


Chicken coccidiosis is one of the most widely reported and economically important parasitic diseases in the poultry industry(
Reference Swaggerty, Genovese and He
1
). It is an intestinal disease that results in clinical and subclinical syndromes such as intestinal mucosal damage, reduced feed efficiency and decreased average daily gain(
Reference Matthews and Southern
2
,
Reference Witlock and Ruff
3
). Coccidiosis has been successfully controlled for decades using various anticoccidial drugs, while large-scale and long-term use of anticoccidial drugs has led to the development of resistance against anticoccidial agents(
Reference Peek and Landman
4
). Owing to the escalating consumer concerns about the use of chemotherapeutic agents as feed additives(
Reference Cox and Dalloul
5
), nutritional optimisation of immune function could be considered as a potential strategy to reduce the impact of coccidial infection. Following coccidial infection, increased mRNA expression level of inducible NO synthase (iNOS) has been reported(
Reference Laurent, Mancassola and Lacroix
6
). It has been shown that plasma levels of the ions
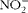 $$NO _{2}^{ - } $$
and
$$NO _{2}^{ - } $$
and
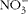 $$NO _{3}^{ - } $$
, stable metabolites of NO, are up-regulated during coccidial infection(
Reference Allen
7
), while plasma concentration of arginine (Arg) is reduced by coccidial infection(
Reference Allen and Fetterer
8
). Therefore, we presumed that due to a high level of iNOS expression, coccidial-infected chickens deplete more Arg than uninfected chickens to limit the production of NO that plays a critical role in parasite killing by direct(
Reference Vespa, Cunha and Silva
9
) or indirect (peroxynitrite, ONOO−)(10) actions.
$$NO _{3}^{ - } $$
, stable metabolites of NO, are up-regulated during coccidial infection(
Reference Allen
7
), while plasma concentration of arginine (Arg) is reduced by coccidial infection(
Reference Allen and Fetterer
8
). Therefore, we presumed that due to a high level of iNOS expression, coccidial-infected chickens deplete more Arg than uninfected chickens to limit the production of NO that plays a critical role in parasite killing by direct(
Reference Vespa, Cunha and Silva
9
) or indirect (peroxynitrite, ONOO−)(10) actions.
Coccidiosis has been developed as an experimental intestinal infection model( Reference Adams, Vahl and Veldman 11 ). Following coccidial infection, a remarkable up-regulation of pro-inflammatory cytokine expression, such as IL-1β and interferon-γ, has been observed( Reference Hong, Lillehoj and Lillehoj 12 ), which mediate the initiation of inflammation( Reference Dinarello 13 ). Inflammation is a defence response against infection, designed to clear foreign pathogens and to inhibit their detrimental effects( Reference Wyss-Coray and Mucke 14 ). Local inflammation recruits phagocytic and non-phagocytic lymphoid cells( Reference Khatri, Palmquist and Cha 15 ) to destroy pathogens, which is important to bird survival. Although the destruction of pathogens occurs intracellularly, release of cytotoxic molecules into the extracellular environment can cause tissue damage( Reference Smith 16 ). In the coccidial infection model, tissue damage is evidenced by intestinal epithelial sloughing and villus tip damage( Reference Witlock and Ruff 3 ). Intestinal mucosal barrier function is regulated by tight junction and adherens junction( Reference Bruewer, Luegering and Kucharzik 17 ). IL-1β has been reported to increase intestinal tight-junction permeability by the NF-κB-dependent pathway( Reference Al-Sadi, Ye and Dokladny 18 ). Increased mucosal barrier permeability exposes underlying immune cells to pathogens, further compromising barrier function( Reference Bruewer, Luegering and Kucharzik 17 ). Coccidial infection has been reported to increase gut permeability in chickens( Reference Rose, Long and Bradley 19 ). Therefore, we presumed that coccidial infection-induced IL-1β production may cause intestinal barrier dysfunction by increasing intestinal barrier permeability, and further induce intestinal mucosal disruption. Due to coccidial infection-induced intestinal mucosal disruption, infected chickens have lower apparent metabolisable energy, amino acid digestibility and N retention, leading to decreased growth rates( Reference Adams, Vahl and Veldman 11 , Reference Persia, Young and Utterback 20 ).
Chicken Toll-like receptor 4 (TLR4) serves as a sentinel by recognising pathogen-associated molecular patterns and triggering inflammation via the myeloid differentiation primary response gene 88 (MyD88)-dependent pathway( Reference Temperley, Berlin and Paton 21 , Reference Keestra and van Putten 22 ). Chickens lack the MyD88-independent pathway due to the absence of Toll/IL-1R-domain-containing adapter-inducing IFN-β (TRIF)-related adaptor molecule( Reference Keestra and van Putten 22 ). A previous in vitro study has shown that TLR4 and MyD88 are involved in chicken Eimeria tenella recognition and subsequent signal transduction( Reference Zhou, Wang and Cao 23 ). Dietary Arg supplementation has been shown to reduce lipopolysaccharide-induced pro-inflammatory cytokine expression via the suppression of the TLR4 pathway in our previous study( Reference Tan, Liu and Guo 24 ), and reduced expression of pro-inflammatory cytokines has been associated with the alleviation of intestinal mucosal disruption in pigs( Reference Liu, Huang and Hou 25 ). Thus, a question arises whether dietary Arg supplementation has a potential effect in partially alleviating coccidial challenge-induced intestinal damage and barrier dysfunction by decreasing the expression levels of pro-inflammatory cytokines.
Mechanistic target of rapamycin (mTOR) complex 1 consists of mTOR, Raptor (regulatory-associated protein of mTOR complex 1) and mLST8 (mammalian lethal with Sec13 protein 8)( Reference Pearce, Huang and Boudeau 26 ). p70 S6 kinase (p70s6k) lies downstream of the mTOR complex 1 signalling pathway, and the activation of p70s6k could enhance ribosome biosynthesis( Reference Christie, Hajduch and Hundal 27 ). Arg administration in cell-culture medium can activate the mTOR signalling pathway( Reference Kong, Tan and Yin 28 ), which mediates the stimulation of cell proliferation( Reference Murakami, Ichisaka and Maeda 29 ) and migration( Reference Rhoads, Niu and Odle 30 ). Therefore, the question remains whether dietary Arg supplementation accelerates the intestinal mucosal renewal process by enhancing intestinal cell proliferation and migration, thereby potentially alleviating intestinal damage and reduction in the growth performance of chickens.
