


Rising global levels of obesity and type 2 diabetes (T2D) are major public health problems facing the human race and are putting considerable load on healthcare resources. Obesity and T2D are closely interlinked, with the rise in T2D mapping the rise in obesity( Reference Larsson, Alicja Wolk and Mantzoros 1 ). Over the centuries, the diet consumed by humans has changed from an energy-dilute diet based on fruit, vegetables and fish to an energy-dense diet of processed foods rich in starch, sugar and fat. The proposal that man currently eats a diet to which, in evolutionary terms, he is poorly adapted was first put forward in 1975 by Voegtlin( Reference Voegtlin 2 ), a concept taken up and developed by Boyd-Eaton & Konner( Reference Boyd Eaton and Konner 3 ) in 1985. They suggested that what they termed the Palaeolithic diet (i.e. diet of humans before the Neolithic Revolution) was likely to be high in protein and rich in plant matter, while containing no cereals or dairy products, and proposed this as a nutrition strategy better fitted for the human genome. It would also be expected that such a diet, while involving cooking, would have less in the way of processing than is found in modern foods. This is based, in part, on the theory that, as farming only resulted in the consumption of dairy and cereal products (that were themselves dependent on more intensive processing) in the past 7000–10 000 years, man has not had sufficient time to evolve to such a drastic change in diet. Recent studies on the health benefits of Palaeolithic-style diets have been conducted over periods of weeks( Reference Frassetto, Schloetter and Mietus-Synder 4 , Reference Boers, Muskiet and Berkelaar 5 ) or months( Reference Lindeberg, Jönsson and Granfeldt 6 – Reference Mellberg, Sandberg and Ryberg 8 ), and found benefits including reduced blood pressure and cholesterol levels and improved glucose tolerance. There is also evidence to suggest improved satiety leading to weight loss for this form of diet( Reference Jönsson, Granfeldt and Erlanson-Albertsson 9 ). However, to date, little mechanistic insight into these observations has been offered, although the reported improvement of glucose tolerance suggests a link to glycaemic control. To investigate this, we designed two test meals (Palaeolithic meal 1 (PAL1) and Palaeolithic meal 2 (PAL2)) to test our hypothesis that acute effects on carbohydrate metabolism of this diet was a key element of the health benefits so far observed, taking as a starting point the basic principles of a Palaeolithic diet but using modern food ingredients. Diets used in previous studies, with emphasis on salad and vegetable preparation ‘from scratch’ would have long-term acceptability and compliance issues for the modern-day consumer. Therefore, we decided to develop Palaeolithic diet principles while using modern foods and meal design, but keeping processing to a minimum to maintain plant structures. These mixed meals therefore consisted of fruit, vegetables and protein, but lacked starchy cereals, dairy products, legumes, and refined fats and sugars. In addition, although cooking of food was required, the plant material chosen for the study was used fresh, where possible, and minimal processing in terms of the plant structure was used. These were compared with a reference meal (REF) based broadly on the current WHO nutritional guidelines. Many Palaeolithic diets propose a high level of animal protein, and to separate the possible effects of this from the fruit and vegetable component, PAL2 was designed to have a lower level of protein that matched that of the REF, while PAL1 was designed to have the higher level of protein used by other authors, but with the same level of carbohydrate.
We investigated the acute effects of the consumption of these Palaeolithic-inspired meals on metabolism. Specifically, we investigated the parameters that are thought to influence glucose control including indices of insulin sensitivity and secretion, as these are the features of meals that would be expected to have a low glycaemic index, based on the highly unprocessed plant matter and low starch content. In addition, we explored satiety, gut hormone and incretin responses. The gastric hormones studied included glucagon-like peptide-1 (GLP-1) and glucose-dependent insulinotropic peptide (GIP) that play a role in the insulin response, while GLP-1 and peptide YY (PYY) are involved in satiety response and ileal brake mechanism. It was hypothesised that Palaeolithic meals would have a lower glucose response, and that this would be indicated by altered incretin and satiety responses.
Materials and methods
The present study was conducted according to the guidelines laid down in the Declaration of Helsinki, and all procedures involving human subjects were approved by Independent Ethics Committee (REC reference 11/LO/0911) NRES Committee South East Coast, Kent, UK. Written and informed consent was obtained from all volunteer subjects. The present trial was registered at www.clinicaltrials.gov, as ‘Glycemic Response After Palaeolithic-type Meals’ (ID no. NCT01418573).
Meal development
The REF was designed based on macronutrient proportions from WHO Technical report 880( 10 ), and contained protein, fruit and vegetables as well as cereals. Polished (but not parboiled) rice was chosen as a carbohydrate/cereal component. Rice was used, as opposed to other plant starch materials such as pasta, as minimal processing of the plant material occurs before cooking and the rice could all be sourced from a single batch for consistency throughout the trial.
PAL1 was based on estimated range ratios for protein and fat that are typical of Hunter–Gatherers and contained no cereals or dairy products, as updated recently( Reference Konner and Boyd-Eaton 11 ), although carbohydrate was slightly higher to allow matching across all three meals.
