


Introduction
Many birds migrate at night, a fact which for over a century has inspired a substantial amount of scientific research (e.g. reviewed in Graber and Cochran Reference Graber and Cochran1959, Farnsworth Reference Farnsworth2005). Technological advances in recent decades have made possible a considerable improvement in our understanding of nocturnal migration. Predominant among these innovations are a variety of observational techniques, each with a different set of advantages and limitations, e.g. radar surveillance (Lack and Varley Reference Lack and Varley1945, Kunz et al. Reference Kunz, Arnett, Cooper, Erickson, Larkin, Mabee, Morrison, Strickland and Szewczak2007, Gauthreaux et al. Reference Gauthreaux, Livingston and Belser2008, Bruderer et al. Reference Bruderer, Steuri, Aschwanden and Liechti2012, Shamoun-Baranes et al. Reference Shamoun-Baranes, Alves, Bauer, Dokter, Hüppop, Koistinen, Leijnse, Liechti, van Gasteren and Chapman2014), thermal infrared cameras (Liechti and Bruderer Reference Liechti and Bruderer1995, Gauthreaux and Livingston Reference Gauthreaux and Livingston2006, Hüppop et al. Reference Hüppop, Dierschke, Exo, Fredrich and Hill2006), ceilometers (Gauthreaux Reference Gauthreaux1969, Avery et al. Reference Avery, Springer and Cassel1976 ), moonwatching (Lowery Reference Lowery1951, Nisbet Reference Nisbet1959, Kiepenheuer and Linsenmaier Reference Kiepenheuer and Linsenmaier1965, Liechti and Bruderer Reference Liechti and Bruderer1995) and, most recently, stereo videos (Huang et al. Reference Huang, Caruana, Farnsworth, Kelling and Ahuja2016). A different approach using sound focuses on the calls of nocturnally migrating birds, which have long been the object of aural studies (Libby Reference Libby1899) and, in the wake of technical progress, of acoustic monitoring studies based on sound recording (e.g. Graber and Cochran Reference Graber and Cochran1959, Evans and Mellinger Reference Evans and Mellinger1999, Larkin et al. Reference Larkin, Evans and Diehl2002, Hüppop et al. Reference Hüppop, Dierschke, Exo, Fredrich and Hill2006, Brandes Reference Brandes2008, Frommolt et al. Reference Frommolt, Hüppop, Bardeli, Hill, Koch, Tauchert and Specht2012).
Each of these approaches has a serious weakness in connection either with the quantification of nocturnal migration or with species identification. The ‘visual’ methods, including radar surveillance, are able to deliver reliable information about nocturnal migration densities, but species identification is almost impossible (Larkin et al. Reference Larkin, Evans and Diehl2002, Gauthreaux and Livingston Reference Gauthreaux and Livingston2006, Schmaljohann et al. Reference Schmaljohann, Liechti, Bächler, Steuri and Bruderer2008). Indeed, in radar studies it is not always possible even to distinguish between birds and other airborne animals like insects or bats (Larkin Reference Larkin1991, Kunz et al. Reference Kunz, Arnett, Cooper, Erickson, Larkin, Mabee, Morrison, Strickland and Szewczak2007), which, however, might change with the development of better analytical techniques (e.g. Zaugg et al. Reference Zaugg, Sapporta, van Loon, Schmaljohann and Liechti2008). Conversely, acoustic-based methods make possible species identification through species-specific vocalisations, but, for example, owing to different calling behaviours among species (Dierschke Reference Dierschke1989, Evans and Rosenberg Reference Evans, Rosenberg, Bonney, Pashley and Cooper2000, Evans and O’Brien Reference Evans and O’Brien2002, Farnsworth Reference Farnsworth2005) the number of individuals or the density of migration cannot be inferred from the number of counted calls.
Nevertheless, acoustic-based field methods have aroused much interest in recent years as protracted digital recording has become increasingly more cost-efficient and easier to handle (Farnsworth Reference Farnsworth2005, Farnsworth and Russell Reference Farnsworth and Russell2007, Brandes Reference Brandes2008, Frommolt et al. Reference Frommolt, Hüppop, Bardeli, Hill, Koch, Tauchert and Specht2012). A modern sound recorder can autonomously record nocturnal calling activity throughout entire nights for complete seasons. The ensuing recordings can later be analysed in laboratory conditions for specific calls to glean information on species composition, calling rates and phenology, making this an important tool for scientific research and environmental impact studies (Hüppop et al. Reference Hüppop, Dierschke, Exo, Fredrich and Hill2006, Kunz et al. Reference Kunz, Arnett, Cooper, Erickson, Larkin, Mabee, Morrison, Strickland and Szewczak2007, Sanders and Mennill Reference Sanders and Mennill2014).