On the basis of the aforementioned question, the present study was conducted to test the hypothesis that additional dietary Arg supplementation higher than those recommended by the National Research Council( 31 ) would have a potential effect in regulating intestinal structure and functionality, and mediating intestinal inflammation in coccidiosis-challenged broiler chickens.
Materials and methods
Birds, feed and experimental design
All animal care and use procedures for the present experiment were approved by the Purdue University Animal Care and Use Committee. A total of 288 male Ross broilers (708; Aviagen, Inc.) aged 1 d old were used for the present study. All chickens were housed in electrically heated battery cages (model no. SB 4T; Alternative Design Manufacturing) in an environmentally controlled room. Battery cage temperature was maintained at 37 ± 1°C for the 1st week and gradually decreased to 27°C in the 3rd week. The lighting schedule was 22 h light–2 h dark throughout the experiment. Chickens were weighed and allocated into groups so that the initial weight of each group was similar. Chickens were provided ad libitum access to drinking water and feed.
The randomised complete block design consisted of a 3 × 2 factorial arrangement of treatments to evaluate dietary Arg concentration (11·1, 13·3 and 20·1 g/kg of Arg, equal to 88·8, 106·4 and 160·8 % of the National Research Council recommendations; analysed concentrations) combined with or without a coccidiosis vaccine challenge, yielding a total of six treatment groups. Each treatment consisted of eight replicate cages, with each replicate containing six birds. The experimental design has been described in our previous study( Reference Tan, Liu and Guo 24 ), in which supplemental dietary Arg (14·2 and 19·0 g/kg; analysed concentrations) attenuated lipopolysaccharide-induced inflammation in broiler chickens. The lowest concentration value of Arg used in the supplementation was 11·1 g/kg. Ingredient formulation, nutrient and analysed dietary Arg concentrations are presented in Table 1. A basal diet was initially mixed, and all nutrients (except for Arg) were formulated to meet or exceed the National Research Council( 31 ) requirements. Final diets were obtained by mixing 98·4 % of the basal ration with a premix (1·6 %) containing different concentrations of Arg and Solka-Floc® (purified cellulose). Diet analyses for amino acid concentration were conducted using HPLC at the University of Missouri Agricultural Experiment Station Chemical Laboratory (Columbia, MO, USA; AOAC International, 2000; method 982.30 E (a, b, c))( 32 ).
Table 1 Composition of the experimental diets (g/kg)

CP, crude protein; ME, metabolisable energy.
* Supplied the following per kg complete diet: Cu, 8 mg; Zn, 75 mg; Fe, 80 mg; Mn, 100 mg; Se, 0·15 mg; I, 0·35 mg; trans-retinyl acetate, 24 mg; cholecalciferol, 6 mg; dl-α-tocopheryl acetate, 7·2 mg; menadione, 1·3 mg; thiamin, 2 mg; riboflavin, 6 mg; cyanocobalamin, 0·025 mg; biotin, 0·0325 mg; folic acid, 1·25 mg; pantothenic acid, 12 mg; niacin, 50 mg.
† Calculated value.
‡ Analysed concentrations.
Coccidial infection and sampling
At 14 d of age, chickens in the coccidiosis-challenged groups were challenged with twenty times the label-recommended individual commercial dose (1000 doses per bottle of freeze-dried vaccine) of coccidial vaccine (Coccivac-B; Schering-Plough Animal Health Corporation) by oral administration, and the unchallenged groups received the diluent. Coccivac-B contains live, attenuated oocysts of E. tenella, E. acervulina, E. maxima and E. mivati. In one previous study( Reference Smyth and Martin 33 ), ten times the label-recommended individual commercial dose of Coccivac-B has been used to induce necrotic enteritis. In our previous studies, twenty-five doses of Coccivac B induced moderate intestinal damage and inflammatory cytokine response in the jejunum (compared with the duodenum and ileum, the jejunum had greater inflammatory response; SA Adedokun, unpublished results). Therefore, twenty doses of Coccivac B were used as the challenge level in the present study. Chickens were weighed by pen at 1, 14 and 21 d of age; meanwhile, feed intake (FI) per pen was recorded to calculate the feed conversion ratio during the experiment. On day 21, one bird was randomly selected from each replicate and euthanised by an overdose of CO2 for tissue sampling.
Intestinal morphological analyses
Jejunal segments were collected at the midpoint between the bile duct entry and Meckel's diverticulum. Tissue was fixed in 10 % buffered formalin and embedded in Tissue Path (Fisher Scientific). Tissue sections (4 μm) were prepared and stained with Alcian Blue and periodic acid–Schiff's reagent as described previously( Reference Horn, Donkin and Applegate 34 ), with some modifications. Briefly, tissue sections were hydrated and stained with Alcian Blue solution (1 g Alcian Blue and 100 ml of 3 % acetic acid, pH 2·5) for 30 min, and rinsed in tap water for 5 min. The sections were then oxidised in 0·5 % periodic acid for 10 min, and rinsed in tap water for 5 min. The tissue sections were placed in Coleman's Schiff reagent solution (Sigma Chemical Company) for 10 min, and washed in lukewarm tap water for 5 min.
The sections were examined using light microscopy. Villus height, villus width and crypt depth were measured from nine villi per bird. Villus height was measured as the distance from the tip of the villus to the crypt mouth. Crypt depth was measured from the base of the villi to the submucosa. Villus width was measured as the average of width at one-third and two-thirds of the villus. Goblet cell counts were taken from the same nine villi per section and averaged, and goblet cell density was calculated as the average of goblet cell counts per μm of villus length. Thickness of the tunica muscularis was measured at nine different locations from the submucosa to the external layer of the intestine.
Mucosal sample analyses
Mucosa was scraped from 10 cm of the jejunum (5 cm proximal to the Meckel's diverticulum) and weighed. Mucosal density was calculated as mucosal weight per cm of jejunum. Mucosal disaccharidase (maltase and sucrase) activities were determined as described previously( Reference Dahlqvist 35 ), with some modifications. Mucosal secretory IgA (sIgA) and secretory IgG (sIgG) concentrations were determined by ELISA (Bethyl Laboratories, Inc.) following the manufacturer's protocol. Protein content of mucosal homogenates was measured colorimetrically by a commercially available kit (Bio-Rad) using bovine serum albumin as the standard. Specific disaccharidase activity and sIgA/sIgG concentrations were expressed as units per g protein.
