


Dietary fat has been implicated in the prevalence of obesity due to its high energy density and palatability(Reference Clegg and Shafat1). Despite this, there is evidence that some lipids may elicit greater satiety than others. Medium-chain TAG (MCT) have been shown to increase satiety compared with long-chain TAG (LCT)(Reference Van Wymelbeke, Himaya and Louis-Sylvestre2–Reference Van Wymelbeke, Himaya and Louis-Sylvestre2) which has been attributed to their quick absorption and metabolism(Reference Bach and Babayan8) and their involvement in the production of ketones which in turn have been shown to yield satiating properties(Reference Marten, Pfeuffer and Schrezenmeir9). In addition to a potential effect on satiety, MCT have been shown to increase energy expenditure (EE) compared with LCT by increasing dietary-induced thermogenesis (DIT)(Reference Clegg, Golsorkhi and Henry6).
Considerable research has been undertaken to understand better the role of exercise in appetite regulation, and it is well known that exercise transiently reduces appetite, a phenomenon referred to as ‘exercise-induced anorexia’(Reference King, Burley and Blundell10). This, combined with the energy expended during exercise, results in significant energy deficits(Reference King, Wasse and Ewens11). The mechanisms of exercise-induced anorexia differ to those of MCT, as acylated ghrelin, a potent stimulator of hunger, is suppressed in response to exercise(Reference King, Wasse and Ewens11, Reference Broom, Stensel and Bishop12), and anorexigenic hormones such as peptide tyrosine tyrosine (PYY), glucagon-like peptide-1 and pancreatic polypeptide are transiently elevated in the postexercise period(Reference Dorling, Broom and Burns13). Thus, it is possible that the combination of MCT and exercise may cumulate in a greater energy-deficit effect via these separate mechanisms, though this is currently not known.
Gastric emptying (GE) plays an important role in the regulation of appetite(Reference Janssen, Vanden Berghe and Verschueren14), as satiety has been shown to be inversely correlated with GE(Reference Hveem, Jones and Chatterton15). A delay in GE increases satiety(Reference Hunt16), and fat has been shown to delay GE to a greater extent compared with other macronutrients, which is mediated by cholecystokinin (CCK)(Reference Boyd, O ’donovan and Doran17). Whereas early findings suggested only fatty acids with chain lengths of twelve and above stimulate CCK release(Reference McLaughlin, Lomax and Hall18), later studies showed that medium-chain fatty acids do stimulate CCK release above baseline but not to the same extent as long chains(Reference Feltrin, Little and Meyer19). Despite this, MCT has been shown to delay GE compared with various sources of LCT(Reference Clegg, Pratt and Markey20), which is believed to be due to MCT having a higher osmolarity than LCT when energy content is equal. Exercise also affects GE, with lower intensities accelerating GE and higher intensities slowing it(Reference Horner, Schubert and Desbrow21). Delayed GE as well as increased production of ketones is believed to be the mechanisms by which MCT increases satiety. These mechanisms are quite different to those of exercise-induced satiety; and as such we believe that in combination, they could be cumulative. The interaction of MCT and exercise on GE, and its implications for appetite control, has not yet been examined.
To our knowledge, no study to date has investigated the combination of MCT and exercise on appetite and subsequent energy intake. As energy balance relies on both EE and energy intake, the research investigating the combination of exercise and satiety becomes of particular interest. Furthermore, as exercise is known to transiently elevate satiety hormones, it is important to examine whether this elevation augments the satiety response to MCT. The present study was designed to examine the possibly cumulative effects of MCT and an acute bout of exercise on satiety, which has not been previously investigated. Therefore, the aim of the present study was to elucidate the effects of MCT, exercise or a combination of the two on subjective appetite sensations, energy intake and overall energy balance.
Methods
Trial registration
ISRCTN registry, ISRCTN15611082, registered on 19 April 2017. Retrospectively registered.
Participants
Inclusion criteria were as follows: male, BMI 18·5−29·9 kg/m2, 18–65 years of age, weight stable for the 3 months leading up to commencement of the study, taking no medication that could affect appetite, non-smokers and unrestrained eaters (as determined by the Three-Factor Eating Questionnaire(Reference Stunkard and Messick22) and Dutch Eating Behaviour Questionnaire(Reference van Strien, Frijters and Bergers23)). After institutional ethical approval, twelve healthy males (age 27·0 (sd 11·4) years, weight 73·9 (sd 12·7) kg, height 175·9 (sd 8·0) cm, BMI 23·8 (sd 2·9) kg/m2, body fat 17·1 (sd 3·8) %), who were recruited through social media, posters and a research activity mailing list, completed a medical questionnaire and gave their written consent.
