


Introduction
Habitability may be quantitatively defined as a measure of the ability of a planet to develop and sustain life (Schulze-Makuch et al. Reference Schulze-Makuch, Méndez, Fairén, von Paris, Turse, Boyer, Davila, António, Catling and Irwin2011); its maximum is set as 1 for a planet where life as we know it has formed, thus it is 1 for the Earth. The requirement for a planet to be called habitable (or potentially habitable)Footnote 1 is that the planet is located within the host's habitable zone (HZ) and has terrestrial characteristics: rocky, with a mass range of 0.1–10 Earth masses and a radius range of 0.5– 2 Earth radiiFootnote 2 . A HZ is conservatively defined as a region where a planet can support liquid water on the surface (Huang Reference Huang1959). The concept of an HZ is, however, a constantly evolving one, and many different variations of it have been since suggested (see, e.g., an excellent review by Lammer et al. (Reference Lammer2009) and references therein and Heller & Armstrong (Reference Heller and Armstrong2014) as a more recent one). Biogenic elements (such as C, H, N, O, P and S) have also been considered as necessary complementary factors for habitability (Chyba & Hand Reference Chyba and Hand2005), but their presence is implied by the existence of water as they are produced in the same stars (Heger & Woosley Reference Heger and Woosley2002; Umeda & Nomoto Reference Umeda and Nomoto2005).
We would like to stress here that throughout the paper, when we talk about detecting life on exoplanets, we still mean life as we know it, the presence of which we are able to establish through predictable changes in planetary atmospheres. Even on Earth, there is a possibility of a different kind of life not based on a usual triad – DNA–protein–lipid; see, for example, the discussion on a ‘shadow’ biosphere by Davies et al. (Reference Davies, Benner, Cleland, Lineweaver, McKay and Wolfe-Simon2009). But just as on Earth we are not able to find it (yet) as we do not know ‘where or what to look for’, we may not be able to distinguish these different kinds of life from the natural environments of exoplanets. Hence, when we talk about biosignatures, we mean only biosignatures that our kind of life produces – oxygen, ozone, nitrous oxide, etc. (e.g., Seager et al. Reference Seager, Schrenk and Bains2012). A planet may host life as we know it (in other words, be not just habitable but inhabited), but we will still not detect it unless it has evolved sufficiently to change its environment on a planetary scale, for instance, through the production of an oxygen atmosphere by photosynthetic organisms. Photosynthesis is currently the only geologically documented biogenic process (see, e.g., Lyons & Reinhard, Reference Lyons and Reinhard2011; Fomina & Biel Reference Fomina, Biel, Allakhverdiev, Rubin and Shuvalov2014 and references therein) that can provide sufficient energy to modify the global planetary (or atmospheric) properties. The large free energy release per electron transfer and stability of the oxygen molecule due to its strong bonding ensures that an oxygen-rich atmosphere provides the largest feasible energy source for complex life (e.g., Catling et al. Reference Catling, Glein, Zahnle and McKay2005). Therefore, by analogy with the Earth, we presume the presence of an oxygen atmosphere as necessary for a planet to host a complex life. Such life would have modified the global planetary (or atmospheric) properties to be noticed from space, and from very far away; after all, the closest potentially habitable planet (PHP) is at about 12 light years (τ Ceti) and we cannot go there to verify. Even Mars might still be inhabited by a primitive subsurface biota which is undetectable without a local and detailed examination. It may also be possible for life to evolve in a manner that we have not anticipated, which, even if it changes the environment globally, would not be detectable simply because we are not looking for those particular changes. For example, aphotic life can exist in the subsurface oceans of Europa or Enceladus, but such life would be currently impossible for us to detect ex situ.
Biological methanogenesis was suggested as a rival to the photosynthesis process in changing the global environment and capable of enriching the exo-atmospheres with biogenic methane (Schindler & Kasting Reference Schindler and Kasting2000; Kharecha et al. Reference Kharecha, Kasting and Siefert2005). Kharecha et al. (Reference Kharecha, Kasting and Siefert2005) has shown that the rate of biogenic methanogenesis in the atmosphere of an Archaen Earth could have been high enough to enrich the atmosphere with high concentration of biogenic methane. However, planets with reduced mantles might enrich their atmospheres by methane abiotically (e.g., Etiope & Lollar Reference Etiope and Sherwood Lollar2013), and thus methane alone cannot guarantee habitability. From this point of view, methanogenic products are a less certain biosignature of Earth-like life than oxygen (Seager et al. Reference Seager, Schrenk and Bains2012). Accounting for a competitive interrelation between metabolic and abiotic origins of methane, a more conservative understanding suggests that only the simultaneous presence of methane along with other biogases is a reliable indication of life (e.g., Selsis et al. Reference Selsis, Despois and Parisot2002; Kaltenegger et al. Reference Kaltenegger, Traub and Jucks2007; Kiang et al. Reference Kiang, Segura, Tinetti, Blankenship, Cohen, Siefert, Crisp and Meadows2007; Kasting et al. Reference Kasting, Kopparapu, Ramirez and Harman2014). It could also be that the planet never develops oxygenic photosynthetic life. In such cases, other biomarkers have been suggested; for example, dimethyl disulphide and CH3Cl may be detected in infrared (IR) in the planetary atmospheres of low-ultraviolet (UV) output stars (Domagal-Goldman et al. Reference Domagal-Goldman, Meadows, Claire and Kasting2011).
