


Summary
Mammals are among the most recognisable and most threatened organisms on Earth. There are more than 6000 living species, ranging from the big (rhinoceroses) to the small (rats), the wondrous to the weird (even venomous, too!) (Figure 4.1). While mammals face accelerating extinction risks as human populations grow and alter the landscapes and climates of the world, hope for survival remains through productive collaborations between scientists, governments and most importantly, local communities and stakeholders. Achieving conservation goals for terrestrial mammals means recognising that:
Our present-day extinctions actually started thousands of years ago!
Extinction filters and centres of risk have changed over time. Today, large-bodied mammals and those with small ranges face the greatest risk, and many of these species are found in the tropics, particularly Southeast Asia.
The most urgent threats include habitat loss and degradation, direct exploitation for bushmeat and trade, human–wildlife conflict, invasive species and infectious diseases. All of these threats intersect with each other and play out in the theatre of global climate change.
In the midst of a mass extinction, some mammals are recovering, and even thriving, and new species are still being described.
The biggest obstacle – and opportunity – for mammal conservation is our own behaviour.
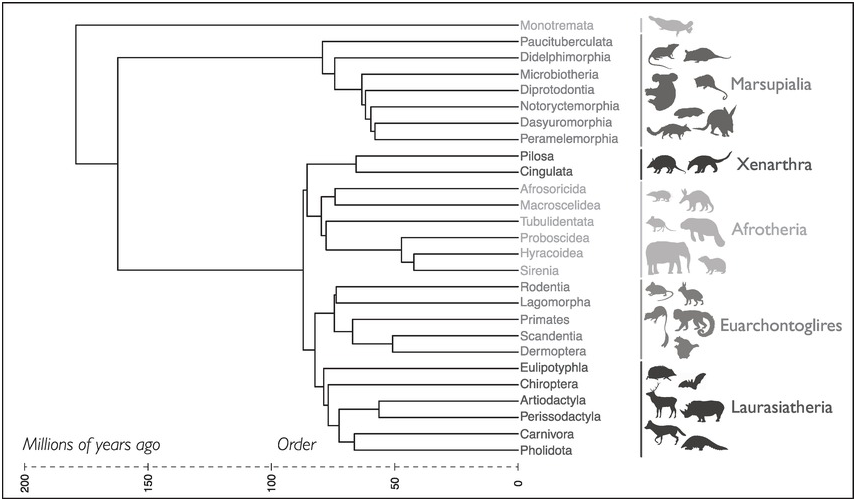
Figure 4.1 Evolutionary relationships of all living mammal groups by Order and Superorder, built using the VertLife mammal tree of 4098 species for which DNA sequences are available (Upham et al., Reference Upham, Esselstyn and Jetz2019). One species was chosen to represent each extant order (27 total) from the maximum clade-credibility topology of the fossil node-dated phylogeny.
The International Union for the Conservation of Nature (IUCN) considers species as threatened within the categories of Critically Endangered (CR), Endangered (EN) and Vulnerable (VU). Unfortunately, so many mammals fall within this elevated threat bracket that they are too numerous to list here. Of the nearly 5800 terrestrial mammals that have been evaluated, two are Extinct in the Wild (EW), 218 are CR, 515 EN, 530 VU, 345 Near Threatened (NT) and 3230 Least Concern (LC) (Figure 4.2). About 15% of all listed species are considered Data Deficient (DD), and these obscure species disproportionately come from the most speciose clades, including rodents and eulipotyphlans (true ‘insectivores’). IUCN recognises 81 species that have gone extinct since 1500 AD: numerous small mammals from island systems such as the Caribbean and the Galapagos, as well as more well-known species such as the thylacine or ‘Tasmanian Tiger’. CR species include a large number of well-known primates including gorillas (Figure 4.3A), gibbons and orangutans, as well as lesser known but equally threatened species including a wide range of rodents (e.g. the social tuco-tuco (Figure 4.3B) and marsupials (e.g. the mountain pygmy possum). A complementary framework for assessing species is the Evolutionarily Distinct and Globally Endangered (EDGE) scheme, which prioritises species whose extinction would represent a disproportionate loss of evolutionary history. The top scoring EDGE mammals are two rare species of long-beaked echidnas.
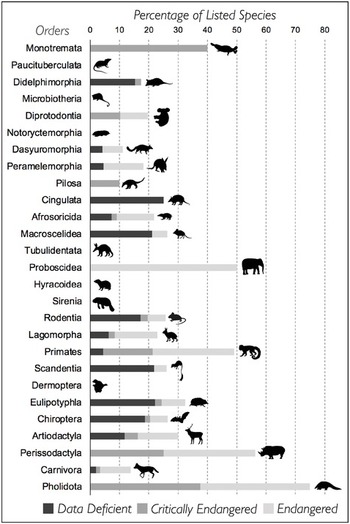
Figure 4.2 Percentage of mammal species, by order, listed as Data Deficient, Critically Endangered and Endangered by IUCN.

(A) Eastern gorilla, Gorilla beringei, Critically Endangered (K. Solari).

(B) Social tuco-tuco, Ctenomys sociabilis, Critically Endangered (J. Hsu).

(C) Jamaican flower bat, Phyllonycteris aphylla, Critically Endangered (A. Soto-Centeno).

(D) Giraffe, Giraffa camelopardalis, Vulnerable (K. Solari).

4.1 Introduction
We live in the ‘Age of Mammals’, a nickname for the Cenozoic Era, reflecting the rise of furry, warm-blooded creatures from below the feet of dinosaurs. The evolution of mammals is also a story of extinction: mammals diversified after the Cretaceous–Paleogene mass extinction approximately 66 million years ago (Upham et al., Reference Upham, Esselstyn and Jetz2019) (Figure 4.1). Closer in time, mammals themselves underwent massive extinctions starting in the Late Pleistocene (roughly the past 125,000 years) with the spread of humans worldwide, and such extinctions stretched through the Holocene (the past 10,000 years) as human populations grew and now continue into the present day.
