


Dendritic cells (DC) are bone marrow-derived antigen-presenting cells capable of inducing protective adaptive immune responses or tolerance. DC are equipped with Toll-like receptors (TLR) and C-type lectin receptors that activate different signalling pathways in response to micro-organisms( Reference van Vliet, den Dunnen and Gringhuis 1 ). TLR expressed on the cell surface predominantly bind to bacterial products, such as lipopeptides and peptidoglycan (TLR-1, -2 and -6), lipopolysaccharide (LPS, ligand of TLR-4) and flagellin (TLR-5). TLR-3, -7, -8 and -9 reside in intracellular organelles and recognise microbial nucleic acids( Reference Takahashi, Yasuhara and Shingo 2 ). In the intestinal mucosa, various subsets of DC are in close contact with the intestinal microbiota and continuously migrate from the lamina propria to the mesenteric lymph nodes, ensuring a balance between immunity and tolerance( Reference Milling, Yrlid and Cerovic 3 ). Lamina propria DC can be divided into two major classes: CD103− DC, which induce T helper (Th)1 and Th17 responses, and CD103+ DC, which induce regulatory T cells( Reference Rescigno and Di Sabatino 4 ). Different types of DC develop from bone marrow progenitors and therefore immature bone marrow-derived DC (BM-DC) are widely used in in vitro studies. Such studies contribute to a better understanding of the mechanism by which micro-organisms modulate immune responses in vivo. Although not identical to intestinal DC, it has been demonstrated that splenic DC from Th1-prone C57BL/6 mice and Th2-prone BALB/c mice respond differently to microbial stimuli( Reference Mirkov, Stojanovic and Glamoclija 5 – Reference Sacks and Noben-Trauth 7 ), and this may in part be explained by differences in the expression of TLR. BALB/c mice express higher levels of Tlr2, Tlr4 and Tlr5 mRNA, whereas C57BL/6 mice express higher levels of Tlr9 mRNA( Reference Liu, Matsuguchi and Tsuboi 8 ). In addition, the expression of the macrophage mannose receptor on BM-DC is mouse-strain specific, and this phenomenon may influence antigen uptake by such cells( Reference Flacher, Douillard and Ait-Yahia 9 ).
Probiotic bacteria modulate DC via surface expression and/or secretion of products that function as ligands for TLR and C-type lectin receptors. The expression of these ligands depends on bacterial species, growth phases and available nutrients, and this explains in part why different species and strains of probiotics vary in their ability to induce cytokines and chemokines( Reference Kim, Park and Ji 10 – Reference Mohamadzadeh, Olson and Kalina 12 ).
We have recently demonstrated that treatment of BALB/c mice with a mixture of probiotic bacteria (i.e. VSL#3) has profound effects on gene expression in the colon of mice subjected to colitis induction, with favourable effects on the development of disease( Reference Mariman, Kremer and van Erk 13 ). However, substantial variability in the response of human subjects to probiotic interventions has been found( Reference Reid, Gaudier and Guarner 14 ), suggestive of an influence of genetic and environmental factors.
To gain more insights into the influence of genetic background, in the present study, we examined BM-DC from two genetically distinct mouse strains and evaluated their responses to the probiotic mixture VSL#3 in vitro compared with those to ultrapure TLR ligands. The results of the present study, obtained by dedicated gene expression profiling of genes related to TLR signalling and by studying cytokine production, indicate that VSL#3 may display both pro- and anti-inflammatory effects, dependent on the mouse strain.
Materials and methods
Mice
C57BL/6 mice (Charles River Laboratories) and BALB/c mice (Janvier) aged 7–11 weeks were used in the present study. All animal experiments were conducted with the approval of the Institutional Animal Welfare Committee, filed as no. DEC2661, in compliance with European Community specifications regarding the use of laboratory animals.