PAL2 was made with identical plant-based ingredients to PAL1, but normalised to the REF for fat, protein and energy in addition to available carbohydrates, by changing the fish, nut and strawberry content.
All the three test meals were normalised to contain 50 g of available carbohydrates by calculation from literature values (available at http://nutritiondata.self.com).
Full ingredients for all the test meals are presented in Table 1. Ingredients for the PAL meals were selected according to the previously defined Palaeolithic principles, in that they included a wide range of fruit and vegetables, but no cereals or dairy products( Reference Boyd Eaton and Konner 3 ). Additionally all the plant-based ingredients were selected as rich in polyphenols (e.g. strawberries( Reference Wang, Wei and Galletta 12 )) and fibre, as we hypothesise that these would be higher in ancestral plants because of their broader genetic variation( Reference Tanksley and McCouch 13 ). As necessity of supply meant that the plant material used was from commercial sources and, therefore, not likely to be as high in phytochemicals due to selective breeding, phytochemical levels were boosted by the addition of cinnamon( Reference Anderson, Broadhurst and Polansky 14 ) and capers( Reference Inocencio, Rivera and Alcaraz 15 ). Fish was used as a protein source across all the three test meals, although PAL1 contained white fish as this enabled boosting of protein levels while keeping the fat content of the meal constant with the other two.
Table 1 Macronutrients and ingredients for the three test meals*

PAL1, Palaeolithic meal 1; PAL2, Palaeolithic meal 2; GAE, equivalents of gallic acid.
* Calculated values were obtained using the United States Department of Agriculture values (available at http://nutritiondata.self.com).
† The calculated values for PAL1 and PAL2 were high, as the analysis of an initial iteration meal with the original calculated values of carbohydrate gave actual values of available carbohydrate that were lower than those expected from the calculation. The recipe was thus altered to give higher carbohydrate values to get closer to the desired target. Values presented are from the analysis of a subset of meals cooked at the time of the study and are lower than those from the initial analysis of an earlier batch, presumably due to seasonal and batch variations found in natural fruit and vegetable-based foods.
‡ Components that were added to the meal only upon serving could be calculated as cooked.
§ Weight was calculated based on group 2 analysis of rice purchased and cooked as described in the Materials and methods section.
Meals were prepared and cooked in single batches and frozen before the start of the study, with ingredients all sourced at the same time. For the Palaeolithic meals (PAL1 and PAL2), the fruit and vegetable content was batch stir-fried as individual portions. Other components such as fish and carrots were precooked and frozen as individual portions to be added on the morning of the study. All food was reheated on the morning of the study and served hot, with the exception of rice for the REF, which was cooked fresh each morning. Samples of each meal were submitted for nutritional analysis (Leatherhead Food Research). To match the carbohydrate content of the REF and to allow for a possible variation between the two types of rice and cooking methods, a sample of long-grain rice bought for the study was cooked in a rice cooker, according to the manufacturer's instructions, macronutrient content analysed and the amount needed for the meal calculated. Then, the same batch of rice and cooking method were used each time on the morning of the study.
Phenolic (Folin) assay
Meals were frozen in liquid N2, freeze-dried, ground in a blender, and the dry powder was stored at − 80°C under N2. Briefly, 20 g of the lyophilate were added to 200 ml of 70 % aqueous ethanol containing 5 g ascorbic acid/l and stirred for 2 h at 22°C in a sealed flask. The suspension was filtered, the residue washed with 50 ml of the same solvent and the filtrates combined. Ethanol was removed by rotary evaporation at 40°C, the residue made up to 100 ml with water and the pH adjusted to 7. The solution was passed through an Amberchrom CG300C column (pre-washed in 500 ml of ethanol and equilibrated with 500 ml of water at pH 7). The column was then washed with 500 ml of water at pH 7, and the phenolics eluted in 750 ml of ethanol. Ethanol was removed by rotary evaporation at 40°C, and the residual water by freeze-drying. The residue was redissolved in 70 % aqueous methanol and assayed by the method of Velioglu et al. ( Reference Velioglu, Mazza and Gao 16 ). Results are expressed as equivalents of gallic acid in mg (mg GAE).
Volunteers
A total of twenty-four apparently healthy male subjects (plus four reserves) aged 18–60 years, with a BMI (calculated as weight/height2) between 18 and 27 kg/m2, were recruited. The sample size calculation was based on postprandial glucose positive incremental AUC over 0–120 min (+iAUC0–120 min) response and variance estimates from a previous meal study run at the same facility. The mean +iAUC in the previous study was 126, and the residual mean square was 37·3. We expected that the control +iAUC in the present study could be different as a full meal was consumed as opposed to just bread in the earlier study; however, we expected that the residual mean square could be similar. A reduction of 37·8 (30 %) in +iAUC0–120 min from the previous control mean would require twenty subjects for the reduction to be detected as significant at P< 0·05 with 80 % power. To allow for dropouts and because of logistical limitations (maximum six subjects per d), a sample size of twenty-four subjects was chosen.
