


Discussing historic fisheries calls for a common store of information, vocabularies, and ideas. This includes working acquaintance with the waters and fishes of post-Roman Europe and the conceptual framework of freshwater and marine ecology used to describe them. For readers not so equipped this chapter lays out that basic knowledge, beginning with characteristic regional waters, going on to their distinctive fishes, and then to flows of energy and materials through those ecosystems and organisms. Streams and lakes are familiar to most people. Runoff of rain and snowmelt is the point in the hydrological cycle when liquid water gains biological significance on the Earth’s surface. Historically seminal medieval fisheries took shape in Europe’s streams and lakes. For these reasons, this discussion starts in fresh water and flows to the sea, then sketches the communities of life forms, especially fishes, living in those habitats. The aim is to identify patterned relationships among aquatic organisms and their surroundings, including humans, in the earlier Middle Ages. The web of life goes around in interconnections, as does the interaction of natural and cultural spheres, so any starting juncture will include links explicable only from other perspectives. With loose ends eventually tied back together, this chapter and the next two establish a baseline – not of humans entering a pristine state of nature but of where medieval Europeans and their fishes started – to recognize and assess later changes.
1.1 Watersheds and SeasFootnote 1
Natural aquatic ecosystems in Europe result from the physical geography and Pleistocene glaciation of the subcontinent. Europe forms a peninsula from the larger land mass of Eurasia, which is a situation ecologists find commonly reducing the variety of all resident life forms (species). Still speaking in the broadest terms, from the north side of the mountains bordering the Mediterranean – Pyrenees, Alps, Balkan ranges – most of the subcontinent slopes to the north and west through uneven (but not abrupt) relief and becomes wetter. How did this topography shape fish life?
Water from rain or snow falling on western and central Europe eventually reaches the sea through one of Europe’s watersheds (river basins), which are the fundamental territorial units of freshwater habitats (see Map 1.1). Those of the north European plain are long and broad, gathering waters from part of the Alps and from many lesser interior mountains and draining northward to the Baltic and North Sea or, west of the Rhine–Maas delta, to the English Channel and Bay of Biscay. Most of the Iberian peninsula also comprises large, west-flowing river basins. Scandinavian waters drain from the central mountain ridge and other uplands westwards to the North Atlantic or south and east to the Baltic, often pausing in extensive post-glacial lakes. From almost at the Rhine and then east through the centre of the continent runs the Danube basin, largest in the west and the only major system to flow from western and central Europe into the Black Sea. The Mediterranean basin has characteristically shorter, and so more numerous, watersheds. Only the Ebro, Rhône, and Po drain extensive areas.Footnote 2 Thus Europe’s terrain and humid temperate climate make for many flowing waters, but fewer and localized permanent natural standing waters. Local and regional variation in gradient, substrate (river bed materials), and seasonal runoff result in reaches of different character, some more dynamic than others and subject to change over time.
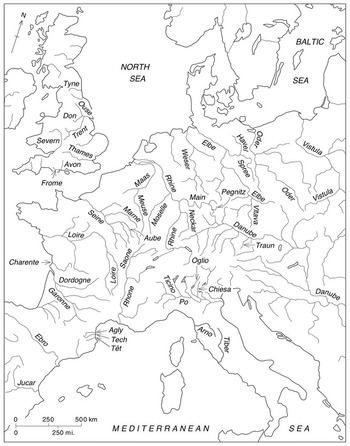
Map 1.1 Some major European rivers (mentioned in this book).
The political authority of Rome over western and southern Europe broke down in the course of the fourth through sixth centuries. So did human numbers and the intensity of land use there. For the next half-millennium or so, native plant communities blanketed much of the irregular and damp landscape north of the Alps. Vegetative cover on the drier and more abrupt Italian and Iberian peninsulas continued to show effects from intensive human use during ancient times.Footnote 3 Some good-sized areas throughout Europe were still under dense wild woodland, where human activity was little to be noticed. More early medieval terrain held open woods and brush lands subject to intermittent and extensive use by scattered human communities and their livestock. Tight growths of woody and large herbaceous plants especially covered the well-irrigated margins of watercourses.Footnote 4 So protected, the land absorbed precipitation and retained both its soils and the mineral plant nutrients in them. Fed by well-filtered and slowly released surface runoff and ground water, many European streams ran clear, cool, and stable. Seasonal high waters inundated only natural floodplains. Even in wetlands, rooted aquatic vegetation absorbed soluble nutrients and kept the water clear. In the late fourth century Gallo-Roman poet Ausonius saw the Moselle “bright as water in crystal goblets,” and his “vision, when it pierces this deep stream, finds the open secrets of the bottom.”Footnote 5 Limpid flows tumble through other poetic Gallic landscapes all the way to 1200.Footnote 6 In Thuringian Wolfram von Eschenbach’s romance Titurel (c. 1210/20), the young hero Schionatulander angled for trout and grayling wading “through the cool and clear-running brook.”Footnote 7 A real-life mid-twelfth-century sportsman, French cleric Gui of Bazoches, contrasted the clear streams still issuing from the Ardennes with those of more agricultural Champagne.Footnote 8
Such evidence of limited human impacts should not, however, be taken to imply static natural riverscapes. Multichannel rivers in early medieval England and high gradient sections of the Danube, for example, continually reworked their beds. Reaches of the Loire and Rhône shifted from incised meanders to braided form with the wetter post-Roman climate, which also brought frequent flooding and alterations to the Po basin during especially the sixth and seventh centuries.Footnote 9 Figure 1.1, though created much later, still depicts the generic complexity of most medieval rivers. Freshwater fish populations confronted changing local conditions and habitats.