Screening
Height (Seca 217, Seca) was measured to the nearest 0·1 cm and body mass and body fat percentage (BC-418 MA; Tanita) to the nearest 0·1 kg and 0·1 %, respectively. Cycling (Lode Corival; Lode B.V.)
 $\dot V$
O2peak was determined using a ramp test where workload increased at a rate of 30 W/min until volitional exhaustion. Expired air was collected and measured using a metabolic stress testing system (CPX Ultima; Medical Graphics Ltd), which has been previously used as an indirect calorimeter during rest and exercise(Reference Winkle, Evans and Dilg24). Heart rate and rate of perceived exertion (RPE)(Reference Borg25) were taken every minute and at the termination of the test, upon exhaustion. Exhaustion was determined when the participants were unable to maintain a pedal cadence of 60 rpm as well as two of the following: heart rate within ≥95 % of age-predicted heart rate (220-age), RPE ≥19, or RER of >1·1(Reference Midgley, McNaughton and Polman26).
$\dot V$
O2peak was determined using a ramp test where workload increased at a rate of 30 W/min until volitional exhaustion. Expired air was collected and measured using a metabolic stress testing system (CPX Ultima; Medical Graphics Ltd), which has been previously used as an indirect calorimeter during rest and exercise(Reference Winkle, Evans and Dilg24). Heart rate and rate of perceived exertion (RPE)(Reference Borg25) were taken every minute and at the termination of the test, upon exhaustion. Exhaustion was determined when the participants were unable to maintain a pedal cadence of 60 rpm as well as two of the following: heart rate within ≥95 % of age-predicted heart rate (220-age), RPE ≥19, or RER of >1·1(Reference Midgley, McNaughton and Polman26).
Protocol
Participants completed four trials: two resting trials, either with the control oil (Con-R) or MCT oil (MCT-R), and two exercise trials with either the control oil (Con-Ex) or MCT oil (MCT-Ex). Trials were performed at 2–10 d apart, and each participant conducted their tests at the same time of day beginning between 07.00 and 09.00 hours. Trials were administered in a randomised order.
In the 24 h preceding the first trial, participants were required to record all food and drink intake, along with any physical activity undertaken, for which they were provided with food and activity record booklets and weighing scales for weighing food items. This was replicated in the 24 h prior to the remaining three trials. Participants were required to avoid strenuous physical activity, alcohol and keep caffeine intake at habitual levels during these 24-h periods. Participants arrived at the laboratory following a 10- to 12-h fast and body mass was measured. Participants rested in a supine position for 30 min before baseline measures of RMR, baseline breath samples for measurement of GE and subjective sensations of appetite on visual analogue scales (VAS) were taken, after which participants consumed the breakfast. Subjects rested in a supine position for either 4 h (Con-R/MCT-R) or 2 h and then cycled for 60 min at 65 %
 $\dot V$
O2peak and then rested for another 1 h (Con-Ex/MCT-Ex), while measures of GE, subjective sensations of hunger and appetite, and EE were taken. At 240 min, participants consumed an ad libitum lunch buffet, before final measurements were taken and participants left the laboratory. Participants were instructed to complete a weighed food diary for the remainder of the day and told to avoid strenuous exercise and alcohol. Finally, they were to complete a questionnaire on gastrointestinal symptoms upon waking the next day. Fig. 1 provides a schematic display of the experimental trials.
$\dot V$
O2peak and then rested for another 1 h (Con-Ex/MCT-Ex), while measures of GE, subjective sensations of hunger and appetite, and EE were taken. At 240 min, participants consumed an ad libitum lunch buffet, before final measurements were taken and participants left the laboratory. Participants were instructed to complete a weighed food diary for the remainder of the day and told to avoid strenuous exercise and alcohol. Finally, they were to complete a questionnaire on gastrointestinal symptoms upon waking the next day. Fig. 1 provides a schematic display of the experimental trials.
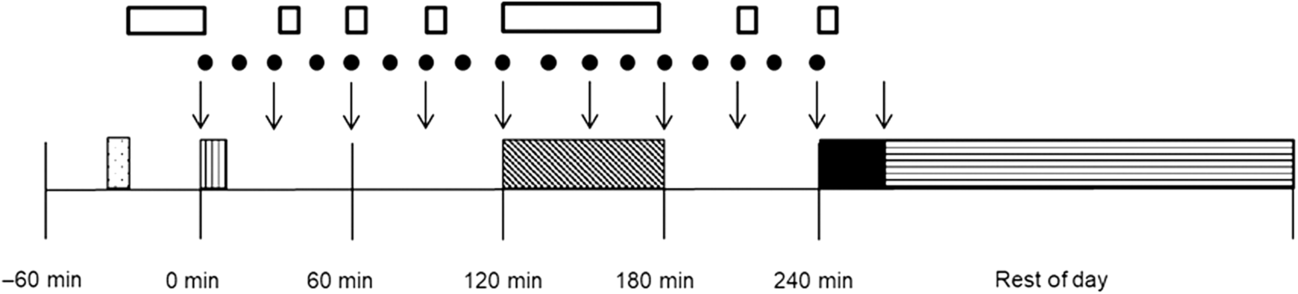
Fig. 1. Schematic representation of the study protocol. (![]() ), Supine rest; (
), Supine rest; (![]() ), breakfast; (
), breakfast; (![]() ), exercise; (
), exercise; (![]() ), ad libitum lunch; (
), ad libitum lunch; (![]() ), diet diary; (↓), visual analogue scale; (
), diet diary; (↓), visual analogue scale; (![]() ), gastric emptying breath sample; (
), gastric emptying breath sample; (![]() ), RMR/energy expenditure.