Isolation and culture of bone marrow-derived dendritic cells
BM-DC were isolated as described previously( Reference Inaba, Inaba and Romani 15 ), with slight modifications. Briefly, bone marrow was flushed from femur and tibia. Cells were passed through nylon mesh to obtain a single cell suspension. After a single wash step, cells were cultured in Roswell Park Memorial Institute medium 1640 containing 10 % fetal bovine serum (Lonza), 2 mm–l-glutamine, 100 U/ml streptomycin, 100 μg/ml penicillin (Gibco) and 50 μm–β-mercaptoethanol (Sigma-Aldrich) in the presence of 20 ng/ml of recombinant mouse granulocyte-macrophage colony-stimulating factor (PeproTech, Inc.). Cells were cultured (106 cells/ml) at 37°C. On days 3 and 6 of culture, two-thirds of the medium were refreshed. After 8 d of culture, non-adherent and loosely adherent BM-DC were collected and used for stimulation experiments. The viability of cells (>95 %) was assessed by trypan blue dye exclusion. Cells were cultured at a density of 106/ml and stimulated with TLR ligands and/or probiotic bacteria at concentrations indicated in the figure legends.
Reagents
Pam3Cys-Ser-(Lys)4 trihydrochloride (Pam3CSK4), polyinosinic:polycytidylic acid (poly I:C), LPS from E. coli K12, flagellin from S. typhimurium, CpGODN1826, and imiquimod and peptidoglycan from E. coli 0111:B4 were purchased from InvivoGen. All these ultrapure TLR ligands were endotoxin free ( < 0·001 endotoxin units/μg), except peptidoglycan, which contained < 125 endotoxin units/ml.
VSL#3, a mixture containing freeze-dried Bifidobacterium longum, Bifidobacterium breve, Bifidobacterium infantis, Lactobacillus acidophilus, Lactobacillus plantarum, Lactobacillus casei, Lactobacillus bulgaricus and Streptococcus thermophilus, was purchased from Ferring Pharmaceuticals. L. plantarum NCIMB8826 and Bifidobacterium animalis ssp. lactis BB-82 were grown at 37°C in Man–Rogosa–Sharpe broth (Scharlau Chemie) until mid-exponential density (OD600 nm= 1). Bacterial cells were washed twice with PBS (pH 7·4). Lyophilised bacteria were prepared by freezing bacterial pellets ( − 80°C) before overnight lyophilisation in a freeze dryer under vacuum (40 mbar). Lyophilised bacteria were stored at − 20°C until use.
RNA isolation and real-time quantitative PCR
Total RNA was isolated from 106 cells using an RNeasy Kit (Qiagen) according to the manufacturer's instructions. RNA (500 ng) was reverse-transcribed into single-stranded complementary DNA using the High-Capacity RNA-to-cDNA Kit (Applied Biosystems) and incubated for 60 min at 37°C and 5 min at 95°C. Complementary DNA was amplified employing a 7500 Fast thermal cycler (Applied Biosystems) using SYBR Green PCR Master Mix (Applied Biosystems). Thermal cycling parameters consisted of 1 min at 50°C and 10 min at 95°C, followed by forty cycles of 15 s at 95°C and 1 min at 60°C. The following primer pairs were used: IL-12 p35 subunit (Il12p35) – 5′-CTG GAG CAT CCG AAT TGC A-3′ (sense) and 5′-CAT CCT CTG AGA TTT GAC GCT TT-3′ (anti-sense); IL-23 p19 subunit (Il23p19) – 5′-GGC AAC TTG GAC CTG AGG AG-3′ (sense) and 5′-CAT GGG CTC TCG GTC CAT AG-3′ (anti-sense); chemokine (C–X–C motif) ligand 9 (Cxcl9) – 5′-CCT AGT GAT AAG GAA TGC ACG ATG-3′ (sense) and 5′-CTA GGC AGG TTT GAT CTC CGT TC-3′ (anti-sense); Chemokine (C–X–C motif) ligand 10 (Cxcl10) – 5′-ATC ATC CCT GCG AGC CTA TCC T-3′ (sense) and 5′-GAC CTT TTT TGG CTA AAC GCT TTC-3′ (anti-sense); β-2-microglobulin (B2m) – 5′-ACC GTG AAA AGA TGA TGA CCC AG-3′ (sense) and 5′-AGC CTG GAT GGC TAC GTA CA-3′ (anti-sense); IL-12 p40 subunit (Il12p40) – 5′-GGA AGC ACG GCA GCA GAA TA-3′ (sense) and 5′-AAC TTG AGG GAG AAG TAG GAA TGG-3′ (anti-sense). Gene expression levels were quantified according to the following formula:
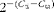 $$2^{ - ( C _{ti} - C _{ta})} $$
, where C
ti is the cycle threshold value of the gene of interest and C
ta is the cycle threshold value of β-actin.