Figure 1.1 A major European river, the Upper Rhine, not yet ‘regulated’.
The view from the Isteiner Klotz looking upriver toward Basel as painted by Peter Birmann about 1800, before the famous engineering of the river begun by Johann Gottfried Tulla in 1817. Though used by small boats and the village on its bank, the Rhine is dotted with islands, sand bars, and other irregular features.
Kunstmuseum Basel, Birmann-Sammlung 1859, Inv. 755. Open access and public use (Gemeinfrei).
Europe, subcontinental peninsula of peninsulas, is deeply penetrated by the sea into which its rivers flow: through salt ponds and marshes as the Po, Rhône, and Tiber; through channels among sand bars and peat bogs as the Rhine in Frisia and Great Ouse in the English fens; or through broad bays with barrier islands as the Weser and Wisła or rocky headlands as the Tejo and Seine. Great streams fed sediments from high mountains; Rhine, Rhône, Po, and Danube, formed extensive deltas. In those estuaries fresh waters transition into marine environments.
Along Europe’s southern margin the Mediterranean and Black Seas have mountainous shorelines and deep waters close to shore.Footnote 10 In their nearly three million square kilometers of water surface the only extensive areas shallower than 200 meters (600 feet, 100 fathoms) are the upper Adriatic and the northwestern quarter of the Black Sea, both lying off large sediment-rich rivers. The Mediterranean itself has a warm, dry climate, and few major sources of fresh water. On balance, more water evaporates from its surface than feeds it, which leaves Mediterranean water salty and dense. Lighter, less salty seawater flows steadily into the Mediterranean from the Black Sea and, at the surface, from the Atlantic. Down below, a strong current carries cold and salty Mediterranean water west past Gibraltar into the Atlantic. With few nutrients from surrounding landscapes, little upwelling, and almost no tidal action, the Mediterranean, especially along its northern shores, is famous for its clear – because naturally infertile – waters. The Black Sea, meanwhile, circulates much surface water from large rivers toward the strong outflow to the Mediterranean, but the shape of its basin and prevailing winds cause little vertical mixing. Below 100 meters low oxygen levels leave the Black Sea lifeless.
Unlike the south, from the inner angle of the Bay of Biscay north to Shetland and the central Norwegian coast western and northern Europe front on the wide continental shelf of the eastern Atlantic Ocean and its extensions, the North Sea and Baltic. Only in a narrow trench off southwestern Norway do soundings greater than 200 meters come nearer to shore than hundreds of kilometers. Hence, the million or so square kilometers of more or less enclosed waters in the European Atlantic, though less than half the area of the Mediterranean and Black Sea, provide two or three times more continental shelf. Even along the Cantabrian and Portuguese coasts, where great depths do reach closer to land, the shallow coastal zone is wider than in most of the western and central Mediterranean. The eastern Atlantic is swept, even into the outer reaches of the North Sea, by the great oceanic circulation of the Gulf Stream; it is agitated twice daily by tides and fed by large temperate rivers. Fresh water flowing into the Baltic, where long cool seasons limit evaporation, lowers salinity there and feeds a steady current running out the Danish straits.
By late Roman times humans had crisscrossed the Mediterranean for millennia, explored all its shores, and settled those they found useful – though winter broke their sailing season and skippers always preferred coasting routes. Comparable long-distance knowledge of movement across northern European seas was acquired only during the Early Middle Ages, as exhibited by settlement of Irish monks, then Norse farming families, on the previously uninhabited Shetland, Faeroes, and Iceland, not to mention movements by other Celtic and Germanic peoples across the protected narrow seas between Scandinavia, the British Isles, and Brittany. Of course none of this hard-gained experience meant systematic knowledge of the oceanic systems, water chemistry, or bottom profiles just described. It is always essential to distinguish between what we can now say or reconstruct of aquatic environments, with its necessary consequences for human activities, and what medievals were then aware of, even when their ignorance in no way impeded, for instance, their ability to catch and eat tuna in the Mediterranean, herring in the Baltic, pike-perch in the Danube, or sturgeon in the Loire.Footnote 11