Subjects were randomly allocated to four cohorts of six subjects. Of these cohorts, two attended each week over a 6-week period, giving three visits per cohort with a 2-week recovery between visits. Allocation of the meals was randomised according to a Williams design within each cohort. Allocation to cohort and of treatment was carried out by an independent statistician who was not involved in the assessment of the study. The investigator and outcome assessor were blinded, while due to the nature of the test material, caregiver and subjects were not blinded. Treatments were pre-randomised to subject numbers. Subjects were assigned a number based on the sequence of entering the study. Subjects were only aware that they were being asked to eat three different meals, no expectations regarding their response to the meals were suggested, and they were unaware as to which meal was reference and which was experimental.
Metabolic study protocol
The intervention elements of the study were carried out at the clinical investigation unit of Leatherhead Food Research (Leatherhead, Surrey, England), which recruited participants in July and August 2011 and assigned them to cohorts. Statistical analysis was performed by Unilever Discover, which were initially blinded to the treatments. Volunteers were asked to fast for 12 h before attending the research unit. All volunteers visited the research unit at 09.00 hours between 19 August and 28 September 2011. On arrival, a cannula was placed in the forearm of all volunteers and baseline measurements were taken. Immediately following this, subjects were fed the meal which they were told to consume within 20 min. If they took longer than 20 min to finish the meal, subjects were excluded at the blind review. Subjects were served the meals in individual booths and were not allowed to talk. Subjects were fed at 10 min intervals for ease of sample taking later.
Study measurements
During each of the three interventions, sixteen venous blood samples were collected via an indwelling cannula (eight samples of 3 and 5 ml for gut hormones and glucose) into BD-P800™ blood (B&D) collection tubes containing a cocktail of protease, esterase and DPP-IV inhibitors and eight samples of 3 and 5 ml for insulin into heparinised tubes. Samples were collected at -15, 15, 30, 45, 60, 90, 120 and 180 min relative to the start of consumption of the meal.
Venous plasma glucose analysis as a primary outcome was conducted on a Horiba ABX Pentra 400 Clinical Chemistry Analyser using the commercially available Glucose PAP CP Kit (reference no. A11A01668; Horiba Medical).
Insulin (primary outcome) in heparinised plasma was analysed using an immulite 2000 Systems analyser, a solid-phase enzyme-labelled chemiluminescent immunometric assay kit (‘ECLIA’ for Roche Elecsys 1010/2010 immunoassay analyser) from Roche Diagnostics GmbH. Its reportable range was 13·89–2083·5 pmol/l with sensitivity 13·89 pmol/l.
Exploratory analysis of gut hormones was performed on blood plasma using MILLIPLEX MAP Human Gut Hormones (HGT-68K 08), EMD Millipore Corporation. This is a multiplex assay kit used to measure ghrelin, leptin, GIP, GLP -1, amylin (active and total), pancreatic polypeptide, PYY and Insulin in human plasma. GLP-1 was repeated by a commercially available ELISA assay (code 27 784; IBL Limited), as the MILLIPLEX MAP system failed to give sufficient sensitivity for this analyte.
Secondary outcome ratings of appetite: feelings (how hungry are you? how full are you? and how strong is your desire to eat?) were scored using a mark on 60-mm scales using EVAS (electronic visual analogue scale)( Reference Flint, Raben and Blundell 17 – Reference Stratton, Stubbs and Hughes 19 ) anchored at the low end with the most negative or lowest-intensity feelings (e.g. extremely unpleasant, not at all), and with opposing terms at the high end (e.g. extremely pleasant, very high, extreme). Volunteers were asked to indicate on a line which place on the scale best reflects their feelings at that moment. Scores were collected so that they could not be used as a further reference for later scores. Volunteers completed the questionnaires pre- and at regular intervals post-consumption ( − 20, 10, 25, 40, 55, 85, 115, 175 min relative to the start of the meal).
Ratings on physical discomfort (headache, stomach discomfort, bloating, heartburn, nausea and belching) were made on a 4-point scale (0 = not, 1 = little, 2 = moderate and 3 = much) or on a line scale at baseline ( − 20 min) and at 55, 115 and 175. Ratings of taste and liking (e.g. general acceptance, mouth feel acceptance, taste acceptance, smell acceptance) were filled on an EVAS 10 min after consumption of the test product( Reference Stubbs, Hughes and Johnstone 18 , Reference Stratton, Stubbs and Hughes 19 ).
Ratings of liking (overall, smell, taste and mouth feel) were scored by means of a mark on 60 mm scales using the EVAS at 15 min.
There were no reports of serious adverse events. However, three subjects each reported a mild and transient gastrointestinal discomfort, of which two were considered to be unlikely to be related to the study and the remaining (PAL1 consumption) was possibly related.