Carter (Reference Carter1983) has pointed out that the timescale for the evolution of intelligence on the Earth (~5 Gyr) is comparable with the main sequence lifetime of the Sun (~10 Gyr). Lin et al. (Reference Lin, Gonzalez Abad and Loeb2014) suggested that intelligent life on expolanets can be detected through the pollution it inflicts on the atmosphere. However, intelligent life, once evolved, is no longer in need of a very precisely defined biosphere – we can already create our own biospheric habitats on planets that are lifeless in our definition of habitability, e.g. Moon or Mars, though we are technologically intelligent for only a 0.0000026% of the time life exists on Earth: 100 years out of 3.6 Gyr. Therefore, intelligent life may not be so easily detectable, especially if they have had a longer time to evolve. However, to answer the most important question of ‘are we alone’, we do not necessarily need to find intelligent life. Even detection of a primitive life will have a profound impact on our civilization. Therefore, we need to concentrate on the period in a planet's history when the emerged life had already influenced the atmosphere of the planet in a way that we can possibly recognize.
We discuss here the importance of the age of the planet in the evaluation of whether that HZ planet contains life and whether that life is detectable. We examine the plausibility of a discovery of a habitable planet with detectable biota among the close (within 600 pc) neighbours of the Sun. We argue that variations in their albedos, orbits, diameters and other crucial parameters make the formation of a significant oxygen atmosphere take longer that the current planetary age and thus, life can be detectable on only half of the confirmed PHPs with a known age.
Initial stages of habitability
Necessary conditions for the developing of life are thought to include rocky surface and liquid water; however, the aspects connected with the stages preceding the onset of biological era are usually left out of consideration. Planetary age as a necessary condition for life to emerge was first stressed by Huang (Reference Huang1959) and implicitly mentioned by Crick & Orgel (Reference Crick and Orgel1973) in their concept of a Directed Panspermia.
In order to understand the importance of planetary age for the evolution of a detectable biosphere we will consider, as an example, the development of cyanobacteria and the related atmospheric oxidation (Irwin et al. Reference Irwin, Méndez, Fairén and Schulze-Makuch2014). This process involves several endothermic reactions and requires sufficiently high temperatures to be activated. In general, the temperature dependence of the photosynthetic rate is rather complicated and conditionally sensitive, with the effective activation energy being of the order of tens of kJ mol−1 (Hikosaka et al. Reference Hikosaka, Ishikawa, Borjigidai, Muller and Onoda2006), much higher than the typical equilibrium temperature on habitable planets. Thus small variations in atmospheric and crust properties can considerably inhibit photosynthesis and increase the growth time of the mass of cyanobacteria. This conclusion may be illustrated through the consideration of the elementary process of carboxylation of RuBP (ribulose-1,5-bisphosphate: C5H12C11P2) in the dark Benson–Bassham–Calvin cycle of photosynthesis (Benson et al. Reference Benson, Bassham, Calvin, Goudate, Haas and Stepka1950; Farquhar et al. Reference Farquhar, von Caemmerer and Berry1980). These photosynthetic reactions, controlled by enzymes, are known to be very sensitive to ambient temperatures with an optimum rate at about 40°C, and a practically zero rate outside the temperature range of 0° < t < 60°C (Toole & Toole Reference Toole and Toole1997). Amongst other fundamental factors RuBP carboxylation is probably the most relevant one, determining the optimal temperature of photosynthesis, and is characterized by the activation energy V C ≃ 30 − 60 kJ mol−1 at the growth temperature (Hikosaka et al. Reference Hikosaka, Ishikawa, Borjigidai, Muller and Onoda2006). We can roughly characterize the RuBP carboxylation by the Arrhenius law
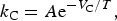 $${k_{\rm C}} = A{{\rm e}^{ - {V_{\rm C}}/T}},$$
$${k_{\rm C}} = A{{\rm e}^{ - {V_{\rm C}}/T}},$$
where k
C is the rate constant, A is the prefactor, and T is the absolute temperature. The characteristic time of RuBP carboxylation is
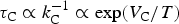 ${{\rm \tau} _{\rm C}} \propto k_{\rm C}^{ - 1} \propto \exp ({V_{\rm C}}/T)$
. Since the RuBP carboxylation is one of the main processes optimizing photosynthetic reactions, τC can roughly characterize the rate of photosynthesis on a planet.
${{\rm \tau} _{\rm C}} \propto k_{\rm C}^{ - 1} \propto \exp ({V_{\rm C}}/T)$
. Since the RuBP carboxylation is one of the main processes optimizing photosynthetic reactions, τC can roughly characterize the rate of photosynthesis on a planet.