$$2^{ - ( C _{ti} - C _{ta})} $$
, where C
ti is the cycle threshold value of the gene of interest and C
ta is the cycle threshold value of β-actin.
PCR array analysis
The expression of genes involved in TLR pathways was studied using the RT2-Profiler PCR Array (mouse TLR signalling pathway) obtained from SABiosciences. This array combines the quantitative performance of SYBR Green-based real-time PCR with the multiple gene profiling capabilities of a microarray. In this analysis, ninety-six-well plates containing gene-specific primer sets for eighty-four relevant TLR pathway genes, five housekeeping genes and two negative controls were used. For each experimental condition, RNA was isolated as described above. Gene expression was normalised to internal controls (housekeeping genes) to determine the fold change in gene expression between test and control samples by ΔΔC t (SABioscience).
Cytokine/chemokine analysis
Cytokines present in the supernatants of BM-DC cultures were quantified 24 h after cell stimulation using commercially available ELISA kits for IL-12 p70 subunit (IL-12p70), IL-12p40, IL-23, IL-10, IL-6 and TNFα (eBioscience). Chemokines (CXCL-9 and CXCL-10) were quantified using a multiplex immunoassay (Invitrogen). Assays were carried out according to the manufacturer's instructions.
Flow cytometric analyses
BM-DC were phenotypically characterised before and after 24 h of stimulation. Cells were collected and washed with PBS. Later, 2 × 105 cells were incubated (30 min, 4°C, protected from light) with monoclonal antibodies. The following dye-conjugated antibodies were purchased from BD Biosciences: CD11c-PerCP/Cy5.5 (N418); CD86-FITC (B7-2GL-1); CD80-Pacific Blue (16-10A1); major histocompatibility complex-phycoerythrin (MHCII-PE) (M5/114.15.2); CD317-allophycocyanin (APC) (927); CD103-PE (2E7). Flow cytometric analyses were carried out using a fluorescence-activated cell sorter (FACSCanto II; BD Biosciences) and FACSDiva software 6.1.2 (BD Biosciences). Gating of positive cells was based on the results obtained with isotype control antibodies.
Statistical analyses
Statistical analyses were carried out using the Mann–Whitney U test or Student's t test if the samples passed the normality tests using RT2 Profiler™ PCR Array Data Analysis (SABiosciences).
Results
Bone marrow progenitors from BALB/c and C57BL/6 mice differentiate into phenotypically distinct dendritic cells
BM-DC from BALB/c and C57BL/6 mice were generated by 8 d of culture under identical conditions in the presence of granulocyte-macrophage colony-stimulating factor. As shown in Fig. 1(a)–(d), BM-DC from BALB/c mice exhibited higher levels of CD103 and CD317 when compared with BM-DC from C57BL/6 mice. Moreover, gene expression profiling of transcripts involved in TLR signalling revealed a higher expression of Tlr3 and interferon (Ifn)b in BM-DC from BALB/c mice (Fig. 1(e)). BM-DC from BALB/c mice also exhibited significantly higher levels of Tlr5, Tlr6 and Tlr9 mRNA when compared with BM-DC from C57BL/6 mice (Fig. 1(f)).

