



Introduction
The analysis of patterns of paleobiogeography depends on a global data set. Efforts to collect information on the age and location of disparate fossil species, such as the Paleobiology Database and the Geobiodiversity Database, among others, are critical in defining global patterns, but these depend on a robust primary literature describing occurrences of fossil taxa.
Reports of Paleozoic crinoids from Mongolia are infrequent, and many of the earliest papers described only stems (Stukalina, Reference Stukalina, Pormnov and Suzukov1973, Reference Stukalina1994, Reference Stukalina1997; Tungalag, Reference Tungalag1998). Webster and Ariunchimeg (Reference Webster and Ariunchimeg2004) described a Lower Devonian (Emsian) fauna from Shine Jinst in southern Mongolia, including the first crinoid cups and thecae from Mongolia. They recognized five distinct taxa but named only Cyathocrinites because of poor preservation. Waters et al. (Reference Waters, Waters, Königshof, Carmichael and Ariuntogos2021) described a Late Devonian (Famennian) echinoderm fauna of twelve crinoid genera and two blastoid genera from southwestern Mongolia. Recently, Carboniferous (Serpukhovian) crinoids have been collected from southwestern Mongolia, but they are too fragmentary for identification (Waters, personal observation, 2020).
In this paper, we describe the new periechocrinitid Gennaeocrinus tariatensis new species from the Emsian of Mongolia. Gennaeocrinus is well known from multiple species from the Devonian of North America and is questionable from western Europe. In terms of Devonian plate tectonics, it is well known from cratonic settings on east/southeast Laurentia and questionably known from similar settings on southwest Baltica and Iberia in the reconstructions of Domeier and Torsvik (Reference Domeier and Torsvik2014). However, G. tariatensis was collected from a collage of small terranes and active island arc complexes in the Central Asian Orogenic Belt on the southeastern margin of Siberia. Thus, its tectonic, paleogeographic, sedimentological, and paleolatitudinal occurrence differs from all other species of the genus.
Geological setting