Total RNA extraction and reverse transcription
Total RNA was extracted from the jejunum using TRIzol reagent (Invitrogen) following the manufacturer's protocol. The concentration of the extracted RNA was determined using a NanoDrop spectrophotometer (ND-1000; NanoDrop Products) at an optical density of 260 nm, and RNA purity was verified by measuring absorbance at an optical density of 260/280. Then, 2 μg of RNA were used for reverse transcription using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems) following the manufacturer's protocol. Synthesised complementary DNA was diluted 1:20 and stored at − 20°C until use.
Real-time quantitative PCR
Expression levels of the following genes were analysed in the jejunum by real-time quantitative PCR: jejunal mucosal immunity-related genes – Mucin-2, β-Defensin-8, IgA and polymeric Ig receptor (pIgR); inflammation-related genes – iNOS, IL-1β, IL-8, TLR4, MyD88, IL-1 receptor type I (IL-1RI) and NF-κB; apoptosis-regulatory genes – B-cell lymphoma 2 (Bcl-2) and Bcl-2-associated X gene (Bax); mTOR complex 1 pathway genes – mTOR, Raptor and ribosomal protein S6 kinase polypeptide 1 (RPS6KB1, encode p70s6k). Primers were designed based on sequences available from public databases (Table 2). Amplification and detection were performed using equivalent amounts of total RNA from jejunum on the Bio-Rad iQ5 detection system (Bio-Rad Laboratories) and the iQ SYBR Green Supermix (Bio-Rad Laboratories). The abundance of the housekeeping genes GAPDH (glyceraldehyde-3-phosphate dehydrogenase), β-actin, RPL-4 (ribosomal protein L4) and HPRT (hypoxanthine phosphoribosyltransferase) in the jejunum was not influenced by the treatments, whose relative expression stability was determined using geNorm and the ΔC
t approach(
Reference Silver, Best and Jiang
36
). GADPH was found to be the most suitable housekeeping gene for the present study, as also used in previous chicken coccidial challenge studies(
Reference Hong, Lillehoj and Lillehoj
12
,
Reference Adedokun, Ajuwon and Romero
37
). Standard curves were generated using the log10 diluted complementary DNA from pooled samples, and revealed that the amplification efficiency values were consistent between the target genes and the housekeeping genes. Real-time quantitative PCR were carried out in ninety-six-well plates at a final volume of 20 μl of the reaction mixture containing 10 μl iQ SYBR Green Supermix, 0·3 μm of each forward and reverse primer and 6 μl complementary DNA. Samples were subjected to the following protocol for all genes: 95°C for 5 min, forty cycles at 95°C for 10 s and 60°C for 20 s, and 72°C for 20 s. The samples were then analysed in triplicate. Relative gene expression data were analysed using the
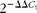 $$2^{ - \Delta \Delta C _{t}} $$
method(
Reference Livak and Schmittgen
38
), and the average ΔC
t value of the pooled sample served as the calibrator for each sample.
$$2^{ - \Delta \Delta C _{t}} $$
method(
Reference Livak and Schmittgen
38
), and the average ΔC
t value of the pooled sample served as the calibrator for each sample.
Table 2 Primers used in real-time quantitative PCR

GAPDH, glyceraldehyde-3-phosphate dehydrogenase; F, forward; R, reverse; iNOS, inducible NO synthase; TLR4, Toll-like receptor 4; MyD88, myeloid differentiation primary response gene 88; IL-1RI, IL-1 receptor type I; Bcl-2, B-cell lymphoma 2 gene; Bax, Bcl-2-associated X gene; mTOR complex 1, mechanistic target of rapamycin complex 1; Raptor, regulatory associated protein of mTOR complex 1; RPS6KB1, ribosomal protein S6 kinase polypeptide 1 (70 kDa); pIgR, polymeric Ig receptor.
* Sequence obtained from Ensembl chicken genome data resources.
Statistical analysis
Data were analysed using the general linear model procedure of SAS 9.3 (SAS Institute, Inc.). Data were subjected to two-way ANOVA in a 2 × 3 factorial arrangement with coccidial challenge and dietary Arg concentration as the main effects and their interactions. The initial body weight (14 d) was significantly affected by dietary Arg concentration; thus, 14 d body weight was used as a covariate in the model to analyse growth performance during coccidial challenge (14–21 d). Polynomial contrasts were used to test the linear and quadratic nature of the response to the analysed dietary Arg concentrations. The IML procedure was used to generate the appropriate contrast coefficients since the Arg levels were not equally spaced. When interactions were significant, polynomial contrasts were performed on the simple means to determine the linear and quadratic responses across the dietary Arg concentrations within the unchallenged and coccidiosis-challenged groups, respectively. When interactions were not significant, polynomial contrasts were performed on the main-effect means across the dietary Arg concentrations (averaged between the unchallenged and coccidial-challenged treatments). Differences with P <0·05 were considered significant, and those with P>0·05 < 0·10 were considered a trend.
Results
Growth performance
Data on growth performance are presented in Table 3. The interaction between coccidial challenge and dietary Arg concentration was found to be significant for body-weight gain (P< 0·05), but no significant interaction was observed for FI and feed conversion ratio (P>0·10). Increasing dietary Arg concentration linearly increased body-weight gain (P< 0·05) irrespective of the treatments. During the challenge period (14–21 d), coccidial challenge dramatically reduced FI by 17·83 % (P< 0·05), and significantly increased the feed conversion ratio (P< 0·001) in coccidiosis-challenged chickens. However, dietary Arg supplementation significantly increased FI (P< 0·001) and linearly decreased the feed conversion ratio (P< 0·001) in chickens challenged with coccidiosis.