Due to these and other advantages of the method (near autonomous maintenance, low equipment costs and ease of deployment even in areas which are remote or difficult of access), the use of such a recorder was thought to be the best suited to shed light on nocturnal bird migration at Besh Barmag, a recently discovered bottleneck site in Azerbaijan (Heiss and Gauger Reference Heiss and Gauger2011). Here, the passage through a 2.5 km wide coastal plain and along the Caspian coast was recorded of an estimated 1.24–1.51 million diurnal migrants in autumn 2011 and 0.65–0.82 million in spring 2012, involving in total 278 species (Heiss Reference Heiss2013). Most of the observed diurnal migrants in autumn 2011 and spring 2012 were passerines, but also substantial numbers of waterbirds were counted in autumn 2011 with lower numbers in spring 2012. Soaring migrants were almost absent in autumn 2011, but minor numbers, compared to other bottleneck sites in the Western Palearctic, were observed in spring 2012 (Heiss Reference Heiss2013).
So far, whereas diurnal migration has been the focus of a number of studies already in the Caucasus region, e.g. near Lenkoran in Azerbaijan (Tugarinov Reference Tugarinov and Kozlova1950), Dagestan in Russia (Butiev et al. Reference Butiev, Mikheev, Shubin and Galushin1983, Mikheev Reference Mikheev and Amirkhanov1985, Lebedeva and Butiev Reference Lebedeva and Butiev1998) and Batumi in Georgia (Verhelst et al. Reference Verhelst, Jansen and Vansteelant2011), nocturnal migration has received no attention whatsoever either in the bottleneck area itself, or on a wider scope, within the Transcaucasian region in general.
To redress this situation I therefore carried out an acoustic monitoring study of the nocturnal bird migration at Besh Barmag to discover on the basis of the number of recorded species and number of calls in how far the diurnal bottleneck-effect of the site extends to nocturnal migrants.
Methods
Sound recordings were obtained at the bird migration bottleneck of ‘Besh Barmag’ (40°59’N, 49°13’E) in Azerbaijan. At this location the foothills of the Greater Caucasus almost reach the shore of the Caspian Sea forming a narrow coastal plain a mere 2.5 km wide through which during daylight hours high densities of birds migrate (Heiss and Gauger Reference Heiss and Gauger2011, Heiss Reference Heiss2013, Reference Heiss2016). To obtain data on nocturnal migration here, sound recordings were made at a fixed point in the coastal plain covering the nights from 2/3 August to 16/17 November 2011 and from 2/3 March to 29/30 May 2012 (Figure 1). For the purposes of this study, ‘night’ is defined as the time between sunset and sunrise, thus including all twilight phases. Due to shifting sunset and sunrise times, night lengths differed between 9 h 08 min on 29/30 May 2012 and 14 h 08 min on 16/17 November 2011 (Figure 1). The recording device used was a Song Meter SM2 Recorder with the omnidirectional SMX-NFC Acoustic Night Flight Call Microphone (Wildlife Acoustics Inc.) with a high-pass filter set at 180 Hz, a pre-amplifier with a gain setting of 48dB, a 32,000/s sample rate and the compression format WAC5 with no detectable loss of information according to Wildlife Acoustics Inc.
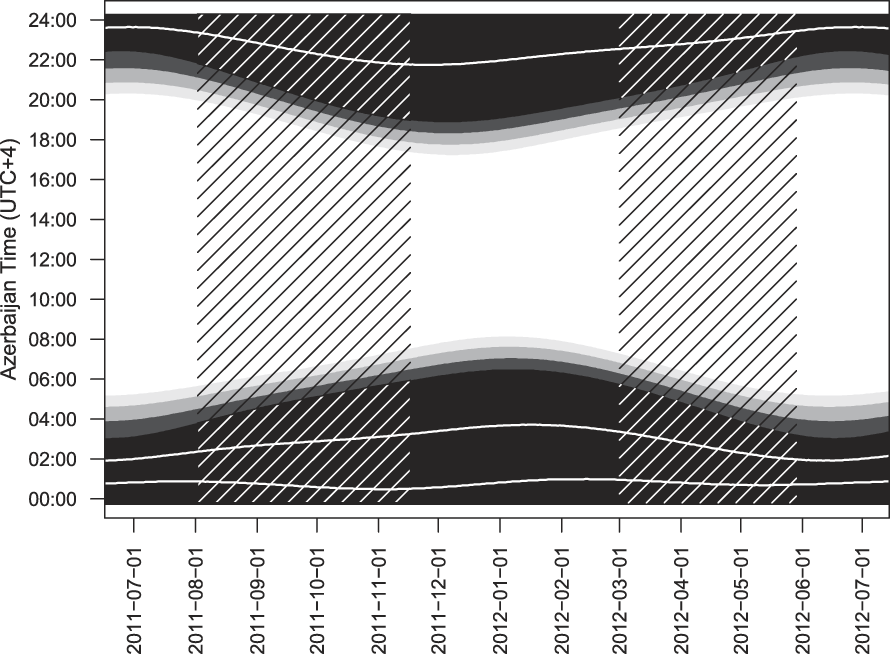
Figure 1. Astronomical data during the periods of field observation (shaded). White area = daytime (solar elevation angle > 0°), light grey = civil twilight (solar elevation angle 0° to -6°), medium grey = nautical twilight (solar elevation angle -6° to -12°), dark grey = astronomical twilight (solar elevation angle -12° to -18°), black = nighttime (solar elevation angle < -18°). The nighttime is subdivided into four equal quarters (white lines).