Statistical analysis
Glucose and insulin concentrations were characterised in terms of the basal and preprandial concentrations (G b and I b, respectively) and, postprandially, the incremental area under the concentration profile for 120 min (for glucose, our concern was with the extent to which each meal raised glucose levels above baseline, therefore only the area above baseline was integrated, glucose +iAUC and insulin iAUC), the maximum concentration attained (glucose C max and insulin C max) and change in concentration between 30 and 60 min following commencement of the meal (glucose Δ30–60 and insulin Δ30–60). In post hoc analysis, indices of basal insulin sensitivity and β-cell function were calculated using homeostasis model assessment as follows( Reference Matthews, Hosker and Rudenski 20 ):
 $$\begin{eqnarray} HOMA-IS = 1/(( G _{b}\times I _{b})/22\cdot 5), \end{eqnarray}$$
$$\begin{eqnarray} HOMA-IS = 1/(( G _{b}\times I _{b})/22\cdot 5), \end{eqnarray}$$
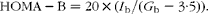 $$\begin{eqnarray} HOMA-B = 20\times ( I _{b}/( G _{b} - 3\cdot 5)). \end{eqnarray}$$
$$\begin{eqnarray} HOMA-B = 20\times ( I _{b}/( G _{b} - 3\cdot 5)). \end{eqnarray}$$
Postprandial insulin sensitivity was expressed by the Matsuda Index, calculated as:
 $$\begin{eqnarray} Matsuda-IS = 1000/( G _{b}\times I _{b}\times G _{mean}\times I _{mean})^{0\cdot 5}, \end{eqnarray}$$
$$\begin{eqnarray} Matsuda-IS = 1000/( G _{b}\times I _{b}\times G _{mean}\times I _{mean})^{0\cdot 5}, \end{eqnarray}$$
where G mean and I mean are the mean postprandial glucose and insulin concentrations( Reference Maki, Kelley and Lawless 21 ). Postprandial β-cell function was expressed as the insulinogenic index, calculated as the insulin concentration 30 min following the start of meal ingestion divided by the accompanying glucose concentration( Reference Maki, Kelley and Lawless 21 ).
Differences between the meals were tested using linear mixed models with subject treated as a random effect (with a variance component error structure) and including fixed effects of baseline (for that meal), subject baseline (i.e. the subject's average baseline overall meals), subject weight, visit number and meal. Each PAL meal was compared statistically with the REF. Because the HOMA-IS, HOMA-B, Matsuda Index and Insulinogenic Index had distributions that were not symmetric, non-parametric Mann–Whitney tests were used to compare the PAL meals with the REF for each of these. The analysis was carried out using SAS Software version 9.3 (SAS Institute).
Results
Of the twenty-four volunteers, twenty-one completed the study. In the blind review, meals where there were protocol deviations (failure to finish the meal in the allotted time) were removed from the per-protocol analysis. After blind review 1, volunteer was removed completely from all analyses, as an obvious outlier in the baseline value for GLP-1 (mean baseline for all subjects 16 pmol/l, excluded subject baseline 997 pmol/l). There were seventeen subjects in the per-protocol analysis set for PAL1 and the same nineteen for both PAL2 and REF. Baseline demographics for the three groups are presented in Table 2.
Table 2 Baseline demographics and clinical characteristics of the study subjects from each group used in the analysis (Mean values and standard deviations)

PAL1, Palaeolithic meal 1; PAL2, Palaeolithic meal 2; REF, reference meal; GLP-1, glucagon-like peptide-1; GIP, glucose-dependent insulinotropic peptide; PYY, peptide YY.
Postprandial glucose and insulin responses
The results for glucose and insulin responses are summarised in Table 3. In post hoc analysis, there were no differences in the basal measures G b, I b, HOMA-IS or HOMA-B before the commencement of each of the three meals. Postprandially, glucose levels with the REF fell between 30 and 60 min to the levels below baseline following the commencement of the meal, and then recovered to basal levels over the next hour (Fig. 1(a)). A similar profile was apparent with PAL1, except that glucose levels remained below baseline from 60 to 120 min. With PAL2, glucose levels also fell from 30 to 60 min but did not fall below baseline and remained at the basal level from 60 to 120 min. Despite these differences, no significant differences were observed in glucose +iAUC120 or insulin iAUC120 between the meals (Table 3, Fig. 1(a) and (b)). Glucose C max did not differ between the meals, but insulin C max was significantly lower (P= 0·040) for PAL2 (mean 264·8 (se 32·42) pmol/l) relative to the REF (mean 339·0 (se 31·40) pmol/l). There was a smaller decline in glucose levels from 30 to 60 min (P= 0·001) for PAL2 (glucose Δ30–60 mean − 0·89 (se 0·14) mmol/l) than the REF (glucose Δ30–60 mean − 1·48 (se 0·14) mmol/l), indicating a slower rate of decline in glucose levels with PAL2. Matsuda-IS did not differ between the meals, but the insulinogenic index was significantly lower (P= 0·02) for PAL2 (median 4·88, interquartile range 3·73–6·73) relative to the REF (median 6·33, interquartile range 4·94–7·86).
Table 3 Postprandial glucose and insulin data for twenty-one subjects after consumption of two Palaeolithically inspired meals (PAL1 and PAL2) compared with a reference meal (REF) (Mean values with their standard errors; medians and interquartile ranges (IQR))

G b, glucose basal; +iAUC, positive incremental AUC; C max, maximum concentration; I b, insulin basal; HOMA-IS, homeostasis model assessment: basal insulin sensitivity; HOMA-B, homeostasis model assessment: basal β-cell function; Matsuda Index-IS, Matsuda Index-insulin sensitivity.
