


INTRODUCTION
The Climate Change Act 2008 (DECC 2009b ) has established a long-term framework to tackle climate change in the UK by setting a target to reduce greenhouse gas (GHG) emissions by at least 80% from 1990 levels by 2050 (DECC 2009b ). The Act also requires Government to set carbon budgets, which are limits on GHG emissions in the UK for consecutive 5-year periods. The first three carbon budgets were set in 2009, following advice from the independent Committee on Climate Change. The Fourth Carbon Budget, which sets the limit on emissions for the 5-year period from 2023 to 2027 was announced on 17 May 2011 (Harvey & Stratton Reference Harvey and Stratton2011). This new target aims to reduce carbon emissions by 50% from 1990 levels, which means that emissions should not exceed 1950 million tonnes of carbon dioxide equivalent (CO2-e). This will put the UK on course to cut emissions by 80% by 2050. Agriculture accounts for 0·06 of all UK GHG emissions and 0·37 of the UK's agricultural emissions are derived from rumen enteric emissions. Of the annual c. 20 Mt CO2-e of agricultural methane (CH4) emissions in the UK, sheep account for c. 4 Mt, dairy cattle c. 5 Mt and beef/veal c. 8 Mt CO2-e (CCC 2010).
Methane, a GHG, is estimated to contribute about 0·24 of anthropogenic global warming, second only to carbon dioxide (Houghton Reference Houghton1997). Rumen methanogenesis, which results in the loss of up to 0·12 of gross energy intake (Johnson et al. Reference Johnson, Hill, Ward, Johnson, Branine, Carmean, Lodman and Khalil1993), is largely responsible for the production of CH4 from ruminants. Sheep CH4 production of c. 20 g/head/day varies according to the diet and is highest when the energy density of the diet is c. 10·5 MJ/kg dry matter (DM) and diet digestibility is c. 0·70 (Pelchen & Peters Reference Pelchen and Peters1998). The development of CH4 mitigation strategies, without causing negative impacts on production, is a major challenge for ruminant nutritionists and geneticists (McAllister et al. Reference Mcallister, Okine, Mathison and Cheng1996; Cottle et al. Reference Cottle, Nolan and Wiedemann2011). The impacts that such strategies might have on flock or herd profitability and efficiency have now become topical in the field of animal science. While CH4 emissions are reported to be greater on improved pasture with higher stocking rates, the additional farm profit gained can exceed the potential cost of additional emissions (Alcock & Hegarty Reference Alcock and Hegarty2006).
An alternative approach to nutritional treatment is to achieve small, cumulative decreases in CH4 production through genetic selection. Between-sheep phenotypic variations in CH4 emissions have been observed in respiration chambers (Blaxter & Clapperton Reference Blaxter and Clapperton1965) and under grazing conditions (Lassey et al. Reference Lassey, Ulyatt, Martin, Walker and Shelton1997; Ulyatt et al. Reference Ulyatt, Baker, Mccrabb and Lassey1999; Pinares-Patino et al. Reference Pinares-Patino, Ulyatt, Lassey, Barry and Holmes2003), where c. 0·85 of the variation in daily CH4 production from sheep grazing temperate pastures was due to variation between animals.
The UK is not close to having a carbon price introduced that would perhaps fit neatly alongside other economic values (EVs) to determine the relative importance of sheep traits related to CH4 emissions in breeding programmes within farm systems. ‘Shadow’ prices of carbon (SPC) of £19–50/t CO2-e have been proposed by Price et al. (Reference Price, Thornton and Nelson2007). A SPC of £19 in 2000 was approximately in line with the price required to reach (globally) a 450–500 ppm CO2-e stabilization scenario. The approach to carbon valuation by Government has recently undergone a major review that concluded in July 2009 before the Copenhagen meeting (DECC 2009a ). The new approach moves away from a valuation based on the damages associated with impacts (i.e. the SPC), instead using it as a basis for the cost of mitigation. The new approach sets the valuation of carbon at a level that is consistent with the UK Government's targets in the short and long term. In December 2008, the EU Climate and Energy Package agreed an aim of achieving 20% reductions in carbon emissions (on 1990 levels) across the EU by 2020, i.e. 0·67% reduction per annum. This equates to c. 0·1 kg CH4/head/yr for sheep if the reduction was achieved solely through breeding. The central price for carbon for those emissions not covered by the EU Emissions trading scheme, such as enteric CH 4, for evaluating the impact of policies are currently set at £51·70/t CO2-e, peaking at £307·78/t CO2-e in 2077 (DECC 2009a ).
