



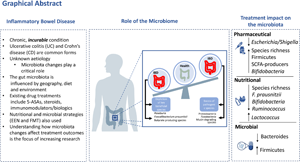
Inflammatory bowel disease (IBD) is comprised of several similar yet clinically distinct entities. The vast majority can be characterised as ulcerative colitis (UC) or Crohn's disease (CD)(Reference Halfvarson, Brislawn and Lamendella1). A small number, approximately 10 % of cases have features of both and are classified as ‘undifferentiated IBD’, which is more common amongst children(Reference Tremaine2). The intestinal inflammation which occurs in IBD results in typical symptoms of abdominal pain, diarrhoea and passage of blood or mucus per-rectally. Sufferers generally will experience attacks or ‘flares’ of disease activity which are interspersed between the periods of relatively symptom-free remission. If untreated, these can culminate in the loss of intestinal function, resulting in complications such as malnutrition.
CD is characterised by inflammation that can involve the entire thickness of the gastrointestinal tract, so-called transmural inflammation (Table 1) and may involve the entire gastrointestinal tract from mouth to the perianal area. Inflammation is discontinuous, resulting in ‘skip lesions’ where active disease is interspersed within the patches of normal-appearing bowel. The most commonly affected area is the ileum and proximal colon. The transmural inflammation can give rise to fistulous tracts that traverse through the bowel wall, sometimes into adjacent bowel or organs. This can manifest in intestinal perforation which requires surgical intervention. The intestinal lumen can also become narrowed due to inflammation or the formation of fibrotic strictures, resulting in bowel obstruction. Acute complications of CD require surgery in 6–16 % of cases, with up to 40 % of patients requiring further resections within 10 years due to disease recurrence(Reference Strong, Koltun and Hyman3). Repeated resection can lead to short gut syndrome with profuse chronic diarrhoea and nutrient deficiencies(Reference Gardiner and Dasari4). One of the most debilitating aspects of CD is involvement of the perianal area by abscesses and fistulae, which are painful and often disfiguring.
Table 1. Comparison of UC and CD location, inflammation, presentation and treatment

In contrast, the inflammation which occurs in UC is limited to the mucosal layer of the colon (Table 1). UC invariably involves the rectum and extends in a proximal and continuous fashion to other regions of the colon. A potentially catastrophic variant of UC is acute severe disease which is defined by a set of clinical and laboratory parameters known as ‘Truelove and Witts Criteria’(Reference Chen, Andrews and Kariyawasam5). Acute severe ulcerative colitis affects 15–25 % of UC patients during their lifetime and carries a 50 % likelihood of requiring total colectomy within 3 years of acute severe ulcerative colitis diagnosis(Reference Dinesen, Walsh and Protic6). Chronic colonic inflammation can result in dysplasia which can progress to malignancy. This occurs to a greater degree in UC compared to CD. The cumulative risk of colorectal cancer in UC patients is 7⋅6 % after 30 years of disease(Reference Rutter, Saunders and Wilkinson7), although increasingly there is a recognition that risk is associated with chronically active disease as much as duration of disease(Reference Choi, Al Bakir and Ding8).
Both UC and CD are associated with pathology in other organ systems, these are known as extra-intestinal manifestations(Reference Lambert, Parnell and Han9). Musculoskeletal extra-intestinal manifestations include sacroiliitis, ankylosing spondylitis, peripheral arthropathy and osteoporosis. In the eye, episcleritis, scleritis and uveitis have been described. Cutaneous manifestations include erythema nodosum and pyoderma gangrenosum. Primary sclerosing cholangitis is the most common liver disease specific to IBD, affecting 4–5 % of patients, and is more commonly associated with UC(Reference Harbord, Annese and Vavricka10).
Role of the microbiome in inflammatory bowel disease
As with most immune-mediated diseases, IBD is considered an idiopathic condition, with no clear aetiologic agent(Reference Chang11). After decades of epidemiologic, genetic, laboratory and clinical studies, the complex interactions between factors influencing IBD pathogenesis are only beginning to be understood. Abundant evidence now suggests that a dysbiotic intestinal microbiota, characterised by an altered ratio of pro- to anti-inflammatory microbes plays a central role in initiating and perpetuating intestinal damage(Reference Yilmaz, Juillerat and Oyas12). Genetic influences including host genetic polymorphisms in a number of genes that are involved in microbial recognition and processing, have also been identified(Reference Jostins, Ripke and Weersma13,Reference Nasser, Bergman and Fulco14) . Environmental factors including lifestyle, diet and medications further affect the balance, often through their impact on intestinal microbiota composition(Reference Yilmaz, Juillerat and Oyas12,Reference Maier, Pruteanu and Kuhn15,Reference Savage16) . It is now generally accepted that IBD results from a ‘perfect storm’ of interactions between a dysbiotic microbiota, aberrant immune system and environmental exposures within a susceptible host (Fig. 1)(Reference Turner, Kumar and Abbas17).