1.2 Which Fishes Lived Where?
Fishes are and were no more uniformly distributed through Europe’s waters than are other animals on land. Most fish cannot survive in the air long enough to get overland from one water body to another. Nor can most fish even move safely between salt and fresh water, for the distinct relations of these to the body chemistry of an aquatic animal can be deadly. Seawater has a higher concentration of salts than is in the blood of fish. To maintain their own more dilute and particular osmotic balance, marine organisms must actually drink water and excrete salts. Those in fresh water face the opposite problem: their kidneys must extract surplus water and pump it out in copious urine. While most fishes can tolerate some range of salinity, only a few can reverse kidney function to live in both fresh and salt water. These are notably certain inshore marine species (called euryhaline) such as flounder and mullet, and diadromous fishes, such as salmon, shad, or eel, whose life cycle stages are adapted to alternate between the two. Most fishes, however, are either marine or freshwater.Footnote 12
1.2.1 In Freshwater EuropeFootnote 13
Freshwater fishes can naturally move from one habitat to another only through a temporary or permanent freshwater link where both temperature and dissolved oxygen levels fall within the tolerance of the species. That meant, until humans built the first major inter-basin canals around 1700, European fishes of fresh water could of their own accord move between watersheds only when geological processes slowly changed the land itself. For instance, the barbel, a resident of moderately flowing rivers on the continent, naturally occurs in Britain only in those eastern watersheds which once joined the Pleistocene Rhine in its route across the then dry or ice-covered floor of what later rising sea levels have made into the North Sea and English Channel.
Given that a fish species could reach a watershed in the past, its presence and distribution there depends on its adaptability to certain environmental factors. Water temperature and dissolved oxygen are critical for cold-blooded, oxygen-respiring animals. Cold water holds more oxygen than warm. As with salinity, each species has a range of tolerance for both variables. Current speed is important in the lives of fishes, too, for it makes them expend energy and through erosion it modifies beds, banks, and water clarity. Suitable food supplies and spawning sites further determine the presence and abundance of a species in a watershed. When any of these factors violates threshold values, the species will be extirpated.
Factors of water temperature are thought to be the greatest single cause for the gross distribution of fishes among European watersheds (and, as we’ll see shortly, in marine environments, too). Cool-water varieties like salmon, trout, whitefishes, and pike were naturally absent from most Mediterranean watersheds or there confined to high-altitude headwaters. Especially in Spain and the Balkans their niches are filled by more tolerant local fishes.
The number of resident freshwater fish species is low throughout Europe and lowest in the northwest.Footnote 14 Many thousands of years ago, a continental ice sheet spread over most of northern and western Europe. As Pleistocene waters chilled below the tolerance level of some, then all, aquatic organisms, those unable to move into salt water died or withdrew to the southeast, the only direction where warm fresh water was accessible. Only cold- and salt-tolerant varieties like salmon, trout, and their kin (collectively called Salmonidae) could survive in oceanic coastal waters and frigid ice-front lakes. Fishes needing warm and fresh water, notably Eurasia’s many representatives of the carp family (Cyprinidae), survived the Ice Age only in the refuge of the lower Danube and other rivers running from the Balkans into the Black Sea and Caspian. From there they could only very slowly recolonize the greater part of peninsular western Europe.Footnote 15 Hence the ancestors of all life forms in early medieval waters had survived or repopulated the frozen antithesis of a freshwater habitat, and the numbers of fish species in European watersheds fall sharply along an east–west gradient.
The clear, cool inland waters of post-glacial western Europe thus held relatively simple fish communities with a low diversity of native species. Most were well adapted to cold, well-oxygenated, alkaline, running-water habitats with low levels of dissolved nutrients. Many fishes in drainages running to the Atlantic, North Sea, and Baltic show adaptations to marine access and to conditions in the ice-front lakes that had joined these basins while the glacier withdrew slowly downslope to the north.Footnote 16 Notably important as fish biomass and for human use in watersheds from the Garonne to the Wisła were sturgeon, shads, and closely related families of salmon, trout, and whitefish. Most of these fishes are anadromous, meaning they spend their adult lives more or less at sea and return to fresh water to reproduce. Two of western Europe’s three species of lampreys are also anadromous. Eel, however, a species of large and growing early medieval economic importance, have the reverse habit of ‘catadromy’, for they enter freshwater as larvae to grow and mature, and return to the sea to spawn and die. Eel, lampreys, sturgeon, and shads were also important natives of western Mediterranean watersheds, but marine migratory salmonids do not occur there. Other ubiquitous (apart from peninsular Italy and Spain), numerous, and oft-eaten natives were the European representatives of two circumpolar genera, the perch, and the pike. About 1080 the monk Ulrich compiled a dictionary of the sign language used at Cluny, then western Christendom’s most respected monastery, during times of compulsory silence. His section on foods named only six fishes, but five of them were from fresh water: salmon, trout, pike, eel, and lamprey.Footnote 17
Other parts of the Old World have many representatives of the huge Cyprinidae family – they comprise 80 percent of China’s 600 freshwater species – which commonly favour warm still waters. By comparison, however, rather few members of this family live in western Europe, and those naturally present, like the barbel, bream, roach, minnow, or gudgeon, are unusually adapted to cool and/or running water. Only barbel and gudgeon, two cyprinids especially suited to swiftly flowing middle reaches of streams, joined perch, pike, salmon, and eel in the catches twelfth-century sporting cleric Gui of Bazoches made in northern French waters.Footnote 18
The post-glacial ice-front lakes for a time united what would become distinct watersheds. As the ice retreated, fishes from the lakes spread downstream to the northwest while migratory species moved along the coasts. (Both groups left relict populations in habitats like cold Alpine lakes.) At the same time, warm-water varieties could work up the Danube system and, using occasional ice lake or swampland inter-basin connections in poorly drained post-glacial Europe, gradually reenter the Atlantic drainage from east to west. The roach made it all the way to central Scotland and to rivers running into the Gulf of Bothnia, and the barbel to the Thames and north Germany. The European catfish or wels spread only so far as the upper Rhine, Elbe, and a small beachhead in southern Sweden. Carp had under wild conditions by the early Middle Ages reached only the middle Danube.