There is a philosophical problem when including carbon price in breeding programmes by allocating it an appropriate EV. In the UK, carbon may be dealt with by cross-compliance for land-based issues (i.e. by imposing a financial penalty for failure to comply with environmental regulations through reducing the Single Farm Payment that farmers receive as part of the Common Agricultural Policy) or by grant (capital or annual maintenance) for compliance (e.g. blocking drains on hill land – based on calculations including a carbon value). This could have an impact on sheep production by, for example, giving a grant to buy rams with a positive estimated breeding value (EBV) for CH4 as a carbon-based trait – which then provides a means of receiving revenue in the sheep marketplace. It is unlikely that farmers will be paid for some improvement in their flock/farm carbon footprint for decades – a mechanism that could better fit into the classical EV selection index approach.
The question remains: how do farmers align a shadow price for anything (carbon, biodiversity benefit, etc.) for which there is intrinsic, but not economic, value against traits with clearer financial values, such as kilograms of lamb? Such valuation of ‘non-market’ values has already been discussed in the context of animal breeding by Olesen et al. (Reference Olesen, Navrud and Kolstad2006) and is worthy of further investigation in this context.
There has been some recognition of the opportunity for reducing CH4 emissions by genetic improvement but few papers on appropriate breeding objective(s) in any livestock species or breed. Breeding objectives should be economically based and the direct costs of CH4 explicitly included in the objective (Cottle et al. Reference Cottle, Van Der Werf and Banks2009, Reference Cottle, Nolan and Wiedemann2011; Hegarty & McEwan Reference Hegarty and Mcewan2010). Similarly, the CH4 emissions metric should be expressed on a basis that is independent of other traits to aid breeder interpretation.
The approach in the current study of hill sheep breeding was to include CH4 with other breeding objective traits and we have a similar investigation underway with terminal sire (meat) sheep. The change in kilograms of CH4 can be compared with changes in selection index response (£) and predicted response in other traits to study internal farm prices for carbon when it currently has no intrinsic value or payment.
The current paper addresses the questions, ‘what are the likely impacts of current breeding programmes in hill sheep farming systems on CH4 emissions’, and ‘what (incentive) price of carbon or other measures are needed to achieve likely government targets in CH4 reduction?’.
MATERIALS AND METHODS
Parameters
The selection index program, MTIndex (Cottle et al. Reference Cottle, Van Der Werf and Banks2009), was modified to construct indexes with the inclusion of CH4 production (kg/yr) as a trait. The genetic parameters and EVs for 10 breeding goals that combine lamb and maternal traits relevant to extensive sheep production in the UK were taken from previous studies (Conington et al. Reference Conington, Bishop, Grundy, Waterhouse and Simm2001, Reference Conington, Bishop, Waterhouse and Simm2004; Lambe et al. Reference Lambe, Bunger, Bishop, Simm and Conington2008) (Tables 1 and 2). Selection index traits not in the breeding objective (e.g. ultrasonic measurements of fat and lean) were implicitly given a zero EV. These traits are typically easier to measure and are indirect criteria for traits in the objective that are too difficult or costly to measure, i.e. fat and muscle as determined by computer tomography (CT). It was assumed that there was one record for all dam traits (Table 1) and one own record and 60 half sib records for weaning weight (WWT), fat and muscle depth, yearling faecal (nematode) egg count (FECn) and CH4 production, when CH4 was included as a selection criterion.
Table 1. Traits in the (modified) hill country index. Traits 1–5 are dam traits, traits 6–11 are lamb traits. Methane is both a lamb and dam trait

* NLW, number of lambs weaned; NLL, number of lamb losses; CT fat, CAT scan fat weight; CT lean, CAT scan lean weight; FECn, Faecal (nematode) log egg count at 12 months.