Mammals are among the most recognisable organisms on the planet and receive a disproportionate share of research attention and conservation efforts among all vertebrates – perhaps because we ourselves are mammals! There are currently ~ 6400 mammal species currently recognised as alive today (Burgin et al., Reference Burgin, Colella, Kahn and Upham2018). While many species that come to mind are threatened, large-bodied and actively monitored (e.g. tigers, elephants, pandas), the majority are actually small, cryptic and understudied in their basic natural history. Large herbivorous mammals exert important effects on vegetation structure, fire regimes and carbon sequestration, and in turn, carnivores regulate the effects of these herbivores from the top down. Small mammals also play key ecological roles by consuming insects, dispersing seeds and influencing disease dynamics; for example, the Virginia opossum eats thousands of ticks a season! Therefore, the loss of mammals – large and small – has cascading effects on the ecosystems around them. These services affect humans, too: for millennia, mammals have been a source of food, labour and raw materials, and they also have aesthetic and religious values to many cultures. Yet today, these different values and uses across different human populations worldwide has led to conflict and decline for many species. The story of mammal conservation today is one of humans interacting with their fellow mammals, and both the problems and the solutions are in our hands.
What makes a mammal a mammal? Despite their wide phenotypic and ecological diversity, mammals are distinguished by several uniting features. Visibly, mammals have hair and mammary glands that produce milk; internally, there are numerous other structures, such as the anatomy of their inner ears and jaw bones, and the lack of nuclei in their blood cells. Mammal groups vary in their number of species, geographic distribution and evolutionary history, presenting conservation challenges and tradeoffs. In this chapter, we will consider the mammals typically featured for conservation, but also make a special effort to highlight the full diversity of mammals, including the cryptic, rare and quite frankly, weird species.
There are three major evolutionary groups of mammals living today (Figure 4.1):
Monotremes. While they lay eggs and lack nipples, it would be incorrect to describe them as ‘primitive’, as they have been evolving for millions of years to fulfill their own specialised lifestyles, including being some of the few venomous mammals on the planet. There are five known living species of Monotremes (Order Monotremata): four echidnas (Family Tachyglossidae) and one platypus (Family Ornithorhynchidae); these two families diverged nearly 50 million years ago from each other. Monotremes are found in Australasia, including Australia and New Guinea. The western long-beaked echidna and Attenborough’s long-beaked echidna are the two highest ranked Evolutionarily Distinct and Globally Endangered (EDGE) species alive today.
Marsupials. The term marsupial derives from having a marsupium, or pouch, though many species don’t have one. Marsupials are characterised by giving birth to relatively undeveloped young after a short gestation period. There are approximately 330 species spread across seven orders: Didelphimorphia, Paucituberculata, Microbiotheria, Dasyuromorphia, Peramelemorphia, Notoryctemorphia and Diprotodontia (Figure 4.1). These species range from the iconic kangaroos, wombats and koalas to the lesser known monito del monte, the only species in an entire order found in southern South America, and the recently extinct ‘Tasmanian tiger’ or thylacine. While their centre of diversity today is Australasia, the group originated in the Western Hemisphere and are found widely in South America. The only marsupial to live in North America is the Virginia opossum.
Placentals. The majority of extant mammals, >5000 species, are placental mammals, named for the distinctive features of their placenta, though there are also metabolic and morphological differences between placental and other mammals. After much debate between conflicting morphological and molecular datasets, the major groups (Superorders) of placental mammals have finally been resolved: Afrotheria (elephants, hyraxes and others), Xenarthra (sloths, armadillos, anteaters), Laurasiatheria (carnivores, ungulates, bats and others) and Euarchontoglires (rodents, primates and others) (Figure 4.1). Placental mammals are found across all continents except Antarctica. The most diverse groups include rodents, bats and eulipotyphlans (true ‘insectivores’).
4.2 A Long Legacy of Extinction and Survival
We are in the midst of an extinction event on par with the ‘Big Five’ mass extinctions of the distant past: a geologic event unfolding on ecological timescales, making it difficult to see how losses can be replaced in the near future (‘extinction without replacement’). For example, recent bat extinctions in the Caribbean would require 8 million years to recover the evolutionary diversity that has been lost (Valente et al., Reference Valente, Etienne and Dávalos2017). Present-day mammal communities represent survivors that have already passed through several extinction filters, and the legacy of the past shapes our perception of present-day mammal diversity and extinction risk (Turvey and Fritz Reference Turvey and Fritz2011). Data points of past abundance represent baselines that can help us trace the decline of species over time. For example, in North America, records of the number of pelts collected during the historical fur harvest industry change how we perceive the present-day population trends (and thus conservation needs) of fur-bearing mammals: population declines of several species are significantly greater when viewed from a baseline of 1850 AD, rather than 1970 AD (Collins et al., Reference Collins, Böhm and Collen2020). Understanding this longer history of extinction – encompassing the Late Pleistocene, Holocene and more recent historical record, with changing human pressures across all three intervals – is necessary to accurately contextualise present-day conservation priorities and reveal hidden opportunities. This view from the past also makes us appreciate that the mammals on the landscape today are in fact survivors, presenting a narrative of hope.