Fig. 1 Phenotypic differences between bone marrow-derived dendritic cells (BM-DC) from C57BL/6 mice and those from BALB/c mice revealed by flow cytometric analysis and gene expression profiling. (a–d) Immature BM-DC generated in the presence of granulocyte-macrophage colony-stimulating factor were stained with phycoerythrin (PE)-labelled anti-CD103 and allophycocyanin (APC)-labelled anti-CD317 and evaluated by flow cytometric analysis. (a) C57BL/6 – 48·6 %; (b) BALB/c – 74·3 %; (c) C57BL/6 – 16·7 %; (d) BALB/c – 64·6 %. Filled histograms show the fluorescence intensity for each marker. Dotted lines indicate staining with isotype control antibodies. The percentage of cells expressing these markers is indicated in each panel. (e) mRNA isolated from immature BM-DC was used to evaluate the expression of genes involved in Toll-like receptor (TLR) signalling. Results (i.e. an algorithm expressing C
t of the gene of interest relative to the C
t of the housekeeping gene) for BM-DC from C57BL/6 mice are plotted against the results for BM-DC from BALB/c mice. Genes that exhibited at least a 3-fold difference in expression between BM-DC from C57BL/6 mice and those from BALB/c mice are indicated. Hspa1a, heat shock 70 kDa protein 1A; Nfkb2, nuclear factor of kappa light polypeptide gene enhancer in B-cells 2 (p49/p100); Tlr3, Toll-like receptor 3; Ptgs2, prostaglandin-endoperoxide synthase 2; IFNβ, interferon β. (f) Differences in Tlr gene expression between BM-DC from C57BL/6 mice (![]() ) and those from BALB/c mice (
) and those from BALB/c mice (![]() ) were evaluated. mRNA isolated from immature BM-DC was amplified by quantitative real-time PCR and C
t values for each transcript were normalised to a panel of six housekeeping genes. These normalised C
t values were inversely correlated with the level of mRNA expression. Data are normalised C
t values from BM-DC from three individual mice, with standard deviations represented by vertical bars. Mean C
t value was significantly different between BM-DC derived from the two mouse strains: * P< 0·05; *** P< 0·001 (Student's t test).
) were evaluated. mRNA isolated from immature BM-DC was amplified by quantitative real-time PCR and C
t values for each transcript were normalised to a panel of six housekeeping genes. These normalised C
t values were inversely correlated with the level of mRNA expression. Data are normalised C
t values from BM-DC from three individual mice, with standard deviations represented by vertical bars. Mean C
t value was significantly different between BM-DC derived from the two mouse strains: * P< 0·05; *** P< 0·001 (Student's t test).
Before studying the response of these cells to probiotic bacteria, we evaluated their response to ultrapure TLR ligands. Flow cytometric analysis of the stimulated BM-DC confirmed a similar up-regulation of CD80, CD86 and MHC class II expression, regardless of the mouse strain or TLR ligand (data not shown). However, BM-DC from BALB/c and C57BL/6 mice exhibited a different cytokine response to several TLR ligands, as shown in Fig. S1 (available online). In particular, the production of IL-12p70 in response to LPS or CpG was higher in BM-DC from C57BL/6 mice. These data indicate that the genetic background strongly influences the phenotype and response of BM-DC to pure TLR ligands.
The probiotic mixture VSL#3 and lipopolysaccharide synergistically induce IL-12p70 and IL-23 production in bone marrow-derived dendritic cells from C57BL/6mice
Probiotic bacteria are far more complex than ultrapure TLR ligands and therefore we investigated to what extent the genetic background of BM-DC would influence their response to VSL#3.
We stimulated BM-DC with increasing numbers of bacteria, in the absence or presence of LPS. In Fig. 2(a)–(c), the production of TNFα, IL-6 and IL-12p70 upon stimulation of BM-DC from C57BL/6 mice is shown. The production of all these cytokines was stimulated by VSL#3 in a dose-dependent manner. Simultaneously, VSL#3 strongly induced the production of IL-10 (Fig. S1, available online). The combination of VSL#3 and LPS exerted an additive effect on the production of all these cytokines. However, at a VSL#3:DC ratio of 10:1, a synergistic induction of IL-12p70 production was observed.