Due to organisational or technical issues (e.g. unexpected depletion of batteries) not all recording sessions in the two study periods covered the entire night. For the subsequent analysis, only recordings containing more than 90% of the full night length were taken into consideration. Thus from autumn 2011 in total, 750 h 31 min of acoustic documentation from 63 nights have been analysed and from spring 2012 714 h 13 min from 67 nights.
The analysis was carried out with the help of the software Songscope (version 4.1.3A, Wildlife Acoustics Inc.) by listening and visually scanning through displayed spectrograms (FFT Size 256, FFT overlap ½, background filter 2s, frequency range 0–14,500 Hz, linear scaling). The use of call recognition software to speed up the call analysis was rejected on the grounds that Songscope would require a comprehensive knowledge of the occurring bird species and their calls for which to search (Agranat Reference Agranat2007, Reference Agranat2009). Other software using e.g. band-limited-energy detectors often produce a high number of false signals (Ross and Allen Reference Ross and Allen2014). The recently released software ‘Kaleidoscope’ (Wildlife Acoustics Inc.) was not available when analysing the data, but promises to be the right tool to investigate long-term recordings, where call types are unknown. Nevertheless, to arrive at the clearest picture of nocturnal migration a manual analysis was necessary, which, although much more time-consuming than the above-mentioned automated search for known call types (Swiston and Mennill Reference Swiston and Mennill2009), guarantees the detection of all sounds relevant to the study.
Each call that was found was annotated and wherever possible identified. Identifications were based on personal knowledge, in reference to various recordings from the http://www.xeno-canto.org website, van den Berg et al. (Reference van den Berg, Constantine and Robb2003), Schulze (Reference Schulze2003), exercising sufficient caution as advocated by Robb (Reference Robb2004), and Bergmann et al. (Reference Bergmann, Helb, Baumann and Engländer2015). If personal knowledge was not sufficient to identify calls, that of other experts (see acknowledgements) was gratefully called upon. Identifications both without and with residual uncertainty (perhaps owing to weak signals, as a result of recording at too great distance, or of obtruding background noise) were categorised according to their quality: a = secure identification (with a subjective identification confidence level of 100%), b = almost secure identification (99% identification confidence), c = identification most likely correct (95% identification confidence). Calls identified with lower confidence levels have been classified as unidentified (in full awareness that many calls of nocturnal migrants in general, and of those in this little-studied region in particular, are unknown). Furthermore, bird songs and any vocalisations of resident bird species have been left out of consideration in order to maximise the probability that the annotated calls are those of (at least potentially) migrating individuals. Thirteen breeding species were completely disqualified in this way: Eurasian Hobby Falco subbuteo, Common Cuckoo Cuculus canorus, Common Barn-owl Tyto alba, Eurasian Scops-owl Otus scops, Little Owl Athene noctua, Northern Long-eared Owl Asio otus, Eurasian Golden Oriole Oriolus oriolus, Eurasian Magpie Pica pica, Crested Lark Galerida cristata, Menetries’s Warbler Sylvia mystacea, Rufous-tailed Scrub-robin Cercotrichas galactotes, Common Nightingale Luscinia megarhynchos and Isabelline Wheatear Oenanthe isabellina. In a number of other species the differentiation of migrating individuals from resident breeding individuals appeared impossible and was thus not attempted. The calls of these species have been included in the analysis in view of the fact that there may be considerable overlap between these two groups (cf. status in Appendix S1 in the online supplementary material). Excluded, however, were calls coming from an extremely vocal breeding colony of Black-winged Stilts Himantopus himantopus from the night of 9/10 April 2012 until the end of the study period in spring 2012, a time during which the level of their calling activity was inadmissibly high. Two bird species, Mute Swan Cygnus olor and Little Bustard Tetrax tetrax, were identifiable by the sound of their wing beats. As neither is otherwise very vocal in flight, each overflying event, which may involve differently sized flocks or single birds, was regarded as a ‘call’ in the subsequent analysis.
For each night of each study period and for each species under consideration a value was established for the number of calls detected. These when added together yielded the seasonal total call numbers. As mentioned above, the recordings obtained on some nights were not analysable thus leaving gaps in the data. In order to arrive at more accurate seasonal totals the missing data were interpolated according to the following simple system. The difference between the number of calls recorded on the night before a gap and the number recorded on the night after the gap was divided by the number of nights in the gap plus one. The resulting average difference per night was then taken as the nightly increment (or decrement) in the number of calls throughout the data gap, thus creating a straightforward linear progression. An example will serve to make this system clear. On the night of 3/4 September (let us suppose) 40 calls of a particular species are recorded in a valid session. No valid data are collected on the following two nights, but a valid session on 6/7 September records 70 calls. In the subsequent analysis the difference of +30 calls will be divided by 3 (the gap of 2 nights + 1), resulting in a gradient of +10 calls per night. The data gap will thus be repaired with values of 40 + 10 (= 50 calls) for the 4/5 September and 40 + 10 + 10 (= 60 calls) for the 5/6 September.