* P values for all tests expect HOMA-IS, HOMA-B, Matsuda Index and Insulinogenic Index were obtained from ANCOVA models with a baseline covariate and 95 % CI. The remaining indices were based on Mann–Whitney non-parametric tests.

Fig. 1 Plasma glucose (a) and insulin (b) concentrations for twenty-one subjects after consumption of Palaeolithically inspired PAL1 (○) and PAL2 (●) and a reference meal (Δ). Values are means, with standard errors represented by vertical bars. * Mean value was significantly different from that of the reference meal (P< 0·05). A Bonferroni adjustment was used to preserve the 0·05 error rate within each time point. To convert insulin in mU/l to pmol/l, multiply by 6·945.
Postprandial gut hormone responses
AUC180 values for all the gut hormones measured are summarised in Table 4. AUC180 values for GLP-1 and PYY (Fig. 2(a) and (b)) were found to be dramatically higher for both PAL meals compared with the REF, while AUC180 values for GIP (Fig. 2(c)) were significantly lower for both PAL meals. Additionally, the ‘shape’ of the GIP response was completely different. The AUC180 for pancreatic polypeptide was significantly higher (P< 0·001) only for PAL1 with respect to the REF, and leptin was significantly lower for PAL2 (P= 0·031). There were no differences between the meals with respect to ghrelin or amylin levels.
Table 4 Postprandial gut hormone and appetite data for twenty-one subjects after consumption of two Palaeolithically inspired meals (PAL1 and PAL2) compared with a reference meal (REF) (Mean values with their standard errors)

PYY, peptide YY; GLP-1, glucagon-like peptide-1; GIP, glucose-dependent insulinotropic peptide; PP, pancreatic polypeptide.
* P values for all tests were obtained from ANCOVA models with a baseline covariate. The remaining indices were based on Mann–Whitney non-parametric tests.

Fig. 2 Postprandial plasma glucagon-like peptide 1 (GLP-1) (a) peptide YY (PYY) (b) and glucose-dependent insulinotropic peptide (GIP) (c) responses after consumption of Palaeolithically inspired PAL1 (○) and PAL2 (●), and a reference meal (REF). Values are means, with standard errors represented by vertical bars. * Mean value was significantly different from that of the reference meal (P< 0·05). A Bonferroni adjustment was used to preserve the 0·05 error rate within each time point. The mean AUC for 180 min for GLP-1 were 1494 (se 66·1) min × pmol/l for PAL1, 1180 (se 64·4) min × pmol/l for PAL2 and 980 (se 63·3) min × pmol/l for the REF (PAL1 v. REF P< 0·001; PAL2 v. REF P= 0·003).
Glucose elimination, insulinogenic index and incretin hormones
Possible explanations for the slower decline in glucose levels with PAL2 were explored in post hoc regression analyses. A slower decline in glucose levels (Δ30–60 more negative) was predicted by a lower insulinogenic index (β = 0·106, P= 0·001) and, in a separate analysis, by PAL2 (β = − 0·492, P= 0·016). When the insulinogenic index was added to the model, PAL2 ceased to be a significant predictor (β = −0·317, P= 0·11) in accord with a lower insulinogenic index contributing the slower decline in glucose levels with PAL2. The GLP-1 response (iAUC between 0 and 30 min) did not predict the insulinogenic index (β = −0·002, P= 0·28), whereas the equivalent measure for GIP was predictive at borderline significance (β = 2·72 × 10− 4, P= 0·074). A lower insulinogenic index was predicted by PAL2 (β = −1·949, P= 0·02). When the GIP response was added to the model, PAL2 ceased to be a significant predictor of the insulinogenic index (β = −1·790, P= 0·14), in accord with the lower GIP response contributing the lower insulinogenic index with PAL2. However, in the stepwise multivariable analysis, with the entry of all potential predictors of the insulinogenic index (BMI, meal type and glucose, GIP, GLP-1 and PYY incremental responses during the first 30 min of the meal), PAL2 was the strongest independent predictor of a low insulinogenic index (β = −2·349, P= 0·002), with initial glucose response as the only other potential independent predictor, albeit at borderline significance (β = 0·013, P= 0·05).
Postprandial satiety electronic visual analogue scale scores
The mean response levels over 175 min for the satiety questions were all statistically significantly different for both PAL1 and PAL2 with respect to the REF (see Table 4). The response level for all the three satiety questions was consistent throughout the time of the study for both PAL meals with respect to the REF, and was still higher for both meals at 175 min (Fig. 3). Percentage changes from the REF for ‘How hungry are you?’ were 44·67 % for PAL1 and 38·44 % for PAL2, for ‘How full are you?’ were − 57·28 % for PAL1, and − 50·59 % for PAL2 and for ‘How strong is your desire to eat a meal?’, 46·94 % for PAL1 and 33·43 % for PAL2. Liking scores for PAL1 and PAL2 were 25·1 and 25·5, respectively, which were significantly lower (P< 0·0001) than those for the REF (60·2).