Table 3 Growth performance of broilers fed diets containing arginine (Arg) concentrations at 11·1, 13·3 or 20·2 g/kg with or without coccidial vaccine challenge from 14 to 21 d of age (Mean values with their standard errors)
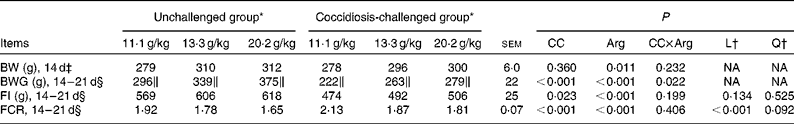
CC, coccidial challenge; L, linear; Q, quadratic; BW, body weight; NA, not assessed; BWG, body-weight gain; FI, feed intake; FCR, feed conversion ratio.
* Unchallenged group: chickens were orally supplemented with sterile saline (0·9 %) at 14 d of age; coccidiosis-challenged group: chickens received the same amount of coccidial vaccine (20 × ; Coccivac-B); means represent eight replicate cages per treatment with six birds per cage.
† When the interaction was not significant, linear and quadratic polynomial contrasts were performed on the main-effect means across the dietary Arg concentrations (averaged between the unchallenged and coccidial-challenged treatments); NA means that these contrasts were not assessed when the interaction was significant.
‡ Analysed by ANOVA.
§ Analysed by ANCOVA with 14 d BW as the covariate.
∥ When the interaction was significant, linear and quadratic polynomial contrasts were performed on the simple means across the dietary Arg concentrations within the unchallenged and coccidiosis-challenged groups, respectively; mean values within the unchallenged or coccidiosis-challenged groups with the symbol ∥ have a linear dose response to Arg concentration (P< 0·05); when there was no significant interaction, symbols are not presented.
Histological characteristics
Data on jejunal morphology and goblet cell counts at 7 d post-challenge are presented in Table 4. The interaction between coccidial challenge and dietary Arg concentration tended to be significant for the villus height:crypt depth ratio (P= 0·10), but no significant interaction was observed for any other histological characteristics (P>0·10). Histological assessment of jejunal samples from the coccidiosis-challenged groups revealed severe disruption of the normal architecture of the jejunum, showing inflammation in the jejunal mucosa. Compared with the unchallenged groups, jejunal samples from the coccidiosis-challenged groups were characterised by a lower villus height and villus height:crypt depth ratio (P< 0·05) and a higher villus width and muscularis thickness (P< 0·05). The inflammatory process was evidenced by crypt and tunica muscularis dilation, severe goblet cell depletion, and villus tip damage.
Table 4 Effect of graded supplementation of arginine (Arg) concentrations (11·1, 13·3 or 20·2 g/kg) on histological measurements from jejunal tissues in broiler chickens on day 7 after coccidial vaccine challenge (Mean values with their standard errors)

CC, coccidial challenge; L, linear; Q, quadratic.
* Unchallenged group: chickens were orally supplemented with sterile saline (0·9 %) at 14 d of age; coccidiosis-challenged group: chickens received the same amount of coccidial vaccine (20 × ; Coccivac-B); means represent eight replicate cages per treatment with one bird per cage.
† When the interaction was not significant, linear and quadratic polynomial contrasts were performed on the main-effect means across the dietary Arg concentrations (averaged between unchallenged and coccidiosis-challenged treatments).
Villus height, villus width and crypt depth
Coccidial challenge significantly decreased the jejunal villus height and villus height:crypt depth ratio (P< 0·05), while increasing dietary Arg concentration significantly increased villus height (P< 0·05). Following coccidial challenge, a significant increase was observed for crypt depth and villus width (P< 0·05). Dietary Arg supplementation linearly increased the crypt depth (P< 0·05).
Tunica muscularis thickness and goblet cell counts
Compared with the unchallenged groups, the tunica muscularis was found to be significantly thicker (P< 0·05) in the coccidiosis-challenged groups. A dramatic reduction was observed for goblet cell counts (per villus) and goblet cell density (per μm of villus height) (P< 0·001) in the coccidiosis-challenged groups compared with the unchallenged groups. Increasing dietary Arg concentration significantly increased goblet cell counts and goblet cell density (P< 0·05).
Mucosal density (per cm of jejunum) and disaccharidase (maltase and sucrase) activities
As shown in Table 5, the interaction between coccidial challenge and dietary Arg concentration was significant for mucosal maltase and sucrase activities (P< 0·05), but no significant interaction was observed for mucosal density (P>0·10). Dietary Arg supplementation linearly increased maltase activity in the unchallenged groups (P< 0·05), and quadratically increased maltase activity in the coccidiosis-challenged groups (P< 0·05), with the activity peaking at the intermediate dose of Arg supplementation (13·3 g/kg) and decreasing at the highest dose of Arg supplementation (20·2 g/kg). Within the coccidiosis-challenged groups, sucrase activity quadratically responded to increasing dietary Arg concentration (P< 0·05) that peaked at the intermediate dose of Arg supplementation (13·3 g/kg) and decreased at the highest dose of Arg supplementation (20·2 g/kg); however, within the unchallenged groups, no linear or quadratic trend was observed. Coccidial challenge significantly reduced mucosal density in the coccidiosis-challenged groups (P< 0·001), but increasing dietary Arg concentration linearly increased mucosal density (P< 0·05).
Table 5 Effects of graded supplementation of arginine (Arg) concentrations (11·1, 13·3 or 20·2 g/kg) on jejunal mucosal density, disaccharidase activity, secretory IgG (sIgG) and secretory IgA (sIgA) concentrations in broiler chickens on day 7 after coccidial vaccine challenge (Mean values with their standard errors)
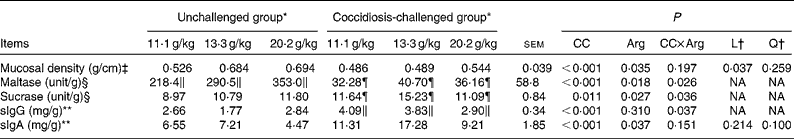
CC, coccidial challenge; L, linear; Q, quadratic; NA, not assessed.
* Unchallenged group: chickens were orally supplemented with sterile saline (0·9 %) at 14 d of age; coccidiosis-challenged group: chickens received the same amount of coccidial vaccine (20 × ; Coccivac-B); means represent eight replicate cages per treatment with one bird per cage.
† When the interaction was not significant, linear and quadratic polynomial contrasts were performed on the main-effect means across the dietary Arg concentrations (averaged between the unchallenged and coccidiosis-challenged treatments); NA means that these contrasts were not assessed when the interaction was significant.