The only problem with this arises when a species under analysis produces an exceptionally high number of calls on one of the nights bracketing the data gap, as this method of interpolation then leads to an overestimation of numbers. To avoid this, single nights contributing more than 50% of a species’ total number of calls in a season have been excluded from these interpolations. In such cases the calculation was based instead on the counted totals of the analysed night before and the analysed night after the night of high activity.
For these calculations all three call identification qualities (see above) were taken into account. Furthermore, on the assumption that environmental background noise would decrease the detectability of calls in the recordings, each night was evaluated separately with respect to different background noise types, such as animals (insects, amphibians, bird songs), wind, rain or surf noise from the shoreline. Data analysis was carried out with the software R version 3.2.3. (R Foundation for Statistical Computing 2015).
Results
A total of 88,455 calls was found in the recordings of which 2,172 (2.5%) have been left unidentified. The vocalisations of 119 bird species were identified, 106 of which can be expected to occur on migration in the study area. The most vocally evident species between sunset and sunrise in autumn 2011 was Black-winged Stilt followed by Eurasian Skylark Alauda arvensis and Western Yellow Wagtail Motacilla flava (Appendix S1). In spring 2012 the Western Yellow Wagtail was the most vocal species (Appendix S1). After the interpolation of values for those nights without valid data, a total estimate was reached of 108,986 calls in autumn 2011 and 33,348 calls in spring 2012, a grand total of 142,334 calls for the two seasons together (Appendix S1).
In absolute terms, in both study periods most bird calls were recorded during morning civil twilight and numbers were lowest in the astronomical evening and astronomical morning twilight (Figure 2). On a relative scale (i.e. calls per hour) the lowest call rates in both recording seasons were in the four night quarters and astronomical twilights. Higher rates are associated with evening and morning nautical and civil twilights, the highest values of 1,027.7 calls/h in autumn 2011 and 477.1 calls/h in spring 2012 (Figure 2) being reached in civil morning twilight.
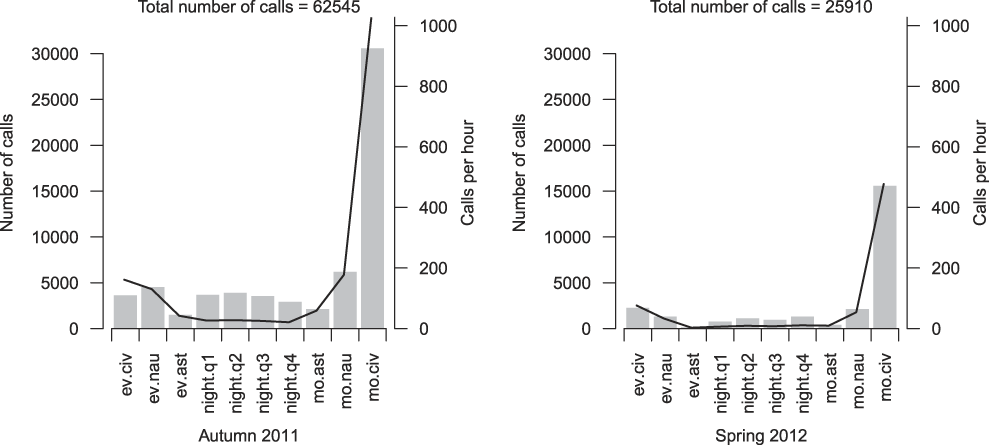
Figure 2. Absolute call numbers (grey bars) and average number of calls per hour (black line) for each twilight and night phase. Autumn 2011 covers 63 recorded nights and spring 2012 covers 67 nights. Abbreviations: ev.civ = evening civil twilight, ev.nau = evening nautical twilight, ev.ast = evening astronomical twilight, night.q1 = first night quarter, night.q2 = second night quarter, night.q3 = third night quarter, night.q4 = forth night quarter, mo.ast = morning astronomical twilight, mo.nau = morning nautical twilight, mo.civ = morning civil twilight. For details see Appendix S2.
With regard to the occurrence of species per night/twilight phase, in both seasons the greatest number of species was recorded during morning civil twilight, with 55 and 50 species for autumn 2011 and spring 2012, respectively. The next highest species numbers occurred, in descending order, in the nautical twilights and the four night quarters. The lowest species numbers were produced by the two astronomical twilight phases (Figure 3). In terms of species per hour, the night quarters yielded the lowest quotients, whereas the twilights returned a higher species density (Figure 3).