Fig. 3 Electronic visual analogue scale (EVAS) response scores over time after consumption of Palaeolithically inspired PAL1 (○) and PAL2 (●) and a reference meal (Δ). (a) Response to question ‘How hungry are you?’, (b) response to question ‘How full are you?’ and (c) response to question ‘How strong is your desire to eat a meal?’. Values are means, with standard errors represented by vertical bars. * Mean value was significantly different from that of the reference meal (P< 0·05). A Bonferroni adjustment was used to preserve the 0·05 error rate within each time point.
Discussion
Acute consumption of plant-rich mixed meals gave a significant increase in the release of appetite-related gut hormones GLP-1 and PYY, and improved satiety, but made little impact on glycaemic control. The incretin hormone GIP was dramatically lower and showed a very different response curve for both PAL1 and PAL2 in comparison with the REF.
Glycaemic response
The primary hypothesis that glycaemic control would be improved with plant-rich meals was not supported by the study findings: no difference between the meals was observed in the glucose +iAUC120; neither was there any difference in insulin iAUC120. However, it should be emphasised that these were acute studies in healthy volunteers with normal glycaemic control. Subtle meal-related differences were observed when the concentration v. time curves were reviewed. Notably, PAL2 was associated with a slower decline in glucose concentrations from 30 to 60 min and the glucose level did not drop below the baseline level. In contrast, both REF and PAL1 were associated with a more rapid decline and, by 60 min, glucose was fallen below the baseline level, albeit to recover during the REF but not during PAL1. This pattern has been linked to a rise in counter regulatory hormones and a deterioration in second-meal glycaemic control( Reference Ludwig 22 ).
The greater control of glucose homeostasis observed with PAL2 could be explained by a lower insulinogenic index and less of an increase in GIP response relative to the REF (although in full multivariable analysis, the effect of GIP was no longer apparent), and might be explained by the positive effect of GIP on β-cell function, as has been reported previously( Reference Harada, Hamasaki and Yamane 23 , Reference Yamane, Harada and Hamasaki 24 ).
A reduction in the insulinogenic index is, conventionally, taken to indicate the impairment of β-cell function; however, the return to basal glucose levels by 60 min following the commencement of PAL2 implies entirely normal glucose tolerance. Moreover, glucose concentrations with PAL2 did not show the exaggerated decline that was apparent with the REF and PAL1, which suggests that the initial insulin response to PAL2 was appropriate and that the responses to the REF and PAL1 were exaggerated. GLP-1 is understood to have a stronger incretin effect than GIP( Reference Kreymann, Williams and Ghatei 25 ), and a clear dose–response relationship between GLP-1 and β-cell response has been reported, although this has only been demonstrated in exogenous infusion studies( Reference Kjems, Holst and Vølund 26 ). PAL1 was associated with the highest GLP-1 response, which might have been expected to contribute to the somewhat higher, albeit non-significant, insulinogenic index and rate of decline in glucose levels, compared with PAL2 (despite PAL1 and PAL2 having considerably lower GIP responses than the REF). However, surprisingly, no relationship was apparent between the GLP-1 and meal insulin responses.
Gastrointestinal hormone responses
Secretion of PYY and GLP-1 from neuroendocrine L-cells in the intestine is stimulated by specific nutrients (dietary fat, protein and SCFA) acting on nutrient-sensing G protein-coupled receptors( Reference Gribble 27 ). This process is complex and not fully understood and involves an array of G protein-coupled receptors, as well as non-receptor-mediated effects, stretch receptors and sensory influences. The interactions between signals generated by food, digestion, enzyme secretion and secretions in the stomach, are all considered factors thought to link the nutrient density and digestibility of a meal to facilitate control of gastrointestinal transit and appetite in order to maximise the absorption of valuable nutrients. The initial GLP-1 levels observed here suggest an early signalling of the presence of glucose in the stomach, as proposed by Schirra et al. ( Reference Schirra, Katschinski and Weidmann 28 ), and then a second response for both PAL meals suggesting the presence of glucose still in the lumen of the lower gut, possibly due to reduced bioavailability, before reaching the lower part of the duodenum. By contrast, the REF shows an initial surge, but no secondary response, suggesting that the majority of glucose has been removed from the gut by the time the meal reaches this area. This low response is probably due, at least in part, to the comparatively small meal size and carbohydrate load( Reference Schirra, Katschinski and Weidmann 28 ), although it is interesting to note that PAL2 is similar for both energy and carbohydrate load and still elicits a secondary GLP-1 response and one that is overall greater than the REF. This suggests that GLP-1 induction in mixed meals is controlled by more than the size of glucose load, as has recently been suggested( Reference Rijkelijkhuizen, McQuarrie and Girman 29 ). GLP-1 response is informed by protein and fat load as well as carbohydrate( Reference Elliott, Morgan and Tredger 30 – Reference Mansour, Hesseini and Larijani 32 ), and this is confirmed by the higher PAL1 response over the other meals. However, the similar levels of both fat and protein across PAL2 and REF make this an unlikely explanation for the differences between these other two meals. While the greater volume and mass of the PAL meals may have played a role in EVAS responses generated here, this alone seems unlikely to be responsible for the concomitant rise in satiety hormones. Studies on PYY and gastric volume alone have shown no link between volume and response( Reference Oesch, Ruegg and Fischer 33 ), while other studies have also shown that the PYY response is linked directly to nutrient content( Reference Adrian, Ferri and Bacarese-Hamilton 34 ). Despite the reported link between the high PYY response and protein levels, the data for the low- and high-protein PAL meals follow the same response up until the last time point.