‡ Based on per cm of jejunum.
§ Based on per g of mucosal protein; one unit of maltase or sucrase activity is equal to 1 μmol of glucose hydrolysed per min, respectively.
∥ When the interaction was significant, linear and quadratic polynomial contrasts were performed on the simple means across the dietary Arg concentrations within the unchallenged and coccidiosis-challenged groups, respectively; mean values within the unchallenged or coccidiosis-challenged groups with the symbol ∥ have a linear dose response to Arg concentration (P< 0·05); when there was no significant interaction, symbols are not presented.
¶ When the interaction was significant, linear and quadratic polynomial contrasts were performed on the simple means across the dietary Arg concentrations within the unchallenged and coccidiosis-challenged groups, respectively; mean values within the unchallenged or coccidiosis-challenged groups with the symbol ¶ have a quadratic dose response to Arg concentration (P< 0·05); when there was no significant interaction, symbols are not presented.
** Based on per g of mucosal protein.
Mucosal secretory IgG and secretory IgA concentrations
The data on the concentrations of jejunal mucosal sIgG and sIgA are presented in Table 5. There was a significant interaction between coccidial challenge and dietary Arg concentration for mucosal sIgG concentration (P< 0·05), but no significant interaction was observed for mucosal sIgA concentration. Chickens in the coccidiosis-challenged groups had linear reductions in sIgG concentration with increasing Arg concentration, but no linear or quadratic trend existed across the dietary Arg concentrations for chickens in the unchallenged groups. The coccidiosis-challenged groups showed higher mucosal sIgA concentration (P< 0·05) than the unchallenged groups. Dietary Arg supplementation significantly influenced mucosal sIgA concentration (P< 0·05), but no linear or quadratic trend was observed.
Gene expression data
Jejunal mucosal immunity-related gene expression
As shown in Table 6, no significant interactions between coccidial challenge and dietary Arg concentration were observed for any mucosal immunity-related gene expression (P>0·10). Coccidial challenge significantly decreased the mRNA expression levels of jejunal Mucin-2 and IgA (P< 0·05), and significantly increased the mRNA expression levels of jejunal β-Defensin-8 (P< 0·05). Dietary Arg supplementation significantly influenced the mRNA expression levels of pIgR (P< 0·05), but no linear or quadratic trend was observed.
Table 6 Effects of graded supplementation of arginine (Arg) concentrations (11·1, 13·3 or 20·2 g/kg) on the relative mRNA expression of jejunal mucosal immunity-related genes in broiler chickens on day 7 after coccidial vaccine challenge (Mean values with their standard errors)
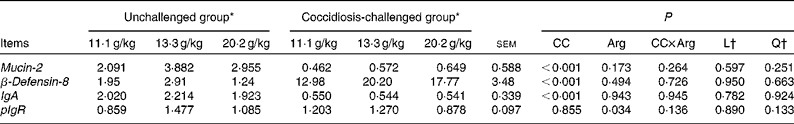
CC, coccidial challenge; L, linear; Q, quadratic.
* Unchallenged group: chickens were orally supplemented with sterile saline (0·9 %) at 14 d of age; coccidiosis-challenged group: chickens received the same amount of coccidial vaccine (20 × ; Coccivac-B); means represent eight replicate cages per treatment with one bird per cage.
† When the interaction was not significant, linear and quadratic polynomial contrasts were performed on the main-effect means across the dietary Arg concentrations (averaged between the unchallenged and coccidiosis-challenged treatments).
Inflammation-related gene expression
As shown in Table 7, there was a significant interaction between coccidial challenge and dietary Arg concentration for the mRNA expression of jejunal TLR4 (P< 0·05), and the interaction tended to be significant for the mRNA expression of jejunal MyD88 (P= 0·10), but no significant interaction was observed for the mRNA expression of jejunal NF-κB (P>0·10). Within the coccidiosis-challenged groups, increasing dietary Arg concentration linearly decreased the mRNA expression level of TLR4 (P< 0·05); however, within the unchallenged groups, no significant linear or quadratic trend was observed (P>0·10). Coccidial challenge significantly increased the mRNA expression levels of jejunal pro-inflammatory cytokine (IL-1β), chemokine (IL-8), iNOS and MyD88 in the coccidiosis-challenged groups (P< 0·05), and significantly decreased the mRNA expression level of IL-1RI (P< 0·05). Dietary Arg supplementation linearly increased the mRNA expression level of jejunal IL-1RI (P< 0·05), and tended to decrease the mRNA expression levels of jejunal IL-1β (P= 0·090) and MyD88 (P= 0·078). No differences were observed in the mRNA expression of NF-κB (P>0·01).
Table 7 Effect of graded supplementation of arginine (Arg) concentrations (11·1, 13·3 or 20·2 g/kg) on the relative mRNA expression of jejunal inflammation-related genes in broiler chickens on day 7 after coccidial vaccine challenge (Mean values with their standard errors)

CC, coccidial challenge; L, linear; Q, quadratic; iNOS, inducible NO synthase; IL-1RI, IL-1 receptor type I; TLR4, Toll-like receptor 4; NA, not assessed; MyD88, myeloid differentiation primary response gene 88.
* Unchallenged group: chickens were orally supplemented with sterile saline (0·9 %) at 14 d of age; coccidiosis-challenged group: chickens received the same amount of coccidial vaccine (20 × ; Coccivac-B); means represent eight replicate cages per treatment with one bird per cage.
† When the interaction was not significant, linear and quadratic polynomial contrasts were performed on the main-effect means across the dietary Arg concentrations (averaged between the unchallenged and coccidiosis-challenged treatments); NA means that these contrasts were not assessed when the interaction was significant.
‡ When the interaction was significant, linear and quadratic polynomial contrasts were performed on the simple means across the dietary Arg concentrations within the unchallenged and coccidiosis-challenged groups, respectively; mean values within the unchallenged or coccidiosis-challenged groups with the symbol ‡ have a linear dose response to Arg concentration (P< 0·05); when there was no significant interaction, symbols are not presented.