Fig. 1. Interactions between the gut microbiota, host and environmental factors in the pathogenesis of inflammatory bowel disease (IBD). The gut microbiota, host and environmental factors modulate and affect each other to form a physiological balance. In IBD, this balance is disturbed e.g. dysbiosis of the gut microbiota which leads to an aberrant immune response. Adapted from Hold et al.(Reference Hold, Smith and Grange94). ATG16L1, autophagy related protein 16-like 1; IL-23R, interleukin-23 receptor; NOD2, nucleotide-binding oligomerisation domain-containing protein 2.
The human gut hosts approximately 1014 bacteria comprising up to 1000 different species, along with viruses, fungi and other microorganisms(Reference Ni, Wu and Albenberg18). The collective genome of these microbes is referred to as the microbiome, which contains 100-fold more genes than the entire human genome(Reference Dave, Higgins and Middha19). The intestinal microbiota is in continual contact with the host immune system, generating inflammatory responses to eliminate pathogens as well as promoting systemic tolerance to the collective microbiota(Reference Chang11,Reference Brandtzaeg20,Reference Franzosa, Sirota-Madi and Avila-Pacheco21) .
Evidence to demonstrate the role of the microbiota in IBD pathogenesis is multifaceted. Animal studies have played a significant role in driving forward our understanding of IBD pathogenesis, especially murine colitis models. Several elegant studies have demonstrated the absolute requirement of the gut microbiota in the development of spontaneous colitis in genetically susceptible animals(Reference Sellon, Tonkonogy and Schultz22–Reference Chiaro, Soto and Zac Stephens24). In the clinical arena, the effectiveness of antibiotics, such as rifaximin, in reducing intestinal inflammation have also been repeatedly demonstrated for some aspects of IBD management including in reducing recurrence of CD after surgical resection(Reference Ambrose, Allan and Keighley25). They are also the mainstay of treatment for inflammation of the ileal pouch after an IBD patient undergoes proctocolectomy(Reference Segal, Oke and Hold26). In patients with CD, diversion of the faecal stream proximally to the inflamed mucosa has also been shown to result in the reduction of inflammation and induction of healing in the excluded parts of the gut, whereas relapse occurs with the restoration of faecal stream and re-exposure to luminal contents(Reference Rutgeerts27,Reference D'Haens, Geboes and Peeters28) . However, whether dysbiosis is the cause, result, contributor or ‘innocent bystander’ of aberrant inflammation remains unclear and remains the topic of ongoing intense study.
The cyclical relapsing and remitting nature of IBD also fuelled the hope that a microbial culprit was waiting to be found; akin to the Helicobacter pylori:peptic ulcer story. Many organisms were proposed but those deemed to have been of most interest over the years include Mycobacterium paratuberculosis, Escherichia coli and various Helicobacter and Campylobacter species(Reference Mukhopadhya, Hansen and El-Omar29–Reference Kaakoush, Holmes and Octavia32). Studies however failed to definitively attribute cause/association of a single culprit as the cause of disease with recognition that the complexity of the lower intestinal microbiota meant it was equivalent to ‘looking for a needle in a haystack’. An important shift in recent times, facilitated by the advancement of next-generation sequencing techniques and culturomics, has been the move towards identification of all species within an ecosystem's microbiota. As a result, instead of focusing on a single species, the current research paradigm considers the entire microbial ecosystem as the potential culprit. This paradigm shift has allowed a picture of the IBD microbiota to emerge; which is typified by lower microbial diversity, altered microbial composition and microbial community instability compared to the non-IBD subjects.
Halfvarson et al. performed one of the pioneering longitudinal cohort studies evaluating intestinal microbiome composition of individuals with CD, UC and healthy controls(Reference Halfvarson, Brislawn and Lamendella1). Stool samples were collected every 3 months for up to 2 years and microbiome composition was determined by 16S rRNA analysis. In keeping with previous studies, microbial composition in controls and IBD formed distinct clusters, with ileal CD patients having the most divergent microbiota profiles compared to healthy controls. Unique insights emerged when patients were evaluated over time. First, although microbiome profiles from healthy controls varied over time, they did so within a relatively limit range. In contrast, IBD microbiomes fluctuated to a much greater extent, occasionally entering the ‘healthy’ zone but not generally remaining there. This observation emphasises that IBD is characterised by a volatility that is not found in the healthy gut. During the study period, seven IBD patients experienced a flare of their disease symptoms and received a course of oral corticosteroids. These patients had greater fluctuations than those on stable medication. However, as no additional samples were collected at the time of the flare, it remains unclear whether there was a specific microbial signature associated with increased disease activity.