Fig. 2 Differences in the responses of bone marrow-derived dendritic cells (BM-DC) from C57BL/6 and BALB/c mice to probiotic bacteria and pure Toll-like receptor (TLR) agonists. Production of (a) TNFα, (b) IL-6 and (c) IL-12 p70 subunit (IL-12p70) by BM-DC from C57BL/6 mice, stimulated with various concentrations of the probiotic VSL#3 (colony-forming units (CFU)/ml) in the absence or presence of 1 μg/ml lipopolysaccharide (LPS), was measured. Synergistic induction of (d) IL-12p70 and (e) IL-23 production depends on the genetic background of the bone marrow donor mouse. IL-12p70 and IL-23 production in BM-DC from C57BL/6 mice (![]() ) or BALB/c mice (
) or BALB/c mice (![]() ) stimulated with bacteria, LPS, or a combination of both is shown as a percentage of cytokine levels secreted in response to LPS alone (C57BL/6: 138 pg/ml IL-12p70 and 117 pg/ml IL-23; BALB/c: 14 pg/ml IL-12p70 and 258 pg/ml IL-23). Data are representative of two to three independent experiments. Values are means of three mice per group, with standard deviations represented by vertical bars. * For C57BL/6 mice, mean value was significantly different from those for VSL#3 only, LPS only and Lactobacillus plantarum only (P< 0·05; Mann–Whitney U test). Relative expression of (f) IL-12 p35 subunit (Il12p35) and (g) IL-23 p19 subunit (Il23p19) genes in BM-DC from C57BL/6 mice was measured after stimulation with LPS (1 μg/ml;
) stimulated with bacteria, LPS, or a combination of both is shown as a percentage of cytokine levels secreted in response to LPS alone (C57BL/6: 138 pg/ml IL-12p70 and 117 pg/ml IL-23; BALB/c: 14 pg/ml IL-12p70 and 258 pg/ml IL-23). Data are representative of two to three independent experiments. Values are means of three mice per group, with standard deviations represented by vertical bars. * For C57BL/6 mice, mean value was significantly different from those for VSL#3 only, LPS only and Lactobacillus plantarum only (P< 0·05; Mann–Whitney U test). Relative expression of (f) IL-12 p35 subunit (Il12p35) and (g) IL-23 p19 subunit (Il23p19) genes in BM-DC from C57BL/6 mice was measured after stimulation with LPS (1 μg/ml; ![]() ), CpG (5 μg/ml;
), CpG (5 μg/ml; ![]() ), Pam3Cys-Ser-(Lys)4 trihydrochloride (Pam3CSK4) (100 ng/ml;
), Pam3Cys-Ser-(Lys)4 trihydrochloride (Pam3CSK4) (100 ng/ml; ![]() ) and polyinosinic:polycytidylic acid (poly I:C) (50 ng/ml;
) and polyinosinic:polycytidylic acid (poly I:C) (50 ng/ml; ![]() ). Results are expressed as a percentage of mRNA expression, induced by stimulation with VSL#3 alone. Values are means of three individual mice, with standard deviations represented by vertical bars. * Synergistic induction was observed only with the combination of VSL#3 and LPS (P< 0·05; Mann–Whitney U test).
). Results are expressed as a percentage of mRNA expression, induced by stimulation with VSL#3 alone. Values are means of three individual mice, with standard deviations represented by vertical bars. * Synergistic induction was observed only with the combination of VSL#3 and LPS (P< 0·05; Mann–Whitney U test).
BM-DC from C57BL/6 mice exhibited a synergistic increase in IL-12p70 and IL-23 production in response to VSL#3 and LPS, whereas such an effect was not observed in BM-DC from BALB/c mice (Fig. 2(d) and (e)). This difference in the response of BM-DC from C57BL/6 and BALB/c mice was confirmed for IL-12p70 with L. plantarum, one of the constituents of VSL#3 (Fig. 2(d) and (e)).