Table 2. Hill breed assumed correlations (phenotypic above, genetic below diagonal). Traits as described in Table 1

Additional traits included in the breeding objective were assigned genetic parameters from the literature or from the UK's Signet breeding service for FECn. These values are included in Tables 1 and 2. Correlations between CH4 and most production traits were not available in the literature. Their assumed mean correlations (Table 2) were set based on the published correlations of other traits with these production traits, where the other trait was known to be highly correlated to CH4. These values did not need changing after discussion with expert colleagues, as they seemed consistent and logical (Cottle et al. Reference Cottle, Van Der Werf and Banks2009). Genetic and phenotypic correlations between methane and any single trait were assumed to be equal. As the CH4 correlation values were unknown, the sensitivity of genetic gains to their variation was a major focus of the current study. The EV of CH4 production (kg/yr) was calculated as £/t CO2-e×21/1000, where 21 is the internationally accepted global warming potential of CH4 in CO2-e (Price et al. Reference Price, Thornton and Nelson2007). The EV of number of lambs weaned (NLW) and number of lambs lost (NLL) included an allowance for assumed carbon price and the predicted lamb offspring CH4 production, which was assumed to be 0·25 of an adult's emissions (allowing for slaughter lamb's having a lower feed intake and excess lambs being sold at 6 months of age).
The EVs were converted to economic weights for a 100-ewe flock, taking into account gene flows and discounting, by the methods described by Conington et al. (Reference Conington, Bishop, Waterhouse and Simm2004). The EV of methane was increased in 100 increments of £5·40/t CO2-e between zero and £535/t CO2-e (c. 25 times the SPC price of £27·00 in 2010 or c. 10 times the central, non-trading sector price of £51·70 in 2010) for most of the program runs. In some runs, the EV of methane was allowed to increase to £1340 (c. 50 times the SPC price) to achieve specific desired gains.
Index calculations
The variance of the selection index (I) was calculated in MTIndex as var(I) (σ I )=var(b′X)=b′var(X)b=b′Pb, where b is the vector of index weights placed on the phenotypic values of traits, X is the array of phenotypic values and P is the phenotypic variance–covariance matrix among traits. The variance of the breeding objective (H) (‘true breeding values’) was calculated as var(H)= σ H =var(v′g)=v′var(g)v=v′Cv, where v is the vector of breeding objective trait economic weights, g is the vector of genotypes or EBVs and C=var(g), the genetic variance–covariance matrix among traits.
The covariance between the index and the breeding objective (cov(I,H)) was calculated as cov(b′X′,v′g)=b′ cov(X,g)v=b ′Gv=b′Pb, so was equal to the variance of the index. The accuracy of the selection index was calculated as the correlation between predicted and true breeding values (r IH )=cov(I,H)/σ I σH =σ I 2/σ I σH =σ I /σ H =SD_Index/SD_Breed_Objective, where σ I is the standard deviation of the index and σ H is the standard deviation of the aggregate genotype. With high accuracies, the standard deviation of the index is almost equal to the standard deviation of the breeding goal.
With selection using a multiple trait selection index I, the average index value of selected parents is calculated as i×σ I =i×r IH ×σ H . The average value of progeny is expected to equal the average index value of parents, i.e. response per unit of selection intensity (R)=i×σ I =i×r IH ×σ H .
Response to each trait was determined by the regression of each trait on the index, so the response to trait j was calculated as
noting that
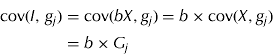
where G j is the jth column of the G matrix. After substituting Eqn (2) into Eqn (1), the response to trait j can be calculated as being equal to i×b′G j /σ I . Responses were converted to genetic change per year by multiplying R by the selection intensity and the inverse of the generation interval. These were calculated from the flock structure, which assumed five age groups of ewes and two age groups of rams mated first at 18 months, with a mating ratio of 50 ewes: 1 ram, a 130% weaning rate and 10% mortality rate from birth to age of selection. The weaning and single lamb mortality rates were based on average figures obtained from 20 years of performance data collected from a hill farm in Scotland. Generation length was thus calculated as 2·92 years and intensity of selection as 1·69.