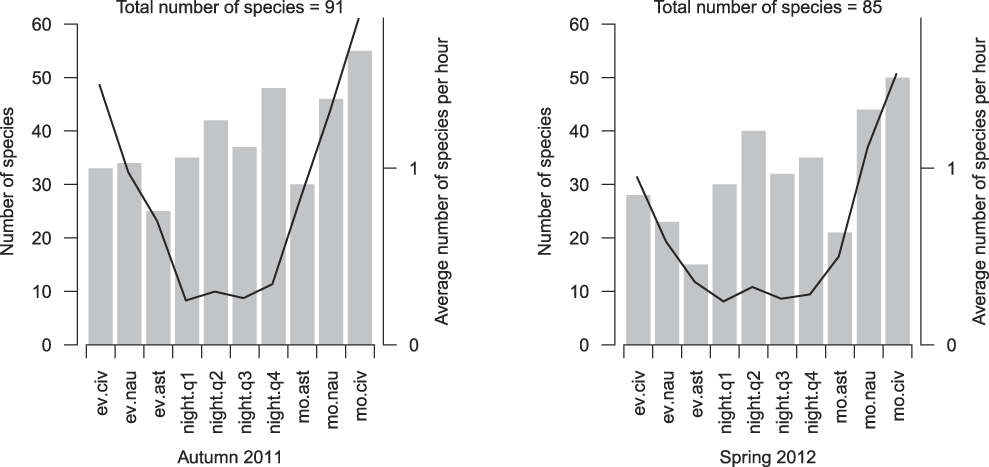
Figure 3. Number of identified species (grey bars) and average number of species per hour for each twilight and night phase (black line). Autumn 2011 covers 63 recorded nights and spring 2012 covers 67 nights. Abbreviations: ev.civ = evening civil twilight, ev.nau = evening nautical twilight, ev.ast = evening astronomical twilight, night.q1 = first night quarter, night.q2 = second night quarter, night.q3 = third night quarter, night.q4 = forth night quarter, mo.ast = morning astronomical twilight, mo.nau = morning nautical twilight, mo.civ = morning civil twilight. For details see Appendix S2.
The intensity of calling activity varied noticeably throughout the two study periods. It reached its peak on 2/3 August 2011 with 4,208 calls recorded during the twilight and night hours. The spring season showed a maximum on the night of 9/10 May 2012 with 1,341 calls.
In general, calling rates were higher in the autumn with an average of 992.8 calls per recorded night, compared to 386.7 calls in the spring. Nocturnal migration in its strictest sense (i.e. night quarters only) was accompanied by more bird sounds in autumn than in spring with averages of 222.7 and 61.9 calls respectively per recorded night (Figure 4).
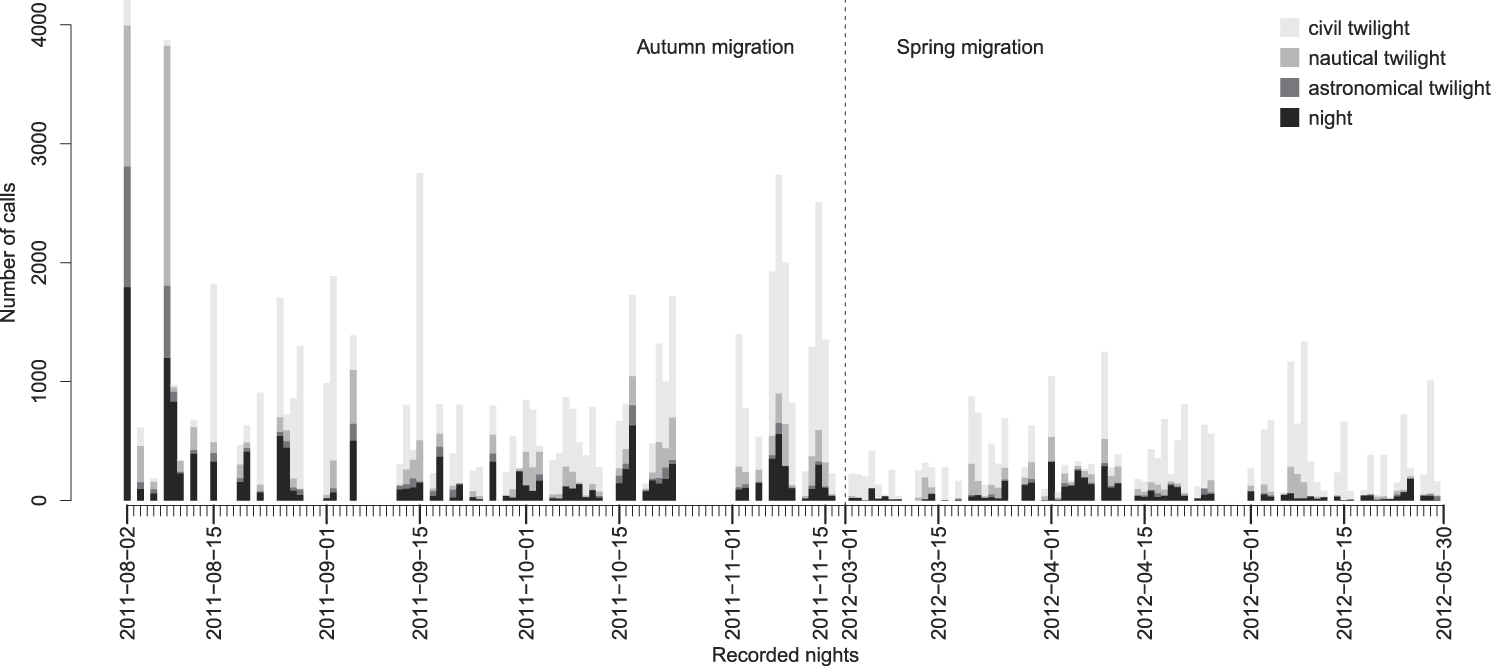
Figure 4. Phenology of all recorded calls differentiated according to the twilight and night phases. For details see Appendix S2.