Other longitudinal studies have explored how microbiota networks relate to long-term disease severity and responsiveness to different IBD treatments(Reference Yilmaz, Juillerat and Oyas12,Reference Franzosa, Sirota-Madi and Avila-Pacheco21,Reference Imhann, Vich Vila and Bonder33–Reference Pascal, Pozuelo and Borruel36) . As the number of these studies increases and the repertoire of multi-omics analysis tools expands, the complexity of the role of the microbiome in IBD is increasingly unravelling. Although changes in microbiota composition are seen based on geography, age and diet, the overarching microbiota changes in studies indicates a global reduction in microbial richness in IBD cohorts with clear separation of CD from healthy controls whilst UC is more heterogenous(Reference Costello, Lauber and Hamady37–Reference Zhou, Xu and He39). In addition, the metabolites, produced by microbes, are increasingly being shown to play a pivotal role in gut homoeostasis through alteration of signalling pathways, immune functioning as well as antimicrobial activity(Reference Dorrestein, Mazmanian and Knight40,Reference McHardy, Goudarzi and Tong41) . In both CD and UC, a greater level of dysbiosis is seen in patients experiencing a disease flare compared to patients in remission but consistent differences are not seen between inflamed/uninflamed tissue from the same patient indicating that microbial changes are a field change rather than simply due to the presence of inflammation.
Take home messages that have come out of multi-omics assessment of IBD patient cohorts include the need to explore wider patient sample sets including serum, stool, biopsies and urine. There is also a growing appreciation that analytical approaches may well need to differ between UC and CD. This is due to the current limited identification of UC-specific biomarkers specific taxa, metabolomic molecules and diagnostic biomarkers, compared to CD, than has been identified by current approaches. The other exciting prospect to come out of multi-omics analysis datasets is the ability to interrogate microbiota changes in response to treatment regimens. Multiple studies have demonstrated the impact of microbes on drug availability and treatment efficacy in many immune-mediated disorders(Reference Ananthakrishnan, Luo and Yajnik42–Reference Vetizou, Pitt and Daillere45). Given the central role of the gut microbiota in IBD pathogenesis, understanding the impacts of therapeutics on gut microbiota and conversely, how the microbiota is related to treatment outcomes is essential to evaluate the role of the gut microbiota as a predictive biomarker for treatment response.
Inflammatory bowel disease treatment options
IBD treatment aims to achieve disease remission and mucosal healing in addition to a reduction in symptoms(Reference Cross and Farraye46). A ‘step-up’ approach to treatment is often used which is based on escalation of drugs from those with a better safety profile and cost-effectiveness but lower efficacy such as 5-aminosalicylic acid (5-ASA) to those that are more potent but with greater risk of adverse effects such as corticosteroids, immunomodulators and biologic agents (Fig. 2)(Reference Aloi, Nuti and Stronati47). However, increasingly there is a recognition that risk is associated with chronically active disease as much as duration of disease(Reference Choi, Al Bakir and Ding8,Reference Aloi, Nuti and Stronati47) . Risk profiling of disease patterns is also used to accelerate step up therapy in patients who have high risk disease.

Fig. 2. ‘Step up’ approach to inflammatory bowel disease (IBD) therapeutics where there is escalation from safer and low-efficacy drugs, such as 5-aminosalicylic acid to more potent drugs such as corticosteroids, immunomodulators and biologic agents based on IBD severity. Adapted from Aloi et al.(Reference Aloi, Nuti and Stronati47). JAK, Janus kinase.
Therapeutic approaches to ulcerative colitis management
First-line UC treatments to induce remission are 5-ASA and/or corticosteroids. Patients who do not respond are considered at high-risk for colectomy and are ‘stepped up’ to more aggressive therapies involving immunomodulators or biologic agents such as anti- TNF-α agents, anti-integrin antibodies or Janus kinase inhibitor (Fig. 2). Anti-TNF-α agents are also used as both induction and maintenance therapy. These agents have been shown to alleviate symptoms, induce mucosal healing, reduce hospitalisations and colectomies. However, loss of response often as a consequence of immunogenicity affects about 30 % of patients within the first year of therapy(Reference D'Haens, Baert and van Assche48). Therefore, combination therapy with an immunomodulator is preferred given its ability to suppress antibody formation. Vedolizumab is the second-line agent for induction of remission in patients non-responsive to anti-TNF-α agents. It has good efficacy, safety profile and lower rates of immunogenicity(Reference Colombel, Sands and Rutgeerts49). Thus, it can often be used without immunomodulator agents.
Therapeutic approaches to Crohn's disease management
It is postulated that there is a short window of opportunity for the treatment of CD that can prevent irreversible bowel damage, hospitalisations, surgeries and disabilities(Reference Colombel, Narula and Peyrin-Biroulet50). On this basis, CD patients with clinical indicators for poor outcomes: younger age of onset, perianal disease or extensive anatomic involvement are treated most aggressively. Current evidence suggests that response/remission rates are higher if a biologic agent is given within 2 years of disease onset(Reference Lichtenstein, Hanauer and Sandborn51) and early use of biologics is associated with significantly reduced rates of hospitalisations, surgeries and complications(Reference Khanna, Bressler and Levesque52). Patients who have treatment failure with anti-TNF-α agents can be switched to ustekinumab or vedolizumab.