), RMR/energy expenditure.
Energy expenditure
The metabolic stress testing system was calibrated according to the manufacturer’s instructions prior to baseline samples using reference gases (Airgas Healthcare) and a 3-litre syringe (Hans Rudolph). Samples were taken using the metabolic stress testing system at 0 (baseline), 30, 60, 90, 120, 210 and 240 min. The sample taken at baseline was for 30 min, the sample at 120 min was continuous for 60 min and all other collections were for 15 min. The first 15, 10 and 5 min of each of these respective samples were discarded and the remainder were analysed. EE and substrate oxidation were calculated from expired air samples using stoichiometric equations(Reference Frayn27).
Gastric emptying
GE was measured by adding 100 mg 13C octanoic acid (Euriso-top) to the breakfast. Octanoic acid is a medium-chain fatty acid retained in food in the stomach but is rapidly absorbed in the duodenum and carried via the portal venous system to the liver where it is completely oxidised. After oxidation, where octanoic acid becomes labelled CO2, it is exhaled in the breath. Breath samples were collected by blowing gently into a 12-ml exetainer (Labco) through a drinking straw and immediately replacing the lid upon withdrawal of the straw, which occurred in duplicate every 15 min for 4 h. The rate of appearance of 13CO2 was measured using isotope ratio MS (ABCA; Sercon Ltd), and results were expressed relative to Vienna Pee Dee Belemnite, an international standard of known 13C abundance. The appearance of labelled CO2 in the breath was used to calculate the time to peak 13C enrichment (13C-TTP) as a marker of GE, according to Van Nieuwenhoven et al. (Reference Van Nieuwenhoven, Wagenmakers and Senden28).
Visual analogue scales
Participants rated their subjective sensations of hunger, fullness, desire to eat (DTE), prospective food consumption (PFC) and nausea using continuous 100 mm VAS, where 0 mm indicated ‘not at all’ and 100 mm indicated ‘extremely’. These were recorded at 0 (baseline), 30, 60, 90, 120, 210 and 240 min (pre-buffet) and 270 min (post-buffet). To test the palatability of the breakfast, participants also rated the appearance, aroma, flavour, pleasantness and texture on 100 mm VAS.
Test breakfast and ad libitum lunch meal
Participants were provided with a breakfast that acted as a vehicle for the oils immediately after baseline measurements were taken. The breakfast consisted of porridge, semi-skimmed milk, honey, the added lipid and 13C octanoic acid, providing 2112 kJ, 56·0 g carbohydrate, 12·7 g protein and 26·1/27·7 g fat (in control trials and MCT trials, respectively). The macronutrient composition of the meal was 44·4 % carbohydrate, 10·1 % protein and 45·5 % fat. The breakfast was designed to mask the flavour of both lipids. In Con-R and Con-Ex, 18·4 g of vegetable oil (Rapeseed oil; Tesco Stores Ltd) was added. In MCT-R and MCT-Ex, 20 g of MCT oil (50–60 % caprylic acid, 30–45 % capric acid, 3 % lauric acid and 2 % caproic acid; Muscleform) was added.
At 4 h, participants were provided with a multi-choice ad libitum meal consisting of pasta with meat-free Bolognese sauce and cheese, white bread, cooked meats, sliced cheese, bananas, apples, oranges, chocolate biscuits and fruit juices. Buffet foods were provided in identical amounts before each meal in excess of expected consumption, and food consumption was determined by weighing each food item before and after the meal. Participants were given 30 min to eat and could not leave early in order to resist the temptation of finishing the meal prior to genuine satiation. Energy and macronutrient content of the food items were ascertained from manufacturers packaging and information.
Statistical analysis
Data were analysed using SPSS version 25 software for windows (SPSS Inc.). AUC values for hunger, fullness, DTE, PFC and nausea v. time curves were calculated using the trapezoidal rule. An ANCOVA with the baseline used as a covariate was used to examine the differences in subjective sensations across the four trials. Two-way repeated-measure ANOVA were used to examine the differences between energy and macronutrient intakes at the ad libitum lunch, for the rest of the day, and over the 24-h period, as well for 13C-TTP. All data were subject to checks for normality using the Shapiro–Wilk test. Where appropriate, post hoc analyses were conducted using the Bonferroni adjustment. Results from ANOVA are reported with the F statistic, along with df and error in subscript parenthesis. Significance was accepted at the α level of 5 %. Data are expressed as means and standard deviations.