4.2.1 Living in the Shadow of Giants: Late Pleistocene Megafauna Extinctions
Fossil discoveries in the nineteenth century led Alfred Russel Wallace to observe that ‘we live in a zoologically impoverished world, from which all the hugest and fiercest and strangest forms have recently disappeared’. The present-day landscape of mammal diversity and the ecosystems they inhabit were shaped by events of the Late Pleistocene, which included major climatic changes of glacial–interglacial cycles and, of paramount importance, the evolution of modern humans and their spread across the globe. Megafaunal mammals (those >45 kg or ~100 lbs) disproportionately went extinct at the end of the Late Pleistocene on continents from the tundra to the tropics, severely reducing the diversity of mammalian groups including sloths, proboscideans, cingulates (e.g. armadillos), ungulates and carnivores (Smith et al., Reference Smith, Smith, Lyons and Payne2018). Many now-threatened species began contracting their ranges during the Late Pleistocene, such as the giant panda, Malayan tapir and tiger in Southeast Asia (Louys, Reference Louys2012). More than 100 genera of megafaunal mammals went extinct in total, with magnitudes of loss varying by continent, with some like South America losing >80% of genera (Smith et al., Reference Smith, Smith, Lyons and Payne2018). Debate still rages over the cause of these losses: was it natural climate change, the arrival of humans, or some synergy of the two? Understanding these ancient extinction forces can yield helpful clues for our present conservation dilemmas, and this is a very active area of research.
4.2.2 The Geography and Selectivity of Extinction
Today, more than 1000 species, or roughly a quarter, of all IUCN assessed mammals are threatened with extinction, with body size and geographic range size acting as the most important predictors of risk (Cardillo et al., Reference Cardillo, Mace and Jones2005). A body size of 3 kg (~7 lbs) appears to be the threshold for which species begin to face multiple, compounding extinction threats. Species smaller than 3 kg are most at risk based on the human impacts they are exposed to in their geographic range (extrinsic risks), whereas those larger than 3 kg are additionally vulnerable to decline given their slow life history traits, including a low reproductive rate and long life span that make it difficult for populations to rebound (intrinsic risks). Given this dichotomy, in general, smaller mammals may benefit from a threatened areas or hotspot approach, whereas larger mammals can require approaches that are species-specific (Cardillo et al., Reference Cardillo, Mace and Jones2005).
A small geographic range can predispose species to be vulnerable to catastrophic events, and in general, extinction risk is higher for species with ranges less than 1 million km2 (Davidson et al., Reference Davidson, Shoemaker and Weinstein2017). The CR social tuco-tuco (a rodent) has an area of occupancy of only 8 km², and the 2011 eruption of the Puyehue-Cordón Caulle volcano near Patagonia nearly halved its population density (Hsu et al., Reference Hsu, Crawford and Tammone2017) (Figure 4.3B). Species with the smallest ranges are disproportionately found on islands (e.g. Madagascar, Sri Lanka and Sulawesi) and tropical mountains (e.g. Andes, Cameroonian and Ethiopian Highlands) (Schipper et al., Reference Schipper, Chanson and Chiozza2008). Some examples include the EN pygmy sloth, found only on the wildlife refuge Isla Escudo de Veraguas of Panama, the EN Nicobar island tree shrew (Order Scandentia) from the Great and Little Nicobar Islands, the EN Seram bandicoot, endemic to the island of Seram in Indonesia, and known only from seven specimens collected in 1920 at a single locality, and the CR Attenborough’s long-beaked echidna, found on a single mountain (Berg Rara) in the Cyclops Mountains of New Guinea.
Extinction risk for mammals has changed over time both spatially and across the tree of life (Turvey and Fritz Reference Turvey and Fritz2011). While in the Holocene, many extinctions were concentrated on islands, today, the largest concentrations of threatened mammals are found in South and Southeast Asia, the tropical Andes in South America, the Cameroonian Highlands and Albertine Rift in Africa, and the Western Ghats in India. These regions share the common feature of being at the intersection of intense anthropogenic pressures, rapidly growing human populations and high species richness, including those species with restricted ranges (Schipper et al., Reference Schipper, Chanson and Chiozza2008; Davidson et al., Reference Davidson, Shoemaker and Weinstein2017). Geographic patterns of risk can differ for different taxa. Large carnivores, such as lions and tigers, have experienced the greatest range contractions across Southeast Asia and Africa (Wolf and Ripple, Reference Wolf and Ripple2017), whereas rodent ‘threat spots’ include the Philippines, New Guinea, Sulawesi, the Caribbean, Chinese temperate forests, the Brazilian Atlantic Forest, and drylands including Togo and the Mongolian–Manchurian steppes (Amori and Gippoliti, Reference Amori and Gippoliti2001).
4.3 Conserving Mammals in a Changing World
As we have seen, correlates of extinction risk and the location of geographic hotspots are not static but can change over time in response to different human and environmental conditions. Mammals in the Anthropocene face multiple global extinction threats that intersect with local ecological, cultural and socioeconomic conditions. Here we identify habitat loss, direct exploitation, human–wildlife conflict, invasive species and disease as the major Anthropocene threats facing mammals, all playing out in the overarching context of climate change.
4.3.1 Habitat Loss, Degradation and Fragmentation
By far the greatest immediate threat to mammalian biodiversity is the loss, degradation and fragmentation of habitat, often linked to agricultural and urban expansion. This affects 40% of IUCN assessed mammal species (Schipper et al., Reference Schipper, Chanson and Chiozza2008). The ecological and behavioural needs of primates make them particularly vulnerable to habitat loss, which is the main driver of threat for species in primate hotspot countries including Brazil, Madagascar and Indonesia (Estrada et al., Reference Estrada, Garber and Mittermeier2018). Under a worst-case scenario of agricultural expansion in the twenty-first century, primate ranges will severely contract by ~60–80% across these countries (Estrada et al., Reference Estrada, Garber and Mittermeier2018). Illegal mining of precious metals and gems has affected many forests in Madagascar, the only place in the world where lemurs are found today (nearly 100 species of them!) (Estrada et al., Reference Estrada, Garber and Mittermeier2018).