Changes in gut microbiota with inflammatory bowel disease therapeutics
5-ASA decreases inflammation through inhibition of NF-κB and pro-inflammatory eicosanoid production, and activation of PPAR-γ(Reference Punchard, Greenfield and Thompson53). However, its effects on microbiota composition remain unclear. A prospective cohort study by Morgan et al.(Reference Morgan, Tickle and Sokol34) observed significant reductions in Escherichia/Shigella abundance and modest increases in Enterococcus abundance in stool samples of one hundred thirty-one IBD patients on mesalamine(Reference Morgan, Tickle and Sokol34). These findings were corroborated by Xu et al.(Reference Xu, Chen and Wu54), who studied microbiota composition between the treatment-naïve UC cohort and 5-ASA-treated patients. They observed that there was a lower abundance of Escherichia/Shigella and an increased abundance of Firmicutes in patients on 5-ASA medication(Reference Punchard, Greenfield and Thompson53). Subsequent administration of 5-ASA for at least 6 months to treatment-naïve patients validated these specific alterations. However, both studies were limited to a single time point. Given that the IBD gut microbiota is typically more dynamic than healthy controls, it is essential to extend assessments of the microbiota over the course of treatments to identify whether these changes are sustained and treatment specific, or whether they simply reflect alterations in the degree of intestinal inflammation.
More recently, Schirmer et al.(Reference Schirmer, Denson and Vlamakis55) investigated changes in the gut microbiota of four hundred and five paediatric UC patients, treated with 5-ASA and corticosteroids, by comparing treatment-naïve baseline to week 4 samples grouped by treatment type and remission status. Eleven operational taxonomic units (OTUs) were associated with 5-ASA use whereas forty-seven OTUs were linked to corticosteroid treatment. Differences in bacterial abundance associated with remission following 5-ASA medication included OTUs belonging to Oscillospira, Eikenella, Rothia mucilaginosa, Clostridiales and Fusobacterium. OTUs associated with corticosteroid response included Actinomyces which increased in the week 4 remission group but decreased in those with sustained disease(Reference Schirmer, Denson and Vlamakis55). The inverse was observed for a Clostridium OTU. Additionally, species including Bifidobacterium, Fusobacterium, Dialister, Blautia producta and Eikenella, showed significant differences in their mean abundances between the remission and no-remission groups(Reference Schirmer, Denson and Vlamakis55). These findings differ from the previous cross-sectional analysis by Morgan et al.(Reference Morgan, Tickle and Sokol34), who observed that Enterococcus was the only genus altered during corticosteroid treatment, therefore suggesting that treatment response can be dependent on other factors including baseline microbiota composition. Nevertheless, this study was limited since the changes in microbial composition and remission status beyond 4 weeks are unknown. Furthermore, findings in the paediatric cohort may not be generalisable to the adult IBD population. Therefore, additional longitudinal prospective studies are needed.
Biologic agents
TNF-α is a pro-inflammatory cytokine produced by macrophages during acute inflammation. TNF-α signals through two transmembrane receptors, TNFR1 and TNFR2, and regulates a number of critical cell functions including cell proliferation, survival, differentiation and apoptosis(Reference Parameswaran and Patial56). TNF-α is considered a ‘master-regulator’ of inflammatory cytokine production because of its pivotal role in orchestrating the cytokine cascade in many chronic inflammatory conditions such as rheumatoid arthritis, psoriasis and IBD. Since the 1990s, the development and use of drugs which inhibit TNF-α action has revolutionised the management of these immune-mediated diseases(Reference Stevens, Chang, Weinberg and Buchholz57). Infliximab, adalimumab, golimumab and certolizumab are TNF-α inhibitors used in the treatment of IBD.