Mongolia is a very large country and occupies a primary position in the Central Asian Orogenic Belt (Fig. 1), the largest Paleozoic orogenic belt that evolved in a manner similar to the modern Indonesia and western Pacific. Mongolia comprises 44 terranes (Badarch et al., Reference Badarch, Cunningham and Windley2002), including cratonic, metamorphic, passive margin, island arc, forearc/backarc, accretionary complex, and ophiolitic terranes (Fig. 2).

Figure 1. The Central Asian Orogenic Belt accretionary zone located between the European and Siberian cratons to the north and the Tarim and North China cratons to the south. Map after Ju and Hou (Reference Ju and Hou2014).
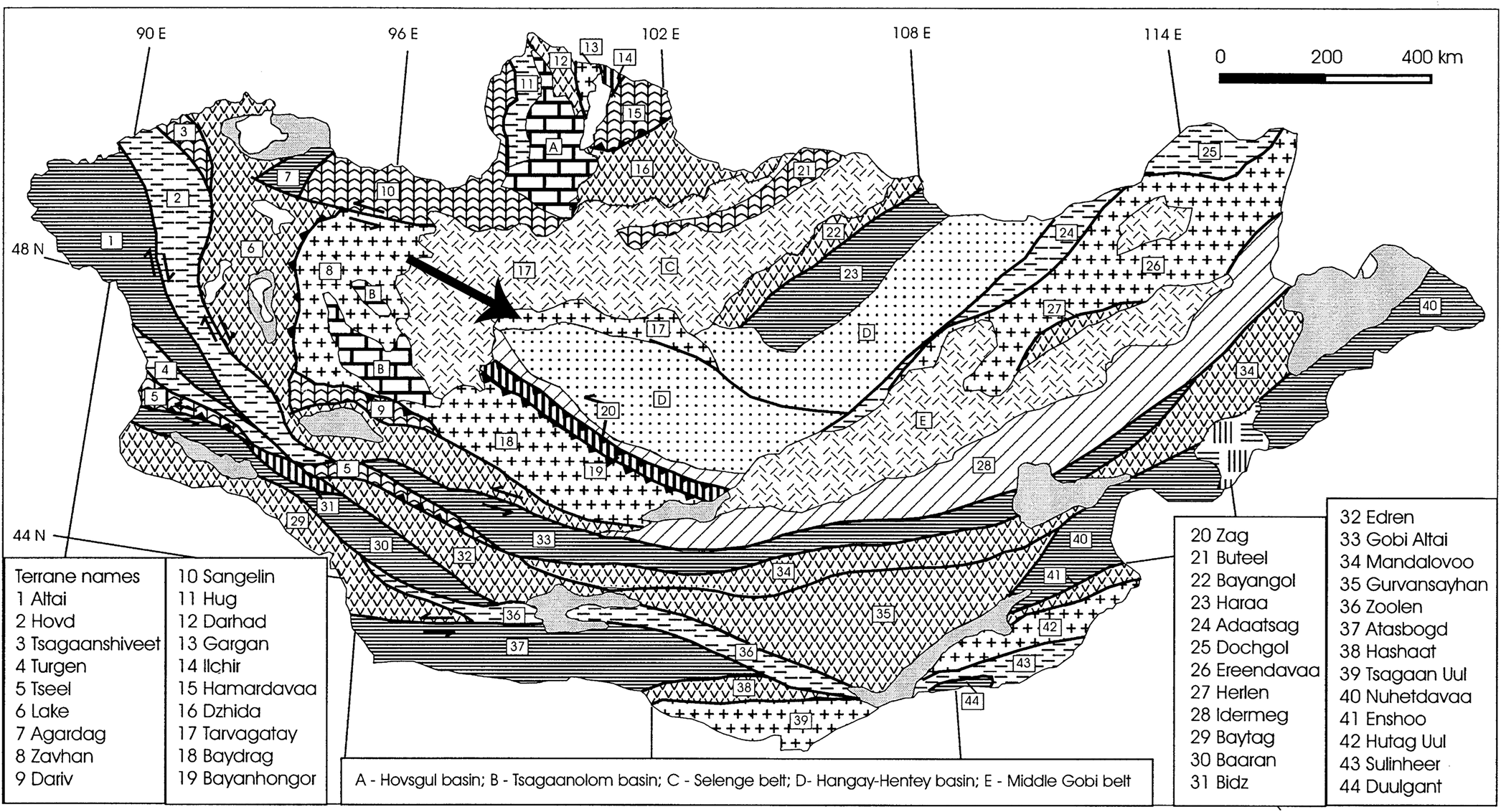
Figure 2. Map of the 44 tectonic terranes of Mongolia (after Badarch et al., Reference Badarch, Cunningham and Windley2002). The crinoid locality, indicated by the arrow, is located on the Tarvagatay Terrane, an arc terrane that accreted to the southern margin of the Siberian Craton.
The crinoid described herein was collected from the Tariat Formation, which is part of the Tarvagatay Terrane of Badarch et al. (Reference Badarch, Cunningham and Windley2002). The Tarvagatay Terrane occurs in the northern Hangay mountains. The Terrane consists of Precambrian basement overlain by lower Cambrian stromatolitic limestone and volcaniclastic rocks. Devonian and Mississippian conglomerate, sandstone, and siltstone containing marine fossils complete the sequence. Alekseeva (Reference Alekseeva1993) described the Tariat Formation as a terrigenous sequence with 70 m of basal conglomerates overlain by 150 m of green sandstones, siltstones, and mudstones with abundant brachiopods and plant fossils in darker siltstones. An overlying terrigenous sequence contains 250 m dark-gray to greenish siltstones and sandstones also with abundant brachiopods. The Tariat Formation is unconformably overlain by lower Carboniferous conglomerates. The Tariat Formation is Emsian in age according to the presence of the strophomenoid Maoristrophia kailensis Shishkina, Reference Shishkina1990, a brachiopod that defines the Mongolian Chulun Horizon (Alekseeva, Reference Alekseeva1993). This horizon correlates with the Salairka Horizon in Siberia and the Polygnathus kitabicus Conodont Biozone in the basal Emsian (Yolkin et al., Reference Yolkin, Gratsianova, Izokh, Yazikov, Bakharev, Alekseeva, Erina, Kim and Shishkina2000).