The sensitivities of these calculated trait annual genetic gains to the assumed CH4: production correlations, which were used in index construction, were studied by randomly sampling 100 times within each of the normal distributions of these correlation values with an assumed standard error of 0·1 for genetic correlations and 0·05 for phenotypic correlations, using a Box & Muller (Reference Box and Muller1958) transformation Z 1= √(−2lnU 1)·sin(2πU 2), where Z 1 is an independent random variable with a normal distribution of standard deviation 1 and U 1 and U 2 are random numbers between 0 and 1. The Z 1 variable was multiplied by the assumed standard error of correlations to generate the normal distributions of correlation values with their assumed standard errors, which generated the covariances and variances used in the selection index calculations as described above. Repeat sampling of independent events allowed simple estimates of the average, as opposed to using a Bayesian approach via Gibbs sampling where each sample is conditional on previous samples (e.g. Ghavi Hossein-Zadeh & Ardalan Reference Ghavi Hossein-Zadeh and Ardalan2010). The assumed distribution of methane correlations was thus considered realistic and the best available until experimentally derived estimates become available. The program had also been modified to allow trait heritabilities to vary in a normal distribution. However, unlike methane correlations with other traits, estimates of CH4 heritability were available (0·3: Pinares-Patino et al. Reference Pinares-Patino, Mcewan, Dodds, Cárdenas, Hegarty, Koolaard and Clark2011; 0·15: P. Vercoe, personal communication) so values of 0·25 or 0·15 were used. The responses were not very sensitive to the choice of 0·15 or 0·25 for CH4 heritability, so only the results derived from an assumed CH4 heritability of 0·25 are presented in the current paper.
As the relative economic value (REV) of carbon in CH4 was increased in discrete increments from its chosen minimum to maximum value, the correlations used in constructing the multi-trait index (from which the trait responses to selection were calculated) were allowed to vary randomly within their expected distributions, as described above. This allowed the sensitivity of the predicted responses to variance in correlations to be assessed from the resultant response graphs (Figs 1–4). The more scattered the points on the graph, the more sensitive the responses to the size of the standard errors of the correlations.

Fig. 1. Changes, without measuring methane directly, in (a) overall index value with carbon payment (SD index) and without carbon payment (SD index-methane) and (b) methane, to changes in carbon price. Flock size of 100 head.

Fig. 2. Cumulative flock methane emissions with different selection indexes. Assumes a starting methane production of 15·3 t CO2-e/100 ewe flock/yr (7·3 kg methane/head). Key to selection index construction. Desired gain of zero for methane – emissions remain static at 7·3 kg/head/yr. All traits zero EV-indirect – all traits except methane given an EV of zero (i.e. maximum reduction in methane possible with assumed methane genetic parameters) with methane not directly measured, i.e. gain made by selection via correlated traits. All traits zero EV-direct – all traits except methane given an EV of zero (i.e. maximum reduction in methane possible) with methane measured directly. Zero EV-indirect – methane given an EV of zero with methane not measured directly. Zero EV-direct – methane given an EV of zero with methane measured directly. A 20% reduction in 30 years – government policy related target of −0·1 kg methane/head/yr resulting from a methane EV set at −£296 with methane measured directly, or −£1396 when methane is not measured directly.

Fig. 3. Methane response predicted for a wide range of carbon prices when methane is not measured directly.

Fig. 4. Changes, when methane is measured directly, in (a) index responses with carbon payment (SD index) and without carbon payment (SD index-methane), (b) methane, to changes in carbon price. Flock size of 100 head.
The index was run unconstrained with a desired methane gain of zero, or with −0·1 kg/head/yr in a 100-ewe flock to calculate the implicit carbon price needed for these reductions and their effects on other traits. This reduction of −0·1 kg CH4/head/yr results in a cumulative methane reduction of 20% over 30 years assuming a starting value for CH4 production of 7·3 kg/head/yr (Cottle et al. Reference Cottle, Van Der Werf and Banks2009).
RESULTS
Methane not included as a selection index trait (i.e. CH4 not measured)
The annual genetic responses to index selection calculated with changes in carbon price without measuring methane directly are shown in Fig. 1 for total index and CH4. Corresponding graphs for predicted annual responses for mature size, CT FAT, CT LEAN, NLW, NLL, WWT, longevity and FECn in relation to carbon price are shown in Appendix 1, Fig. A1a–h, respectively.
Figure 1 a shows that at very high carbon prices, relative financial gain is calculated to rise with increasing carbon price after financial gain reaches a minimum around the point of zero predicted CH4 gain, which occurs at a carbon price of £271·65/t CO2-e. This rise in financial returns with a high carbon price is due to the lower financial returns from lower numbers of lambs weaned, lower dams’ lambs’ average WWT and lower WWT being outweighed by the higher financial responses for lower mature size, lower CT fat, higher CT lean and lower CH4 production. The main financial impact is via the lower CH4 production. If the financial response from lower CH4 is removed (if a payment was not actually received or a penalty not imposed) the financial response from other trait responses to changing carbon prices declines (Fig. 1 b). This decline represents a loss of income from genetic gain in production traits due to the effects of a carbon penalty on reduced selection pressure on these traits.