The range of variation in τC on a habitable planet due to the uncertainty in the equilibrium temperature T e is
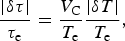 $$\displaystyle{{\vert{{\rm \delta} {\rm \tau}} \vert} \over {{{\rm \tau} _{\rm e}}}} = \displaystyle{{{V_{\rm C}}} \over {{T_{\rm e}}}}\displaystyle{{\vert{\rm \delta} T\vert} \over {{T_{\rm e}}}}, $$
$$\displaystyle{{\vert{{\rm \delta} {\rm \tau}} \vert} \over {{{\rm \tau} _{\rm e}}}} = \displaystyle{{{V_{\rm C}}} \over {{T_{\rm e}}}}\displaystyle{{\vert{\rm \delta} T\vert} \over {{T_{\rm e}}}}, $$
with τ e being a characteristic time of photosynthesis at T e. The equilibrium temperature T e, in turn, is calculated using planetary parameters inferred from the observations,
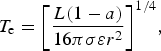 $${T_{\rm e}} = {\left[ {\displaystyle{{L(1 - a)} \over {16{{\rm \pi} {\rm \sigma} {\rm \varepsilon}} {r^2}}}} \right]^{1/4}}, $$
$${T_{\rm e}} = {\left[ {\displaystyle{{L(1 - a)} \over {16{{\rm \pi} {\rm \sigma} {\rm \varepsilon}} {r^2}}}} \right]^{1/4}}, $$
where the uncertainties in the parameters determine the uncertainty in its estimate,
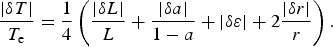 $$\displaystyle{{\vert{\rm \delta} T\vert} \over {{T_{\rm e}}}} = \displaystyle{1 \over 4}\left( {\displaystyle{{\vert{\rm \delta} L\vert} \over L} + \displaystyle{{\vert{\rm \delta} a\vert} \over {1 - a}} + \vert{{\rm \delta} {\rm \varepsilon}} \vert + 2\displaystyle{{\vert{\rm \delta} r\vert} \over r}} \right). $$
$$\displaystyle{{\vert{\rm \delta} T\vert} \over {{T_{\rm e}}}} = \displaystyle{1 \over 4}\left( {\displaystyle{{\vert{\rm \delta} L\vert} \over L} + \displaystyle{{\vert{\rm \delta} a\vert} \over {1 - a}} + \vert{{\rm \delta} {\rm \varepsilon}} \vert + 2\displaystyle{{\vert{\rm \delta} r\vert} \over r}} \right). $$
Here L is the luminosity of the central star, a and ε the planet's albedo and emissivity, r the orbital radius, and σ the Stephan–Boltzmann constant. It is readily seen that the actual time of the onset of photosynthesis for a given habitable planet might differ significantly from the value calculated from largely uncertain parameters that were, in turn, derived from observables. Indeed, uncertainties in estimates of the equilibrium temperature |δT|/T
e are heavily amplified for habitable planets with V
C/T
e ≃ 10 − 20 for V
C ≃ 30 − 60 kJ mol−1 and T
e ~ 300 K, such that even relatively low observational errors in deriving the parameters in equation (4), of 5% each, might result in 50−100% error in the estimates of the overall photosynthesis rate. If one considers oxidation of the Earth atmosphere as a process tracing the developing photosynthesis, the characteristic time for the growth of biota on early Earth can be estimated as the oxidation time,
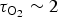 ${{\rm \tau} _{{\rm O}_2}} \sim 2$
Gyr (Kasting Reference Kasting1993; Wille et al.
Reference Wille, Kramers, Nägler, Beukes, Schröder, Meisel, Lacassie and Voegelin2007; Fomina & Biel Reference Fomina, Biel, Allakhverdiev, Rubin and Shuvalov2014). Therefore, a 50% error in τC may delay the possible onset of biological evolution on a planet by 1 Gyr, i.e. biogenesis might not start earlier than 3 Gyr from the planetary formation. In general, however, the problem of the photosynthetic process is much more complex, depending on many factors determined by thermal and non-thermal processes on a planet (Shizgal & Arkos Reference Shizgal and Arkos1996; Hikosaka et al.
Reference Hikosaka, Ishikawa, Borjigidai, Muller and Onoda2006), and might be even more sensitive to variations in physical conditions. Even on the early Earth, physical conditions could have been such as to preclude the onset of biogenesis over a long time (Sagan Reference Sagan1974; Maher & Stevenson Reference Maher and Stevenson1988; Solomatov Reference Solomatov and Canup2000).
${{\rm \tau} _{{\rm O}_2}} \sim 2$
Gyr (Kasting Reference Kasting1993; Wille et al.
Reference Wille, Kramers, Nägler, Beukes, Schröder, Meisel, Lacassie and Voegelin2007; Fomina & Biel Reference Fomina, Biel, Allakhverdiev, Rubin and Shuvalov2014). Therefore, a 50% error in τC may delay the possible onset of biological evolution on a planet by 1 Gyr, i.e. biogenesis might not start earlier than 3 Gyr from the planetary formation. In general, however, the problem of the photosynthetic process is much more complex, depending on many factors determined by thermal and non-thermal processes on a planet (Shizgal & Arkos Reference Shizgal and Arkos1996; Hikosaka et al.
Reference Hikosaka, Ishikawa, Borjigidai, Muller and Onoda2006), and might be even more sensitive to variations in physical conditions. Even on the early Earth, physical conditions could have been such as to preclude the onset of biogenesis over a long time (Sagan Reference Sagan1974; Maher & Stevenson Reference Maher and Stevenson1988; Solomatov Reference Solomatov and Canup2000).
From this point of view, the planetary habitability index (PHI) recently proposed by Schulze-Makuch et al. (Reference Schulze-Makuch, Méndez, Fairén, von Paris, Turse, Boyer, Davila, António, Catling and Irwin2011) in the form
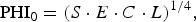 $${\rm PH}{{\rm I}_0} = {(S \cdot E \cdot C \cdot L)^{1/4}},$$
$${\rm PH}{{\rm I}_0} = {(S \cdot E \cdot C \cdot L)^{1/4}},$$
can be generalized with explicit inclusion of the age of the planet t as
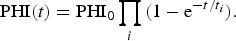 $${\rm PHI}(t) = {\rm PH}{{\rm I}_0}\prod\limits_i {(1 - {{\rm e}^{ - t/{t_i}}})}. $$
$${\rm PHI}(t) = {\rm PH}{{\rm I}_0}\prod\limits_i {(1 - {{\rm e}^{ - t/{t_i}}})}. $$
In equation (5), S defines a stable substrate, E the necessary energy supply, C the polymeric chemistry and L the liquid medium; all the variables here are in general vectors, while the corresponding scalars represent the norms of these vectors. In equation (6), the index i denotes a chemical chain relevant for further biochemical evolution, and t i is its characteristic time. It is obvious that the asymptotic behaviour – approaching the maximum habitability – is controlled by the slowest process with the longest t i .