Sample size calculations
The sample size used in the present study was estimated to be sufficient to detect a significant difference in energy intake between conditions. The anticipated effect size was based on compiled data from Schubert et al.(Reference Schubert, Desbrow and Sabapathy29) examining energy intake and relative energy intake after exercise; and based on these data and an α level of 5 %, a sample of twelve participants would have more than 95 % power to detect a difference of 2894 kJ in relative energy intake. The data from Schubert include ad libitum intake after exercise, with a minimum of one and a maximum of four meals provided in studies after exercise; with relative energy intake calculated as the total energy intake minus the gross energy expenditure. G*Power(Reference Faul, Erdfelder and Lang30) was used to perform all calculations.
Results
Energy and macronutrient intake
There were no significant main effects or interactions for energy intake between the trials at the ad libitum lunch (all P ≥ 0·05), for the rest of the day (all P ≥ 0·05), or for total intake (all P ≥ 0·05) (Table 1). No differences were observed in carbohydrate (all P ≥ 0·05), fat (all P ≥ 0·05) or protein (all P ≥ 0·05) intakes at any of these time points (Table 1).
Table 1. Energy and macronutrient ingestion at the ad libitum lunch, during the remainder of the trial day (from weighed food records), and total intake over the entire day in all trials*
(Mean values and standard deviations)
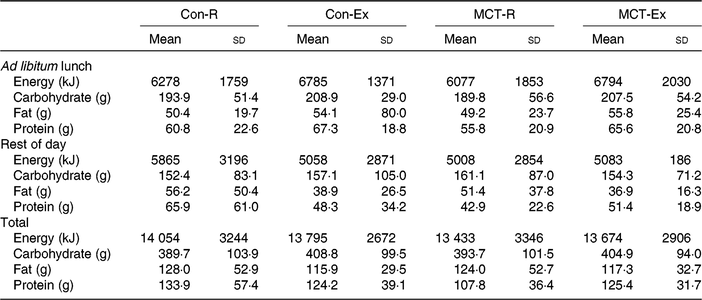
Con-R, resting trial with control oil; Con-Ex, exercise trial with control oil; MCT-R, resting trial with medium-chain TAG oil; MCT-Ex, exercise trial with medium-chain TAG oil.
* No significant differences were observed between trials in any aspect of intake.
Energy expenditure and substrate oxidation
EE increased in the postprandial period (0–2 h) for all trials, peaking at 30 min in the resting trials and at 120 min the exercise trials. The rate of postprandial EE changed over time (F 10,110 = 229·362, P < 0·001) and between exercise and rest (F 1,11 = 241·516, P < 0·001) but were not affected by MCT (F 1,11 = 1·361, P = 0·268; Fig. 2(a)). There was a significant difference in total EE between breakfasts (F 1,11 = 6·092, P = 0·031) and between conditions (F 1,11 = 227·875, P < 0·001), with greater energy expended over the 4-h testing period after MCT breakfasts (124·8 kJ, 95 % CI 13·5, 236·0) and during exercise trials 2960·6 kJ, 95 % CI 2528·9, 3392·2; Fig. 2(b)).
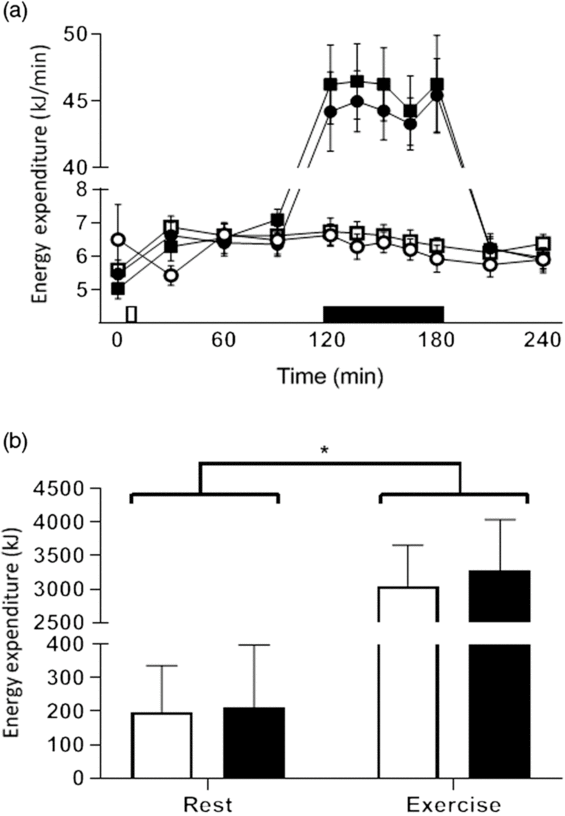
Fig. 2. Postprandial change in energy expenditure (a). On the x axis, the white rectangle indicates the test breakfast and the black rectangle indicates the hour of rest/exercise (in resting trial with control oil (Con-R)/resting trial with medium-chain TAG oil (MCT-R) and exercise trial with control oil (Con-Ex)/exercise trial with medium-chain TAG oil (MCT-Ex), respectively). Total energy expenditure (b) in all trials. Data are means, with vertical bars indicating standard errors of the mean (for clarity; a) and standard deviations (b). (a) (![]() ), Con-R; (
), Con-R; (![]() ), Con-Ex; (
), Con-Ex; (![]() ), MCT-R; (
), MCT-R; (![]() ), MCT-Ex. (b) (
), MCT-Ex. (b) (![]() ), Control; (
), Control; (![]() ), medium-chain TAG. * Significant at P < 0·05.