Apoptosis-regulatory gene expression
As shown in Table 8, no significant interactions between coccidial challenge and dietary Arg concentration were observed for apoptosis-regulatory gene expression (P>0·10). Coccidial challenge tended to down-regulate the mRNA expression of the anti-apoptosis gene Bcl-2 in the jejunum (P= 0·066). The mRNA expression of jejunal Bcl-2 quadratically responded (P< 0·05) to increasing Arg supplementation, which peaked at the intermediate dose of Arg supplementation (13·3 g/kg) and decreased at the highest level of Arg supplementation (20·2 g/kg). No significant difference was observed in the mRNA expression of Bax (P>0·10).
Table 8 Effect of graded supplementation of arginine (Arg) concentrations (11·1, 13·3 or 20·2 g/kg) on the relative mRNA expression of jejunal apoptosis-regulatory genes and mechanistic target of rapamycin (mTOR) complex 1 pathway genes in broiler chickens on day 7 after coccidial vaccine challenge (Mean values with their standard errors)
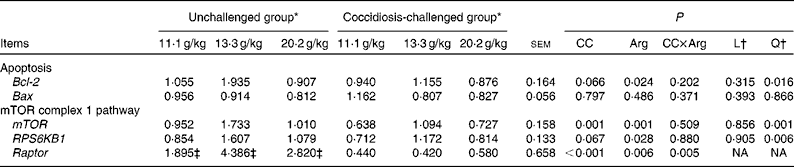
CC, coccidial challenge; L, linear; Q, quadratic; Bcl-2, B-cell lymphoma 2 gene; Bax, Bcl-2-associated X gene; RPS6KB1, ribosomal protein S6 kinase polypeptide 1 (70 kDa); Raptor, regulatory-associated protein of mTOR complex 1; NA, not assessed.
* Unchallenged group: chickens were orally supplemented with sterile saline (0·9 %) at 14 d of age; coccidiosis-challenged group: chickens received the same amount of coccidial vaccine (20 × ; Coccivac-B); means represent eight replicate cages per treatment with one bird per cage.
† When the interaction was not significant, linear and quadratic polynomial contrasts were performed on the main-effect means across the dietary Arg concentrations (averaged between the unchallenged and coccidiosis-challenged treatments); NA means that these contrasts were not assessed when the interaction was significant.
‡ When the interaction was significant, linear and quadratic polynomial contrasts were performed on the simple means across the dietary Arg concentrations within the unchallenged and coccidiosis-challenged groups, respectively; mean values within the unchallenged or coccidiosis-challenged groups with the symbol ‡ have a quadratic dose response to Arg concentration (P< 0·05); when there was no significant interaction, symbols are not presented.
Mechanistic target of rapamycin complex 1 pathway gene expression
As shown in Table 8, the interaction between coccidial challenge and dietary Arg concentration was significant for the mRNA expression of Raptor (P< 0·05), but no significant interactions were observed for the mRNA expression of mTOR and RPS6KB1 (P>0·10). The mRNA expression of Raptor responded quadratically within the unchallenged groups (P< 0·05), which peaked at the intermediate dose of Arg supplementation (13·3 g/kg) and decreased at the highest dose of Arg supplementation (20·2 g/kg); however, no significant linear or quadratic trend was observed within the coccidiosis-challenged groups (P>0·10). Coccidial challenge significantly decreased the mRNA expression level of jejunal mTOR (P= 0·001), and tended to decrease the mRNA expression level of RPS6KB1 (P= 0·067). The mRNA expression of jejunal mTOR and RPS6KB1 quadratically responded to increasing Arg concentration (P< 0·05), peaking at the intermediate dose of Arg supplementation (13·3 g/kg).
Discussion
Coccidiosis is an economically important disease in the poultry industry, and the economic loss caused by this disease is due to the reduction in growth performance and the cost involved in treatment and prevention( Reference Peek and Landman 4 ). In the present study, coccidial challenge remarkably increased the mRNA expression levels of IL-1β and IL-8 that were involved in jejunal inflammation. Inflammation is an essential part of the innate immune system, and involved in the recruitment of phagocytic cells to phagocytise and destroy infectious agents, clearing of cellular debris from the site of damage, and secretion of chemicals that attract other cell types to produce new tissue( Reference Khatri, Palmquist and Cha 15 , Reference Dee, Puleo and Bizios 39 ). During coccidial infection, survival followed by the growth performance of chickens becomes a priority. Therefore, jejunal inflammation plays an important role in the survival of chickens via processing and clearing of the pathogen with subsequent tissue repair. In the present study, coccidial challenge-induced inflammation caused severe mucosal disruption that was characterised by a decreased villus:crypt ratio, which in turn caused a reduction in growth performance.
Intestinal mucins are secreted by goblet cells and serve as a barrier to protect the epithelial cell layer from direct contact with bacteria( Reference Johansson, Phillipson and Petersson 40 ). In the present study, coccidial challenge caused severe depletion of goblet cells and reduction in the mRNA expression level of jejunal Mucin-2, with a concomitant 20 % reduction in mucosal density, indicating that coccidial challenge caused mucosal barrier dysfunction. The reduction in goblet cell counts and mRNA expression level of Mucin-2 probably degraded the protective mucin layer, thereby exposing jejunal epithelial cells to bacteria/coccidial oocysts that induced villus damage observed in the present study. Further inflammatory responses, explained in part by a higher expression level of IL-1β, have the potential to increase jejunal tight-junction permeability( Reference Al-Sadi, Ye and Dokladny 18 ).
The availability of Arg in the microenvironment (at the tumour site or sites of infection) has been reported to regulate the cell-cycle progression of T cells via the regulation of cyclin D3 and cyclin-dependent kinase 4( Reference Rodriguez, Quiceno and Ochoa 41 , Reference Zea, Rodriguez and Culotta 42 ). However, it is not yet clear whether Arg has a direct effect on goblet cells or enterocyte replication and/or a role in directing cell fate by a similar mechanism. Due partly to increased goblet cell counts, mucosal density linearly increased with increasing Arg concentration, indicating that dietary Arg supplementation may have partially protected the integrity of the intestinal mucosal barrier.