Mammals in Southeast Asia face devastating rates of habitat loss due to forest clearance for agriculture, international logging concessions, and oil palm and other agroforestry crops (e.g. rubber, biofuels, wood pulp) (Hughes, Reference Hughes2017). These causes of deforestation, combined with mining and dam construction, have destroyed more than 50% of the original forest cover in some places (e.g. the Philippines, parts of Indonesia), and up to 98% original forest loss is projected for some countries in the next decade (Hughes, Reference Hughes2017). It is estimated that more than 100,000 individual CR Bornean orangutans (half the population) were lost due to logging, deforestation or the construction of industrialised plantations from 1999–2015 (Voigt et al., Reference Voigt, Wich and Ancrenaz2018). Habitat loss in this region also affects smaller, lesser known species such as the EN flat-headed cat, which is rapidly losing its native Indonesian wetlands and lowland forest habitats to oil palm plantations and clearance of coastal mangroves (Wilting et al., Reference Wilting, Cord and Hearn2010).
Many small mammals are also impacted by habitat loss, particularly those whose already small ranges unfortunately intersect with anthropogenic activities. Numerous shrews (Order Eulipotyphla) are threatened by habitat destruction, including the CR Nelson’s small-eared shrew of Veracruz, Mexico and the CR Jenkin’s shrew of India’s Andaman Island. Their convergent cousins, the afrosoricid insectivores (Orders Afrosoricida and Macroscelidea) (Figure 4.1), are similarly threatened by habitat loss. The EN golden-rumped sengi relies on Kenyan forests with abundant leaf litter, such as the Arabuko-Sokoke Forest, which has been degraded by tree felling. Perhaps surprisingly, of the 21 known species of golden moles (Family Chrysochloridae), more than half are listed at an elevated threat level due to urbanisation, mining and agriculture, including the possibly extinct De Winton’s golden mole of South Africa. Even bats, which are more vagile due to their flight capabilities, can vary in sensitivity based on roosting behaviours. Two CR bat species, the Jamaican flower bat (Figure 4.3C) and the Jamaican greater funnel-eared bat, are now each restricted to a single remaining cave roost, demanding urgent conservation attention to avoid the loss of their entire species.
4.3.2 Direct Exploitation
Humans exert direct pressure on mammal populations by removing wild individuals for food, trade, traditional medicine and sport, all within the context of a globalising economy and changing cultural norms. Such direct exploitation currently threatens more than 300 species of terrestrial mammals (Ripple et al., Reference Ripple, Chapron and López-Bao2016). Those species threatened by overharvesting are typically large bodied and charismatic, such as elephants, primates and rhinoceroses, and their selective removal from an ecosystem can result in ‘empty forest syndrome’, creating landscapes devoid of large mammals. The drivers of direct exploitation are complex, and solutions require strengthening legal protections coupled with increased support of local communities in ways that do not recapitulate colonial power dynamics. Efforts to increase global awareness and change attitudes, such as World Pangolin Day, and emerging technologies in wildlife crime forensics are additional tools in the conservationist’s toolbox.
Humans have been consuming mammals for millennia, with outcomes ranging from extinction to sustainable harvest. However, commercialisation, global supply chains and easier access to firearms have fundamentally altered these traditional practices; for example, drug cartels are now involved in poaching due to its profitability. Bushmeat hunting (in contrast with legal, regulated hunting) is a major conservation challenge because it is linked to economic inequality, food insecurity and cultural norms, and conservation responses must include alternative livelihoods and protein sources. Mammalian orders with the most species threatened by hunting include primates, bats, carnivores, even- and odd-toed ungulates, and rodents (Ripple et al., Reference Ripple, Chapron and López-Bao2016). Civil wars and illegal mining in many primate-rich areas, such as the DRC, have led to elevated bushmeat exploitation affecting numerous species including CR Eastern lowland gorillas (Figure 4.3A). Popular as trophy hunting prizes, giraffes (Figure 4.3D) in Uganda have alarmingly declined by 90% over 30 years, and more than 39,000 giraffe derivatives were imported into the US from 2006–2015 (Brown et al., Reference Brown, Bolger and Fennessy2019). Small mammals are also exploited for both their meat and fur. The Russian desman, an aquatic mole (Family Talpidae), was overharvested for its water-resistant fur throughout the 1800s until the Soviet government enacted a hunting moratorium in 1957 (Rutovskaya et al., Reference Rutovskaya, Onufrenya and Onufrenya2017), giving the species a chance to recover.
International trade is formally controlled by the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES). More than 800 mammal species are currently regulated by CITES, but many species are traded illegally and smuggled across borders in response to global economic forces. For example, demand for ivory in Chinese markets drives poaching of African elephants (Figure 4.3E) (Hauenstein et al., Reference Hauenstein, Kshatriya, Blanc, Dormann and Beale2019). Pangolins (Order Pholidota) are an unfortunate icon of the international illegal wildlife trade and, as the most trafficked mammal on the planet, have been described as literally being eaten to death. Pangolins are often called scaly anteaters due to their keratin scales and specialised diet of ants and termites, and are found across Africa and Asia. While they have been consumed on a local scale for decades, they are now highly sought after in the Chinese Traditional Medicine market and sold as delicacies in restaurants, resulting in African species being shipped for consumption and use in Asia. Recent estimates suggest that the equivalent of more than 895,000 pangolins were trafficked globally between 2000 and 2019 (Challender et al., Reference Challender, Heinrich, Shepherd, Katsis, Challender, Nash and Waterman2020). All pangolin species were listed in the CITES Appendix I at CoP17 in 2016, establishing an international trade ban, though conservation solutions must target changing attitudes and norms in Asian markets to be successful.
Many mammals are traded as pets and for use in entertainment. Hundreds of thousands of primates are thought to be traded as pets each year, including the unfortunate viral video star, the CR Javan slow loris (Nijman et al., Reference Nijman, Nekaris, Donati, Bruford and Fa2011). There are more tigers in captivity in the United States than in the wild, yet less than 5% of the estimated 10,000 tigers are held within accredited facilities, and many are severely inbred (Nyhus et al., Reference Nyhus, Tilson, Hutchins, Tilson and Nyhus2010). White tigers are a particularly egregious case of genetic mismanagement, as the inbreeding required to produce a white coat is accompanied by birth defects including club feet and crossed eyes (Figure 4.4A). Raising awareness of the ethical issues surrounding captivity can lead to meaningful policy change through public pressure on governments, such as the continued amendments strengthening the Animal Welfare Act in the United States, though such protections still do not apply to less broadly charismatic species (e.g. rodents).