Treatment with TNF-α inhibitors has been shown to have a significant impact on faecal microbiota community profiles in CD. In both paediatric and adult studies, a compositional shift towards a healthier gut microbiota has been seen within 6 weeks of treatment. In addition, increases in SCFA producing bacterial species including Roseburia, Odoribacter, Fusobacterium and Prevotella have been reported with the more pro-inflammatory Klebsiella, Escherichia and Enterococcus genera decreasing significantly in patients achieving remission(Reference Zhou, Xu and He39,Reference Lewis, Chen and Baldassano58–Reference Magnusson, Strid and Sapnara60) . In general, TNF-α inhibitors treatment success is associated with higher bacterial diversity/richness at baseline and a decrease in Actinomyces and increase in Lactococcus and Roseburia, following 6 weeks of drug. The findings have been broadly confirmed in both adult and paediatric studies. Kolho and Sipponen(Reference Kolho and Sipponen61) assessed the effect of various TNF-α inhibitors in paediatric IBD patients, demonstrating that by week 6 of TNF-α inhibitor treatment, microbial diversity amongst patients and their similarity to the microbiota of controls increased in the responder group, but not amongst non-responders(Reference Kolho, Korpela and Jaakkola62). Furthermore, the increase in microbial diversity also correlated with an improved long-term outcome. Interestingly, there were six groups of bacteria whose abundance at baseline predicted the response to infliximab. These included genus-level groups Bifidobacterium, Clostridium colinum, Eubacterium rectale, uncultured Clostridiales and Vibrio. These bacteria have been associated with health as they are known to have immune-stimulatory properties and are found in high abundance during early childhood(Reference Huttenhower, Kostic and Xavier63). Responders also had a lower abundance at baseline of Streptococcus mitis. High microbial diversity at levels equivalent to controls was significantly associated with a sustained therapeutic response and lower calprotectin levels at 3 months, thus suggesting the potential of microbiome analysis to predict treatment outcomes. The recent longitudinal Swiss IBD cohort study also found that amongst CD patients receiving TNF-α inhibitors, increased Bifidobacterium, Collinsella, Lachnospira, Lachnospiraceae, Roseburia, Eggerthella taxa and reduced Phascolarctobacterium were associated with treatment success(Reference Yilmaz, Juillerat and Oyas12). One study also observed microbiota changes associated with infliximab discontinuation(Reference Rajca, Grondin and Louis64). In a study of thirty-three CD patients in stable remission on combined immunomodulator and infliximab therapy, infliximab was discontinued as a part of planned therapy de-escalation with faecal microbiota composition evaluated at baseline, 8 weeks, 6 months and at 18 months, by which time nineteen patients (58 %) had relapsed. There was no significant fluctuation in faecal microbiota composition across time-points in either relapsers or non-relapsers; there was also no correlation between microbial signals and inflammatory markers. Bacterial signals which corresponded with relapse however included reduced the numbers of Faecalibacterium prausnitzii, Bacteroides members and Clostridium coccoides (Reference Rajca, Grondin and Louis64).
In UC, Magnusson et al.(Reference Magnusson, Strid and Sapnara60) studied the microbiota composition of fifty-six biologic-naïve adult UC patients who commenced anti-TNF therapy. Based on stratification into responders v. non-responders after 12–14 weeks of treatment they found that responders had lower dysbiosis indices and a higher abundance of F. prausnitzii at baseline compared to non-responders(Reference Magnusson, Strid and Sapnara60). Furthermore, a longitudinal increase in F. prausnitzii was observed in responders at weeks 2 and 6. Additionally, the study analysed microbiota and proteomic data from faecal and mucosal biopsy samples respectively and found that at baseline, responders had detectable expression of several antimicrobial peptides or proteins while non-responders had expression of a protein which inhibited antimicrobial peptide expression. This difference in antimicrobial response was postulated to be a dysbiosis indicator and higher baseline expression of these proteins was potentially a predictor of anti-TNF-α therapy response.
Other biologic agents: anti-integrin antibody, anti-IL-12/IL-23 antibody
Vedolizumab is a humanised anti-α4β7 integrin monoclonal antibody that selectively blocks trafficking of memory T cells to inflamed gut tissue by inhibiting the α4β7-mucosal addressin cell adhesion molecule-1 interaction(Reference Rietdijk and D'Haens65). Approved for treating patients with moderately to severely active UC and CD, vedolizumab is generally considered safer than other biologics due to its gut-specific mode of action.
Recently, Ananthakrishnan et al.(Reference Ananthakrishnan, Luo and Yajnik42) evaluated the effects of vedolizumab on gut microbiota composition in forty-three patients with UC and forty-two patients with CD. In CD patients, five taxa significantly decreased in relative abundance between baseline and week 14 in patients achieving remission. These taxa include Bifidobacterium longum, Eggerthella, Ruminococcus gnavus, Roseburia inulinivorans and Veillonella parvula. In UC, only one taxon, Streptococcus salivarius, significantly increased in abundance in patients who did not achieve remission. The α diversity was significantly higher whereas the β diversity was lower in CD patients at baseline achieving remission by week 14. In particular, R. inulinivorans and a Burkholderiales species were significantly more abundant at baseline in patients achieving remission. Additionally, responders at week 14 demonstrated greater persistence of their microbiota changes at 1 year compared to non-responders, suggesting that early changes in microbiome could be an indicator of sensitivity to treatment and initial response. When the authors examined the trajectory of metabolic pathways during vedolizumab therapy, they noted more pronounced trends compared to microbial composition. This suggests that understanding the net metabolic and immunologic effect of resident microbiota, rather than that of single organisms is key to unravelling the mechanism of intestinal damage in IBD.