Chicken maltase is mainly produced in the upper half of the villus( Reference Uni, Platin and Sklan 43 ); thus, the reduction in mucosal maltase activity in the present study was due to coccidial challenge-induced damage to the upper villus. Various mechanisms have been related to the alterations in mucosal disaccharidase activities, including changes in gene expression within enterocytes and changes in the rates of maturation in enterocytes after infection( Reference Solaymani-Mohammadi and Singer 44 ). In the present study, dietary Arg supplementation linearly increased the crypt depth, plausibly increasing the replicating pool of enterocytes to replace the damaged enterocytes, but no trend was observed in jejunal villus height. Additionally, Arg supplementation concomitantly increased mucosal maltase activity. Therefore, the regulatory effect of dietary Arg supplementation on mucosal maltase activity was either via the direct regulation of maltase gene expression and/or through the regulation of villus growth. In the present study, sucrase activity responded quadratically with increasing dietary Arg concentration within the coccidiosis-challenged groups, but its cause remains unclear.
Previous studies showed that the activation of the TLR4 signalling pathway triggered IgG( Reference Yang, Murillo and Delannoy 45 ) and β-defensin( Reference Vora, Youdim and Thomas 46 ) responses. In the present study, the activation of TLR4 by coccidial challenge triggered an increase in mucosal sIgG concentration and jejunal β-Defensin-8 mRNA expression. Dietary Arg supplementation linearly reduced the mRNA expression level of TLR4 in the coccidiosis-challenged groups, which is consistent with our previous study using a lipopolysaccharide challenge model( Reference Tan, Liu and Guo 24 ). The suppression of the TLR4 signalling pathway decreased mucosal sIgG concentration; however, no difference was observed for β-Defensin-8, whose level was high in the jejunum of broiler chickens( Reference Hong, Song and Lee 47 ).
sIgA is generated by the cooperation of two types of cells: plasma cells secrete dimeric IgA and epithelial cells transport IgA into the lumen( Reference Johansen and Kaetzel 48 ). The transport of IgA is mediated by pIgR. iNOS plays a role in the regulation of IgA class-switch recombination, and adoptive transfer of iNOS+ dendritic cells restored IgA secretion in iNOS− / − mice( Reference Tezuka, Abe and Iwata 49 ). Thus, the increase in mucosal sIgA concentration by coccidial challenge in the present study was probably mediated by the activation of iNOS. Chicken NF-κB was involved in the activation of iNOS expression in macrophages( Reference Dil and Qureshi 50 ), but no difference was observed in the present study. Dietary Arg supplementation has been reported to increase mucosal sIgA concentration in rats( Reference Shang, Wang and Lai 51 ) and intestinal IgA-secreting cell numbers in piglets( Reference Zhu, Liu and Xie 52 ). In the present study, mucosal sIgA concentration and jejunal pIgR mRNA expression showed a similar changing trend with dietary Arg concentration. Thus, the regulatory effect of Arg supplementation on mucosal sIgA concentration may be mediated by the regulation of either IgA secretion and/or IgA transportation. Intriguingly, pre-treatment with citrulline also enhanced mucosal sIgA production in mice( Reference Batista, Nicoli and Santos Martins 53 ), suggesting that NO may participate in the regulation of sIgA secretion.
Cytokine and chemokine responses play a crucial role in immune defence against infection( Reference Withanage, Wigley and Kaiser 54 ). Following coccidial challenge, strong increases in pro-inflammatory cytokines, such as IL-1β, were observed in the present study. The effects of IL-1β are mediated by IL-1RI( Reference O'Neill and Dinarello 55 ). A previous study has shown that IL-1 synergises with IL-23 to act as an upstream regulator of the pro-inflammatory cytokine IL-17, whose production is significantly lower in IL-1RI− / − mice than in wild-type mice( Reference Sutton, Brereton and Keogh 56 ). Interestingly, coccidial challenge decreased the mRNA expression level of IL-1RI in the present study. Following E. tenella infection, chicken IL-17 mRNA expression has been shown to be down-regulated in the caecum( Reference Kim, Jeong and Park 57 ). Similar findings were observed in our unpublished results. Therefore, it is possible that the suppression of IL-1RI following coccidial challenge may have decreased the mRNA expression level of IL-17. IL-1RI is highly expressed in regulatory T cells( Reference Li, Kim and Boussiotis 58 ); thus, the decrease in the mRNA expression level of IL-1RI in the present study may be due to coccidial challenge-induced regulatory T-cell depletion. Meanwhile, increasing dietary Arg concentration linearly increased the mRNA expression level of IL-1RI, which may be associated with attenuated mucosal barrier dysfunction.
Coccivac-B contains the three most common pathogenic Eimeria species( Reference Sun, Pang and Jia 59 ): E. tenella; E. acervulina; E. maxima. Previous studies have reported that TLR4 expression is activated by E. tenella infection( Reference Zhou, Wang and Cao 23 , Reference Zhang, Liu and Ma 60 ) but reduced by E. acervulina infection( Reference Kim, Lillehoj and Min 62 ), and MyD88 expression is activated by E. maxima and E. tenella infection( Reference Hong, Lillehoj and Lillehoj 12 , Reference Zhou, Wang and Cao 23 , Reference Zhou, Hu and Wang 61 ). In the present study, mRNA expression levels of TLR4 and MyD88 were notably increased by coccidial challenge, indicating that the inflammatory response was probably triggered by the TLR4/MyD88 signalling pathway, which is consistent with previous study( Reference Zhou, Wang and Cao 23 ). Meanwhile, dietary Arg supplementation linearly reduced the mRNA expression level of TLR4 in the coccidiosis-challenged groups, and tended to reduce the mRNA expression levels of MyD88 and IL-1β, suggesting that Arg supplementation may have a potential inhibitory effect on coccidial challenge-induced inflammation via the suppression of the TLR4 signalling pathway. An attenuated inflammatory response in pigs has been reported to be associated with the mitigation of lipopolysaccharide-induced intestinal morphology impairment( Reference Liu, Huang and Hou 25 ). Therefore, the suppression of the TLR4 pathway in the present study may partially contribute to the mitigation of jejunal villus impairment. Chicken TLR4 is capable of recognising foreign pathogens and triggering the inflammatory cascade to protect against infection. TLR4-deficient mice have been shown to be defective in their ability to clear the virus( Reference Kurt-Jones, Popova and Kwinn 63 ). Thus, the suppression of the TLR4 pathway by dietary Arg supplementation may potentially compromise host defence against infection.