With zero CH4 EV, the current situation in the UK, it is expected that a flock of 100 hill sheep would be genetically increasing CH4 production by c. 0·3 t CO2-e/yr. This is a relatively modest amount but it is in the opposite direction to that desired by government policy and the cumulative gain builds up (Fig. 2). The carbon price would need to be around £272/t CO2-e to result in a selection index for hill sheep that had zero gains predicted in CH4 production.
The index response is relatively linear, when CH4 is not directly measured, until carbon price rises above £500/t CO2-e when it becomes fairly static as carbon price increases (Fig. 3), if all production traits have the EVs assumed in the current paper. If all traits other than CH4 are given an EV of zero, which is unrealistic, CH4 can be reduced much more rapidly to the detriment of other traits, with a 20% reduction in accumulated CH4 emissions being reached in 13 years, if CH4 is measured, or 26 years if it is not measured directly (as shown in Fig. 2).
Methane measured and included as a selection trait
There is feed bin equipment available commercially that is able to measure CH4 emissions in cattle (S. Zimmerman, personal communication) and 2 h static chambers are being studied with sheep (Goopy et al. Reference Goopy, Hegarty, Robinson, Chilliard, Glasser, Faulconnier, Bocquier, Veissier and Doreau2009). If CH4 could be measured cost effectively and directly in individual sheep, a carbon price of £49·70/t CO2-e provides the incentive needed to keep CH4 production static, i.e. zero genetic change (shown in Fig. 4 a) via index selection. The predicted changes in CH4 in relation to carbon pricing when it is measured directly is shown in Fig. 4 b and the responses to the other goal traits in the index are shown in Appendix 1, Fig. A2 a–h.
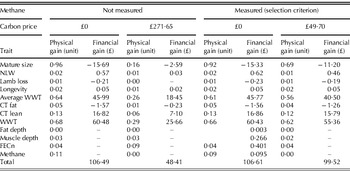
The predicted physical and financial gains when carbon price is zero or set at the value needed to achieve static CH4 emissions are shown in Table 3. The reduced production that is predicted to result from the carbon price for static CH4 emissions is less significant if CH4 is measured, as the carbon price needed to achieve static CH4 emissions is much lower than when CH4 is not measured directly.
Table 3. Hill sheep predicted gains (per year) in the breeding objective traits: mature size, NLW, lamb loss, longevity, average WWT, CT fat, CT lean, WWT, fat depth, muscle depth, FECn and methane with and without methane available as a selection criteria for a carbon price of zero, £49·70 or £271·65
DISCUSSION
Selection implications for production and profit
The UK government's ‘whole economy’ target reduction of 50% by 2027 can be achieved by hill sheep breeders via genetic selection with or without direct measurement of CH4, based on the assumed values for genetic parameters used in the current paper. However, such a gain would come at a large cost to the farmer in lost production in other traits and would need significant compensation to provide incentives for the required breeding programme changes and financial sustainability for the hill sheep breeding enterprises. The reduced progress in other traits as a direct result of including CH4 in the breeding programme arises mainly because of predicted improvements in lamb output due to improvements in the number of lambs reared, i.e. higher rates of lamb survival. The predicted increase in CH4 from hill sheep (3 kg CO2-e/ewe/yr), when using current selection indices that have implicit zero carbon prices, needs to be balanced against the predicted improvements in the number of lambs surviving to weaning, i.e. improved animal welfare, that is expected when using current selection indexes that have an implicit zero carbon price.