Recently, it has been shown that TIR-domain-containing adapter-inducing interferon-β (TRIF)- and myeloid differentiation primary response gene (88) (MyD88)-dependent TLR ligands act in synergy to induce the release of IL-12p70 in BM-DC( Reference Krummen, Balkow and Shen 16 ). However, VSL#3 did not exhibit synergy with the MyD88-coupled TLR ligands CpG and Pam3CSK4 or the TRIF-coupled TLR ligand poly I:C with regard to the induction of Il12p35 and Il23p19 mRNA expression (Fig. 2(f) and (g)).
Synergy in the induction of IL-12p70 and IL-23 production was hence specific for the combination of VSL#3 and LPS and was only observed in BM-DC from C57BL/6 mice.
TLR gene array analysis of bone marrow-derived dendritic cells identifies the stimulating and blocking activities of VSL#3
To identify mechanisms by which probiotic bacteria may modulate the innate immune response, we analysed the mRNA expression of eighty-four genes involved in TLR signalling. A time course study revealed a peak in the levels of Il12p35, Il12p40 and Il23p19 mRNA after 4 h of stimulation with VSL#3 and LPS (data not shown). Therefore, RNA was isolated from BM-DC after 4 h of culture with or without LPS, VSL#3, or a combination of both. In Fig. 3, the relative gene expression in BM-DC from C57BL/6 and BALB/c mice is shown. Results obtained for three individual mice per stimulus are each shown as a column in this heat map. Hierarchical clustering of transcripts in BM-DC from C57BL/6 mice revealed genes that are co-regulated. In BM-DC from both mouse strains, LPS and VSL#3 down-regulated a cluster of genes encoding TLR, e.g. Tlr5, Tlr4 and Tlr8. Moreover, both VSL#3 and LPS induced a set of pro-inflammatory gene transcripts, such as Tnf, Il1a and Il6, regardless of the mouse strain.

Fig. 3 Differences in gene expression between bone marrow-derived dendritic cells (BM-DC) from C57BL/6 mice and those from BALB/c mice. The supervised hierarchical clustering of genes related to Toll-like receptor signalling after stimulation of BM-DC with medium, lipopolysaccharide (LPS; 1 μg/ml), VSL#3 (107 colony-forming units/ml), or a combination of both is shown by the clusterogram. Relative expression levels for each individual gene are shown as minimum (green) and maximum (red). Columns represent the gene expression profiles of BM-DC from C57BL/6 and BALB/c mice after stimulation for 4 h (three individual mice per strain and experimental condition). For the description of gene symbols, see Table 1. -, Unstimulated cells.
Table 1 Gene description

In addition to the synergistic induction of IL-12 and IL-23 production (Fig. 2(d)–(g)), the TLR signalling array revealed that the combination of VSL#3 and LPS synergistically induced the expression of several other genes involved in innate immunity. In Fig. 4, the three representatives of this cluster–lymphotoxin A (Lta), Ifng and Ifnb – that were synergistically induced in BM-DC from C57BL/6 mice, but not in BM-DC from BALB/c mice, are shown.

Fig. 4 Synergistic induction of a cluster of genes in bone marrow-derived dendritic cells (BM-DC) from C57BL/6 mice, but not in those from BALB/c mice, by the probiotic mixture VSL#3 and lipopolysaccharide (LPS). The expression levels (i.e. C t values obtained from Fig. 3) of (a) lymphotoxin A (Lta), (b) interferon (Ifn)b and (c) Ifng in BM-DC from C57BL/6 or BALB/c mice were determined after stimulation with VSL#3 (107 colony-forming units/ml), LPS (1 μg/ml), or a combination of both. Bar graphs represent the fold induction compared with unstimulated BM-DC from C57BL/6 mice. Values are means, with standard deviations represented by vertical bars. For C57BL/6 mice, mean value was significantly different from that for LPS only: * P< 0·05; ** P< 0·01; *** P< 0·001 (Student's t test).
Importantly, a set of LPS-induced gene transcripts was suppressed by VSL#3 in BM-DC from both BALB/c and C57BL/6 mice. Myd88, Cxcl9 and Cxcl10 were the most important representatives of this cluster.