), medium-chain TAG. * Significant at P < 0·05.
No effect of breakfast was observed on carbohydrate oxidation (F 1,11 = 1·367, P = 0·267), but greater carbohydrate oxidation was observed in exercise trials (11·2 g/min, 95 % CI 9·9, 32·3; F 1,11 = 89·095, P < 0·001). No interaction of breakfast and condition was observed (F 1,11 = 0·769, P = 0·399). Fat oxidation was not affected by condition (F 1,11 = 0·183, P = 0·980); however, there was a trend for greater fat oxidation after MCT breakfasts (7·9 g/min, 95 % CI −1·1, 16·8, F 1,11 = 3·765, P = 0·078). No interaction of breakfast and condition was observed (F 1,11 = 2·014, P < 0·184).
Energy balance
After accounting for energy expended during the trials by both rest and exercise, participants remained in an energy-deficit state in both exercise trials compared with the resting trials (−2393·0 kJ, 95 % CI −3635·6, −1150·4, F 1,11 = 17·967, P = 0·001), but no effect of breakfast was observed on energy balance (Con-R: 6091 + 1781 kJ; MCT-R: 5866 + 1818 kJ; Con-Ex: 3739 + 1453 kJ; MCT-Ex: 3432 + 2281 kJ; F 1,11 = 0·797, P > 0·05), and no interaction of breakfast and condition was observed (F 1,11 = 0·19, P > 0·05; Fig. 3). Inspection of the data showed that seven of the twelve participants achieved an energy deficit in MCT-R compared with CON-R, and seven of the twelve participants achieved an energy deficit in MCT-Ex compared with CON-Ex. Energy deficits were all lower in both exercise trials compared with all resting trials.
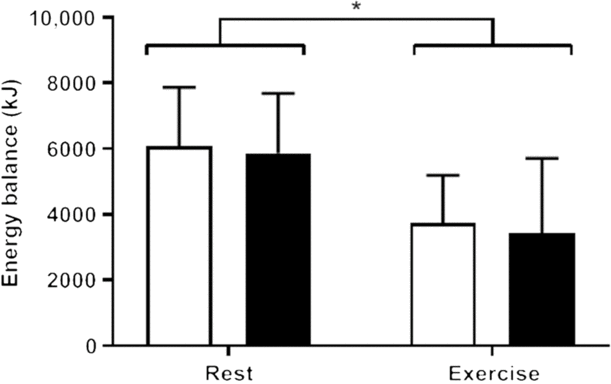
Fig. 3. Energy balance during each trial calculated as energy intake (from the test breakfast and ad libitum lunch) minus energy expenditure. (![]() ), Control breakfast; (
), Control breakfast; (![]() ), medium-chain TAG breakfast. Data are means, with vertical bars indicating standard deviations. * Significant at P < 0·05.
), medium-chain TAG breakfast. Data are means, with vertical bars indicating standard deviations. * Significant at P < 0·05.
Subjective sensations of appetite
No differences were observed in any of the calculated AUC for any of the VAS scores (all P ≥ 0·668) and no effect on ratings of nausea (F 3,43 = 0·952, P = 0·424). In all tests, feelings of satiety increased sharply after consumption of the breakfast and gradually decreased until the ad libitum lunch. Exercise did not affect this pattern (Fig. 4).
