketron and Waters2014:331–332). Additionally, metabolic disease is indicated by the presence of cribra orbitalia and porotic hyperostosis. Nearby was found the mandible of an extinct genus of peccary, with ancient cuts and scratches made with stone tools (Stinnesbeck et al. Reference Stinnesbeck, Stinnesbeck, Mata, Olguín, Sanvicente, Zell, Frey, Lindauer, Sandoval, Morlet, Nuñez and González2018). The Muknal skeleton, approximately 50 percent complete, consists of the skull, mandible, a clavicle, and a few long bones. Stinnesbeck and colleagues (Reference Stinnesbeck, Stinnesbeck, Mata, Olguín, Sanvicente, Zell, Frey, Lindauer, Sandoval, Morlet, Nuñez and González2018:9–10) present a thorough and compelling discussion, arguing that the partial remains were intentionally deposited there secondarily, as part of an extended mortuary ritual. The secondary burial deposit is dated to 10,235 to 9639 cal b.p. (LUR-1240: 8890 ± 100, from Stinnesbeck et al. Reference Stinnesbeck, Stinnesbeck, Mata, Olguín, Sanvicente, Zell, Frey, Lindauer, Sandoval, Morlet, Nuñez and González2018:Table 1), based on dates on charcoal recovered directly below the skeletal remains (recalibrated from Stinnesbeck et al. Reference Stinnesbeck, Stinnesbeck, Mata, Olguín, Sanvicente, Zell, Frey, Lindauer, Sandoval, Morlet, Nuñez and González2018 with OxCal v. 4.4.2 [Bronk Ramsey et al. Reference Bronk Ramsey2020] using the IntCal20 calibration curve [Reimer et al. Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey, Butzin, Cheng, Edwards, Friedrich, Grootes, Guilderson, Hajdas, Heaton, Hogg, Hughen, Kromer, Manning, Muscheler, Palmer, Pearson, van der Plicht, Reimer, Richards, Scott, Southon, Turney, Wacker, Adolphi, Büntgen, Capano, Fahrni, Fogtmann-Schulz, Friedrich, Köhler, Kudsk, Miyake, Olsen, Reinig, Sakamoto, Sookdeo and Talamo2020]).
El Templo was a poorly preserved, but mostly articulated male, approximately 25–30 years old at time of death (González González et al. Reference González González, Rojas, Acevez, Avilés, Analco, del Río, Luna, Velázquez, Stinnesbeck, Terrazas, Benavente, Leshikar-Denton and Erreguerena2008a:135). Despite the poor preservation, approximately 70 percent of the skeleton was recovered. Based on the position of the body, prone and limbs akimbo, González González and colleagues (Reference González González, Terrazas, Stinnesbeck, Benavente, Avilés, Rojas, Padilla, Velásquez, Acevez, Frey, Graf, Ketron and Waters2014) posit an accidental death. Though no dating analysis is reported for this context, the proposed early date appears to be based on the location of the articulated individual, whose bones were cemented to the floor while the cave was dry, in a submerged chamber that would have been flooded during the Early Holocene (González González et al. Reference González González, Rojas, Acevez, Avilés, Analco, del Río, Luna, Velázquez, Stinnesbeck, Terrazas, Benavente, Leshikar-Denton and Erreguerena2008a:138, Reference González González, Rojas, Terrazas, Benavente, Stinnesbeck, Aviles, de los Ríos and Aceves2008b:1).