Paleobiogeography
The tectono-stratigraphic compilations of Mongolia by Badarch et al. (Reference Badarch, Cunningham and Windley2002) and Windley et al. (Reference Windley, Alexeiev, Xiao, Kröner and Badarch2007) recognized the Main Mongolian Lineament, which divides the country into an early Paleozoic domain in the north and a late Paleozoic domain in the south. The Tarvagatay Terrane is part of the Mongol–Okhotsk Belt north of the Main Mongolian Lineament. The Tariat Formation was deposited in a trench–accretionary wedge environment in the Mongol–Okhotsk Ocean to the south (in current orientation) of the Siberian Craton (Bussien et al., Reference Bussien, Gombojav, Winkler and von Quadt2011).
Emsian paleobiogeography is based primarily on brachiopods and shows differentiation into the Rhenish–Bohemian Realm, the Appalachian Realm, and the high-latitude Malvinokaffric Realm. Some Devonian workers combine the Rhenish–Bohemian Realm into the Appalachian Realm. Brachiopods with a cosmopolitan distribution from the Mongol–Okhotsk fauna support the conclusions of Hou and Boucot (Reference Hou, Boucot, McKerrow and Scotese1990) that they are part of the Old World Realm even though geographic distances between the areas are quite large. Paleobiogeographic reconstructions by Torsvik and Cocks (Reference Torsvik and Cocks2017) placed the Mongolian terranes, including the Tarvagatay Terrane, along the southern margin of the Siberian Craton at a latitude of approximately 50°N. The Old World Realm is primarily equatorial to mid-latitudes, and the Malvinokaffric Realm is located in the high latitudes of the Southern Hemisphere (Torsvik and Cocks, Reference Torsvik and Cocks2017).
Emsian faunas have a high degree of cosmopolitan distribution of taxa where adequate data are available. The Emsian marked the acme of a remarkable spread of reefs, the largest known and most latitudinally widespread reefs in the Phanerozoic according to Copper and Scotese (Reference Copper, Scotese, Chan and Archer2003). Judging from the distribution of reefs, they concluded that the Middle Devonian (Emsian–Givetian) had a supergreenhouse climate. However, Joachimski et al. (Reference Joachimski, Bresig, Buggisch, Talent, Mawson, Gereke, Morrow, Day and Weddige2009), concluded that the Middle Devonian had sea surface temperatures closer to 23–25°C and was not a supergreenhouse climate according to oxygen isotopes in conodont apatite. Regardless of which temperature model is correct, Emsian sea levels were high, and geographically disparate faunas contained many similar, cosmopolitan genera.
Materials and methods
The holotype of Gennaeocrinus tariatensis n. sp. was collected from an outcrop of the Tariat Formation at 48°12′23″N, 100°1′49″E (measured by GPS) in Arkhangai Aimag, Tariat Somon, Mongolia (Fig. 3). The specimen was a mold preserved in a greenish–gray fossiliferous siltstone. Descriptions of the specimen are based on a latex cast prepared at the Paleontological Center of the Mongolian Academy of Science and photographed with ammonium chloride sublimate.

Figure 3. Field photo of outcrops of the Tariat Formation located in the Arkhangai Aimag, Tariat Somon, Mongolia. The Tariat is an Emsian fossiliferous unit composed primarily of conglomerates, sandstones, and siltstones. The arrow points to the crinoid locality.
Repository and institutional abbreviation
The latex cast of the holotype is reposited in the collections of the Mongolian University of Science and Technology (MUST) research collections in Ulaanbaatar, Mongolia.