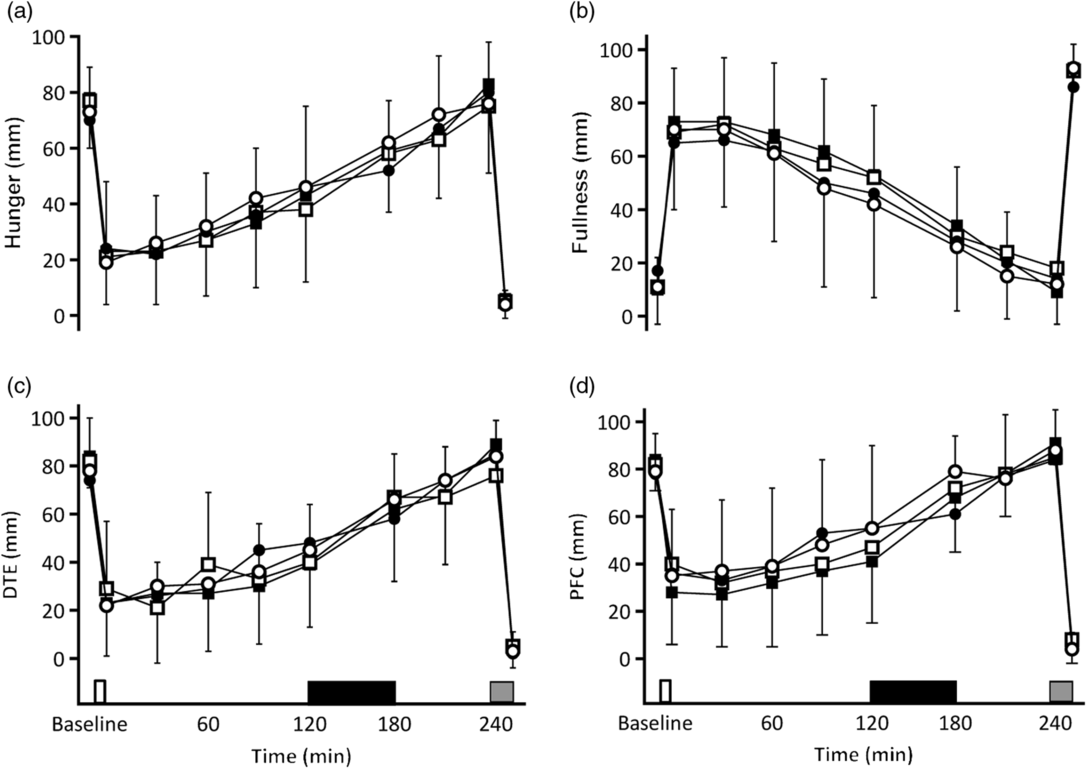
Fig. 4. Subjective sensations of hunger (a), fullness (b), desire to eat (DTE) (c) and prospective food consumption (PFC) (d). On the x axis, the white rectangle indicates the test breakfast, the grey rectangle indicates the ad libitum meal, and the black rectangle indicates the hour of rest/exercise (in resting trial with control oil (Con-R)/resting trial with medium-chain TAG oil (MCT-R) and exercise trial with control oil (Con-Ex)/exercise trial with medium-chain TAG oil (MCT-Ex), respectively). (![]() ), Con-R; (
), Con-R; (![]() ), MCT-R; (
), MCT-R; (![]() ), Con-Ex; (
), Con-Ex; (![]() ), MCT-Ex. Data are means, with vertical bars indicating standard deviations.
), MCT-Ex. Data are means, with vertical bars indicating standard deviations.
Gastric emptying
Exercise led to shorter 13C-TTP, regardless of the breakfast consumed (Con-Ex: 70·9 (SD 10·2) min; MCT-Ex: 85·8 (SD 13·2) min) compared with the resting trials (Con-R: 116·6 (SD 24·6) min; MCT-R: 149·0 (SD 28·0) min; F 1,11 = 137·299, P < 0·001). MCT also delayed 13C-TTP compared with CON (F 1,11 = 24·780, P < 0·001), but no interaction of breakfast and condition was observed (F 1,11 = 3·000, P > 0·05).
Palatability
No difference in appearance, aroma, flavour, pleasantness, or texture (all P > 0·05) was observed between any of the trials.
Discussion
The primary aim of the present study was to examine the combined effect of MCT and aerobic exercise on satiety and the overall energy balance. It was found that although exercise resulted in approximately 2400–3100 kJ deficit compared with resting conditions, no cumulative effect of exercise and MCT was observed. Furthermore, neither MCT nor exercise affected appetite sensations or energy intake. From a weight management perspective, the substitution of MCT for LCT could be used as a viable method for increasing EE in the absence of moderate to high physical activity. However, as there was no dietary compensation after exercise, exercise can be used to produce greater deficits in energy balance. In line with the previous findings examining GE(Reference Clegg, Pratt and Markey20), MCT led to delayed 13C-TTP in rest and exercise conditions, and exercise promoted GE and led to the fastest 13C-TTP.