Balancing welfare benefits of having higher lamb survival v. increases in CH4 output can be undertaken using economic considerations if appropriate weightings of the societal benefits from having better welfare are included in the economic comparisons and a carbon price is used to place a value on the higher CH4 emissions. The topic of valuing traits with a societal value, such as animal welfare is discussed by Olesen et al. (Reference Olesen, Gjerde and Groen1999, Reference Olesen, Navrud and Kolstad2006). Using research market methods such as contingent valuation and conjoint analyses, a value can be placed on traits with no clear market EV by rating consumer preferences based on their willingness to pay for goods with perceived benefits in animal welfare or environmental enrichment. Such methodology has been placed in a breeding goal context for livestock by Nielsen et al. (Reference Nielsen, Christensen and Groen2005, Reference Nielsen, Christensen and Ødegard2006, Reference Nielsen, Amer and Olesen2008) and Nielsen & Amer (Reference Nielsen and Amer2007), and is discussed further in an environmental breeding goal context by Wall et al. (Reference Wall, Simm and Moran2010). The use of such methodology could well inflate the carbon price so that traits affected by the price rank higher in relation to other traits in the breeding goal. However, the expected responses in CH4 output rely heavily on the assumed correlations between CH4 production and other traits in the breeding goal. These assumptions are critical to the predicted rates of response in all traits. To deal with such uncertainty, a sensitivity analysis was carried out, using correlation estimates with a standard error of 0·1. However, when standard errors were altered to 0·5 (i.e. less certainty in assumed parameters), the results (not reported) could not be readily interpreted because the points were scattered very widely on the response graphs with very low R 2 values for the linear relationship between trait gains and carbon price. Thus, the results from the current study were sensitive to the assumed correlations, which are to date poorly documented. Further research is needed to estimate the genetic parameters associated with CH4 production to better predict the outcomes of including CH4 in breeding programmes.
When CH4 was not measured, an index resulting in a zero CH4 gain would cost the farmer c. £60/100 ewe flock in lost genetic gain per year in other traits. As genetic gain is cumulative, this loss increases non-linearly in total value with time. For example, over a 10-year period a loss of £60 per annum in genetic gain is equal to a cumulative, non-discounted loss of £3300 (£60×55) or a 5% discounted loss of £2851. Over a longer 30-year period this increases to a cumulative, non-discounted loss of £29 760 or a 5% discounted loss of £19 049. Such a penalty to farmers indicates that many farmers could struggle financially if a government policy of static change in CH4 production in sheep flocks were to be adopted. Farmers who performance record their livestock and use an index to achieve zero CH4 change at maximum profit in a high carbon price scenario would be purposely breeding smaller, less productive (per head) animals with lower feed intake and methane emissions. This would be counter-intuitive to many breeders, whose pride in improving their animals is an important component of their job satisfaction as well as income. It is unlikely that the sheep industry would broadly support the adoption of such a breeding strategy, even if the carbon financial penalties were compensated for by some form of government subsidy. It is also unlikely that any of the UK devolved Governments would support any scheme that would lead to reduced productivity. This is particularly the case in Scotland, following a recent report that since 2004, both the rate of labour productivity (output per agricultural worker) and land (output per ha) have been declining (Barnes et al. Reference Barnes, Bevan and Revoredo-Giha2011). It should be noted that indices have been calculated that would maximize the EV resulting from sheep breeding, if carbon was given different non-zero prices in future. This should not be construed as suggesting that the breeding of less profitable sheep is the means to reduce GHG emissions. It does, however, suggest that farmers would probably be adversely affected by carbon prices.
The current results suggest that CH4 production per head has probably been increasing in hill sheep flocks using the UK ‘Hill 2’ breeding index, where CH4 and feed intake have not been used as selection criteria. Due to the assumed correlations between CH4 and other traits, it is anticipated that even if CH4 was measured, CH4 would still increase at a similar rate (c. 3 kg/head/yr) as it is likely that CH4 is highly correlated with the main traits that are predicted to improve with index selection. Further work is already under way at Scottish Agricultural College (SAC) to obtain estimations of these correlations to improve understanding of the impacts of various selection indexes. The robustness of the results depends on how close the assumed CH4 correlations are to actual correlations when experimental estimates of them become available.