Three individuals were found in the Chan Hol cave, which forms part of the Toh Ha system and is accessed through the Chan Hol cenote, approximately 15 kilometers south of Tulum. Located 530 m northeast of the cenote entrance, Chan Hol I (or “El Joven”) was partially articulated, with the body placed on the left side and legs flexed, while the right arm was extended to the right. Though 90 percent complete, poor preservation limits the analytical potential of these remains, so much so that age and sex could not be assessed (González González et al. Reference González González, Terrazas, Stinnesbeck, Benavente, Avilés, Rojas, Padilla, Velásquez, Acevez, Frey, Graf, Ketron and Waters2014:Table 19.1). A 14C analysis on bone apatite produced a date of 11,165–10,745 cal b.p. (KIA-43520: 9589 ± 49 [González González et al. Reference González González, Terrazas, Stinnesbeck, Benavente, Avilés, Rojas, Padilla, Velásquez, Acevez, Frey, Graf, Ketron and Waters2014:Table 19.2] recalibrated with OxCal v. 4.4.2 [Bronk Ramsey et al. Reference Bronk Ramsey2020] using the IntCal20 calibration curve [Reimer et al. Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey, Butzin, Cheng, Edwards, Friedrich, Grootes, Guilderson, Hajdas, Heaton, Hogg, Hughen, Kromer, Manning, Muscheler, Palmer, Pearson, van der Plicht, Reimer, Richards, Scott, Southon, Turney, Wacker, Adolphi, Büntgen, Capano, Fahrni, Fogtmann-Schulz, Friedrich, Köhler, Kudsk, Miyake, Olsen, Reinig, Sakamoto, Sookdeo and Talamo2020]). No description of the purification methods used in radiocarbon-dating the bone sample are given for either publication (González González et al. Reference González González, Rojas, Acevez, Avilés, Analco, del Río, Luna, Velázquez, Stinnesbeck, Terrazas, Benavente, Leshikar-Denton and Erreguerena2008a, Reference González González, Terrazas, Stinnesbeck, Benavente, Avilés, Rojas, Padilla, Velásquez, Acevez, Frey, Graf, Ketron and Waters2014), so we are unable to assess the reliability of the results.
Chan Hol II was located approximately 1,200 meters west of the cenote entrance in a small cave tunnel. After its discovery, but before it could be studied, the site was looted and most of the bones from the approximately 80 percent complete skeleton were taken, including the nearly complete cranium and many intact long bones (González González et al. Reference González González, Terrazas, Stinnesbeck, Benavente, Avilés, Rojas, Padilla, Velásquez, Acevez, Frey, Graf, Ketron and Waters2014; Stinnesbeck et al. Reference Stinnesbeck, Becker, Hering, Frey, González, Fohlmeister, Stinnesbeck, Frank, Mata, Benavente, Avilés, Aceves, Zell and Deininger2017). Much of the analysis is based on the few pictures taken by divers at the time of the individual's initial discovery. Visible skeletal features suggest the individual is possibly a young adult male, and though the body was clearly articulated, the informal position likely indicates that the placement was not intentional. A recent study by Stinnesbeck and colleagues (Reference Stinnesbeck, Becker, Hering, Frey, González, Fohlmeister, Stinnesbeck, Frank, Mata, Benavente, Avilés, Aceves, Zell and Deininger2017) places the date at around 11,000 years b.p. (9050 b.c.), using U-series techniques (with U concentration values ranging between 0.2605 and 12.78 ppm and 232Th concentrations of <10 ng g-1) and stable isotope analyses on a stalagmite overgrowing the pelvis. However, without a direct date on cultural remains, this U-Th date offers a date only sometime after the original human activity, rather than a direct indication of the timing of the human activity itself.
Chan Hol III is the most recently discovered early skeleton in this region (Stinnesbeck et al. Reference Stinnesbeck, Rennie, Olguín, Stinnesbeck, González, Frank, Warken, Schorndorf, Krengel, Morlet and González2020). Approximately 30 percent complete and not in its original position, the skeleton belongs to an adult female, likely in the age range of 20–40 years. Many of the teeth were lost antemortem, some in association with dental abscesses, and caries were present on two of the remaining molars. Other pathologies include arthritis on the humeral head and on several vertebrae, as well as signs of infection on the cranium that perhaps are related to trauma or may represent treponemal disease. The cranium shows evidence of three traumas to the posterior and lateral portion of the skull, resulting in bone loss and remodeling. Finally, strontium isotope analysis indicates the individual was of local (upper Yucatan Peninsula) origin. Due to a lack of preserved collagen, the individual was assigned a minimum age of 9900 yr b.p. (7950 +/−100 b.c.), using 230Th/U-analysis of flowstone directly covering the phalanges (Stinnesbeck et al. Reference Stinnesbeck, Rennie, Olguín, Stinnesbeck, González, Frank, Warken, Schorndorf, Krengel, Morlet and González2020:14). Initial U concentrations of 140–157 ng/g were reported, alongside 232Th concentrations of <10 ng g-1 (Stinnesbeck et al. Reference Stinnesbeck, Rennie, Olguín, Stinnesbeck, González, Frank, Warken, Schorndorf, Krengel, Morlet and González2020:5).