Apoptosis, the process of programmed cell death, is critical for development and protection against pathogens( Reference Adams and Cory 64 ). In E. acervulina-infected chickens, dynamic apoptosis has been observed in duodenal villi( Reference Major, Toth and Goldova 65 ). Increased production of NO after induction of iNOS has been reported to induce apoptosis in rat parenchymal cells( Reference Arstall, Sawyer and Fukazawa 66 ). In the present study, coccidial challenge increased the mRNA expression level of iNOS that may have altered jejunal apoptosis. Enhanced apoptosis has been reported to induce the exfoliation of epithelial cells in the chicken caecum( Reference Takeuchi, Kitagawa and Imagawa 67 ). Thus, jejunal mucosal disruption observed in the present study may partially be due to coccidial challenge-induced apoptosis. However, apoptosis is also essential for chickens to rid the body of infected cells, which is an auto-defensive response of chickens for survival. Bcl-2 has been shown to prevent apoptosis by blocking cytochrome c release( Reference Yang, Liu and Bhalla 68 ), while Bax has been proven to induce apoptosis( Reference Finucane, Bossy-Wetzel and Waterhouse 69 ). In the present study, compared with the lowest (11·1 g/kg) and the highest (20·2 g/kg) dose of Arg supplementation, supplemental Arg near the requirement level (13·3 g/kg) increased the mRNA expression level of jejunal Bcl-2, but no difference was observed for Bax mRNA expression. Arg as a source of NO has been reported to inhibit apoptosis in Nb2 lymphoma cells by a Bcl-2-mediated pathway( Reference Dodd, Limoges and Boudreau 70 ), and the blockade of the Arg/NO pathway increased apoptotic cell death in rats( Reference Yagnik, Takahashi and Tsoulfas 71 ). Thus, dietary Arg supplementation may have a potential inhibitory effect on coccidial-induced apoptosis via a NO-dependent, Bcl-2-mediated pathway.
NO, synthesised from Arg as well as citrulline, is endowed with the unique property of initiating and preventing apoptosis( Reference Brune, von Knethen and Sandau 72 ). Low-dose NO (0·1 mm-S-nitroso-N-acety1-N-dl-penicillamine, SNAP) administration as a pre-treatment can prevent mouse macrophage-like cells from apoptosis by the cooperation of heat shock proteins and DnaJ homologue, while high-dose NO administration (1·5 mm-SNAP) has been established to initiate apoptosis( Reference Gotoh, Terada and Mori 73 ). Immune-modulating nutrients can be helpful as well as harmful if used inappropriately( Reference Klasing 74 ). Dietary Arg supplementation has been reported to increase plasma NO levels in chickens( Reference Khajali, Tahmasebi and Hassanpour 75 ). In view of the promotional effect of high-dose NO administration on apoptosis, caution should be exercised when supplementing a high dose of Arg to prevent coccidial challenge-induced apoptosis.
Due to jejunal mucosal disruption, coccidiosis-challenged chickens showed lower mRNA expression level of mTOR. Amino acids have been shown to activate mTOR complex 1 via Ca2+/CaM signalling to hVps34 in HeLa cells grown in Dulbecco's modified Eagle's medium (Sigma)( Reference Gulati, Gaspers and Dann 76 ). In the present study, dietary Arg supplementation quadratically activated mTOR and RPS6KB1 mRNA expression, and quadratically increased Raptor mRNA expression in the unchallenged groups. These results indicated that the mTOR complex 1 pathway is sensitive to dietary Arg supplementation in chickens. A similar finding has been reported in cultured chicken hepatocytes, in which the mTOR/p70s6k pathway is activated by l-leucine administration( Reference Lee, Jo and Lee 77 ). mTOR complex 1 has been reported to regulate CD8+ T-cell differentiation( Reference Araki, Turner and Shaffer 78 ). Dietary Arg supplementation has been reported to increase the percentage of circulating CD8+ T cells( Reference Abdukalykova, Zhao and Ruiz-Feria 79 ). Thus, the up-regulation of the percentage of CD8+ T cells by dietary Arg supplementation was probably through the activation of the mTOR complex 1 pathway.
In chicken intestinal mucosa, a continuous renewal process occurs as enterocytes proliferate within the crypt and along the villus, and migrate along the crypt–villus axis( Reference Uni, Platin and Sklan 43 , Reference Uni, Geyra and Ben-Hur 80 ). Dietary Arg supplementation has been reported to increase jejunal villus height and crypt depth in pigs( Reference Wu, Ruan and Gao 81 ). Similar findings were observed in the present study. Arg administration activated the mTOR/p70s6k signalling pathway in rats, which is essential to intestinal cell migration and may play a role in intestinal repair( Reference Rhoads, Niu and Odle 30 ). In the present study, dietary Arg supplementation activated the mTOR complex 1 pathway, by which Arg accelerated enterocyte migration and the mucosal renewal process, and further enhanced jejunal mucosal recovery that was evidenced by increased villus height and crypt depth. The observed increase in villus height with Arg supplementation represents an augmentation in the surface area available for nutrient absorption, which enhances the feed efficiency of chickens with increasing Arg concentration. Thus, the partial mitigation of jejunal mucosal disruption observed in the present study may also be due to the enhancement of the jejunal mucosal renewal process by Arg supplementation via the activation of the mTOR complex 1 pathway.
In conclusion, the present study showed that the effects of dietary Arg supplementation on the partial alleviation of jejunal mucosal disruption were exhibited as a potential inhibitory effect on inflammation through the suppression of the TLR4 pathway, and an enhancement of the jejunal mucosal renewal process via the activation of the mTOR complex 1 pathway.
Acknowledgements
The authors thank Sunday A. Adedokun, Bertrand Grenier, Qian Zhang, Xi Chen and Kolapo Ajuwon for their assistance.
The present study was partially supported by China Scholarship Council. The funder had no role in the design and analysis of the study and in the writing of this article. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the US Department of Agriculture. US Department of Agriculture is an equal opportunity employer.
The authors’ contributions are as follows: J. T. conducted the animal trial, performed the sample analyses and wrote the manuscript; S. L. assisted with all data analyses; T. J. A. contributed to the study design, conduct of the study and preparation of the manuscript; Y. G. and S. D. E. contributed to the experimental design and preparation of the manuscript.
None of the authors has any conflicts of interest to declare.