Systematic paleontology
The superfamilial classification used here follows Cole (Reference Cole2017), Wright (Reference Wright2017), and Wright et al. (Reference Wright, Ausich, Cole, Peter and Rhenberg2017); family-level classifications follow Moore and Teichert (Reference Moore and Teichert1978). Morphologic terminology follows Ubaghs (Reference Ubaghs, Moore and Teichert1978) and Ausich et al. (Reference Ausich, Wright, Cole and Sevastopulo2020). The plating of interrays is given by the number of plates in each range from proximalmost plate to the last range before the tegmen. In the posterior interray, the primanal is indicated by “P,” and the first interradial plate in regular interrays is indicated by “1.”
Subclass Camerata Wachsmuth and Springer, Reference Wachsmuth and Springer1885
Infraclass Eucamerata Cole, Reference Cole2017
Order Monobathrida Moore and Laudon, Reference Moore and Laudon1943
Family Periechocrinidae Bronn, Reference Bronn1849
Genus Gennaeocrinus Wachsmuth and Springer, Reference Wachsmuth and Springer1881
Type species
Actinocrinus kentuckiensis Shumard, Reference Shumard1868, by original designation.
Other species
G. carinatus Wood, Reference Wood1901; G. carinatus crassicostatus Goldring, Reference Goldring1923; G. chilmanae Kesling, Reference Kesling1968; G. chilmanae Kesling, Reference Kesling1968; G. cornigerus (Lyon and Casseday, Reference Lyon and Casseday1859); G. decorus Goldring, Reference Goldring1923; G. dulukae Kesling, Reference Kesling1969; G. eucharis (Hall, Reference Hall1862); G. facetus Rowley, Reference Rowley and Green1903; G.? germanicus Schmidt, Reference Schmidt1941; G. goldringae Ehlers, Reference Ehlers1925; G. maxwelli Johnson and Lane, Reference Johnson and Lane1969; G. mourantae Goldring, Reference Goldring1934; G. nyssa (Hall, Reference Hall1862); G. perculiaris Goldring, Reference Goldring1923; G. percarinatus Goldring, Reference Goldring1935; G. romingeri Kesling, Reference Kesling1964; G. sculptus Rowley, Reference Rowley and Green1903; G. seversoni Laudon, Reference Laudon1973; G. similis Goldring, 1935; G. simulans Rowley, Reference Rowley and Green1904; and G. variabilis Kesling and Smith, Reference Kesling and Smith1962.
Occurrence
Previously, this genus was known from the Emsian to Givetian in North America, with questionable occurrences in Germany and Spain. The specimen described herein is from the Emsian of Mongolia.
Remarks
Wachsmuth and Springer (1881, p. 160) proposed Gennaeocrinus “for a little group of crinoids from the Upper Devonian, which cannot be satisfactorily referred to any established genus.” They noted differences in the plating of the anal area and the number and arrangement of both anal and interradial plates. As presently understood, Gennaeocrinus is a common periechocrinid crinoid from the Lower and Middle Devonian of North America with questionable occurrences in Germany and Spain (Ausich and Kammer, Reference Ausich, Kammer, Ausich and Webster2008; Webster and Webster, Reference Webster and Webster2014), but this Devonian genus has a high morphological disparity with, for example, aboral cup shapes varying from very low cone or bowl shape to medium globe shape and a wide range in aboral plate sculpturing, presence or absence of a basal ridge and a basal concavity, plating arrangements in all interrays, the number of fixed secundibrachials, and number of free arms. A comprehensive review of Gennaeocrinus is needed, but this is beyond the scope of the present study.
The generic assignment of this new Mongolian species would be questionable, except that it belongs to a group of Gennaeocrinus species that includes G. kentuckiensis (Shumard, Reference Shumard1868), the type species of the genus. Although this new specimen is a partial calyx, its morphology is distinct from other species and is considered a new species. The Gennaeocrinus species grouping to which G. tariatensis n. sp. belongs includes G. comptus Rowley, Reference Rowley and Green1903, G. kentuckiensis (type species), G. maxwelli, G. percarinatus, G. romingeri, and G. sculptus. These seven species all have a calyx shape of low cone/bowl to very low cone/bowl, multiple radiating ridges connecting to like ridges on adjoining calyx plates, basal concavity absent, and posterior interray plating P-3- (where known). As delineated in the following, species diagnostic characters for this group of species include calyx shape, presence or absence of an arcuate ridge on the radial plates, presence or absence of a central node on calyx plates where radiating ridges coalesce, presence or absence and character, if present, of a rim around the base of the calyx, presence or absence of prominent ray ridges, proximal regular interray plating, presence or absence of an anitaxial ridge, presence or absence or spinose tegmen plates, and number of free arms per ray.