Other factors delaying the onset of habitability
Sagan (Reference Sagan1974) was the first to stress that harmful endogenous and exogeneous processes in the early Earth could postpone emergence of life on it. Such processes could be important even in the very initial primitive episodes of biogenesis and delay the formation of biota for up to billions of years. It is known from the 182W isotope dating that the late heavy bombardment of Earth, Moon and Mars lasted till about 3.8 Ga (Schoenberg et al. Reference Schoenberg, Kamber, Collerson and Moorbath2002; Moynier et al. Reference Moynier, Koeberl, Quitté and Telouk2009; Robbins & Hynek Reference Robbins and Hynek2012). The Martian primitive atmosphere is believed to have been lost through catastrophic impacts about 4 Ga (e.g., Melosh & Vickery Reference Melosh and Vickery1989, Webster et al. Reference Webster2013). Evidence of a heavy bombardment in other exoplanet systems exists: collision-induced hot dust was detected in several young planetary systems. Spectral signatures of warm water- and carbon-rich dust in the HZ of a young ~1.4 Ga MS star η Corvi (Lisse et al. Reference Lisse, Wyatt, Chen, Morlok, Watson, Manoj, Sheehan, Currie, Thebault and Sitko2012), and of host dust in seven sun-like stars (Wyatt et al. Reference Wyatt, Smith, Greaves, Beichman, Bryden and Lisse2007) indicate recent frequent catastrophic collisions between asteroids, planetesimals or even possible planets (Song et al. Reference Song, Zuckerman, Weinberger and Becklin2005). Out of these seven stars, five are young systems within their first Gyr of life.
It is also well-known that solar-type stars remain very active in the first billion years of their life, sustaining conditions that are hostile to the survival of the atmosphere and to the planetary habitability. G-type stars, within the first 100 Myr of reaching ZAMS, produce continuous flares of extreme-UV (EUV) radiation up to 100 times more intense than the present Sun, and have much denser and faster stellar winds with an average wind density of up to 1000 times higher. Low-mass K- and M-type stars remain X-ray and EUV-active longer than solar-type stars, where EUV emission can be up to 3–4 times and 10–100 times, respectively, higher than G-type stars of the same age; and active M-type stars could keep stellar winds in the HZ that are at least 10 times stronger than that of present Sun (France et al. Reference France2013).
In recent simulations by Schaefer and Sasselov (Reference Schaefer and Sasselov2015) of the development of oceans on super-Earths, it was shown that though these planets keep their oceans for longer than the Earth (up to 10 Gyr), it also takes longer for them to develop the surface ocean due to the delayed start of volcanic outgassing that returns water back to the surface from the mantle. For super-Earths 5 times the Earth's mass, that would take about a billion years longer. The oceans are believed to have established on Earth 750 Myr after formation, therefore super-Earths would have their surface water established at only 2 Gyr after formation. After all, the Great Oxygenation Event about 2.5 Ga (Anbar et al. Reference Anbar, Duan, Lyons, Arnold, Kendall, Creaser, Kaufman, Gordon, Scott, Garvin and Buick2007) was most likely induced by oceanic cyanobacteria, which allowed life to emerge on land about 480 − 360 Ma (Myr ago) (Kenrick and Crane, Reference Kenrick and Crane1997).
Potentially habitable planets
At the time of writing, more than 1900 exoplanets have been confirmed (Extrasolar Planets Encyclopaedia, June 2015) with another 4000 waiting for confirmation (NASA Exoplanet Archive). The majority of detected planets are in the vicinity of the Sun, and their hosts are mostly young Population I (Pop I) stars with ages of hundreds of Myr to a few Gyr. The age distribution of the host stars with measured ages is shown in Fig. 1. 58% of the host stars have ages of 4.5 Gyr and less, and more than one-third (~38%) are younger than 3 Gyr. Simple statistics shows the median age of ~3.8 Gyr.
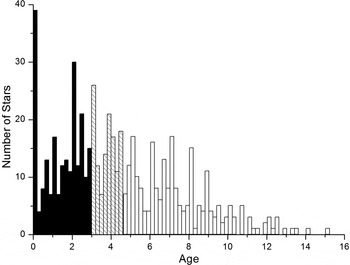
Fig. 1. Age distribution of the stars hosting confirmed planets (total 583 hosts with known ages at time of writing). We highlight the number of stars with ages below 3 Gyr in black, and between 3 and 4.5 Gyr as hatched. The predominance of young host stars is clearly seen, which could be the effect of observational selection (Shchekinov et al. Reference Shchekinov, Safonova and Murthy2013). This figure was made using the Extrasolar Planets Encyclopaedia data.
The fact that more than one-third of the planetary systems, discovered by ongoing exoplanetary missions, are younger than ~3 Gyr is not surprising, because the continuous star formation (SF) in the Galactic disk supplies young stars, and the fraction of hosts younger than 3 Gyr represents that very fraction of Pop I stars that would be born provided the SF rate is nearly constant during the whole period of the thin disk formation. Most of the current exoplanet missions suffer from an observational bias – they mostly detect systems that are younger than the age at which life is presumed to have appeared on the EarthFootnote 3 .
Incidentally, Fig. 1 shows a deficit of stars with ages t > 6 Gyr. Assuming that Pop I stars, i.e. the thin Galactic disc, have started forming at about 10 Ga (Chen et al. Reference Chen, Hou and Wang2003; Carraro et al. Reference Carraro, Geisler, Villanova, Frinchaboy and Majewski2007), one might expect the presence of such old stars in our vicinity in the proportion corresponding to the SF history in the early Galaxy. The most conservative assumption implies a constant SF rate, in which case one should expect the number of planet-hosting stars with ages t > 6 Gyr of about 40%. It is, however, believed that the SF was more active in the early epochs (Bouwens et al. Reference Bouwens, Illingworth, Franx and Ford2007), therefore, the fraction of hosts older than 6 Gyr should be correspondingly higher. The reason for the decline in the number of the hosts in this age range is unclear and might, in particular, indicate that planetary systems lose planets with age.