Sac Actun System
Fragments of at least two ancient skeletons were found dispersed in the debris mound of Cenote Pat Jacinto (also known as “El Pit”), which is connected to the Sac Actun System, the deepest system in the survey area and one of the deepest known in the entire Yucatan Peninsula (González González et al. Reference González González, Terrazas, Stinnesbeck, Benavente, Avilés, Rojas, Padilla, Velásquez, Acevez, Frey, Graf, Ketron and Waters2014:332–334). El Pit I consists of a calotte and a collection of postcranial material assumed to belong to the same individual. A pronounced nuchal crest suggests the individual is male, and the dentition is consistent with a young adult. The cranium shows evidence of trauma in life in the form of a small lesion above the left eye. The second individual, El Pit II, consists of a partial cranium and fragmentary postcranial elements of a likely child or juvenile.
González González and colleagues (Reference González González, Terrazas, Stinnesbeck, Benavente, Avilés, Rojas, Padilla, Velásquez, Acevez, Frey, Graf, Ketron and Waters2014:333) argue for the antiquity of both individuals from Cenote Pat Jacinto, though they were found in the debris mound of the cenote near the cave entrance. In the case of El Pit I, the partial skull has a mesocranial shape and lacks intentional modification, both somewhat unusual in Pre-Contact Maya from the area, and a radiometric 14C dating of bone apatite provided a preliminary calibrated age of approximately 13,325–13,105 cal b.p. (KIA-43524: 11,332 ± 64 [González González et al. Reference González González, Terrazas, Stinnesbeck, Benavente, Avilés, Rojas, Padilla, Velásquez, Acevez, Frey, Graf, Ketron and Waters2014:Table 19.2] recalibrated with OxCal v. 4.4.2 [Bronk Ramsey et al. Reference Bronk Ramsey2020] using the IntCal20 calibration curve [Reimer et al. Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey, Butzin, Cheng, Edwards, Friedrich, Grootes, Guilderson, Hajdas, Heaton, Hogg, Hughen, Kromer, Manning, Muscheler, Palmer, Pearson, van der Plicht, Reimer, Richards, Scott, Southon, Turney, Wacker, Adolphi, Büntgen, Capano, Fahrni, Fogtmann-Schulz, Friedrich, Köhler, Kudsk, Miyake, Olsen, Reinig, Sakamoto, Sookdeo and Talamo2020]). However, given the issues inherent in dating bone apatite, with the carbonate in apatite being subject to isotopic exchange with dissolved environmental carbon, this method is considered less reliable than collagen dates (Zazzo and Saliège Reference Zazzo and Saliège2011:52). Therefore, without a corresponding date we cannot assess the reliability of this date without a comparison with other 14C dates or cultural materials. No particular reason is given for assigning El Pit II an ancient date, though presumably this relates to its proximity to the El Pit I individual. In addition, no descriptions of the purification methods were given for the radiocarbon dating of the bone apatite for this sample; therefore, the reliability of this date cannot be assessed.