Increased feelings of hunger have been cited as a practical reason as to why dietary strategies may be difficult to adhere to or are unsuccessful(Reference King, Wasse and Ewens11). Foods with enhanced satiety have been outlined as having benefits to weight management(Reference Chambers, McCrickerd and Yeomans31). MCT have previously been shown to increase satiety and decrease energy intake when compared with ‘traditional’ LCT(Reference Van Wymelbeke, Himaya and Louis-Sylvestre2–Reference Rolls, Gnizak and Summerfelt4,Reference Kinsella, Maher and Clegg32) , though these are not unequivocal findings(Reference Poppitt, Strik and MacGibbon33,Reference Rizzo, Masic and Harrold34) . The apparent increase in satiety is possibly due to their fast absorption, as MCT are transported directly to the liver via the portal vein; avoiding chylomicron transport in the lymphatic system(Reference Bloom, Chaikoff and Reinhardt35). This rapid absorption, by avoiding deposition into adipose tissue(Reference Bach and Babayan8), means they are preferentially utilised as an available fuel source. This readily available fuel source may initiate a glucose-sparing effect due to the production of ketone bodies(Reference Zhang, Kuang and Xu36), which is thought to be appetite suppressing(Reference Mayer37). MCT have also been shown to increase EE through DIT(Reference Clegg, Golsorkhi and Henry6), again due to the rapid metabolism of MCT. MCT also achieve greater DIT through activation of the sympathetic nervous system as evidenced by increased urinary noradrenaline(Reference Dulloo, Fathi and Mensi38). Clegg et al.(Reference Clegg, Golsorkhi and Henry6) found MCT resulted in 51 % greater DIT than LCT (calculated as an increase in EE as a percentage of the test meal energy), and Dulloo et al.(Reference Dulloo, Fathi and Mensi38) reported 5 % increased 24-h EE after MCT ingestion. Similarly, Scalfi et al.(Reference Scalfi, Coltorti and Contaldo39) and Flatt et al.(Reference Flatt, Ravussin and Acheson40) reported daily EE increases of about 119 kJ and 439 kJ after MCT, respectively. In the study by Clegg et al.(Reference Clegg, Golsorkhi and Henry6), DIT was calculated as the increase in EE as a percentage of the test meal energy. This was not calculated in this case as it was not relevant for the exercise trials and would be confusing to interpret. Although the amount of MCT given in the present study was lower than those given by those two early studies(Reference Scalfi, Coltorti and Contaldo39,Reference Flatt, Ravussin and Acheson40) , we still report a significant effect of MCT on EE over the 4-h postprandial period.
Exercise has also been shown to induce an energy deficit, due to the large amount of energy expended which is not fully compensated for by food intake(Reference King, Wasse and Ewens11). The meta-analysis by Schubert et al.(Reference Schubert, Desbrow and Sabapathy29) confirms this, as they report no change in energy intake after exercise. King et al.(Reference King, Wasse and Ewens11) showed that similar (approximately 4800 kJ) deficits created by exercise or energy restriction resulted in different responses, as restriction led to compensatory mechanisms whereas exercise did not. This was due to an increase in the hunger-promoting hormone acylated ghrelin and a decrease in the satiating PYY3-36. Ghrelin stimulates hunger through activation of the lateral hypothalamus via the vagal nerve(Reference Date, Murakami and Toshinai41) by producing neuropeptide Y and agouti-related peptide, which are potent orexigenic peptides(Reference Chen, Trumbauer and Chen42). However, only the acylated form of ghrelin exerts its effects, as this form binds to the growth hormone secretagogue receptor type 1a. Acylated ghrelin has been shown to be suppressed after aerobic exercise(Reference Broom, Stensel and Bishop12); however, this has more recently been shown to be only with higher intensity exercise(Reference Broom, Miyashita and Wasse43). The exercise protocol in the present study, therefore, is not likely to have affected acylated ghrelin, which would explain, in part, the lack of effect on appetite sensations or energy intake. Exercise and MCT apparently create energy deficits through different physiological mechanisms, but the results of the present study show that these are not synergistic and therefore do not lead to a greater, cumulative, energy deficit.
GE in the present study was delayed with MCT, which corroborates previous research(Reference Clegg, Pratt and Markey20). Studies have reported that MCT accelerates GE(Reference Beckers, Jeukendrup and Brouns44), which is in line with the findings that GE is delayed by LCT and not MCT(Reference Feltrin, Little and Meyer19). However, Clegg et al.(Reference Clegg, Pratt and Markey20) point out that few studies consider the higher osmolarity of MCT which may delay GE, even in the absence of gut hormone-mediated GE. Exercise of moderate (45–70 %) intensities accelerate GE, whereas higher intensities delay it(Reference Horner, Schubert and Desbrow21). The increase in gastric motility is dependent on vagal nerve activity, as demonstrated by the lack of increase in vagotomised rats(Reference Wang, Kondo and Suzukamo45). The lack of appetite or energy intake response in the present study may have been influenced by the increased rate of GE, as delayed GE has been linked to increased satiety(Reference Hunt16). It is possible that higher intensities of exercise may delay GE, which may yield a satiety effect. Further work should aim to examine whether the delaying of GE by MCT synergises with the delaying of GE stimulated by higher intensity exercise and what implications this has for energy intake.