About 40 PHPs are currently documentedFootnote 4 , though extrapolation of Kepler's data shows that in our Galaxy alone there could be as many as 40 billion PHPs (Petigura et al. Reference Petigura, Howard and Marcy2013). In Table 1, we show the data for PHPs for which the host ages were available in the literature. The fraction of young planetary systems is nearly consistent with the age distribution of Pop I stars: among the 33 confirmed habitable planets with known ages more than half are ≲3.5 Gyr old.
Table 1. Host ages for confirmed PHPs

a The radii of these planets are >1.7 Earth's, however, it is still too soon to exclude them from the list, according to Torres et al. (Reference Torres2015), since there are many uncertainties in the modelling of the transition from rocky to hydrogen/helium planets, and these planets may be rocky.
References to ages: 1. The Extrasolar Planet Encyclopaedia (http://exoplanet.eu); 2. Angalada-Escudé et al. (Reference Anglada-Escudé2012); 3. Anglada-Escudé et al. (Reference Anglada-Escudé2013); 4. Borucki et al. (Reference Borucki2013); 5. Anglada-Escudé et al. (Reference Anglada-Escudé2014); 6. Mamajek & Hillenbrand (Reference Mamajek and Hillenbrand2008); 7. Tuomi & Anglada-Escudé (Reference Tuomi and Anglada-Escudé2013); 8. Open Exoplanet Catalogue (http://www.openexoplanetcatalogue.com); 9. Nordström et al. (Reference Nordström, Mayor, Andersen, Holmberg, Pont, Jørgensen, Olsen, Udry and Mowlavi2004); 10. Tuomi et al. (Reference Tuomi, Anglada-Escudé, Gerlach, Jones, Reiners, Rivera, Vogt and Butler2013a); 11. Pepe et al. (Reference Pepe, Lovis, Ségransan, Benz, Bouchy, Dumusque, Mayor, Queloz, Santos and Udry2011); 12. Metcalfe (Reference Metcalfe2013); 13. Wittenmyer et al. (Reference Wittenmyer2014); 14. Torres et al. (Reference Torres2015); 15. NASA Exoplanet Archive at http://exoplanetarchive.ipac.caltech.edu; 16. Gaidos (Reference Gaidos2013); 17. Crossfield et al. (Reference Crossfield2015); 18. Tuomi et al. (Reference Tuomi2013b).
It seems reasonable to update the definitions in footnote 1 on page 1 as
-
1. PHP – a rocky, terrestrial-size planet in an HZ of a star.
-
2. Habitable planet – a rocky, terrestrial-size planet in an HZ with detected surface water and some of the biogenic gases in atmosphere.
-
3. Inhabited planet – the best case scenario: a rocky, terrestrial-size planet in an HZ with simultaneous detection of species such as water, ozone, oxygen, nitrous oxide or methane in atmosphere, as proposed by e.g. Sagan et al. (Reference Sagan, Thompson, Carlson, Gurnett and Hord1993) or Selsis et al. (Reference Selsis, Despois and Parisot2002).
We may expect only a primitive form of biota on the youngest planets (≲2 Gyr) in Table 1, which would not be detectable. Biogenesis could have started, or even progressed to more advanced stages with an oxidized atmosphere, on older planets with ages from 2 to 4 Gyr. In the former case, one can expect that methane from metabolic reactions has already filled the atmosphere, while in the latter case, oxygen molecules at some level can appear in the atmosphere – though atmospheric oxygen on Earth appeared about 2.5 Ga, the Earth itself became visibly habitable only about 750–600 Ma, when the biosphere became active and complex enough to modify the environment to be noticed from space (e.g., Méndez et al. Reference Méndez, González, Jimenez, Pérez and Bracero2013). The traces of these gases may, in principle, be observed in sub-mm and micron wavelengths, provided the planets are orbiting low-mass stars (0.5–0.8 M ⊙). Even if one-third of the low-mass stars in the sky host planets (Tutukov & Fedorova Reference Tutukov and Fedorova2012), there may be as many as a thousand planets within a 10 pc vicinity with ages ranging from Myr to a few Gyr.
The age of a planet is of primary importance for developing the future strategy of looking for life on PHPs. Since space programs are extremely expensive and require extensive valuable telescope time, it is crucial to know in advance which planets are more likely to host detectable life. Young planets will not have atmospheres abundant in products of photosynthetic processes, and many planets, though residing in the HZ, may not actually be habitable for life as we know it. For example, the host stars in the Degenerate Objects around Degenerate Objects (DODO) direct imaging search for sub-solar mass objects around white dwarfs (Hogan et al. Reference Hogan, Burleigh and Clarke2009) are rather young with an average age of only 2.25 Gyr. The target star selection of the Darwin (ESA) mission is restricted to stars within 10 − 25 pc (Kaltenegger & Fridlund Reference Kaltenegger and Fridlund2005), and two space missions that are currently under study, the NASA Transiting Exoplanet Survey Satellite (TESS) mission and ESA's PLAnetary Transits and Oscillations of stars (PLATO) mission, will only survey bright F, G, K stars and M stars within 50 pc (e.g., Lammer et al. Reference Lammer2013), sampling therefore only the thin Galactic disc stars – young Pop I hosts. The main focus of Exoplanet Characterization Observatory (EChO) (Drossart et al. Reference Drossart2013) is the observation of hot Jupiter and hot Neptune planets, limited due to the mission lifetime constraints to bright nearby M stars (Tinetti et al. Reference Tinetti2012). Most known habitable planets cannot have an existing complex biosphere although they may develop it in the future, because most currently known PHPs are found around relatively young Pop I stars. We feel that it is reasonable to fix a period of ~4 Gyr as the minimum necessary time for the formation of complex life forms under optimal conditions, as evidenced by the Earth's biosphere. Direct observations of planetary atmospheres in IR and sub-mm wavebands would be a promising method for tracing biogenesis. Planned future IR and sub-mm observatories could provide such observations (see the discussion in Section ‘Observational prospects’ below.).