Belize
Recent work at two rock shelters in the Maya Mountains of southern Belize has uncovered a large number of very early burials (Figure 1; Hill et al. Reference Hill, Moes, Trask, Kate, McKenzie, Edgar, Kennett and Prufer2018; Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020; Prufer et al. Reference Prufer, Alsgaard, Robinson, Meredith, Culleton, Dennehy, Magee, Huckell, Stemp, Awe, Capriles and Kennett2019). Basic information has been published for all 52 individuals, providing estimates of sex and age, as well as brief descriptions of the burial contexts, including skeletal preservation, body position, and any indications of disturbance or secondary placement (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020: Supplement 1). Because the rock shelters were used for burial over a period spanning thousands of years, most of the individuals are partially disturbed or are represented by commingled or isolated bones. A plot of radiocarbon dates identifies three Late Paleoindian individuals (PSUAMS-4290: 8565 ± 45; UCIAMS-151854: 8290 ± 35; PSUAMS-4800: 7900 ± 35) dating to 9660–8590 cal b.p., two Early Archaic individuals (PSUAMS-3206: 6485 ± 30; PSUAMS-3205: 6455 ± 30) dating to 7465–7310 cal b.p., and at least 25 Late Archaic individuals dating from 5710 cal b.p. (UCIAMS-151853: 4905 ±25) to 3985 cal b.p. (PSUAMS-2681:3735 ± 20; Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020:Figure 2). Collagen samples were purified using either ultrafiltration or XAD chromatography based on sample preservation (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020:9). This collection is exciting not only because of its five earliest individuals, which are contemporaneous with the northern Yucatan samples, but for the large number of Late Archaic individuals, which are the only ones known from this time period in the Maya region. Initial dietary isotope work has traced changes in patterns of maize consumption over this span, identifying its introduction to the local population by 4000 b.p. (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020).
The three Late Paleoindian individuals, all of whom were recovered from excavations in the Mayahak Cab Pek rock shelter, include a partially disturbed but well-preserved 30- to 50-year-old adult male buried in a semi-flexed position, with his head to the east and facing south; a mostly disarticulated cluster of bones in fair condition representing a middle to old adult female; and an adult of unknown sex found in a commingled context with at least seven other individuals (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020:Supplement 1). Two Early Archaic individuals recovered from the Saki Tzul rock shelter were in the same burial feature. The two males, one of whom was 35–55 years old and the other 35–45 years old, were both arranged in flexed positions with heads to the north/northeast. Based on the burial descriptions by Kennett and colleagues (Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020:Supplement 1), they do not appear to have been buried at the same time, though it is certainly possible that their close proximity was intentional.
There are 25 Late Archaic individuals found at both the Mayahak Cab Pek rock shelter and the Saki Tzul rock shelter, with various degrees of preservation. These individuals’ ages range from infant (<6 months) to old adult (>70 years), with males and females present in equal numbers. Though the burial population is relatively small, the age distribution is consistent with an expected mortality curve, with adults and subadults in relatively equal proportions (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020:Supplement 1). While this small dataset is not appropriate for paleodemographic analysis, it at least suggests that the use of these burial sites was not restricted by aspects of social identity related to age or sex. Further aDNA analyses could identify patterns of genetic relatedness among the burials to determine whether mortuary use of the sites was limited to particular lineages. Finally, the consistent mortuary use of these rock shelters for simple, mostly unadorned burials over a period spanning thousands of years provides an interesting record of the antiquity of similar cave-related mortuary practices that have been documented during the Preclassic and Classic periods at rock shelter sites in the Caves Branch and Roaring Creek river valleys of central Belize (Figure 3; Wrobel et al. Reference Wrobel, Morton, Michael, Biggs and Andres2017).

Figure 3. Members of the Bladen Paleoindian and Archaic Archaeological Project (BPAAP) documenting the skeleton of Burial MHCP.17.1.8. The 9,600 year old male is the oldest skeleton in the region excavated from stratified contexts and directly dated. Image courtesy of Keith Prufer.