Neither breakfast nor exercise condition had any statistically significant effect on appetite, though there are trends for increased fullness after the MCT breakfast. The present study utilised a relatively small sample size, and it is possible this was not enough to detect differences in subjective sensations of appetite, though our sample size was similar to other studies that report a satiating effect of MCT(Reference Van Wymelbeke, Himaya and Louis-Sylvestre2–Reference Rolls, Gnizak and Summerfelt4). Similar to Clegg et al. (Reference Clegg, Golsorkhi and Henry6), the breakfast in the present study was relatively large (1476·9 kJ) in comparison with the dosage of MCT (694·5 kJ), compared with other studies that used a smaller breakfast vehicle(Reference Kinsella, Maher and Clegg32) or larger MCT dose(Reference Van Wymelbeke, Himaya and Louis-Sylvestre2). The optimal MCT bolus for enhancing satiety has yet to be elucidated; however, it has previously been shown that MCT decrease intake without affecting appetite sensations(Reference Coleman, Quinn and Clegg5). While there was no difference in palatability scores between the breakfasts, scores were generally low for all of the variables of all trials (e.g. flavour scores were 51 (SD 35) mm, 51 (SD 35) mm, 55 (SD 30) mm and 51 (SD 29) mm, for Con-Ex, Con-R, MCT-Ex and MCT-R, respectively). This indicates that the breakfasts were generally not liked, which may have interfered with the satiety scores(Reference Clegg46). Future studies should utilise more palatable vehicles for the comparison of LCT and MCT.
The present study has several limitations. The sample size in the present study was estimated to be sufficient to detect differences in energy intake based on data from exercise studies; however, this may not have been sufficient to detect subtler changes in energy intake induced by MCT. The present study utilised recreationally active, healthy young males, and so these findings cannot be applied to other populations such as overweight/obese populations where decrease in food intake may be more relevant and GE and appetite regulation have been shown to be altered(Reference Verdich, Lysga and Madsen47). As discussed, higher intensities of exercise may affect GE and appetite to a greater extent than the moderate exercise used in the present study. From a practical standpoint, 1 h of higher intensity exercise may not be achievable by individuals engaging in weight management strategies; thus, future studies should also aim to examine the combination of lower (<45 %) intensity exercise along with MCT. This may help to elucidate whether MCT solely increase EE at rest, or whether the combination of MCT with lower intensity exercise can also increase EE. Despite allowing for a degree of external validity and reducing the burden to the participant, food records are well known to be subject to participant bias, which may result in underreporting of intake(Reference Livingstone and Black48). To produce more reliable results, future work could consider including all meals under controlled laboratory conditions to confirm these findings. Finally, the accuracy and reliability of the CPX ultima for measurement of RMR have been questioned, as it has been reported to yield significantly different results and not correlate to the Deltatrac Metabolic Monitor(Reference Cooper, Watras and O’Brien49), an indirect calorimeter that has shown to be valid and reliable for RMR. However, the CPX ultima, unlike the Deltatrac, is designed for use in exercise and as such they are difficult to compare. Our own pilot testing indicated a within-subject variance of 6·9 %, which is better than that reported in Cooper et al.(Reference Cooper, Watras and O’Brien49); and as all comparisons were completed within individual, we believe it to be reliable for the purposes of the present study.
MCT and MCT-based products have gained popularity in recent years, and there is some evidence that these can be used in weight management strategies. By enhancing satiety, there is potential to decrease energy intake and thus increase adherence to dietary manipulation that aims to achieve an energy deficit. Another method of creating this deficit is to increase EE, either by exercise or by products that increase DIT. This is the first study to date to examine the potential combination of MCT with simultaneous exercise to produce greater energy deficits through a cumulative increased satiety and EE. MCT increase EE through increased DIT, and exercise results in energy deficits due to the lack of dietary compensatory mechanisms. Combining MCT and exercise does not appear to be of any benefit, and thus MCT should be used in periods of low activity in order to increase EE.
Acknowledgements
T. M. was supported by an Oxford Brookes University Nigel Groome Scholarship, which also funded the present study. This research received no external funding.
T. M. and M. C. conceived the study. All authors contributed to the study design. T. M., A. E. C. and M. C. undertook the data collection. All authors contributed to data analysis. T. M. and M. C. wrote the manuscript. All authors read and approved the manuscript.
There are no conflicts of interest.






 Open access
Open access