The analysis of the sound recordings in both study periods suffered to varying degrees from background noise caused by wind, sea and, to a much lesser extent, by rain, which had a comparatively low impact on the recordings. At the appropriate season, background noise was also caused by the activity of local animals, such as insects (e.g. crickets) that occurred in autumn 2011 until mid-October. In spring 2012, from April onwards the recordings were affected by insects, amphibians and birdsong (Figure 5).
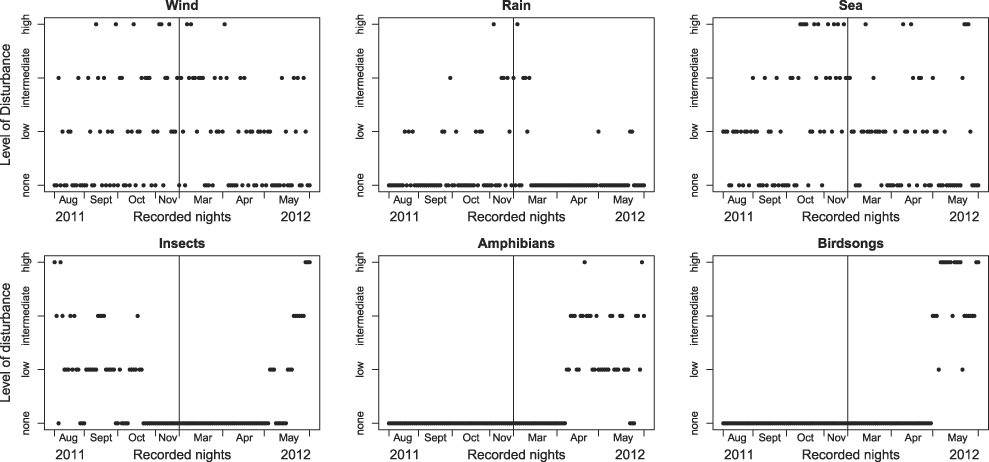
Figure 5. Level of disturbance from various background noise sources liable to disrupt the sound analysis.
Discussion
Species and their identification
In this acoustic-based study, the only measureable criteria available to evaluate the presence of a nocturnal bird migration bottleneck are the abundance of species and, later discussed, the number of calls occurring during the observation hours.
About one-third of the total number of species recorded at Besh Barmag, including diurnal observations and breeding birds (Heiss Reference Heiss2013), were acoustically identified, in all 119 species (listed in Appendix S1). However, this figure includes a number of typically diurnal species extending their migration into the half-light of dusk and dawn, twilight phases whose inclusion in this study was imperative in order to take account of several nocturnal species which frequently start migration at dusk and continue till around dawn (e.g. herons, thrushes). Other studies, e.g. radar results and references in Dinevich et al. (Reference Dinevich, Matsyura and Leshem2003), showed that nocturnal migration often starts shortly after sunset and lasts until sunrise and thus overlaps the migration activity of typically diurnal species. The number of ‘true’ nocturnal migrants is therefore lower. As can be seen in Appendix S1, for 27 species there were no records in the darkest phases of the night (night quarters and astronomical twilights), and a further 22 species occurred with less than 5% of their total calling volume. In contrast, 17 species were recorded exclusively in these dark night phases (Appendix S1), 13 of which, however, were visually observed on migration during daylight hours. In summary, out of 106 species 80 displayed nocturnal calling behaviour (Appendix S1).
This figure must be regarded as very conservative as it admits only those species of which at least one call could be assigned to identification confidence category ‘a’ (= 100% certain; see above). It is to be expected that a certain number of the calls of category ‘b’ (= 99% certain) and ‘c’ (= 95% certain) will have been correctly identified, which will lengthen the list. Furthermore, calls that were completely unknown are quite likely to belong to yet further species absent from the initial list. Altogether, these undetermined calls may refer to about 20 additional species. It should be also mentioned that many nocturnal migrating species are silent on passage (e.g. Sylviidae) and will never be acoustically recognised at night.