Finally, and quite apart from issues of water temperature, Europe’s peninsular character has held up the westward spread of freshwater fishes with more easterly distributions. The migratory Atlantic salmon is absent from Mediterranean and Black Sea watersheds. In the latter its niche is filled by the huchen or ‘Danubian salmon’, a non-migratory form with congeners in central Asia. Likewise neither cold nor heat barred from the Rhine, Rhône, and other western watersheds the pike-perch or zander, close relative of the North American walleye and occupant of a broad Pontic, Russian, Scandinavian, and east-central European range.
One side of that peninsular Europe, the Baltic Sea, can better be understood as one of the world’s largest estuaries, a body of water where fresh and salt mix. This makes for a hard environment, but for fishes which can adapt, also a fecund one. The Baltic at least once held major populations of some cyprinids and pike in its freshest eastern reaches, salmon, sea trout, and sturgeon throughout, and in the saltiest west, at times legendary numbers of herring and cod.
1.2.2 Marine Zoogeography
The global distribution of marine fishes is less well understood than are freshwater varieties, if only because more species cover larger areas with less visible boundaries among them. In place of explanatory principles marine zoogeographers offer descriptive regions set by oceanographic conditions and continental shelves, where currents and especially water temperature seem influential.
Fishes in historically relevant nearshore European waters chiefly form a ‘Mediterranean–Eastern Atlantic’ temperate region, bounded fuzzily on the north along a line from Iceland to northern Norway by a distinctive circumpolar Arctic or boreal region. In the south, the Mediterranean fauna, with many (540) species, has much in common with the Atlantic south of the English Channel, where summer water temperatures also stay above 12°C. Fishes from typically tropical families, the mullets and sea breams or porgies, for instance, are common inshore, and so are the sardine/pilchard and anchovy. Low salinity and restricted marine access let only some of the Mediterranean fishes populate the Black Sea. More northerly Atlantic waters are consistently colder, averaging between 12° and 6°C, and containing important representatives of coldwater fish families, the codfishes, flatfishes, herrings, and salmonids. Only some of these range into the Bay of Biscay. Colder waters typically contain fewer species and very large numbers of individuals in those species.
Two other marine regions need mention. Open offshore surface waters provide a different kind of migratory habitat to distinctive, sometimes world-girdling fish families. South of the British Isles and in the Mediterranean the tuna or tunny, mackerels, and swordfish, are familiar, at least in warmer months. Further north this role is more that of adult salmon. Finally, deep-sea fishes, those which live on and below the continental slopes, seem relatively undiversified – and remained all but unknown to human fishers before the industrial age.
1.3 Ecosystems, Habitats, and Communities
Even in native waters few fishes live everywhere. Variables of current speed, light penetration, and nutrient levels separate different habitat zones to which various species are more or less adapted. Inland, the basic division is between moving and still waters; the former, where constant movement and mixing dominate, contain what ecologists call ‘lotic’ habitats, and the latter, more influenced by depth and water chemistry, ‘lentic’ habitats. In such terms marine environments might be conceived as oversized and chemically distinctive lakes, with their own analogous habitat zones and nomenclatures (pp. 44–45 below).Footnote 19
In the lotic habitats of running waters European biologists commonly distinguish four zones of decreasing current speed, each named after a typical resident fish.Footnote 20 The trout zone is in mountainous headwaters where steep gradients cause very fast currents. These make for much dissolved oxygen but so erode the banks and bottom as to leave them mostly rock and gravel. Trout and young salmon are the most common fish species. As the bed flattens and currents slow slightly, the stream enters the grayling zone. Oxygen levels remain high, but more gravel is found, and sheltered places hold enough sand or silt for rooted aquatic plants. Trout or grayling are the most abundant fishes, but commonly joined by cyprinids like barbel, dace, and chub. Third, the barbel zone is found where the gradient becomes gentle and the current moderate. The bottom is predominantly soft, and rooted plants are common. In a mixed fish fauna the cyprinids dominate; if water temperatures remain low, trout and grayling may still occur, but piscivorous pike and perch are also important. Finally, the bream zone is named for the slow-water cyprinid native to western Europe. In quiet, warm, and weedy waters they are accompanied by roach, rudd, tench, predatory fishes, and now the introduced carp. Not until this last zone in the lowest and gentlest reaches of rivers are depth or circulation likely to impede constant temperature, light, and nutrient levels throughout the water column. Streams rising in bogs or having high gradients in middle or lower reaches will depart from the normal zonal pattern and can accordingly show unusual distributions of fishes.