Another newer biologic agent to enter the scene of IBD treatment is ustekinumab, an inhibitor of IL-12 and IL-23. These are related cytokines that have been implicated in the pathogenesis of several immune-mediated disorders including IBD. They are heterodimers made up of a common p40 subunit complexed to unique p35 (IL-12) or p19 (IL-23) sub-units. Ustekinumab is a human monoclonal antibody that specifically binds the p40 subunit of IL-12/23, thus preventing IL-12 and IL-23 from binding to their cell surface receptor complexes, thereby blocking the T helper (Th) 1 (IL-12) and Th17 (IL-23) inflammatory pathways(Reference Feagan, Sandborn and Gasink66). Ustekinumab has proven efficacy in the treatment of moderate to severe CD and UC which are refractory to other therapies. Its long-term safety of doses used in the treatment of IBD is not yet well-established, but is believed to be similar to TNF-α inhibitors(Reference Sandborn, Gasink and Gao67).
Doherty et al.(Reference Doherty, Ding and Koumpouras68) conducted a multicentre randomised placebo-controlled phase 2B trial which analysed microbiota changes associated with ustekinumab treatment. The findings followed the trend reported in the majority of anti-TNF-α studies, with an increase in microbial diversity particularly SCFA-producing bacteria in treatment responders. Following 6-weeks of treatment, the relative abundance of Ruminococcaceae, Faecalibacterium, Blautia, Clostridium XIVa and Roseburia was higher, and the proportion of Shigella and Escherichia was lower amongst ustekinumab responders compared to non-responders. The α-diversity measures were also increased amongst ustekinumab responders at week 22, whereas no significant change was measured amongst non-responders or those who received placebo. In addition, baseline microbiota profiles were shown to predict response to ustekinumab. Baseline α-diversity amongst ustekinumab-treated patients in remission at 6 weeks was 1⋅7 times higher compared to those with persistent CD activity. Faecalibacterium and Bacteroides were significantly more abundant at baseline amongst patients who achieved in remission 6 weeks compared to those who did not.
Total parenteral nutrition and exclusive enteral nutrition
Dietary antigens have potential to stimulate mucosal immunity, therefore eliminating them through bowel rest with has been utilised as an early therapy in IBD(Reference Graham and Kandil69). Total parenteral nutrition where the patient is fasted whilst a mixture of lipids, glucose, amino acids, salts with added dietary minerals and vitamins is administered intravenously, emerged in the 1980s as an important strategy for the treatment of moderate to severe CD. In a prospective study, CD patients were treated with total parenteral nutrition and bowel rest. The majority achieved initial remission, but relapse was common once food was re-introduced(Reference Muller, Keller and Erasmi70). Conversely, there are very few data regarding the effectiveness of enteral therapy for UC. A small randomised trial of hospitalised patients with severe UC did not find any differences in response rates to corticosteroid therapy with a polymeric diet compared with total parenteral nutrition(Reference Gonzalez-Huix, Fernandez-Banares and Esteve-Comas71).
In recent decades, exclusive enteral nutrition (EEN) has superseded total parenteral nutrition as a safer dietary strategy for treating CD. EEN utilises diets composed of elemental, semi-elemental or defined formulae. It has proven efficacy for inducing remission and is often utilised as first-line therapy, particularly for paediatric patients with CD(Reference Caprilli, Gassull and Escher72,Reference Sandhu, Fell and Beattie73) . EEN's utility in UC is less well-established.
Despite its efficacy for CD, the mechanism of EEN action has not been fully characterised. Interestingly, there does not appear to be major differences in EEN efficacy based on the composition of the formula, with a Cochrane meta-analysis finding similar efficacy of formulas with variable degrees of protein hydrolysis in treating CD(Reference Zachos, Tondeur and Griffiths74). Several hypotheses have been proposed to explain the efficacy of EEN for CD. The most likely mechanisms include direct anti-inflammatory effects and improvement of intestinal barrier function. Modulation of gut microbiota has also been proposed although current data are sparse. In a small case series of nine paediatric CD patients treated with polymeric EEN, all experienced significant shifts in intestinal microbiota composition(Reference Lionetti, Callegari and Ferrari75). Another study measured the faecal abundance of five key bacterial taxa from CD patients treated with EEN compared with healthy controls on a standard diet. At baseline, bacterial diversity present was comparable. At week 8 of follow-up, CD patients displayed reduced microbial diversity, and this persisted for several months after cessation of EEN(Reference Leach, Mitchell and Eng76). The authors observed a paradoxical reduction in ‘protective’ gut bacterial species such as F. prausnitzii and metabolites including butyrate usually associated with reduced gut inflammation. Similar reduction in F. prausnitzii abundance was reported amongst healthy volunteers placed on EEN(Reference Benus, van der Werf and Welling77). These findings challenge the previous notion that increased microbiome diversity and higher numbers of F. prausnitzii are central to gut health.