In this context, we have undertaken the project of updating the catalogue of Nearby Habitable Systems (HabCat) constructed for SETI by Turnbull & Tarter (Reference Turnbull and Tarter2003a) for the search for potentially habitable hosts for complex life. A complete characterization of all the stars within a few hundred (or even a few tens of) parsecs, including their masses, ages, and whether they have planetary systems (including terrestrial planets), was not realizable at that time. Our aim was to find out the information on these stars: their ages and whether they have planets and if they could be potentially habitable.
To begin with, we have taken the HabCat II, a ‘Near 100’ subset – a list of the nearest 100 star systems of the original HabCat (Turnbull & Tarter Reference Turnbull and Tarter2003b), as a basis for our project. These stars were scrutinized for information on their age, nearby planets etc., which were missing in the original catalogue but are important now due to their impact on selecting the targets for future space missions. Out of 100 nearby (within 10 pc) objects in the HabCat II, we have found the age data for 50 stars. This list is being cross-correlated with the Hypatia Catalogue, which is a project to find abundances for 50 elements, specifically bio-essential elements, for the stars in the HabCat (Hinkel et al. Reference Hinkel, Timmes, Young, Pagano and Turnbull2014). Our goal is to compile a list of the most probable planets that may allow future missions to search our neighbourhood for habitable/inhabited planets more efficiently. The preliminary result of this project is presented in Table A.2 in Appendix B.
Old planetary systems
General census
Most of the old planetary systems were discovered serendipitously. Only in 2009 were targeted surveys of metal-poor stars initiated (Setiawan et al. Reference Setiawan, Klement, Henning, Rix, Rochau, Rodmann and Schulze-Hartung2010). In spite of that, quite a few old (≳9 Gyr) planetary systems are currently known. Shchekinov et al. (Reference Shchekinov, Safonova and Murthy2013) attempted to compile a list of such system (see their Table 1) on the basis of metallicity, considering stars [Fe/H] ≤ −0.6. They, however, missed many previously known systems with ages determined by several different methods, including metallicity abundances, chromospheric activity, rotation and isochrones. Combining their table with other studies (Saffe et al. Reference Saffe, Gómez and Chavero2005; Haywood Reference Haywood2008 and latest updates of online exoplanet catalogues) brings the census of planetary systems with ages ≳9 Gyr to 116 planets (90 host stars; see Appendix A for the table of these systems). It is possible that the number of such hosts is much larger since we have counted only those stars where estimates from different methods were comparable. For example, in the list of NASA Exoplanet Archive candidates to PHPs, out of 62 hosts with estimated ages, 28 are older than 10 Gyr.
The majority of old planets were detected by the radial velocity method which is biased to detect preferentially massive planets due to a limited sensitivity. The continuously increasing precision of radial-velocity surveys may in future change this picture, and the first example of that is the detection in mid-2014 of the terrestrial planet (~5 Earth masses) orbiting extremely old (10–12 Gyr) Kapteyn's star (Anglada-Escudé et al. Reference Anglada-Escudé2014). The most remarkable thing is that this planet lies in the HZ. The star also has another super-Earth outside the HZ.
Potential habitability of old planetary systems
The improved precision has also resulted in the rejection of three previously reported old planets HIP 13044 b and HIP 11952 b, c (e.g., Setiawan et al. Reference Setiawan, Klement, Henning, Rix, Rochau, Rodmann and Schulze-Hartung2010, Reference Setiawan, Roccatagliata, Fedele, Henning, Pasquali, Rodríguez-Ledesma, Caffau, Seemann and Klement2012) as a genuine signal (Jones & Jenkins Reference Jones and Jenkins2014). However, it still leaves the number of old planets of at least 117 (92 hosts, see Table A.1 in the Appendix A) with 11 super-Earths (namely, Kepler-18 b; 55 Cnc e; Kapteyn's b, c; MOA-2007-BLG-192L b, OGLE-2005-BLG-390L b and five planets of Kepler-444) and all the rest gas giants, which do not fall into the category of habitable planets. However, because giant planets typically harbour multiple moons, the moons may be habitable and may even lie in the domain of a higher habitability, or even ‘superhabitability’ (Heller & Armstrong Reference Heller and Armstrong2014). For example, Schulze-Makuch et al. (Reference Schulze-Makuch, Méndez, Fairén, von Paris, Turse, Boyer, Davila, António, Catling and Irwin2011) estimate the PHI for Jupiter to be only 0.4, while it is around 0.65 for Titan. There are 33 potentially habitable exomoons with habitable surfaces listed by HEC (excluding possibility of subsurface life), which have on average ESI higher than the PHPs. Heller et al. (Reference Heller2014) have shown that the number of moons in the stellar HZ may even outnumber planets in these circumstellar zones, and that massive exomoons are potentially detectable with current technology (Heller Reference Heller2014). Even though Population II (Pop II) stars are normally two order of magnitude less abundant in metals, they may harbour up to ten potentially habitable rocky Earth-size subsolar objects each (Shchekinov et al. Reference Shchekinov, Safonova and Murthy2013), either as planets or as moons orbiting gaseous giants. Planets can form at metallicities as low as Z ~ 0.01Z ⊙ due to the centrifugal accumulation of dust (Shchekinov et al. Reference Shchekinov, Safonova and Murthy2013). However, Pop II stars could have formed in the metal-enriched pockets resulting from a non-perfect mixing in young galaxies when the Universe was as young as a few hundreds of Myr (Dedikov & Shchekinov Reference Dedikov and Shchekinov2004; Vasiliev et al. Reference Vasiliev, Dedikov and Shchekinov2009). They would be able to form planets in a traditional way, and our Galaxy may have a vast number of rocky planets residing in habitable zones. Such planets had longer time for developing biogenesis. Recently discovered five rocky planets orbiting 11.2 Gyr old star Kepler-444 (Campante et al. Reference Campante2015) seems to confirm the previously suggested (Shchekinov et al. Reference Shchekinov, Safonova and Murthy2013) hypothesis.