Biologists commonly stratify the lentic habitats of still waters (lakes, ponds) by depth and light penetration into three major zones, which they label littoral, limnetic, and benthic. The littoral zone extends from the shore to the depth limit of locally prevalent rooted aquatic plants. It contains many nutrients, the warmest water of the lake, and the greatest variety of fish species. The limnetic or pelagic zone is the lakeward extension of the littoral. Its open surface waters are well-lighted and thus can hold large populations of drifting microscopic plants, plankton, on which certain fishes, such as some whitefishes and cyprinids, can feed. Like littoral waters, the upper layer of the limnetic is subject to rapid changes through wave action or air temperature. Beneath the limnetic zone the dark benthic (profundal) zone goes to the bottom of the lake. It receives nutrients from above, but is commonly cold and often short of oxygen. The profundal zone is chiefly inhabited by bottom-feeding fishes and their predators, such as European catfish or burbot, the freshwater cod with a circumpolar range. The relative importance of each zone depends on the predominant physical features of each lake or lake region. Deep and steep-sided lakes may have no littoral zone and shallow ones nothing else.
Unlike rivers, ponds and lakes are ephemeral features of geologically ‘young’ landscapes. As the land erodes, all still waters accumulate nutrients, become more fertile, and slowly fill in. This natural process, called eutrophication, moves lakes through three conventional stages. In Europe lakes at different stages are naturally characteristic of certain physiographic regions. Mountain lakes are commonly deep and narrow, hence typically cold habitats with low levels of dissolved nutrients and rooted plants. Biologists call infertile habitats oligotrophic. They are suited for salmonids, plankton-eating whitefishes, and associated predators. Unexploited oligotrophic lakes can contain quite large biomass concentrated in large, old, and slow-growing adult fishes. Where the land is more rolling and fertile, as around the post-glacial lakes of northern Germany and Poland, the still waters hold more nutrients and more shoreline vegetation. This middle group, called mesotrophic lakes, supports a mix of salmonids and cyprinids. Finally there are the highly fertile eutrophic lakes, relatively shallow, soft-bottomed, and full of plant life. In Europe they are naturally associated with slow-moving reaches of rivers (the bream zone) and inhabited by the same fishes.
At a scale smaller than whole watersheds, then, European regions and localities contain characteristic freshwater habitat zones and characteristically abundant or rare fish species. Comparable differences occur in marine ecosystems, where a general classification of habitats as coastal (neritic), offshore surface (called variously pelagic, epipelagic, or oceanic), and bottom-oriented continental shelf (sublittoral, benthic) seems to organize the environments historically engaged by human activities.Footnote 21
The sea is a moving and changing environment with patterned uniformities, which exhibit more variation where seawater contacts air or earth. Hence marine ecosystems classify into habitat zones oriented to the surface (neritic and epipelagic) and to the bottom (benthic). The neritic zone is the water over the continental shelf, so extending downward to about 200 meters. This water is well supplied with light, oxygen, nutrients, and wave and current action, all subject to seasonal variation. Such variety supports many habitat niches and hence the greatest diversity of marine fishes. Herrings, eels, tunas, and mackerels are familiar inhabitants of the neritic zone. Continuation of this same 200-meter-thick, well-lighted and well-oxygenated layer out beyond the continental shelf produces the epipelagic zone, a huge and featureless expanse of open ocean waters. Most fishes – think of tunas, mackerels, and swordfishes, but also of sardines and anchovies – concentrate in nutrient-rich areas of upwelling or near coasts. Beneath the neritic zone are the bottom-oriented habitats of the benthic, from the intertidal area, exposed to the air up to half of every day, down to the edges of the continental slope, where even seasonal changes are barely perceptible. Across most of this submarine expanse, wave action and the makeup of the bottom (substrate) rank behind only temperature in distinguishing habitats for bottom-oriented (demersal) fishes. Important in European waters are the flatfishes – flounder in estuaries, plaice inshore, halibut well out on the shelf, etc. – and codfishes – whiting along shore, often with small cod and pollack not much further out, adult cod and hake in deeper waters, the former northwards in the Atlantic, the latter more common south of the British Isles, and ling favouring the deepest zones on the slope. Waters below 200 meters lack light and seasonal changes, while temperature declines and pressure increases with depth. Numbers of fish species and individuals diminish, too. All organisms there must eat (once-)living objects fallen from above, whatever they can catch on brief feeding forays upwards, or one another.
Finally, a transitional and fluctuating quality dominates estuarine ecosystems, where fresh and salt waters meet in a turbid, sandy river mouth, a deep, clear, rocky fjord, or perhaps a maze of channels, bars, and weed or shellfish beds. Diadromous fishes use estuaries to move between spawning and feeding areas. The layering of dense salt water under lighter fresh water and their eventual mixing by currents and tides cause high biological productivity, so many fishes use estuaries as nurseries for their young. Consequently, estuaries often hold large seasonal concentrations of certain species.