Two individuals, one from Saki Tzul (7465–7320 cal b.p.) and one from Mayahak Cab Pek (9660–9135 cal b.p.), were included in a large-scale analysis of ancient genomes from Central and South America (Posth et al. Reference Posth, Nakatsuka, Lazaridis, Skoglund, Mallick, Lamnidis, Rohland, Nägele, Adamski, Bertolini, Broomandkhoshbacht, Cooper, Culleton, Ferraz, Ferry, Furtwängler, Haak, Harkins, Harper, Hünemeier, Lawson, Llamas, Michel, Nelson, Oppenheimer, Patterson, Schiffels, Sedig, Stewardson, Talamo, Wang, Hublin, Hubbe, Harvati, Delaunay, Beier, Francken, Kaulicke, Reyes-Centeno, Rademaker, Trask, Robinson, Gutierrez, Prufer, Salazar-García, Chim, Gomes, Alves, Liryo, Inglez, Oliveira, Bernardo, Barioni, Wesolowski, Scheifler, Rivera, Plens, Messineo, Figuti, Corach, Scabuzzo, Eggers, DeBlasis, Reindel, Méndez, Politis, Tomasto-Cagigao, Kennett, Strauss, Fehren-Schmitz, Krause and Reich2018). The latter burial was disarticulated (Prufer et al. Reference Prufer, Alsgaard, Robinson, Meredith, Culleton, Dennehy, Magee, Huckell, Stemp, Awe, Capriles and Kennett2019), and thus, presumably, was a secondary interment. The genetic analysis demonstrated that the Belizeans, as well as other early individuals from Chile and Brazil, share a direct line of ancestry to the lineage of Anzick 1, a Paleoindian infant from southern Montana, which suggests a shared ancestry with Western Clovis culture peoples. The study also finds that these early Belizean individuals share alleles equally with modern indigenous people in the Maya area, lower Central America, and South America (Prufer et al. Reference Prufer, Alsgaard, Robinson, Meredith, Culleton, Dennehy, Magee, Huckell, Stemp, Awe, Capriles and Kennett2019). The authors argue that this finding is consistent with evidence for widespread population replacements throughout Central and South America by a distinct lineage, which began sometime prior to around 3450 cal b.c. This lineage, as represented by the individuals found in the rock shelter, represents a line of ancestors to all modern Maya groups.
THE EMERGING BIOLOGICAL PICTURE OF THE EARLIEST HUMANS FROM THE MAYA AREA
Though there are a growing number of individuals representing preceramic contexts in the Maya area (Table 1), most have only the most basic of descriptions. In particular, analyses of most individuals from the submerged cave sites are limited because of poor preservation and/or looting. In contrast, those from the Belizean rock shelters appear to be well-preserved, providing great promise for future research. Thus, our current model of biological variability of Paleoindian and Archaic groups in the Maya area must be considered preliminary, and any broader interpretations about the ancient populations from which they are derived highly speculative. However, when viewed in combination with the rich environmental and material culture data (reviewed in the other articles in this Special Section), we see a distinct biocultural picture of these groups beginning to emerge.
Studies of early skeletons from the New World, including those from the Maya area, have thus far focused primarily on debates over the origins of the earliest Americans. Specifically, while cranial and dental morphology of Holocene Native Americans closely resembles groups from northeast Asia, the shapes of crania pre-dating 10,000 years ago demonstrate a surprising amount of morphological diversity that in some cases more closely resembles phenotypes found in Australia, the Pacific Rim, and even Africa. Similarly, in a recent analysis of Naia's nonmetric dental traits, Cucina and colleagues (Reference Cucina, Vega Lizama and Chatters2021) found that while her teeth fit within Sinodont morphology (consistent with east Asian ancestry, rather than Pacific island Sundadont ancestry), the earliest North American individuals show great diversity that does not consistently fall within the Sinodont–Sundadont dichotomy. Their findings suggest that the peopling of the Americas involved (not surprisingly) complex microevolutionary processes that included gene flow, drift, and natural selection, but they further fit within a growing literature that disputes whether this dental typology accurately distinguishes these regional groups.