Gennaeocrinus tariatensis new species
Figure 4
Holotype
The holotype and only known specimen is IV.JW.2013.1-5 reposited in the collections of the Paleontological Center of the Mongolian Academy of Sciences, Ulaanbaatar, Mongolia.

Figure 4. Gennaeocrinus tariatensis n. sp. (1) Photograph of the holotype of Gennaeocrinus tariatensis preserved as a mold in a greenish-gray siltstone. (2) Latex cast of the holotype showing prominent ornament of radiating ridges that cross plate boundaries. (3) Plate diagram of Gennaeocrinus tariatensis prepared by W.I.A. (4) Enlargement of the latex cast of Gennaeocrinus tariatensis showing subtle plate boundaries largely obscured by strong radiating ridges. Scale bars = 1 cm.
Diagnosis
Low to very low bowl calyx shape, multiple stellate ridges connecting to like ridges on adjoining plates throughout the calyx, arcuate ridge on radial plates absent, central node on calyx plates absent, basal ridge around base of calyx absent, prominent ray ridges absent, regular interray plating 1-3, anitaxial ridge absent or very weak, nature of tegmen plates and free arms per ray unknown.
Occurrence
Tariat Formation (Emsian), Arkhangai Aimag, Tariat Somon, Mongolia.
Description
Calyx low to flat bowl shaped, arms probably grouped; calyx plate sculpturing multiple radiating ridges from plate centers and connecting with like ridges on adjacent plates; central nodes are not formed where ridges coalesce at plate centers. Basal circlet flat cone shaped, a small percentage of overall calyx height, column cicatrix occupying most of basal circlet, individual sutures between basal plates not clearly preserved. Radial circlet projects outward, presumably slightly visible in lateral view, a small percentage of overall calyx height, radial plates presumably five, hexagonal or heptagonal, approximately as wide as high, in lateral contact except in the CD interray.
Regular interrays do not interrupt radial plate circlet, in contact with tegmen. First interradial plate hexagonal, higher than wide, approximately the same size as radial plates and first primibrachial plates; second range with two plates, more distal plating not preserved, but more than two plates. Interradial regions connect to tegmen.
Primanal higher than wide, probably hexagonal, positioned between the C and D radial plates, CD interray much wider that regular interrays, plating P-3-6-?.
First primibrachial fixed, hexagonal wider than high, smaller than radial plates and approximately the same size as the first interradial plate in regular interrays and the second primibrachial; second primibrachial axillary, presumably pentagonal or heptagonal; fixed secundibrachials present.
Tegmen and free arms unknown. Proximal column circular, but other aspects unknown.
Etymology
A species of Gennaeocrinus from the Tariat Formation (Emsian).