1.4 Food Webs and Other Relationships
Like all of Earth’s widespread ecosystems, those in water depend on energy from sunlight, which powers the winds and currents circulating air and water and which green plants, large and small, use to produce organic material (a store of chemical energy) and oxygen from water and carbon dioxide. Only plants, from single-celled drifting plankton to well-anchored kelp or massive oaks dropping leaves into a brook, are ‘producer organisms’, and that only when exposed to sunlight. The ‘euphotic’ zone where light is available penetrates at most some 200 meters into clear seawater, and much less into water clouded or stained with suspended sediment, soluble chemicals, ice, or drifting plankton. In lentic fresh waters and in the sea, then, plants can live only in the littoral or neritic zones along the shore or in the (epi)pelagic surface layer; elsewhere they stop making food and oxygen and first temporarily (in nighttime respiration), then permanently (in decomposition) absorb oxygen to break down organic hydrocarbons. Turbid waters may limit the productive euphotic zone to only a few meters.
Survival of green plants, the basis of all life on Earth, demands air, water, and also access to the relatively rare soluble forms of nitrogen and phosphorus, mineral salts or ions of nitrates and phosphates. These nutrient chemicals are obtained in solution from terrestrial runoff and concentrated in the bodies of plants and animals. Solid organic wastes, including dead bodies, gradually sink to the floor of a still water body and thus out of the euphotic zone, where plants can reuse their constituent minerals. Inshore and nearshore marine regions where soluble nutrients are continually recharged with the runoff of large rivers – like, for instance, the North Sea, Bay of Biscay, or Gulf of St Lawrence – can support richer rooted and planktonic flora than can more sterile waters – the Mediterranean or the epipelagic areas of mid-ocean. Aquatic life also flourishes where natural mechanisms – upwellings of deep water like the Labrador Current or the annual ‘turnover’ of water in temperate-zone lakes before freezeup in fall and after breakup of ice in the springFootnote 22 – stir mineral nutrients back up from the sterile and Stygian depths.
All animals consume and transform plant materials. The plant-eating herbivores (primary consumers) are followed by succeeding trophic levels of animal-eating predators (secondary, tertiary, etc. consumers). Most fishes occupy an identifiable niche or level in this interlocking web of eating and being eaten. Besides the invertebrate animals which do much primary consumption in many aquatic ecosystems, European fishes filling that role include the minnow in the trout zone, whitefishes in the limnetic zone of oligotrophic lakes, and the roach in warm and fertile waters. Top-level predators, the uneaten eaters, in the same habitats include large trout in cold streams and lakes, pike, and catfish. Cyprinids of large adult size such as barbel, carp, or bream commonly function as herbivores (and important prey for predators) when young, and when larger, as low-level predators consuming many invertebrates along with their diet of plant materials. Along coasts from Brittany to Scandinavia tiny young cod near bottom inshore and tiny young sand eels offshore eat zooplankton, so are secondary consumers; adult cod eating adult sand eels are at a higher trophic level. Western Mediterranean pelagic tuna often eat Spanish mackerel, which commonly eat anchovies and sardines, which as adults eat mainly zooplankton, which eat phytoplankton.
Because living things convert only about 10 percent of the energy from one trophic level to the next, organisms nearest the base of the food web (producers or primary consumers) have necessarily the greatest total biomass and numbers of individuals; those with the most intervening stages are reciprocally low in biomass and numbers. The relationship can be visualized as a trophic pyramid (Figure 1.2)Footnote 23 Predators are characteristically larger in individual size but fewer in number and less in total mass than their prey. Predators and prey coexist in nature – sometimes the interaction controls the numbers of both – but the relationship is not necessarily stable and environmental changes of any origin can cause great dislocation.
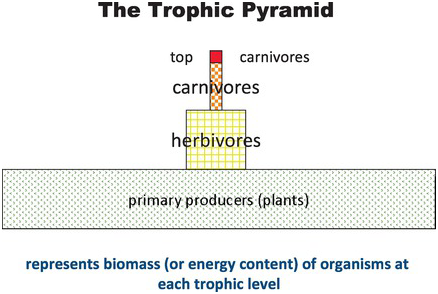
Figure 1.2 The trophic pyramid.
In natural ecosystems the food web is completed through the activities of detritivores (waste-consuming organisms) and the process of decomposition, which break down complex tissues and recycle the material components of life (minerals, carbon, water) back into food for plants. Organisms removed from a watershed take the embodied nutrients with them. Lotic (running-water) ecosystems continually export nutrients downstream and so must receive regular inputs from the surrounding terrestrial environment.
Food webs are not the only historically significant patterned relationship among different species or individuals of the same species. Different animals compete for access to resources which their ecosystem holds in limited supply: think of food sources, spawning sites, even, in some situations, oxygen. Generally speaking, two species with identical habitat requirements cannot coexist; one or both must adapt or die out.Footnote 24 Among fishes, patterns of competition commonly shift with phases of a species’ life cycle. The small fishes vying for zooplankton or insect food in the shallows of a sluggish English river may include young pike which as adults will eat their one-time competitor roach or loach. Adult salmon and trout choose gravels of different size to make spawning sites in the same reach of a stream, but for a year or two after hatching their offspring will compete to live from the same drifting or bottom-dwelling invertebrates.