(A) Kenny, a captive white tiger (Panthera tigris), was born with skull and jaw malformities due to inbreeding. While most tigers with this appearance would have been euthanised, Kenny was rescued by Turpentine Creek Wildlife Refuge (https://www.turpentinecreek.org/).

(B) Sacred sika deer (Cervus nippon), with antlers removed, peruse a store and are tolerated by the shop owner in Nara, Japan (A. Mychajliw).

(C) A female Hispaniolan solenodon (Solenodon paradoxus) killed by a dog near Puerto Escondido, Dominican Republic – extended teats suggest that she was nursing (A. Mychajliw).

(D) A pika (Ochotona roylei) in Spiti Valley, Himachal Pradesh, India, stuck between a rock and a hard place – rising temperatures and high elevation hypoxia (K. Solari).

(E) Despite their abundance in places such as Costa Rica, Xenarthrans such as the northern tamandua (Tamandua mexicana) remain understudied (K. Solari).
4.3.3 Human–Wildlife Conflict
As human populations grow, so does the potential for human–wildlife conflict through competition for the same resources, such as prey or habitat, or when wild mammals are perceived as a threat to the safety of humans or their property. Often, contentious wildlife management decisions are made in the shadow of political tensions and the power dynamics of human communities, such as the massive battles between ranchers and conservationists over grey wolves in Yellowstone National Park, United States. In North America and Europe, years of carnivore absence have led to a human population that lacks the social norms for responsibly behaving in places where carnivores are returning, leading to increased attacks reported in the media, which can reinforce negative attitudes (Penteriani et al., Reference Penteriani, del Mar Delgado and Pinchera2016). Because human–wildlife conflicts often centre on perceived, rather than real, threats and are enmeshed in complex sociopolitical landscapes, social science research is critical for conservation progress on this front.
Humans are considered ‘super predators’ – both consuming prey and killing off other predators to avoid competition. This manifests most clearly in the conflict associated with livestock predation by wild mammalian carnivores, which often results in retaliatory killings of the offending (or more often, suspected) individual. Carnivores are increasingly co-occurring with humans and livestock in shared spaces, particularly in the Global South where people rely directly on livestock and multiuse forests for their livelihoods. Government support for predator control and eradication has resulted in several historical carnivore extinctions and massive range extirpations, such as the range reduction of wolves in Asia, North America and Europe, lions and wild dogs in Africa, tigers in Asia and jaguars in the Americas (Wolf and Ripple, Reference Wolf and Ripple2017). Historic carnivore extinctions associated with hunting bounties to ‘protect’ livestock include the thylacine, the Hokkaido wolf of Japan, the California grizzly bear and the Falkland Islands wolf. While common approaches to preventing livestock depredation include lethal control, other options can include translocation of problematic individuals, construction of barriers and the use of trained guard dogs. Governments and non-governmental organisations can also offer compensation to farmers who have lost individuals to predation events to ameliorate feelings of resentment and support livelihoods, despite economic losses.
Carnivores are not the only species that may conflict with humans. Given their massive size, elephants often have major impacts on villages and agriculture, with records of crop raiding in Asia dating back centuries. In Africa, strategies to avoid conflict have taken advantage of elephants’ natural fear of bees. Farmers who place beehives along their fences can simultaneously protect their crops and generate new sources of income, creating a conservation win-win scenario (Ngama et al., Reference Ngama, Korte, Bindelle, Vermeulen and Poulsen2016).
While humans and wildlife conflict with certain cultural practices, others can promote conservation. Temples and shrines have served as important places for positive human–wildlife interactions for hundreds of years. The sacred herd of sika deer at the Kasuga-taisha Shrine in Nara, Japan (a UNESCO World Heritage Site) attracts thousands of visitors annually, generating income for local communities through tourism and the sale of special crackers (鹿煎餅, shika-senbei) for feeding the docile deer (Figure 4.4B). This particular deer herd has been protected for hundreds of years; killing of a deer was punishable by death up until the mid 1600s, and the deer remain protected under national treasure status, even as populations grow and spill outside of the shrine’s perimeter onto city streets. Bucks’ antlers are proactively removed each year to avoid injuries to humans, especially during mating season when males become aggressive. This relationship formed over hundreds of years provides insight into the strategies – both from the biological and social sciences – that do and don’t work in facilitating co-existence.
4.3.4 Introduced and Invasive Species
Invasive species can impact mammals directly through predation, or indirectly, by competing with species for shared resources or altering ecosystem structures, leading to habitat degradation and loss. While mammals are highly threatened by the activities of introduced species, they themselves actually represent some of the most destructive invasive species globally: black and brown rats, feral cats and dogs, mongoose and domestic livestock, among others. These invasive mammals have insidiously significant and wide-ranging effects on island species. The numerous historical extinctions of Caribbean small mammals, including an entire endemic family of island shrews (Order Eulipotyphla), have been linked to the introduction of mongooses, cats and dogs associated with European colonial practices (Turvey et al., Reference Turvey, Kennerley, Nuñez-Miño and Young2017a). Species such as the EN Hispaniolan hutia and the EN Hispaniolan solenodon (Figure 4.4C) remain severely threatened by predation, though recovery could be feasible where dog populations are controlled (Turvey et al., Reference Turvey, Kennerley, Nuñez-Miño and Young2017a).