More accurate statements would require far more knowledge about nocturnal calls, and concomitantly, a reduction of the volume of unknown calls. One possible method of augmenting reference material might be to use temporarily captured birds in a ‘recording chamber’. Here, nocturnal calls stimulated naturally by zugunruhe or triggered artificially by sound reproduction devices can be systematically recorded and analysed (Lanzone et al. Reference Lanzone, DeLeon, Grove and Farnsworth2009). More information about the nocturnal species inventory, which would help guide the identification of some of the unknown calls, could be gained by mist-netting in the shrubby coastal woodland in this area, which is a good stop-over site for migrants (Heiss and Gauger Reference Heiss and Gauger2011). Sanders and Mennill (Reference Sanders and Mennill2014) found that captured species and numbers are correlated with nocturnal calling activity, showing that this method is also capable of providing statistical material about potential nocturnal callers. Another method of approaching a more complete species inventory might be the search for potential nocturnal migrants among the victims of collision with power lines, which are widespread in the study area, as well as at sites such as tall and brightly lit towers about 80 km away in the capital Baku. The list of casualties at such towers often provides valuable information about nocturnal species composition and is also linked with levels of nocturnal migration activity (Graber Reference Graber1968).
For the moment, though, in the absence of this kind of additional data, all that can safely be asserted here is that 106 plus c.20 nocturnal migrant species are liable to be heard between sunset and sunrise. A comparison of this number with results from other study sites appears to be difficult on account of differences in recording techniques, study design and availability of local knowledge of migrant calls. Nonetheless, the numbers from similar studies elsewhere, e.g. 67 species in Canada (Sanders and Mennill Reference Sanders and Mennill2014), 30–50 species in the Gulf of Mexico (Farnsworth and Russell Reference Farnsworth and Russell2007) and 97 species in the German Bight, a figure which also took diurnal vocalisations into account (Hüppop et al. Reference Hüppop, Dierschke, Hill and Hüppop2012), are all slightly or considerably smaller than the number from the Azerbaijan site, which is therefore presumably a good location for nocturnal migration.
Nocturnal call numbers
Here, too, the inclusion of twilight phases led to inflated totals through the interference of the calls of diurnal migrants. The majority of the recorded calls (54.7% in autumn 2011 and 68.8% in spring 2012) were in fact made in the civil twilights (Figure 2), and although some of the calls occuring during these phases certainly came from nocturnal migrants, most by far were attributable to diurnal species. For example, in the civil twilight of autumn 2011, the most vocal migrant, the Yellow Wagtail, accounted for 6,830 calls, and a further 5,323 during the same phase in spring 2012 (Appendix S1). There is, however, no denying that the 17,640 calls recorded in the darkest night phases (night quarters and astronomical twilights) in autumn 2011 and the 4,672 in spring 2012 (Figure 2) do indicate a considerable amount of nocturnal movement. Particularly conspicuous were the nocturnal calling activities of herons (Great Bittern Botaurus stellaris, Little Bittern Ixobrychus minutus, Grey Heron Ardea cinerea, Purple Heron Ardea purpurea and Black-crowned Night Heron Nycticorax nycticorax), rails (Common Moorhen Gallinula chloropus and Eurasian Coot Fulica atra, plus an additional five species in fewer numbers), waders (Little Ringed Plover Charadrius dubius, Black-winged Stilt, Green Sandpiper Tringa ochropus and Wood Sandpiper Tringa glareola) and thrushes (Song Thrush Turdus philomelos) (Appendix S1).
Call numbers alone and unsupported by visual observation (ceilometers, radar etc.) are, of course, insufficient to provide reliable information about the number of migrating individuals (Graber and Crochan Reference Graber and Cochran1960, Hüppop et al. Reference Hüppop, Dierschke, Hill and Hüppop2012), a considerable limitation of the acoustic data collection method. As far as the nocturnal calling activity of birds is concerned, studies have shown that the range of behaviours both within and among species (Dierschke Reference Dierschke1989, Evans and Rosenberg Reference Evans, Rosenberg, Bonney, Pashley and Cooper2000, Evans and O’Brien Reference Evans and O’Brien2002, Farnsworth Reference Farnsworth2005) and individuals (Farnsworth Reference Farnsworth2005) varies between the extremes of all members of a group calling continually and no member of the group calling at all. Furthermore, even members of those species that call most frequently may just happen to be silent when passing over the microphone (Kunz et al. Reference Kunz, Arnett, Cooper, Erickson, Larkin, Mabee, Morrison, Strickland and Szewczak2007). Consequently, it can by no means be assumed that a migrant species whose nocturnal calls are heard frequently is common or that a species which is rarely or never heard is not.
Furthermore, not all of the vocal activity giving rise to the acoustic data can be accepted as an indication of migratory movement. Locally breeding or resting birds are particularly prone to overinflate call count results, their nocturnal vocalisations being easily misinterpreted as signs of great migration activity. Although some of the error from this source was eliminated by the exclusion from the analysis of known 13 breeding species (see Methods), some of the calls that were taken into account may have been made by individual members of migrant species breeding undetected in the study area and may thus be totally unconnected with nocturnal migration.
As a consequence, the call numbers of suspected breeding species (also listed in the Methods section) should be interpreted with great caution as the settled presence in the study area of any breeding individuals in the spring period (April–May) and the early autumn period (August) will result in a distortion of the nocturnal acoustic footprint.