To explore associations between the gut microbiota during EEN, Quince and colleagues utilised 16S rRNA sequencing and shotgun metagenomics to determine microbial composition of faecal samples from CD and healthy children. From the CD patients, faecal samples were collected before and during EEN, and after return to habitual diet. Microbial diversity was lower in CD than in controls before EEN. During EEN, the microbial diversity of CD children faecal samples further decreased. The reduction in diversity became apparent after only 15 d on EEN with lowest microbial diversity levels observed by 30 d of EEN treatment. A slight recovery towards the end of EEN and complete recovery of microbial diversity measures to pre-treatment levels were seen when patients returned to a regular diet. During EEN, the microbial community structure became less similar to that of controls compared to pre-EEN samples. The vast majority of changes represented a reduction in relative abundance, with some of the most negatively impacted genera being Bifidobacterium, Ruminococcus and Faecalibacterium. Their abundance was already lower at baseline in CD children compared to controls. The only genus that increased with EEN treatment was Lactococcus. In terms of microbial metabolic pathways, the abundance of genes involved in biotin and thiamine biosynthesis decreased during EEN, indicative of a reduction in bacteria that bear genes encoding for these vitamins, such as Bifidobacteria and E. coli spp., or changes in the synthesis of SCFA that require these vitamins. Conversely, pathways involved in spermidine/putrescine biosynthesis increased(Reference Quince, Ijaz and Loman78). These pathways are known to play a major role in cell growth, and are considered a marker of cell renewal and epithelial healing(Reference Slezak, Hanske and Loh79). Tjellstrom and colleges also reported an increase in SCFA production amongst CD patients commenced on EEN(Reference Tjellstrom, Hogberg and Stenhammar80).
In the recently published CD-TREAT study, Svolos and colleagues evaluated the effects of an individualised food-based diet, with similar composition to EEN, on the gut microbiota, inflammation and clinical response(Reference Svolos, Hansen and Nichols81). Using a combination of animal studies and human studies including five children with relapsing CD, they created a solid food EEN by the exclusion of certain dietary components (e.g. gluten, lactose and alcohol) and matching of others (macronutrients, vitamins, minerals and fibre) as closely as possible using ordinary food. When compared to conventional EEN, the CD-TREAT diet induced similar effects on faecal microbiome composition, metabolome and mean total sulphide. Sulphides is a bacterial product known to break down mucus barrier and been linked to intestinal inflammation(Reference Ijssennagger, van der Meer and van Mil82). Similar effects on bacterial load reduction and SCFA composition were also comparable between the two diets. Amongst the children with CD, 8 weeks on the CD-TREAT diet led to clinical improvement in 80 % and clinical remission in 60 %(Reference Svolos, Hansen and Nichols81). Although a pilot study which requires extensive validation and extended timeframe evaluation, the landmark study clearly demonstrates how through careful consideration of nutrition it is possible to achieve equivalent if not superior effects on IBD symptoms, thus providing additional less toxic therapeutic options for disease remission and management.
Probiotics
The success of probiotics in the management of IBD ranges from mixed results to considerable potential and is dependent on the strains used and disease subtype targeted. The most encouraging studies have been in the non-pathogenic strain of E. coli Nissle 1017, as well as VSL#3, which contain four strains of Lactobacilli (L. casei, L. plantarum, L. acidophilus and L. delbrueckii subsp. bulgaricus), three strains of Bifidobacteria (B. longum, B. breve and B. infantis) and one strain of Streptococcus (S. salivarius subsp. thermophilus). These probiotics have been shown to be effective against recurrence of pouchitis after surgery, as well as in the induction and maintenance of remission in UC(Reference Orel and Kamhi Trop83).
Faecal microbiota transplantation
In simple terms, faecal microbiota transplantation (FMT) replaces the subject's dysbiotic gut microbiota with microbes from ‘healthy’ donors. As early as 1989, FMT was being used to treat IBD, sometimes with dramatic responses. A systematic review, published in 2012, containing twenty-six subjects found that nineteen patients experienced symptomatic improvement, thirteen ceased taking IBD medications within 6 weeks and fifteen had no active disease 3–36 months following FMT(Reference Anderson, Edney and Whelan84).
Bennet and Brinkman's initial report documented the complete clinical remission of a case of UC for at least 6 months following FMT through retention enema(Reference Bennet and Brinkman85). A paediatric case series subsequently found that seven out of nine patients with mild-to-moderate UC disease activity experienced clinical improvement, and three achieved clinical remission within 1 week after a 5-d course of daily FMT enema(Reference Kunde, Pham and Bonczyk86). In contrast, no clinical improvement was observed in two smaller studies(Reference Suskind, Singh and Nielson87,Reference Kump, Grochenig and Lackner88) .