Analysis of craniofacial morphology of early central Mexican individuals shows similarities to several circum-Pacific populations, as well as to Paleoindians from Brazil and an Archaic sample from Colombia (González-José et al. Reference González-José, Neves, Lahr, González, Pucciarelli, Hernández and Correal2005). Chatters and colleagues (Reference Chatters, Allen and Jones2014) argue that Naia's cranial and dental morphologies do not clearly resemble those of modern Native American groups. Although a later morphometric reanalysis of Naia by Azevedo and colleagues (Reference Azevedo, Bortolini, Bonatto, Hünemeier, Santos and González2015) did note similarities to modern Native American morphology, this analysis is based on a two-dimensional image from Chatter et al.'s (Reference Chatters, Kennett, Asmerom, Kemp, Polyak, Blank, Beddows, Reinhardt, Arroyo, Bolnick, Malhi, Culleton, Luna, Rissolo, Morell-Hart and Stafford2014) publication, without taking into consideration potential distortion as a result of the water, and thus it should be viewed with caution. Regardless, it is safe to say that the morphology of early skeletons is diverse, and that the reasons for this variability are still being explored.
While this ambiguity in the cranial and dental data has fueled debate over the nature of the peopling of the Americas and the origin of the earliest migrants, the relatively recent incorporation of ancient DNA from Mesoamerica into these analyses has the potential to greatly inform this research. For instance, the recent genetic study by Posth and colleagues (Reference Posth, Nakatsuka, Lazaridis, Skoglund, Mallick, Lamnidis, Rohland, Nägele, Adamski, Bertolini, Broomandkhoshbacht, Cooper, Culleton, Ferraz, Ferry, Furtwängler, Haak, Harkins, Harper, Hünemeier, Lawson, Llamas, Michel, Nelson, Oppenheimer, Patterson, Schiffels, Sedig, Stewardson, Talamo, Wang, Hublin, Hubbe, Harvati, Delaunay, Beier, Francken, Kaulicke, Reyes-Centeno, Rademaker, Trask, Robinson, Gutierrez, Prufer, Salazar-García, Chim, Gomes, Alves, Liryo, Inglez, Oliveira, Bernardo, Barioni, Wesolowski, Scheifler, Rivera, Plens, Messineo, Figuti, Corach, Scabuzzo, Eggers, DeBlasis, Reindel, Méndez, Politis, Tomasto-Cagigao, Kennett, Strauss, Fehren-Schmitz, Krause and Reich2018) shows a clear link between early groups in southern Belize and Eurasian populations. Though mtDNA data from Naia have been reported by Chatters and colleagues (Reference Chatters, Allen and Jones2014), a later study identified at least two contamination events associated with their analysis and thus the results must be discounted (Prüfer and Meyer Reference Prüfer and Meyer2015).
There has also been some debate about whether the small sample of Mexican Lowland human skeletons from submerged caves form a genetically circumscribed regional population. Some studies (Herrera et al. Reference Herrera, Peart, Hernandez, Spradley and Hubbe2017; Hubbe et al. Reference Hubbe, Mata, Herrera, Benavente Sanvicente, González, Sandoval, Olguín, Núñez and Von Cramon-Taubadel2020) have found a significant amount of cranial morphological variability within this group, consistent with the genomic study by Posth and colleagues (Reference Posth, Nakatsuka, Lazaridis, Skoglund, Mallick, Lamnidis, Rohland, Nägele, Adamski, Bertolini, Broomandkhoshbacht, Cooper, Culleton, Ferraz, Ferry, Furtwängler, Haak, Harkins, Harper, Hünemeier, Lawson, Llamas, Michel, Nelson, Oppenheimer, Patterson, Schiffels, Sedig, Stewardson, Talamo, Wang, Hublin, Hubbe, Harvati, Delaunay, Beier, Francken, Kaulicke, Reyes-Centeno, Rademaker, Trask, Robinson, Gutierrez, Prufer, Salazar-García, Chim, Gomes, Alves, Liryo, Inglez, Oliveira, Bernardo, Barioni, Wesolowski, Scheifler, Rivera, Plens, Messineo, Figuti, Corach, Scabuzzo, Eggers, DeBlasis, Reindel, Méndez, Politis, Tomasto-Cagigao, Kennett, Strauss, Fehren-Schmitz, Krause and Reich2018), which showed that there were several dispersals likely involving different lineages over a period of perhaps 4,000 to 5,000 years. In contrast, González González and colleagues (Reference González González, Terrazas, Stinnesbeck, Benavente, Avilés, Rojas, Padilla, Velásquez, Acevez, Frey, Graf, Ketron and Waters2014:332) note the presence