Australian mammals – both marsupial and placental – are particularly threatened by non-native predators. Similar to the Caribbean, feral cats have been proposed as the cause of more than 20 Australian endemic mammal extinctions historically, including the lesser bilby and the entire family of pig-footed bandicoots (Family Chaeropodidae) (Woinarski et al., Reference Woinarski, Burbidge and Harrison2015). This is why many conservationists suggest that if you have a pet cat, you should keep them inside to protect native species. Many of Australia’s threatened mammals now occur only on small islands or fenced enclosures, and these predator-free environments represent important places for reintroductions. The EN numbat (Family Myrmecobiidae), a termite specialist, nearly disappeared in the 1970s, but experimental fox control programmes showed significant numbat recovery potential. As a result, numbats are now successfully bred in captivity for reintroduction to predator-free reserves across southwestern Australia and translocated to two fenced mainland islands in eastern Australia.
New archaeological research continues to deepen our understanding of just how long humans have been moving species outside of their native ranges. In fact, the earliest documented translocation occurred 20,000 years ago (Hofman and Rick, Reference Hofman and Rick2018)! Such studies paint a complex picture of the underappreciated or unrecognised human components of mammal histories, including species we, perhaps ironically, now protect today. Dingoes were brought to Australia by people more than 3000 years ago and likely had significant ecological effects on native species, potentially precipitating the extirpation of Tasmanian devils from Australia. Similarly, Indigenous peoples likely introduced foxes to the previously canid-free California Channel Islands ~7000 years ago, eventually evolving into the ‘endemic’ and beloved island fox (Hofman and Rick, Reference Hofman and Rick2018). These examples reflect the truly coupled long-term history of humans and other mammals and are a window into what co-existence looks like over hundreds and even thousands of years.
4.3.5 Infectious Diseases and Spillovers
Our increasingly interconnected world is putting species together in new ways, in turn allowing them to share parasites, viruses, bacteria and other agents of disease across biogeographic boundaries. The Christmas Island rat, which went extinct around 1900, represents the earliest mammal extinction directly attributable to a human-mediated disease, as invasive ship rats were accompanied by pathogenic trypanosomes (Wyatt et al., Reference Wyatt, Campos and Gilbert2008). More generally, the expansion of domestic mammals and bushmeat consumption, changing habitat conditions, due to deforestation, and shifting climates are all working to reshape the disease landscapes that species inhabit and lead to increasing disease spillovers of relevance to wildlife and human health.
The global popularity of domestic dogs is accompanied by an increase in the extent and transmission of canine distemper virus to wild canids, such as the EN Ethiopian wolf (Marino et al., Reference Marino, Sillero-Zubiri and Deressa2017) and can impact numerous carnivore species aside from canids if not managed through vaccination. Plague, introduced to US shipping ports by invasive rodents in 1900, decimates immunologically naive black-tailed prairie dog populations, which are the main food item for EN black-footed ferrets. As a result, the ferrets are hit with a one-two punch of prey depletion and direct plague mortality (Salkeld, Reference Salkeld2017). The oral vaccine for plague can be delivered to prairie dogs in the form of peanut-butter-flavoured baits, but such dissemination requires collaboration between US federal agencies and private landowners, which can be contentious. Yet, hope for combating such diseases comes from the case of rinderpest, which once spilled over from domestic livestock and caused massive mortality events in African ungulates, but is now considered to be globally eradicated (Morens et al., Reference Morens, Holmes, Davis and Taubenberger2011).
Bats are often associated with zoonotic diseases in the minds of the public due to viral spillovers including Hendra, Nipah, Ebola, Marburg and now, COVID-19. These spillover events occur as a result of habitat loss and degradation, which changes bat behavioural patterns and species interactions. Hunting of bats and other co-occurring mammals for bushmeat further increases the likelihood of transmission to humans (Schneeberger and Voigt, Reference Schneeberger, Voigt, Voigt and Kingston2016). Humans have also facilitated the spread of diseases affecting bats, such as the deadly white-nose syndrome (WNS) in North America, a fungus that infects the skin and causes bats to wake up too frequently during hibernation (Schneeberger and Voigt, Reference Schneeberger, Voigt, Voigt and Kingston2016). WNS has killed millions of bats since its introduction to a New York cave by tourists in 2006 and prompted the listing of several mouse-eared bat species under the US Endangered Species Act due to imminent threat of disease-related extinction. From this hard lesson, we have learned the vital importance of sterilising equipment and other precautions to avoid bringing pathogens from one location to another, whether in research or tourism.
Given their close relationship to us, non-human primates are particularly vulnerable to zoonotic diseases transmitted by humans. Ebola outbreaks killed over 90% of western gorillas (Figure 4.3A) and 80% of chimpanzees in the Lossi Sanctuary of Republic of Congo between 2002 and 2004 (Bermejo et al., Reference Bermejo, Rodríguez-Teijeiro and Illera2006). In Kibale and Gombe Stream National Park of Tanzania – where Jane Goodall famously worked – human respiratory viruses are the leading cause of death for chimpanzees. In the new era of COVID-19, researchers must be extremely vigilant to avoid spillover of this highly infectious disease to the very non-human primate communities they are trying to protect.
4.4 Climate Change: Move, Adapt or Go Extinct
Anthropogenic climate change is transforming our planet and no mammal species is outside the reach of its impacts. Globally, hotspots of climate change risk for mammals include areas in Africa (northeastern South Africa, northeastern Tanzania, southwestern Kenya), the Yunnan region of China, the mountains of New Guinea and the western Amazon Basin in South America (Pacifici et al., Reference Pacifici, Visconti and Butchart2017). In the past, species responded to natural climate change by either adapting to stay in place or moving to a new place to track their preferred ecological conditions; if neither of those options were feasible, the outcome was extinction. Today, these survival options are even more complicated: adaptation may be hindered by a population’s low genetic diversity resulting from other threats (e.g. fragmentation and inbreeding) or may not be possible given how rapidly temperatures are rising relative to reproductive rates. Range shifts may not be possible because of anthropogenic barriers such as habitat conversion, roads and fences. As species continue to shift outside of protected areas and move across political boundaries, planning in the context of climate change requires conservation innovation and international collaboration. In the Western Hemisphere, anywhere from ~9–39% of mammals at a given location may not be able to shift their ranges to match the velocity of suitable climate conditions changes (Schloss et al., Reference Schloss, Nuñez and Lawler2012), and these impacts are worsened for dispersal-limited taxa, including primates, shrews and moles. Such species may benefit from a controversial but promising approach of assisted colonisation, in which managers translocate populations outside of their current range in anticipation of climate impacts.