Direct measurements of metallicities and abundance pattern in the early Universe have recently become possible with the discovery of extremely metal-poor (EMP) stars with metallicities as low as 10−5 of the solar value – these objects are believed to represent the population next after the Population III (Pop III) stars (Beers & Christlieb Reference Beers and Christlieb2005). The relative abundances observed in the EMP stars are shown to stem from the explosions of Pop III intermediate-mass SNe with an enhanced explosion energy about 5 × 1051 erg (Umeda & Nomoto Reference Umeda and Nomoto2005). These stars are also often found to be overabundant in CNO elements. Interestingly, their relative abundance (Aoki et al. Reference Aoki2006; Ito et al. Reference Ito, Aoki, Beers, Tominaga, Honda and Carollo2013) is consistent with the abundance pattern of the Earth crust (Taylor & McLennan Reference Taylor and McLennan1995; Yanagi Reference Yanagi2011) and the chemical composition of the human body (see, e.g., Nielsen Reference Nielsen, Fischer, L'Abbé, Cockell and Gibson1997).
Though Earth is rich in chemistry, living organisms use just a few of the available elements: C, N, O, H, P and S, in biological macromolecules: proteins, lipids and DNA, which can constitute up to 98% of an organisms’ mass (e.g., Alberts et al. Reference Alberts2002). Apart from hydrogen, these ‘biogenic’ elements are all produced by the very first massive Pop III stars. Detection of substantial amount of CO and water in the spectrum of z = 6.149 quasar SDSS J1148 + 5251 shows, for example, that at ~800 Myr after the Big Bang, all the ingredients for our carbon-based life were already present. The initial episode of metal enrichment is believed to have occurred when the Universe was about 500–700 Ma – the absorption spectra of high-redshift galaxies and quasars show significant amount of metals, in some cases up to 0.3 of the solar metallicity (e.g., Savaglio Reference Savaglio2006; Finkelstein et al. Reference Finkelstein2013). The abundance pattern of heavy elements in this initial enrichment contains a copious amount of elements sufficient for rocky planets to form within the whole range of masses (Bromm et al. Reference Bromm, Coppi and Larson1999; Abel et al. Reference Abel, Bryan and Norman2000; Clark et al. Reference Clark, Glover, Smith, Greif, Klessen and Bromm2011; Stacy et al. Reference Stacy, Bromm and Loeb2011).
Therefore, planets formed in the early Universe and observed now as orbiting very old (≳9 Gyr) Pop II stars, may have developed and sustained life over the epochs when our Solar System had only started to form. In this way, the restricted use of six ‘biogenic’ elements may be considered as a fossil record of an ancient life – it is well known that at the molecular level, living organisms are strongly conservative. The general direction of the biological evolution is in the increase of complexity of species rather than (chemical) diversity (Mani Reference Mani, Saviotti and Metcalfe1991). For example, paradoxically, both oxygen and water are destructive to all forms of carbon-based life (e.g., Bengston Reference Bengston1994). The presence of water reduces the chance of constructing nucleic acids and most other macromolecules (Schulze-Makuch & Irwin Reference Schulze-Makuch and Irwin2006). The toxic nature of oxygen necessitated the evolution of a complex respiratory metabolism, which again shows the strong chemical conservatism at the molecular level in that the living organisms developed the protection mechanisms to circumvent these problems rather than use other compounds.
Observational prospects
Recently, a 13.6 Gyr star was detected placing it as the oldest star in the Universe (SMSS J031300.36–670839.3, Keller et al. Reference Keller2014); the age was estimated by its metallicity [Fe/H] ≤ −7.41. In spite of that, this star, believed to have formed from the remnants of the first-generation SN, was found to contain carbon, metals such as lithium, magnesium, calcium, and even methylidyne (CH). It is quite possible that such stars have planets that are directly observable in micron wavelength range. Such EMP stars are known to have low masses and, as such, the orbiting planets could be seen directly in the IR.