The suppressive effect of VSL#3 on LPS-induced Cxcl9 and Cxcl10 expression was verified by quantitative real-time PCR (Fig. 5(a) and (b)). Down-regulation was also confirmed from the protein levels of CXCL-10 (Fig. 5(c)): levels of CXCL-9 were below the detection limit.

Fig. 5 Suppression of lipopolysaccharide (LPS)-induced chemokine induction by the probiotic mixture VSL#3 and Lactobacillus plantarum. The mRNA expression levels of (a)Cxcl9 and (b) Cxcl10 in bone marrow-derived dendritic cells (BM-DC) from C57BL/6 mice were determined after stimulation with VSL#3 in the absence or presence of LPS (1 μg/ml). Data are presented relative to unstimulated cells and are representative of three independent experiments. Values are means of three individual mice, with standard deviations represented by vertical bars. Mean value was significantly different from that for LPS alone: * P< 0·05; *** P< 0·001 (Mann–Whitney U test). (c) CXCL-10 production after 24 h of stimulation was determined by multiplex assay. Data are given as concentrations measured in culture supernatants of BM-DC from three individual mice. Values are means, with standard deviations represented by vertical bars. *** Mean value was significantly different from that for LPS alone (P< 0·001; Mann–Whitney U test). The effect of L. plantarum and Bifidobacterium lactis on LPS-induced expression of (d) Cxcl9 and (e) Cxcl10 in BM-DC from C57BL/6 mice was investigated. Data are given as fold change of three individual mice, compared with unstimulated cells. Values are means, with standard deviations represented by vertical bars. * Mean value was significantly different from that for LPS alone (P< 0·05; Mann–Whitney U test). (f) CXCL-10 production after 24 h of stimulation was determined by multiplex assay. Data are given as concentrations in culture supernatants corresponding to three individual mice. Values are means, with standard deviations represented by vertical bars. ** Mean value was significantly different from that for LPS alone (P< 0·01; Mann–Whitney U test).
VSL#3 is composed of different members of two lactic acid genera, i.e. lactobacilli and bifidobacteria. Therefore, we studied the effects of representative members, i.e. L. plantarum and Bifidobacterium lactis. Whereas L. plantarum suppressed LPS-induced CXCL-10 expression at both the mRNA and protein levels and Cxcl9 expression at the mRNA level, B. lactis failed in this respect (Fig. 5(d) and (e)).
Discussion
Probiotic bacteria and prebiotic supplements have successfully been used in the treatment of various inflammatory disorders, including allergies and intestinal disorders( Reference Ozdemir 17 , Reference Arslanoglu, Moro and Schmitt 18 ). However, results of clinical trials are not consistent and variation in efficacy most probably depends on numerous factors, including bacterial strain type (single strain v. a mixture of strains), dosing regimen, delivery method and host factors, such as genetics, age, diet and disease state( Reference Gourbeyre, Denery and Bodinier 19 ). Several trials have shown the beneficial effects of lactobacilli, bifidobacteria and E. coli Nissle 1917 on the induction of remission as well as the maintenance of remission in patients with ulcerative colitis( Reference Cary and Boullata 20 ). However, the efficacy in Crohn's disease appears less evident( Reference Jonkers, Penders and Masclee 21 ). In the present study, we employed a simplified model system to evaluate the effects of VSL#3 – a complex mixture of TLR ligands – on BM-DC and identified both pro- and anti-inflammatory effects. These apparently contradictory findings may be due to the simultaneous presence of eight probiotic strains. On the other hand, similar contradictory effects were observed with L. plantarum, i.e. one single probiotic strain. In fact, even individual TLR ligands may induce a quick and transient production of pro-inflammatory cytokines such as IL-12, followed by the production of anti-inflammatory cytokines such as IL-10, as part of a suppressive mechanism essential for limiting the extent of inflammation. The balance between (the levels) of these cytokines may differ between individuals and represents a major determinant for the severity of the subsequent inflammation process. Besides, probiotic bacteria may mediate additional mechanisms in vivo that control the severity of inflammation, e.g. enhancement of intestinal barrier function, alteration of epithelial signalling, competition with pathogens, and secretion of SCFA such as acetate, propionate and butyrate, and also exert effects on immune cells distinct from DC( Reference Chang, Hao and Offermanns 22 – Reference Brestoff and Artis 26 ).