Anthropogenic climate change is accompanied by an increased frequency of extreme weather events. In Mexico, threatened species including two subspecies of spider monkeys, two howler monkeys and the silky anteater are all vulnerable to population mortality events associated with hurricane exposure (Ameca et al., Reference Ameca, Mace, Cowlishaw and Pettorelli2019). In the Pacific Islands, increasingly frequent tropical cyclones have devastated insular bat populations (80–90% of flying fox populations) through direct mortality and alteration of resources (Scanlon et al., Reference Scanlon, Petit, Tuiwawa and Naikatini2018).
Small mammals with restricted ranges on islands, mountains or already arid areas are extremely sensitive to climate change. The Bramble Cay melomys, a rodent, is now considered the first mammal extinction caused directly by anthropogenic climate change (Waller et al., Reference Waller, Gynther, Freeman, Lavery and Leung2017). The species was formerly restricted to Bramble Cay (Maizab Kaur), an ~4 ha sand cay located in Australia’s Torres Strait. This low elevation and small range made it vulnerable to sea-level rise and storm surges, which destroyed its habitat and also caused direct mortality (Waller et al., Reference Waller, Gynther, Freeman, Lavery and Leung2017). Pikas (Order Lagomorpha) (Figure 4.4D) are cold-temperature specialists distributed across mountain ranges in Asia and North America. The impact of climate change on pikas is twofold, placing pikas in a vice between two different selective pressures: at lower elevations, warming temperatures (hyperthermia) may be lethal, and at higher elevations, low oxygen levels (hypoxia) may be lethal. American pikas are already climbing higher in elevation in the US Great Basin at a rate of ~145 m per decade (Beever et al., Reference Beever, Ray, Wilkening, Brussard and Mote2011). Genetic research provides us with some hope, luckily – pikas may be able to flexibly alter how they express genes related to oxygen use, permitting them to occupy somewhat higher elevations (Solari et al., Reference Solari, Ramakrishnan and Hadly2018).
Climate change is not purely its own force, but rather intersects with and amplifies all other ongoing threats to mammals in the specific local contexts where they live. Large mammals that migrate may need to alter their migration pathways in response to climate change. Caribou live at high latitudes in circumpolar regions of Europe, Siberia and North America, and play critical roles in the livelihoods of Indigenous peoples. The boreal caribou, a subspecies of caribou in Quebec, Canada, will have its migratory movements reduced due to climate change in a way that exacerbates existing fragmentation due to road construction. Positively, the removal of secondary roads inside protected areas could, however, offset these losses as a compensatory conservation action and benefit the subspecies (Bauduin et al., Reference Bauduin, McIntire, St-Laurent and Cumming2018). Climate change also influences the potential geographic ranges and activity periods of pathogens and their vectors. In Kazakhstan, the shocking death of >200,000 individual CR saiga (a type of antelope) over a three-week period in 2015 was due to an outbreak of haemorrhagic septicemia during a calving aggregation. This disease is caused by the bacterium Pasteurella multocida, which is normally endemic in saiga, but lethal levels were caused by abnormally high temperatures and humidity (Kock et al., Reference Kock, Orynbayev and Robinson2018).
4.5 The Future of Mammals
If we do not act, the future of mammal communities will look very different from the recent past. Assuming all threatened mammals eventually go extinct, ~20–50% of all mammals will have been lost since 125,000 years ago, and the largest mammal on the planet in a few hundred years may very well be a domestic cow, representing the lowest average body mass for terrestrial mammals in 45 million years (Smith et al., Reference Smith, Smith, Lyons and Payne2018). But, with active management efforts that take into account the needs of both local communities and wild mammals, coupled with increasingly effective captive breeding strategies, there is hope for preserving many now-threatened species and even for discovering previously cryptic species with new genetic and monitoring tools.
4.5.1 There’s More to Discover
Despite being among the most well studied of any taxa, many mammal species are insufficiently assessed by IUCN and therefore are ‘Data Deficient’, hindering our projections of conservation need (Figure 4.2). Spatial and taxonomic biases in international research agendas, often linked to funding priorities, means that many threatened species have not been assessed and may be lost before we can even appraise their status. Conversely, this also means that there may be species who are faring better than expected! While there is good research coverage of primate and carnivores – likely linked to their charismatic appearances and behaviours – small mammals such as shrews and rodents are poorly studied. For example, nearly 20% of the 2000 species of rodents, the 100 species of didelphimorphid marsupials and the 500 species of eulipotyphlans are Data Deficient (Figure 4.2). An entire Superorder, Xenarthra (Figure 4.4E), suffers from a lack of basic field-based natural history studies, and new genetic and morphological studies continue to expand the number of small-range endemic species.
4.5.2 Not Extinct Yet: Just in the Wild
IUCN currently lists two mammals as Extinct in the Wild (EW). While it is possible that these species will go extinct, they represent major opportunities for conservation innovation in developing tools for bringing species back from the brink. And it has been done before: through captive breeding of just 12 wild individuals, the Przewalski’s horse, or takhi, moved from EW in the 1960s to being listed as EN today, with nearly 200 individuals spread across reintroduction sites in the Gobi steppe near the Mongolian–Chinese border.