Competition also occurs within a single species and population – over food, cover, nesting sites, and so on. Young carp eat much the same bottom invertebrates as do adults, and spread the total carp biomass across many small individuals. Young pike eat insects and smaller fish and so convert abundant but small-sized food into a more efficient meal for large pike – whose cannibalistic proclivities were familiar to medieval observers.Footnote 25 The hazard of competitive and predatory interaction between age classes is one reason many fishes have breeding grounds and nursery habitat distinct from adult feeding areas – which, in turn, makes those species dependent on more than one habitat. Note further that such patterns of interaction across the species’s life cycle can relate to the structure of fish populations. Schools of pelagic herrings in the North Sea, for example, are successively made up of particular dominant year classes, great numbers of fish all born in the same especially good year for survival and growth of the larvae – rather like the baby-boomer cohort among late twentieth-century North Americans and Europeans. Among inland lake fishes – charr are a prime example – good conditions for small fish but limited food supplies will spread the biomass across large numbers at the lower end of the size range; limited reproductive success, on the other hand, will let large, even possibly slow-growing older fish dominate. The latter is also true of sturgeons, which take decades to reach sexual maturity.
From a broader perspective, considering flows of energy and competition over them is just one way to think about the structure, preferences, and vulnerabilities of aquatic communities. Especially recent work in community and restoration ecology groups species, irrespective of their evolutionary relationships, in terms of overlapping habitat requirements into ‘ecological guilds’. Trophic levels may thus be cross-cut by such shared preferences as reproduction conditions (spawning on gravel, vegetation, sand, or open water with or without current or nest building), current velocity, migratory or schooling behaviours, and relative tolerance or sensitivity to water quality, temperature, etc. ‘Guild’ membership means certain environmental changes can have similar effects on unrelated fish species, resulting in alterations to an entire community.Footnote 26
Another ecological principle emphasizes the importance of ‘edge’ between two adjoining habitats as affording access to resources in both. This helps explain the rich concentration of fish life near the surface, where light and oxygen come from the air, in the open sea and open lakes; in the well-mixed and ever-changing tidal zones, with their many closely defined habitat niches; and in churned-up, nutrient-rich waters of estuaries. Plant life and herbivorous organisms attract an ascending pyramid of predators. The attraction works in other directions, too. Rich populations of fishes in Europe’s streams and coastal inlets attracted many air-breathing predators, birds, mammals, and humans, to what were in the early Middle Ages still relatively natural boundaries between water and land.Footnote 27
Humans who insert themselves into aquatic ecosystems by taking fish for food become a new apex predator atop the trophic pyramid, but one with potentially distinctive impacts. Aquatic food webs, especially those exploited by humans, are characteristically longer than terrestrial ones, as people in most cultures eat few aquatic plants or herbivores but rather the larger carnivorous fishes at the top of the food chain. Human choices of prey species may be set by biological variables (prey numbers, habits, accessibility), or be determined by such cultural aspects as taste, prestige, or technical knowledge. Human fishers can anticipate future concentrations of favoured fishes or wait and select even rare varieties yielding fewer calories than the work necessary to catch them.Footnote 28 Different methods of catching fishes may be more or less selective of species, habitat, fish size, or habits in a given aquatic community.Footnote 29 The chosen bait can strongly affect which species are taken with hook and line: medieval fishers knew well that cyprinids liked bread paste with strong herbal flavours and pike a shiny little fish. The season and habitat fished have more influence on the catches made with basket traps or a trawl: among the oldest written records of fishing in lakes near Salzburg is a mid-ninth-century agreement which distinguished the autumn run of lake-dwelling trout from the spring fishery for albuli pisces (probably cyprinids, possibly Coregonid whitefishes).Footnote 30 Size of mesh in a net determines the share of small individuals taken, but fish of all sizes are available for gathering after being stunned with a submarine explosion or a poison. Hence different fisheries can have highly variable effects on an aquatic ecosystem.
Two effects of predation may be especially connected to humans. Human preferences for larger fish, whether cultural or technical in origin, often have distinctive and detectable results. In part because fish, unlike other vertebrate animals, grow continually throughout their lives, and in part from human cultural and device biases, even moderate fishing pressure removes the largest individuals and reduces the probability of others surviving to achieve great age and size. This commonly affects the naturally larger and less numerous predator species, with the further consequence of triggering a ‘trophic cascade’: as natural apex predators are removed, numbers of lower-ranking carnivores increase and can, in turn, consume more of their own (still smaller) prey … ultimately even altering any limits the feeding of herbivores may have imposed on plant biomass. Secondly, when human predators (fishers and/or consumers) find themselves deprived of once favoured large carnivorous fishes, they may respond by ‘fishing down the food chain’, that is, targeting and developing a taste for more numerous species from lower trophic levels (perch or roach in place of pike or pike-perch; sardine or herring instead of tuna or cod).Footnote 31 Precise local balances might then determine the effects of a fishery on its supporting ecosystem. Consequences also follow from ways humans finally consume the fishes they catch and allow both waste and consumed nutrients eventually to re-enter an aquatic ecosystem. From a metabolic perspective it matters if the nutrients are being restored to the system from which they had been removed or added to some other waters. It is rare for effluents from small and scattered human populations to have impacts that are more than local or short-term. From this historical distance, at least, none are readily visible in early medieval Europe.