More recently, following the landmark study by van Nood et al.(Reference van Nood, Vrieze and Nieuwdorp89) describing the superiority of FMT for the treatment of recurrent Clostridioides difficile infection, a number of studies have been published describing FMT treatment in IBD. Interestingly, FMT does not appear to be effective in CD, with all positive studies assessing UC cohorts. Moayyedi and colleagues randomised seventy patients with active UC to receive weekly FMT or placebo enemas(Reference Moayyedi, Surette and Kim90). By week 7, clinical and endoscopic remission was achieved in nine of thirty-eight (24 %) patients in the FMT arm v. two of thirty-seven (5 %) in the placebo arm, with no difference in adverse event rates reported between the groups. Interestingly, seven of the nine patients in remission after FMT received faecal material from a single donor, highlighting the possibility of the FMT ‘super donor’(Reference Moayyedi, Surette and Kim90). After 6 weeks of treatment, there was a statistically significant change in microbiota composition, with an increased microbial diversity in the FMT group compared with the placebo group. Donor stool enrichment for the family Lachnospiraceae and the genera Ruminococcus was associated with successful treatment. A further study by Rossen and colleagues randomised fifty UC patients to receive either donor or autologous (patient own) FMT by nasoduodenal infusion but the study failed to detect a difference. The clinical response rate was 52 % in the control and 43⋅5 % in the treatment group (P = 0⋅58). At 12 weeks after treatment, the diversity index amongst responders in both groups increased, whereas no change in diversity was detected amongst non-responders. Microbial composition of responders in the treatment group shifted towards that of their donors by week 12, with regain of Clostridium clusters IV, XIVa, and XVIII, and reduction in Bacteroidetes. Responders in the control group also displayed a change in microbiome composition, but unlike FMT-treated responders, this shift was mostly associated with an increase in abundance of Bacilli, Proteobacteria, and Bacteroidetes(Reference Rossen, Fuentes and van der Spek91).
Two subsequent studies utilised pooled stool from healthy donors. Paramsothy and colleagues randomised eighty-one patients to receive donor stool or placebo(Reference Paramsothy, Kamm and Kaakoush92). The index dose was administered via colonoscopy, and this was followed by an intensive regimen of daily enemas for 8 weeks. The primary outcome of steroid-free clinical remission was achieved in 27 % of donor-stool recipients v. 8 % of those assigned placebo (P = 0⋅021), with no difference in adverse effects. Stool microbial profiles of patients after donor FMT shifted from a dominance of Bacteroides spp. to Prevotella spp., bringing them closer to the donor's profile. Several microbial taxa were associated with remission including Barnesiella spp., Parabacteroides spp., Clostridium cluster IV and Ruminococcus spp. Conversely, Fusobacterium spp. and Sutterella spp. were associated consistently with the lack of FMT response(Reference Paramsothy, Kamm and Kaakoush92).
Costello and colleagues were the first to utilise anaerobically processed pooled healthy donor stools for FMT in seventy-three patients with mild to moderately active UC(Reference Costello, Hughes and Waters93). The rationale for this approach was that anaerobic processing helped to achieve a microbial profile which more closely mimicked intra-luminal conditions. With one colonoscopic FMT dose followed by two subsequent enemas, patients receiving donor stool demonstrated higher rates of clinical and endoscopic remission at 8 weeks compared to the placebo arm which received autologous stool. A total of 42 % of remitters also maintained steroid-free remission to 12 months. As expected, diversity increased following donor FMT and this was maintained up to 8 weeks following treatment. An increased abundance of Anaerofilum pentosovorans and Bacteroides coprophilus was strongly associated with disease improvement following donor FMT. SCFA levels were not significantly different between treatment groups at weeks 4 or 8 and did not predict clinical outcome(Reference Costello, Hughes and Waters93).
Conclusions
We have come a long way in our understanding of IBD pathogenesis and the involvement of the gut microbiota, but there is still a long way to go and defining optimal treatment strategies for patients remains a challenge. Lessons learnt along the way include the need to appreciate the bigger picture by allowing the study design to capture the differing aspects of this complex disease. This includes the need to focus on well-phenotyped patient cohorts, use of prospective longitudinal cohorts, appreciation of inflammatory and treatment confounders as well as considering differences due to geography, age and diet. Even with the consideration of these requirements, there remains the need to ensure robust standardised scientific methodology is applied consistently to ensure the quality of the findings.
Financial Support
The IBD research programme at the Microbiome Research Centre, UNSW is supported by funds from Crohn's Colitis Australia in the form of a scholarship to N. W. We have also received support from St George and Sutherland Medical Research Foundation. G. H. is also supported by funds from the Australian Research Council (DP210103897).
Conflict of Interest
None.
Authorship
The authors had sole responsibility for all aspects of preparation of this paper.