The dramatically reduced GIP levels observed for both PAL meals are particularly intriguing, and may, in part, be explained by the available carbohydrate constituting a higher level of fructose in the PAL meals( Reference Vozzo, Baker and Wittert 35 ). However, previous work on mixed meals with free fructose suggest that the presence of both fructose and glucose would have caused only a slightly lower peak in the GIP response( Reference Teff, Elliot and Tschöp 36 ), rather than the very delayed response that is observed here. GIP has also been shown to be induced by lipid as well as glucose, although studies on protein vary( Reference Elliott, Morgan and Tredger 30 ). However, again, while total fat was higher in PAL1, this cannot account for the striking difference observed between PAL2 and REF, which had similar fat levels.
The overall data suggest that some aspect of the PAL meals renders glucose less accessible, which is the mechanism by which GIP is induced( Reference Ebert and Creutzfeldt 37 ); however, this can only be hypothesised as the glucose measurements cannot distinguish here between glucose absorption and uptake. While delayed gastric emptying alone could explain the delayed GIP response for the PAL meals, the later raised GLP-1 response suggests the delivery of unabsorbed glucose to the lower duodenum where there is higher L cell and lower K cell abundance. The authors speculate that a possible explanation for this could be flavonoids in the PAL meals, inhibiting glucose absorption in the jejunum( Reference Alvarado 38 , Reference Dao, Waget and Klopp 39 ), as delayed gastric emptying alone would be expected to give a GLP-1 response that was even further delayed than that observed here for GIP. However, it seems unlikely that delayed response alone can account for the magnitude of the differences in both GLP-1 and PYY responses observed between PAL2 and REF. This hypothesis would allow for the high GLP-1 level, in that glucose would be transported to the lower part of the duodenum unabsorbed where the presence of glucose in the lumen would induce the higher levels of GLP-1 observed here. This would also be supported by the higher levels of phenolic compounds extracted from PAL1 and PAL2 when compared with the REF (57·2, 86·1 and 4·47 mg GAE, respectively). As the extraction process for this did not use lipophilic extraction solvents and acid hydrolysis, which remove cell wall-bound hydroxycinnamic acids, and used Amberchrom column chromatography, these phenolics would be expected to be largely flavonoids.
Recent studies have shown a link between high-energy density foods and long-term PYY levels( Reference Hill, Rolls and Roe 40 ), but none (to our knowledge) have shown an overall long-term pattern of response for GLP-1 and GIP levels. The higher volume of the meals from ingredients as opposed to added water or other artificial means of increasing volume would suggest that it is volume coupled with nutrient content, which is then released as the food passes through the gut, that is responsible for the hormone responses observed here. It is tempting to speculate that the rate of gastric flow may play a role in delayed delivery, as both PYY and GLP-1 have been implicated in the ileal brake; however, this needs confirmation and is an area we plan to pursue in further studies.
Of the remaining hormones measured in the present study (pancreatic polypeptide, amylin, ghrelin and leptin), while pancreatic polypeptide showed a increased response for both PAL meals, none of the other hormones showed any consistent significant differences for the AUC between the three meals (see Table 4).
There is no known single lifestyle intervention that can ameliorate obesity and T2D risk factors. However, bariatric surgery has been shown to result in lowering of GIP with a concomitant rise in PYY and in some cases also GLP-1, as discussed above, and these factors are thought to play a role in insulin sensitivity and appetite regulation( Reference Le Roux, Aylwin and Batterham 41 , Reference Korner, Besslerb and Inabnetb 42 ). A similar reduction in GIP and rise in PYY, along with a related improved insulin sensitivity, has also been found as a result of lifestyle change studies( Reference Kelly, Brooks and Solomon 43 , Reference Solomon, Haus and Kelly 44 ).
Independently of their role in insulin and glucose control, there are also emerging data showing that both GIP and GLP-1 can play a role in other negative physiological responses to diet, such as fatty liver and obesity( Reference Yabe and Seino 45 ). The reduced GIP response, in particular, would justify further study in the light of data on knockout mice, which suggest that this hormone has a role to play in how the body metabolises and stores fat( Reference Seino and Yabe 46 ). However, further studies on the longer-term effects of this would be needed to link GIP response to longer-term weight loss in human subjects.