Another foreseeable source of overestimation are the calls of birds spending the night within range of the microphone in one or other of the attractive resting habitats nearby, which include steppe (e.g. for larks and Little Bustard), scrub (e.g. for finches, buntings and bee-eaters) and a coastal lagoon (e.g. for waders and ducks). These may be contact calls from individuals resting or feeding at night, but may also come from local movements, probably flights between different feeding grounds. Or they may be caused by disturbance at night e.g. from nocturnal predators. In the dark, the calls of a group of disturbed waders in flight, for example, might be difficult to differentiate from those of a flock on passage. On the other hand, nocturnal events of this nature involving typically diurnal migrants are easier to interpret. For example, the remarkable vocal outbursts staged by European Bee-eater Merops apiaster and Blue-cheeked Bee-eaters Merops persicus were more likely to have been rooted in this kind of nocturnal disturbance than to have come from a migrating flock (Appendix S1 and S2).
In contrast to other study sites where, especially in unfavourable weather conditions with limited visibility (Graber and Cochran Reference Graber and Cochran1959, Reference Graber and Cochran1960, Evans and Mellinger Reference Evans and Mellinger1999, Farnsworth and Russell Reference Farnsworth and Russell2007), significant numbers of nocturnal migrants gathered, attracted by the presence of artificial light sources e.g. lighthouses (Jones and Francis Reference Jones and Francis2003), towers (Avery et al. Reference Avery, Springer and Cassel1976) or cities (Watson et al. Reference Watson, Wilson and Mennill2016), the Besh Barmag bottleneck area suffered no interference from light trespass. Call counts were therefore not biased by the commonly observed behaviour of birds repeatedly flying around light sources at night and frequently uttering flight calls evocative of heavy migration, a consideration which should be borne in mind in comparisons with the results obtained e.g. at oil platforms (Farnsworth and Russell Reference Farnsworth and Russell2007) and offshore platforms (Hüppop et al. Reference Hüppop, Dierschke, Hill and Hüppop2012).
Interference to the opposite effect (i.e. an underestimation of call numbers), albeit of minor extent, is expected to have taken place through the masking of calls by ambient background noise (Figure 5). The detection of low frequency calls was hampered by low frequency sounds produced by wind or surf noise from the sea. Especially when distant, the calls of birds may have been discounted in spectrograms as those of similar sounding crickets or amphibians. The overall impact of background noise in this study can, however, be regarded as low. Plainly then, the number of calls recorded is a parameter with many uncertainties and imponderables on which no evaluation of Besh Barmag as a nocturnal migration bottleneck can be solely based.
Conclusion and outlook
The results obtained by this study indicate a considerable amount of nocturnal movements. Although the nocturnal call rates are consonant with results from radar studies at other sites (Larkin et al. Reference Larkin, Evans and Diehl2002, Farnsworth et al. Reference Farnsworth, Gauthreaux and van Blaricom2004), Besh Barmag will require a radar study of its own. For this task the deployment of vertical radar detection, although rather expensive, is an obvious choice, the clarity of the results it obtains by capturing individual bird signals at night brings to light the true magnitude of nocturnal migration (Lack and Varley Reference Lack and Varley1945, Desholm et al. Reference Desholm, Fox, Beasley and Kahlert2006, Kunz et al. Reference Kunz, Arnett, Cooper, Erickson, Larkin, Mabee, Morrison, Strickland and Szewczak2007, Gauthreaux et al. Reference Gauthreaux, Livingston and Belser2008, Bruderer et al. Reference Bruderer, Steuri, Aschwanden and Liechti2012). Bottleneck sites where radar (or other visual methods) has already proved successful include Israel (Bruderer et al. Reference Bruderer, Underhill and Liechti1995, Bruderer and Liechti Reference Bruderer and Liechti1995, Dinevich Reference Dinevich, Leshem and Matsyura2005), Falsterbo (Alerstam Reference Alerstam1972, Zehnder et al. Reference Zehnder, Åkesson, Liechti and Bruderer2001) and the Strait of Messina (M. Panuccio pers. comm.).
A further advantage of radar is to supplement important information about migration altitude, which is essential in environmental impact studies. At Besh Barmag, for instance, the observed heights of passing diurnal migrants were low (Heiss Reference Heiss2016) and it would be of the greatest interest to know whether this was also the case with nocturnal migrants in the pitch-black of night when the risks of collision with obstacles is even higher.
Supplementary Material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0959270917000454
Acknowledgements
I wish to thank Elchin Sultanov (Azerbaijan Ornithological Society) for his support during the fieldwork and for useful literature and additional information. Vasil Ananian, Victor P. Belik and Elchin Sultanov provided valuable data on nocturnal migration in their respective ‘home’ regions. Nils Kjellén, Michele Panuccio, Jean-Paul Urcun, Noam Weiss and Edward Bonavia supplied details of nocturnal research activity at bottleneck sites within the Western Palearctic. I thank also Patrick Franke, Roland Neumann, Lukas Pelikan, Magnus Robb and Nina Seifert for their help in identifying unfamiliar and confusable bird calls. Mike Wilson helped with literature. Ken Wilson checked the English. This study was supported financially by the German Academic Exchange Service (DAAD).