Remarks
Gennaeocrinus tariatensis n. sp. is represented by one specimen, which is an incomplete calyx with breaks in plating in places. Normally such a specimen would be insufficient as the foundation of a new species. However, as previously noted, this specimen can be confidently placed in Gennaeocrinus, and its morphology is unique among species of Gennaeocrinus. The following characters distinguish Gennaeocrinus tariatensis within the grouping of similar species: a low to very low bowl calyx shape; arcuate ridge on radial plates absent; central node on calyx plates absent; sharp ridge around base of calyx absent; ray ridges absent, proximal normal interray plating 1-2-; absent or weak anitaxial ridge; tegmen unknown; character of tegmen plates and number of free arms per ray unknown. By contrast, G. comptus has a very low cone calyx shape; arcuate ridge on radial plates present; central node on calyx plates absent, discontinuous, sharp ridge around base of calyx; prominent ray ridges; proximal normal interray plating 1-3?-; prominent anitaxial ridge; one central spine on tegmen; six arms per ray. G. kentuckiensis (type species) has a low bowl shape; arcuate ridge on radial plates absent; central node on calyx plates; continuous, low ridge around base of calyx; prominent ray ridges; proximal normal interray plating 1-2- or 1-3?-; weak anitaxial ridge; spines on tegmen plates; and eight arms per ray. G. percarinatus has a low(?) bowl calyx shape; presence or absence of arcuate ridge on radial plates unknown; central node on calyx plates; sharp ridge around base of calyx discontinuous, small, sharp; prominent ray ridges; proximal normal interray plating 1-2-; weak anitaxial ridge; tegmen unknown; and number of free arms unknown. G. romingeri has a very low bowl shape; arcuate ridge on radial plates absent; central node on calyx plates; sharp ridge around base of calyx absent; prominent ray ridges; proximal normal interray plating 1-2-; weak anitaxial ridge; tegmen unknown; and four free arms per ray. G. sculptus has a low bowl shape; arcuate ridge on radial plates absent; central node on calyx plates; sharp, continuous ridge around base of calyx unknown; prominent ray ridges; proximal normal interray plating 1-2-; anitaxial ridge absent; tegmen spines absent; six free arms per ray. G. maxwelli is a very poorly preserved, silicified specimen, and characters other than recognizing its presence within this grouping of species cannot be discerned with confidence.
Discussion
Gennaeocrinus is a monobathrid crinoid that is well known from the Emsian and Givetian of North America with possible occurrences from Germany and Spain. By contrast, G. tariatensis n. sp. is the first species of the genus found in the Central Asian Orogenic Belt (CAOB). It was collected from Emsian terrigenous sediments on a convergent margin terrane in the Mongol–Okhotsk Belt, which accreted onto the Siberian Craton. In terms of paleolatitude, previously known species of Gennaeocrinus have an equatorial distribution, but Gennaeocrinus tariatensis occurs at 50°N. The occurrence of Gennaeocrinus tariatensis significantly increases the paleogeographic range of Gennaeocrinus, but information is still lacking to hypothesize on patterns of migration. The occurrence of Gennaeocrinus in the Mongol–Okhotsk fauna is not completely unexpected as Emsian brachiopods have a high degree of cosmopolitanism and significant similarity between taxa in the Old World Realm and the Mongol–Okhotsk fauna (Hou and Boucot, Reference Hou, Boucot, McKerrow and Scotese1990).
Webster and Arunichimeg (Reference Webster and Ariunchimeg2004) described an Emsian crinoid fauna from the Chuluun Formation in the Shine Jinst area of Mongolia. In contrast to the terrigenous nature of the Tariat Formation, the Chuluun Formation is dominated by carbonates with mound- and reef-forming stromatoporoids and corals in addition to crinoidal grainstones. This locality is south of the Main Mongolian Lineament and is located on either the Mandavooloo or Gobi Altai Terrane. In contrast to assertions by Webster and Arunichimeg (Reference Webster and Ariunchimeg2004) that the Chuluun crinoids were the most northerly known Emsian crinoid fauna, recent paleogeographic reconstructions (Torsvik and Cocks, Reference Torsvik and Cocks2017) of the CAOB place the Mandavooloo and Gobi Altai Terranes at a latitude of 25–30°N. Gennaeocrinus tariatensis is the most northerly known Emsian crinoid, with a paleolatitude of approximately 50°N.
Acknowledgments
J.A.W.'s participation in fieldwork in Mongolia was funded in part by a grant to M.D. Brazeau, Imperial College, London, by the European Research Council. Y.A. Ariunchmeg provided critical assistance during the fieldwork. S. Gonchigdorj provided access to historical literature on the Devonian of Mongolia. We also thank P. Dittoe for helping attain access to crinoid literature when OSU libraries were closed. We thank the two reviewers who improved this manuscript.





 Open access
Open access