The number of EMP stars is estimated to be about 250,000 within 500 pc in SDSS database (Aoki et al. Reference Aoki2006), so the mean distance between them is about 10 pc. If each EMP star hosts an Earth-size planet, the flux from the planet at a distance d in the IR range (λ ~ 10 µm) evaluated at the peak frequency (Wien's law) ν T = αkT/h, is
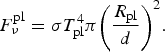 $$F_\nu ^{{\rm pl}} = {\rm \sigma} T_{{\rm pl}}^4 {\rm \pi} {\left( {\displaystyle{{{R_{{\rm pl}}}} \over d}} \right)^2}. $$
$$F_\nu ^{{\rm pl}} = {\rm \sigma} T_{{\rm pl}}^4 {\rm \pi} {\left( {\displaystyle{{{R_{{\rm pl}}}} \over d}} \right)^2}. $$
we can rewrite this flux as
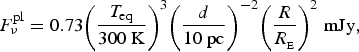 $$F_\nu ^{{\rm pl}} = 0.73{\left( {\displaystyle{{{T_{{\rm eq}}}} \over {300\;{\rm K}}}} \right)^3}{\left( {\displaystyle{d \over {10\;{\rm pc}}}} \right)^{ - 2}}{\left( {\displaystyle{R \over {{R_{_{\rm E}}}}}} \right)^2}\;{\rm mJy}, $$
$$F_\nu ^{{\rm pl}} = 0.73{\left( {\displaystyle{{{T_{{\rm eq}}}} \over {300\;{\rm K}}}} \right)^3}{\left( {\displaystyle{d \over {10\;{\rm pc}}}} \right)^{ - 2}}{\left( {\displaystyle{R \over {{R_{_{\rm E}}}}}} \right)^2}\;{\rm mJy}, $$
where T eq is the equilibrium temperature of a planet and R is its radius. For the Sun/Earth system, the ratio of the fluxes at a distance of 10 pc is
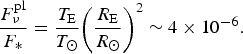 $$\displaystyle{{F_\nu ^{{\rm pl}}} \over {{F_{\ast}}}} = \displaystyle{{{T_{\rm E}}} \over {{T_ \odot}}} {\left( {\displaystyle{{{R_{\rm E}}} \over {{R_ \odot}}}} \right)^2} \sim 4 \times {10^{ - 6}}. $$
$$\displaystyle{{F_\nu ^{{\rm pl}}} \over {{F_{\ast}}}} = \displaystyle{{{T_{\rm E}}} \over {{T_ \odot}}} {\left( {\displaystyle{{{R_{\rm E}}} \over {{R_ \odot}}}} \right)^2} \sim 4 \times {10^{ - 6}}. $$
However, if we consider a super-Earth with M ~ 5M E, R ~ 2R E and T eq = 300 K, orbiting the star with T = 3000 K and R ~ 0.1 R ⊙ – an M dwarf, we get an improvement of
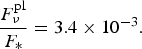 $$\displaystyle{{F_\nu ^{{\rm pl}}} \over {{F_{\ast}}}} = 3.4 \times {10^{ - 3}}. $$
$$\displaystyle{{F_\nu ^{{\rm pl}}} \over {{F_{\ast}}}} = 3.4 \times {10^{ - 3}}. $$
It seems challenging to detect such a weak contribution to a total flux from a planet even in the IR. There is, however, a possibility to distinguish the emission from the planet in IR molecular features, such as CH4 or O2, tracing either initiated biogenesis or developed metabolism. Detection of direct IR emission from O2 on exoplanets going through the initial epoch of biogenesis, or which are already at a stage with developed biota, was discussed in Churchill & Kasting (Reference Churchill and Kasting2000) and Rodler & Lopez-Morales (Reference Rodler and Lopez-Morales2014), respectively. Rich IR to sub-mm spectra of methane (Niederer Reference Niederer2012; Hilico et al. Reference Hilico, Loete and Champion1987) also allow to optimistically view the future detection of this biosignature. Even at the low temperatures of EMP stars, T * ~ 3000 K, these molecules are unlikely to survive in sufficient amount in their atmospheres. Therefore, if such emission is observed from an EMP star, it should be considered as a direct indication of an orbiting rocky planet that has already entered the habitable epoch with growing PHI (equation (6)). The most promising way to identify habitable (inhabited) planets seems to look for simultaneous presence of water, O2, O3, CH4 and N2O in atmospheric spectra (e.g., Selsis et al. Reference Selsis, Despois and Parisot2002; Kaltenegger et al. Reference Kaltenegger, Traub and Jucks2007; Kiang et al. Reference Kiang, Segura, Tinetti, Blankenship, Cohen, Siefert, Crisp and Meadows2007). Though such observations can be used to detect planets with highly developed habitability orbiting old EMP stars, the expected fluxes in the IR are still below current sensitivity limits and might be only possible in the future. For example, the future Millimetron space observatory planned for launching in next decade (estimated launch 2025) will have the detection limit of 0.1 µJy in 1 h observation in 50–300 µm range (Kardashev et al. Reference Kardashev2014). A molecular CH4 absorptions at ~50 µm can be detected by Millimetron in 3 h observations (Equation (10)) if a nearby (within 10 pc) habitable super-Earth planet transits an M-dwarf.
Summary
-
• Age of a planet is an essential attribute of habitability along with such other factors as liquid water (or an equivalent solvent), rocky mantle, appropriate temperature, extended atmosphere, and so forth. The knowledge of the age of a ‘habitable’ planet is an important factor in developing a strategy to search for complex (developed) life.
-
• Nearly half of the confirmed PHPs are young (with ages less than ~3.5 Gyr) and may not have had enough time for evolution of sufficiently complex life capable of changing its environment on a planetary scale;
-
• Planets do exist around old Pop II stars, and recently discovered EMP stars (belonging presumably to an intermediate Pop II.5) are good candidates for direct detection of orbiting planets in the IR and sub-mm wavelengths. Though currently only very few such PHPs are known, old giant planets may have habitable worlds in the form of orbiting moons.
-
• IR and sub-mm observations of terrestrial planets orbiting low-mass old stars are a promising way to trace biogenetic evolution on exoplanets in the solar neighbourhood.
Acknowledgements
Y. S. acknowledges the hospitality of RRI and IIA, Bangalore, when this work has been initiated. The authors thank Tarun Deep Saini for his useful comments, IIA Ph. D. student A. G. Sreejith for help with graphics and IIA internship student Anuj Jaiswal for his contribution in the project of updating the HabCat. The authors also thank the referees for their valuable comments which led to considerable improvement in the paper. This research has made use of the Extrasolar Planets Encyclopaedia at http://www.exoplanet.eu, Exoplanets Data Explorer at http://exoplanets.org, NASA Exoplanet Archive at http://exoplanetarchive.ipac.caltech.edu and NASA Astrophysics Data System Abstract Service.
Appendix A. The list of stars with estimated ages ≥9 Gyr
Table A.1. Stars with measured/estimated masses of ≥9 Gyr

Appendix B. The ‘Near 100’ – a subset from the nearest 100 star systems
Table A.2. Selection from the ‘Near 100’ of the stars with estimated ages