But in contemporary view and in historical retrospect the study of fisheries abounds with observational hazards deeper still than that between one elemental medium and another. Whether held by present-day scientific observers, past participants, or environmental historians looking to reconstruct past conditions, human cultural representations are susceptible to at least two diametrically opposed delusions. One, the shifting baseline syndrome, arises from the short professional lifespan of experiential observers, fishers or scientists, for whom conditions of their youth become personal benchmarks for subsequent assessments of change. The syndrome tends to suppress perception of trends occurring over longer periods of time than the subject could witness. A possible shifting baseline mindset calls for critical care in using both recent scientists’ assertions of no meaningful changes to a fish stock or ecosystem prior to a certain recent event and past narrators’ anecdotes of stability ‘since time immemorial’.Footnote 32
The other misapprehension assumes the great and stable abundance of a mythic past before personal experience. According to a tale well-reported in European and North American sources since at latest the seventeenth century, for instance, not so long ago Atlantic salmon used to be so abundant that household servants demanded contractual or even legislative protection against being served salmon more than once (or twice or thrice) a week. Yet no contemporary testimony or evidence of such documents has ever verified this retrospective dream.Footnote 33 So, too, back in the early twelfth century Cosmas of Prague, first Czech historiographer, wrote down how the Czech people first entered Bohemia (fifth/sixth century?) and there found “tasty fishes, healthy to eat … from waters full of fishes beyond measure.”Footnote 34 While the myth of hyperabundance imagines a pristine past and degenerate present, shifting baselines limit change to what the subject could her- or himself observe. Both attitudes and assumptions formulate an unchanging ideal past and a human-driven present of one-way decline. Unchecked, both discourses are inimical to good science and to good history. Evidence of the past itself, that of palaeoscience and of the historical written and material record, better reflects and echoes both stabilities and changes.
Humans are not the sole drivers of massive change to aquatic or other ecosystems. Events as local and abrupt as entry of a new species or as large and perhaps slow as a new pattern of runoff or general climate change can, did, and do alter basic ecological relations to the point of what ecologists call a ‘regime’ change or shift.Footnote 35 Species once keystones can be eliminated or so reduced in biomass that changes cascade up and down the trophic pyramid. In the post-glacial long run, for instance, both the Baltic and the Mediterranean experienced periods disconnected from direct access to the Atlantic, becoming respectively freshwater or highly saline lakes with fish assemblages unlike those of late antiquity or today.
1.5 Rome and After
Early medieval fish populations are not to be thought as a pristine nature in stable habitats unsullied by human activity. By about 500 CE, Europeans had been taking and eating fish for millennia, with or without written records. The classical Greco-Roman Mediterranean is being explored by marine environmental historians despite how literary proclivities of elite Roman writers, lack of any operational records for ancient fisheries enterprises, and relative paucity of domestic or market refuse yielding large samples of consumption together curtail opportunities to establish genuine ecological perspectives. Recent research has found fish a more important dietary element than was previously thought. Fresh fish were an elite dish, supplied by local artisanal fishers from nearby waters. Processed fish products had wider distribution in salted form, especially in the various fermented sauces generically called ‘garum’. Sustained by urban demand scores of industrial scale processing sites around and beyond the western Mediterranean made sauce out of the tiniest specimens, bycatches, and at least scraps of tuna and other large varieties. Both segments of the industry relied on ecological knowledge of local fishers, some of it possibly even passed on into modern times. The best-documented Roman fisheries were exclusively a marine activity which in the West faded together with Roman naval superiority.Footnote 36
At least two ecological impacts mark the historical record before the end of Roman times.Footnote 37 While archaeological evidence from previously uninhabited early Neolithic Cyprus and of cods taken in the early sixteenth century from the formerly unexploited Grand Banks both show initially remarkable abundance of very large old specimens, even pre-Roman fish remains from Europe lack any such features. But regionally confined populations of especially desirable fishes did incur further more particular and long-lasting damages during Greco-Roman antiquity. Roman writers blamed high prices on depleted coastal fisheries.Footnote 38 As remarked in the Introduction, long-term archaeozoological study of sturgeon remains from the Rhône estuary shows heavy commercial fishing reducing first the average size, age, and then the numbers caught of this large and long-lived but slow-growing favourite of Roman gourmets. By late antiquity local overfishing had essentially removed sturgeon from the Provençal catch and dining table. Subsequent release of pressure still meant that stock needed centuries to recover its pre-Roman abundance.
In general, then, human influence on aquatic life in Europe during the post-Roman, early medieval centuries up to about 1000 CE was diffuse and marginal, limited with respect to habitat even in the most-impacted Mediterranean to the irreversible hydrological effects of classical land-use patterns, and otherwise to local or superficial results of long-standing but rarely intensive fisheries. Ecologies in and around Europe’s fresh and coastal waters were largely consequences of natural drivers, processes, and their long-term changes. These had shaped a subcontinent of cool running waters with stable seasonal cycles in diverse riverine habitats, a sparsely differentiated fish fauna much adapted to marine access, and extensive tracts of marine coastal and inshore habitats both in the warm but nutrient-poor Mediterranean and the nutrient-rich but cold western and northern seas. But grounds for human pursuit of fish were shifting.