Although we allowed bone marrow progenitors from BALB/c v. C57BL/6 mice to differentiate in vitro into immature DC under controlled and identical conditions, we observed that VSL#3 and several TLR ligands induced a more pronounced pro-inflammatory response in cells from C57BL/6 mice, whereas those from BALB/c mice exhibited more IL-10 production. These observations are consistent with differences between the two mouse strains in terms of cellular and humoral immune responses to different pathogens( Reference Scott, Natovitz and Coffman 27 – Reference Coffman 29 ). No explanations for these differences could be obtained from the levels of Tlr gene expression in the present study. Moreover, we observed that VSL#3 augmented the induction of IL-12p70 and IL-23 production by LPS in a synergistic fashion in BM-DC from C57BL/6 mice, but not in those from BALB/c mice. Furthermore, several other genes involved in cellular immunity were synergistically up-regulated after combined stimulation with VSL#3 and LPS. This indicates that this mixture of probiotic bacteria has the potential to amplify inflammatory responses dependent on the genetic background of the host.
In parallel studies employing human DC, we demonstrated that LPS-induced phosphorylation of transducers and activators of transcription 1 (STAT-1) was inhibited by VSL#3, whereas phosphorylation of NF-κB was not affected (R Mariman and L Nagelkerken, unpublished results). A similar mechanism could explain the inhibition of secretion of chemokines, such as CXCL-9 and CXCL-10, by VSL#3, along with a robust induction of IL-12 and IL-23 production.
It should be noted that the effects of VSL#3 are not representative for individual probiotic strains, as has been shown by comparing L. plantarum and B. lactis in our in vitro studies.
Differential immune-modulating capacities of specific probiotic bacterial species in vivo are likely to be caused by the distinct repertoire of microbe-associated molecular patterns or effector molecules, which are expressed in a strain-specific manner( Reference van Baarlen, Troost and van der Meer 30 ).
Our findings also indicate that certain probiotic strains may be ineffective in vivo or may even display adverse effects during ongoing inflammation( Reference Besselink, van Santvoort and Buskens 31 , Reference Sanders, Akkermans and Haller 32 ). On the other hand, our extensive characterisation of gene expression during the induction of trinitrobenzene sulphonic acid colitis in BALB/c mice did not reveal that VSL#3 increased Il12p40 or Il23p19 mRNA levels in colonic tissue (not even in the acute phases of the model)( Reference Kremer, Mariman and van Erk 33 ). As has been mentioned above, this might be due to simultaneous induction of anti-inflammatory cytokine production and dampening of the local inflammatory response through mechanisms distinct from TLR signalling.
Importantly, in this in vitro study, we confirmed that probiotic bacteria may down-regulate the production of chemokines in agreement with our in vivo findings in the recurrent trinitrobenzene sulphonic acid colitis model, where we also showed less recruitment of inflammatory cells and a lower disease severity( Reference Kremer, Mariman and van Erk 33 ).
Altogether, our data indicate that the net effect of probiotic bacteria in vivo depends on a complex of factors that vary between individuals and may depend on their genetic background and health status.
Supplementary material
To view supplementary material for this article, please visit http://dx.doi.org/10.1017/S000711451400169X
Acknowledgements
The authors thank Frans Tielen and Arianne Plomp for their excellent technical assistance.
The present study was carried out in the framework of and with financial support from the Dutch Top Institute Pharma, project D1-101. The experiments were conceived and designed by R. M., B. K. and L. N. The experiments were performed by R. M. Data were analysed by R. M., B. K. and L. N. The paper was written by R. M., B. K., F. K. and L. N. The authors declare that the study was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.