of cranial and facial similarities between several of the better-preserved individuals. Similarly, Stinnesbeck and colleagues (Reference Stinnesbeck, Rennie, Olguín, Stinnesbeck, González, Frank, Warken, Schorndorf, Krengel, Morlet and González2020) point out the consistently mesocephalic cranial morphology of all 10 early skeletons found from the Yucatan, which is distinct from the dolicocephalic cranial morphology of contemporaneous Paleoindians from Central Mexico, suggesting two different regional Paleoindian populations. Considering the extremely mobile nature of early hunter-gatherer groups, along with the current sample's time depth spanning several thousand years, it is difficult to draw any conclusive picture regarding patterns of regional genetic variability. Though methodological improvements may allow more complete sequencing of the poorly preserved DNA in the handful of skeletons currently available, this issue cannot be resolved without larger and more representative samples.
Pathology data have been reported for only a few of the better-preserved skeletons. These data have revealed interesting aspects of individual life histories and appear to be generally consistent with the data from previous analyses of Paleoindian groups, which typically characterize them as having short and brutal lives (Chatters Reference Chatters, Allen and Jones2014). For instance, trauma data are available for five of the individuals (three females and two males), and three display evidence of interpersonal violence, including healed bone fractures. Head traumas are particularly common, with four showing healed cranial contusions. In his survey of early skeletons from North America, Chatters also found that almost all of the trauma among Paleoindians occurred on males, and because almost all injuries were found on the upper, ventral body, he suggests they probably resulted from blows directed at the head. In contrast, he found very little evidence for trauma among females and of the three reported female cases in his study, none were head traumas. Two were on the back (suggesting they were fleeing) and one was Naia's broken arm. Chatters (Reference Chatters, Allen and Jones2014:82) interprets these sparse data as suggesting a pattern of “face-to-face violence between males and abuse of females”. Interestingly, the new data from Mexico show an even distribution—one female (Naharon I) and one male (El Pit I) had cranial trauma to the front of their heads, while the cranial trauma to Muknal occurred on the back of his head, and to the Chan Hol III female on the back and side of her head.
Other pathology data from a few of the individuals point to general health problems experienced by members of early groups. A rare instance of infectious disease reported for the Chan Hol III skeleton, in the form of severe alteration of the posterior parietal and occipital bones, is possibly the result of treponemal bacteria (Stinnesbeck et al. Reference Stinnesbeck, Rennie, Olguín, Stinnesbeck, González, Frank, Warken, Schorndorf, Krengel, Morlet and González2020). Porotic hyperostosis and cribra orbitalia provide evidence of metabolic disease on Naharon I and Muknal. Arthritis on the lumbar vertebrae of the 25- to 30-year-old Naharon skeleton, the young adult Chan Hol III skeleton, and on the appendicular skeletons of individuals from Saki Tzul and Mayahak Cab Pek suggests intense physical stress, and such early degeneration is typical of hunter-gatherers (Chatters Reference Chatters, Allen and Jones2014; Hill et al. Reference Hill, Moes, Trask, Kate, McKenzie, Edgar, Kennett and Prufer2018). Metabolic stress, as indicated by porotic hyperostosis and cribra orbitalia, is present on two individuals, and caries are present on Naia and on a couple of the remaining teeth of Chan Hol III and the Muknal individual, both of whom also showed antemortem tooth loss associated with periodontal disease. While these ailments are typically associated with agricultural communities, they have been reported in other Paleoindian skeletons (Chatters Reference Chatters, Allen and Jones2014; Cucina et al. Reference Cucina, Herrera and Chatters2019), suggesting a complex diet that included carbohydrates and sugars.