The two currently EW species are both ungulates. The scimitar-horned oryx, an antelope adapted to arid areas that once ranged across North Africa, nearly disappeared by the 1990s. Today there are more than 15,000 individuals in captivity, many of which are thought to have resulted from a founder population of ~50 individuals from Chad (Woodfine and Gilbert, Reference Woodfine, Gilbert, Bro-Jorgensen and Mallon2016). Studbook-managed breeding programmes and trial reintroductions into fenced areas in Morocco, Chad, Senegal and Tunisia are showing signs of success. The Père David’s deer was formerly endemic to China and its survival shows just how much luck can be involved in dodging extinction. The species first became known to Western scientists in 1864, when the French missionary Père David encountered a small herd in the Nanyuan Royal Hunting Garden south of Beijing. Unfortunately, the remaining 20–30 individuals were eaten by soldiers during the 1900 Boxer Revolution – but (very!) luckily, several deer had been sent to Europe a few years prior, and the very last 18 deer in existence formed a breeding herd in England. Despite inbreeding, the population grew, and the first captive animals were sent to the Beijing Zoo in 1956, and there are now >2000 individuals in China across fenced areas of Beijing, Dafeng, Shishou and Yuanyang. This species was perhaps both imperilled but also saved by its cultural importance as a symbol of high status for people in China (Turvey et al., Reference Turvey, Barnes, Marr and Brace2017b), with a history of management from the fifth century BCE that now stretches into the present day.
Even in cases where it seems like hope may be lost, new and improving assisted reproductive technologies may help species persist. The CR northern white rhino, a subspecies of white rhinoceros, is possibly extinct in the wild: the living population consists of two females, Najin and Fatu, at a private sanctuary in Kenya. While no living males persist, a sliver of hope of saving this subspecies can be found in technological advancements that transform northern white rhino cells into gametes that can be used in in vitro fertilisation with a southern white rhino surrogate (Tunstall et al., Reference Tunstall, Kock and Vahala2018).
4.5.3 Global Markets and Local Impacts
As we have seen, our increasingly connected world means that choices made at a supermarket or website on one side of the planet can affect mammal species on the other. This does not always have to be a negative transaction – rather, this should empower us to recognise the role we currently play in affecting local- and global-scale mammal conservation, and alter our behaviours if necessary by supporting conservation organisations, advocating for science-based government policies and most simply, modifying our day-to-day choices as consumers. While 40% of IUCN assessed species are impacted by habitat loss (Schipper et al., Reference Schipper, Chanson and Chiozza2008), consumer decisions and technological advances in agriculture can blunt this trend through increasing crop yields, adjusting land zoning practices and implementing conservation-based agricultural trade (Tilman et al., Reference Tilman, Clark and Williams2017). Large carnivores are not only losing habitat directly to deforestation for livestock grazing, but they are also persecuted and, in some cases, actively culled, to protect livestock. Therefore, reducing meat consumption in the Global North represents a step that many individuals can take to benefit wild mammals and wild carnivores in particular. For example, Tilman et al. (Reference Tilman, Clark and Williams2017) found that halving per capita meat consumption by 2060 would lead to significant conservation benefits for medium- and large-bodied mammals in the tropics, predominantly South America and Southeast Asia – the very places where mammals need conservation action the most.
4.5.4 Rewilding, Recovery and Resilience
Mammal conservation success happens when local communities are included in decision-making about the wildlife that they live alongside. This requires expanding the disciplines considered when implementing management strategies, and also changes what narratives we tell about mammal survival in a changing world. Indeed, carnivores are recovering in many places and there are many suitable reintroduction sites globally, from Mongolia, Canada and Thailand, to Namibia, Indonesia and Australia (Wolf and Ripple Reference Wolf and Ripple2017). The return of carnivores in turn could help restore other components of ecosystems through their cascading ecological impacts across the food webs. Similarly, successful wildlife policies in Europe have led to 90% of the region having at least one native wild ungulate (Linnell et al., Reference Linnell, Cretois and Nilsen2020), and the ‘rewilding’ of large herbivores is now seen as a major component of ecological restoration policy in Europe.
Ongoing human demographic trends means that more people now live in cities than anywhere else in the world, and such concentrated urbanisation may provide new conservation opportunities for mammals. Ageing human populations are resulting in countryside depopulation in places such as Europe and Japan, making space for the recovery of large mammals in unused agricultural lands (Tsunoda and Enari, Reference Tsunoda and Enari2020). Within urban areas, even the megacity of Los Angeles, California, supports a diverse carnivore guild that persists in the midst of millions of people. The iconic mountain lion named P-22 likely crossed two freeways to take up residence below the Hollywood sign in the city’s famous Griffith Park, and despite likely predating a captive koala from the Los Angeles Zoo, P-22 remains beloved by many Angelinos, highlighting the central importance of human emotion and attitudes in conservation success.
While mammal diversity appears to be inexorably eroding, not all mammal stories are centred on loss. It is important to remember that of the nearly 6000 mammal species on Earth, more than half of the mammal species alive today are not considered to be threatened with extinction (3230 species are of Least Concern). And, since 1990, scientists have actually described more than 800 species of mammals, including large and charismatic species such as the olinguito, the Skywalker gibbon and the little black tapir (Burgin et al., Reference Burgin, Colella, Kahn and Upham2018). Species once thought to be extirpated or extinct have reappeared following decades of absence, such as the short-tailed chinchilla in Bolivia, the silver-backed chevrotain in Vietnam and the Nelson’s spiny pocket mouse in Mexico. Even the Machu Picchu arboreal tree rat virtually rose from the grave; known only from skulls in ancient Incan tombs for decades, it was unexpectedly photographed alive in 2014! From the humble beginnings of scurrying around dinosaurs’ feet, mammals have come to dominate the planet’s ecosystems, and their history of resilience prepares them (and by extension, us) for survival, even in the face of seemingly inescapable change.