Appetite response
The satiety scores for the three test meals were significantly improved across all the three questions for both PAL meals (Table 4; Fig. 3), and yet, despite the higher energy and protein content of PAL1, there was no significant difference between these two meals until the final time point recorded. In fact, satiety quotient per unit energy( Reference Green, Delargy and Joanes 47 ) for all the three EVAS questions at 175 min shows that PAL2 gives the greatest level of satiety per kJ. Both PAL meals also showed higher levels of PYY, which has been shown to play a key role in appetite and energy intake reduction( Reference Batterham, Heffron and Kapoor 48 ), although levels for both PAL meals are very similar, contradictory to reports that this hormone is energetically governed( Reference Batterham, Cowley and Small 49 ). GLP-1 affects the ileal brake, resulting in slower gastric emptying( Reference Schirra, Katschinski and Weidmann 28 , Reference Wishart, Horowitz and Morris 50 ), and this could also have been a factor in the higher satiety ratings recorded for both PAL meals. A major similarity of the two PAL meals is the overall lower energy density caused by the minimal processing and cooking of the plant matter, causing the PAL meals to have a higher integral water content, although total dry weights of the matched PAL2 and REF were almost identical (HFJ Bligh and KJ Hunter, unpublished results). While the high volume and weight of the two PAL meals are likely to be a contributing factor to the initial EVAS scores observed here, it seems unlikely that this alone can account for the high and sustained level of PYY and GLP-1, which are observed to map the EVAS scores. However, this does suggest that the low energy density of the Palaeolithic-type meals from the high plant content is a contributory factor in the recorded satiety in the present and other studies, rather than just high protein, as has been suggested previously( Reference Mellberg, Sandberg and Ryberg 8 , Reference Jönsson, Granfeldt and Erlanson-Albertsson 9 , Reference Cordain, Eaton and Miller 51 ).
It is interesting to note that GLP-1 agonists are currently being explored as treatments for both diabetes and obesity, and the present study gives some indication that the levels of both GLP-1 and PYY could also be controlled nutritionally. The higher fibre content of the PAL meals, combined with the fact that this fibre would still be complex with the plant cell structure, would provide another contributor to the satiety and appetite scores. However, it is difficult to see how this alone can account for the incretin responses observed here, as the presence of fibre in some studies appears to suppress GLP-1( Reference Karhunen, Juvonen and Huotari 31 , Reference Mansour, Hesseini and Larijani 32 ) and increased fibre alone does not necessarily lead to enhanced satiety( Reference Barr and Wright 52 ). It should also be noted that Jönsson et al. ( Reference Jönsson, Granfeldt and Erlanson-Albertsson 9 ) appeared to also discount fibre as the sole explanation for their satiety data. There has also been some suggestion that phenolic content can also play a role in appetite response, and this may also have been a factor given the high phenolic load of the PAL meals (Table 1)( Reference Trigueros, Peña and Ugidos 53 ). By contrast, many modern processed foods are not only energy dense, but rapidly and easily digested, and thus do not maintain satiation and satiety for prolonged periods( Reference Prentice and Jebb 54 ).
Critical evaluation of the study
The EVAS score data were not the primary objective of the present study, and, as such, no ad libitum meal was undertaken. Further studies are currently in progress, and these will include ad libitum meals as part of the further investigation into both the satiety and anorectic hormone responses observed here.
The lack of palatability may have played a role in the EVAS scores recorded; however, this may well have been exacerbated by the time of day the meals were consumed, which was necessitated by the need for an overnight fast before consumption. While palatability recorded in the actual study was low, earlier tasting sessions did not raise this as an issue and while palatability probably affected early scores, it seems unlikely that this would have been sustained throughout the period studied or affected the hormone responses observed here.
There was no attempt to tailor the energy content of the meal to the BMI of the subjects, which may have played some role in individual responses. However, due to the complexity of ingredients of the PAL meals and the need to prepare these in advance to reduce batch variation of raw materials, it was not possible to energy match these to the energy requirements of the participants.
Overall conclusion
A preliminary acute-effects study is consistent with the view that rationally designed Palaeolithic-type meals based on modern foods show potential for delivering benefits in the reduced risk of obesity and even T2D. The findings presented here suggest that this may be mediated through the actions of PYY and GLP-1. Relevant features in meal design, which may be responsible for these effects, include low energy density, high fibre and high polyphenol content. Longer-term studies are needed to confirm this and elucidate the mechanisms of action.
Acknowledgements
The authors thank Sonia Pombo and colleagues at Leatherhead for conducting the study, Joan Lane for guidance on protocols and ethics, and Professors M. Thomas and M. Richards for helpful discussion. The authors also thank Professor Peter Wilde for critical reviewing of the manuscript
The present study was fully funded by Unilever.
The authors' responsibilities are as follows: J. C., M. J. B., H. F. J. B. and K. J. H. were involved in formulating the original research question; H. F. J. B., K. J. H., M. J. B., P. M., J. C. and T. P. J. M. worked on the design of the study; T. P. J. M. and H. F. J. B. were involved in the practical aspects of the study including meal design and preparation; H. F. J. B., K. J. H., D. H. and D. C. S. T. performed the venous glucose and hormone analysis; P. M. performed the statistical analysis and created the graphs; H. F. J. B., K. J. H., G. F., I. F. G., M. J. B. and K. M. were involved in the interpretation of the study; I. F. G. and G. F. suggested and performed the insulin modelling; H. F. J. B., K. J. H., G. F., I. F. G., J. C. M. J. B. and P. M. were involved in the drafting and reviewing of the manuscript. All authors read and approved the final manuscript.
All authors with the exception of G. F. and I. F. G. are employed by Unilever. G. F. is a consultant for Unilever.








 Open access
Open access