Stature estimates were available for three adult females (and for Naia, who was likely still growing) and these appear to be well within the range of other Paleoindians. At 152 cm, Las Palmas I matched the mean stature of 152.3 cm for Paleoindian females reported by Chatters (Reference Chatters, Allen and Jones2014:85; González González et al. Reference González González, Rojas, Acevez, Avilés, Analco, del Río, Luna, Velázquez, Stinnesbeck, Terrazas, Benavente, Leshikar-Denton and Erreguerena2008a:133), while at approximately 140 centimeters, Naharon I is slightly smaller than the shortest individual reported by Chatters (González González et al. Reference González González, Rojas, Terrazas, Benavente, Stinnesbeck, Aviles, de los Ríos and Aceves2008b:13). Chan Hol III, at 163 cm, is the tallest of these, and the second tallest in Chatters’ survey (Stinnesbeck et al. Reference Stinnesbeck, Rennie, Olguín, Stinnesbeck, González, Frank, Warken, Schorndorf, Krengel, Morlet and González2020). Naia is reported as being gracile and 149 cm +/− 4 cm (Chatters et al. Reference Chatters, Allen and Jones2014:Supplement).
While our current picture of diet is mostly based on interpretations of dental pathologies, a recent isotopic study was carried out on the 52 burials from the rock shelters in southern Belize that span the Late Paleoindian, Archaic, and Formative periods, providing a unique view of changes over time leading up to domestication (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020). The study found no evidence of maize consumption prior to 4,700 years ago, and then an increase in use over the next 700 years. By 4,000 years ago, maize was a staple, making up around 70 percent of the diet.
Finally, the complex mortuary treatments for some of these individuals are somewhat difficult to interpret, in large part because our current sample is very small and confined to two discrete regions. As a result, we do not know if the obviously intentional placement of many of these individuals in caves is an indication of special treatment. The lack of more human skeletons in cave systems that have been the focus of extensive exploration seems to indicate that burial in deep caves was rare. Further surveys and deep excavations of rock shelters, like those carried out in Belize, will no doubt turn up more early contexts.
CONCLUSIONS
This review identifies some intriguing themes among Paleoindian and Archaic skeletons in the Maya area. Continued work in the submerged caves of the Yucatan will no doubt identify new burials, while work being carried out in southern Belize has already produced burials from the Paleoindian and Archaic periods, which are being actively studied and soon will be published. However, given the time depth involved and the poor preservation that is typical of most Mesoamerican contexts, it is likely that sample sizes will remain small and that mostly isolated early skeletons will be geographically limited. Additionally, it should be emphasized that any summary statements about this group of individuals, who span a period of more than 10,000 years, is certainly oversimplified. Despite the detailed data derived from current and future analyses, none of these early individuals alone can stand as representative of what was surely a broad range of cultural and biological adaptations by ancient populations to the diverse environments comprising Late Pleistocene and Early Holocene Mesoamerica.
RESUMEN
Este artículo presenta una revisión de los restos óseos tempranos encontrados hasta ahora en el área maya que datan de los períodos paleoindio y arcaico. Mientras que son pocos en número, varios individuos encontrados en cuevas sumergidas en México y en abrigos rocosos en Belice han sido objeto de análisis intensivos, lo que ofrece un panorama emergente de la vida en la región antes de la transición a la agricultura.
ACKNOWLEDGEMENTS
Many thanks to Raúl Alejandro López Pérez for help with proofreading, and to Andrea Cucina, Keith Prufer, and two anonymous reviewers for their very helpful comments on earlier drafts of this article. Thanks also to Alejandro Terrazas and Mark Hubbe for allowing us to use their excellent map of the early sites in the Yucatan, and to Keith Prufer for the image of rock-shelter excavations in the Bladen Reserve.





 Open access
Open access
{kind=link}