A more effective approach could be to select animals with a high feed efficiency (EBV weight gain:EBV feed intake) that are not necessarily smaller and less productive per head. That is, reduce CH4 indirectly via the lower feed intake per kg meat produced rather than reducing CH4 directly. Index modelling by Cottle et al. (Reference Cottle, Nolan and Wiedemann2011) suggests that this indirect approach would work in cattle breeding. There is evidence that there is genetic variation for emissions intensity (CH4 per unit of feed eaten) in sheep (Hegarty et al. Reference Hegarty, Alcock, Robinson, Goopy and Vercoe2010) and cattle (Arthur et al. Reference Arthur, Donoghue, Herd and Hegarty2009). The Sheep Cooperative Research Centre research programme (http://www.sheepcrc.org.au/; verified 17 November 2011) includes searching for cheaper methods to measure CH4, recognizing that direct measurement of methane is difficult (Robinson et al. Reference Robinson, Bickell, Toovey, Revell and Vercoe2011). Feed intake was not studied in the current paper because only traits that are included in current UK hill sheep selection indexes (with the exception of CH4) were included in the analyses. Feed intake has been included as a selection criterion trait in an associated study of UK terminal sheep breeds that is in preparation. A method is under development (patent pending: PCT/AU2010/001054) to facilitate the estimation of pasture feed intake in grazing animals and, if successful, this would make selecting directly for feed efficiency and indirectly for reduced CH4 production at pasture a practical possibility.
Wall et al. (Reference Wall, Simm and Moran2010) also concluded that a target reduction in GHG emissions could be met through improving the efficiency of production, as demonstrated by the 21% difference in CH4 output that already exists between the selection (high genetic merit) and control (average genetic merit) lines of cows at SAC's Langhill dairy herd. Wall et al. (Reference Wall, Simm and Moran2010) predicted that the rate of CH4 reduction could be further accelerated in dairy cows using direct measurements of CH4 production, if such measurements were included as breeding goals and given appropriate weightings in multiple trait, environmentally based selection indices.
Further studies
The current results do not take into account the cost of measuring CH4 or changes in livestock numbers that graze a set land area as a result of changes in traits such as NLW. To incorporate such measurement costs and changes in stock numbers resulting from selection to calculate flock profit requires more sophisticated modelling. Conington et al. (Reference Conington, Bishop, Waterhouse and Simm2004) quantified the economic impact of having fewer replacement females due to a higher flock productive lifespan within different land constraint parameters, as a result of improving ewe longevity. They found that in addition to having lower replacement costs, veterinary and medicine costs were also reduced. Additional benefits were also seen for grass growth due to a reduction in the demand for grazing, which would be cumulative over time. The costs of improving longevity were marginally higher feed costs per ewe (from keeping a mature ewe v. a replacement female), higher feed costs from having higher average litter size (due to having an older age structure) and a lower wool clip value from mature v. replacement female fleece. Such detailed modelling would be better justified and more credible, when CH4 genetic parameters are better defined in future, in particular the correlations between CH4 production and traits that are most important for efficient sheep production. Most of these correlations are unknown for both sheep and cattle, so selection index modelling results need to be treated with appropriate caution and a sensitivity approach, as reported in the current paper, is best used.
Non-breeding options
A greater impact on reduction of GHG emissions from hill farms may be obtained from changes made in other parts of the farming system, such as pasture management. When a broader perspective than the individual flock is being considered, life cycle analysis (LCA) of emissions can be carried out (Peters et al. Reference Peters, Rowley, Wiedemann, Tucker, Short and Schulz2010). Such an analysis should set the scene more broadly in terms of which farm management activities have the greatest impact on reducing GHG emissions and should also include consideration of levels of soil and pasture carbon sequestration. These LCA studies will enable more comprehensive comparisons between different farm systems to be made, including breeding programmes. As hill flocks generally have limited opportunities to improve or renew pastures, or to use alternative crops such as brassicas to finish lambs for slaughter, the options available for hill sheep breeders to reduce their carbon footprint are largely confined to reducing wastage (e.g. improving lamb survival through reducing lamb losses) and improving the efficiency of production (e.g. improving kg of lamb sold per kg of ewe kept on the farm through management and genetic selection). The current paper is a first attempt at providing information on possible impacts of including CH4 in UK hill sheep selection indices and first estimates of the carbon prices probably needed to stimulate a change in farmer practice.
The authors acknowledge the advice and assistance of Professor Julius van der Werf.
APPENDIX 1

Fig. A1 Changes in traits and overall index value to changes in carbon price when methane is not measured (a) mature size, (b) CT fat, (c) CT lean, (d) NLW, (e) NLL, (f) WWT, (g) longevity and (h) FECn.

Fig. A2 Changes in traits and overall index value to changes in carbon price when methane is measured directly (a) mature size, (b) CT fat, (c) CT lean, (d) NLW, (e) NLL, (f) WWT, (g) longevity and (h) FECn.









