


INTRODUCTION
Several species of single-celled eukaryotes are important causes of death and disease in humans and domestic animals. These include such ancient scourges as the malaria parasites; trypanosomes, agents of sleeping sickness and Chagas disease; Leishmania parasites and numerous agents of primarily intestinal disease of humans and animals, including Cryptosporidium, Entamoeba and Giardia. Understanding the biology, and particularly the life cycles, of these parasites has been an important step in combating the diseases they cause. However, despite considerable research, many aspects of their biology remain mysterious.
A key part of the life cycle of an organism is reproduction. In protists, particularly parasitic protists, the sexuality or otherwise of numerous species has been hotly debated for many years (Tibayrenc et al. Reference Tibayrenc, Kjellberg and Ayala1990; Tibayrenc and Ayala, Reference Tibayrenc and Ayala2002). More recently, particularly with the advent of high throughput DNA sequencing technologies, evolutionary and population genomics have provided new evidence about reproduction in these organisms.
Here, we review some recent studies of sex and genetic exchange in parasitic protists and consider how the various reproductive modes of unicellular parasites may have adapted to differing parasitic lifestyles. Parasites considered in this review include important representatives of several major clades. Apicomplexans include malaria parasites of the genus Plasmodium as well as Babesia and Theileria and intestinal parasites of the genera Cryptosporidium, Toxoplasma and Eimeria. The Excavata include kinetoplastid parasites of the genera Trypanosoma and Leishmania as well as diplomonad parasites of the genus Giardia and parabasalid parasites of the genus Trichomonas. The Amoebozoa include enteric parasites of the genus Entamoeba.
SEX IN PARASITIC PROTISTS
True sex, consisting of cell fusion, nuclear fusion and meiosis is found only in eukaryotes. It is closely linked to the exchange and recombination of genetic material among individuals because it brings DNA molecules with different genealogies into contact, so that crossing over and exchange may occur. It has been argued that the evolution of sex is intimately associated with the origin of eukaryotes and arose much later than recombination, which occurs in all organisms (Cavalier-Smith, Reference Cavalier-Smith2002, Reference Cavalier-Smith2010).
Sex is widespread among eukaryotes and the reason for its maintenance is a central question of evolutionary biology. The benefits of sex, such as purging the genome of deleterious mutations and bringing together advantageous mutations, must be set against its fitness costs: mates have to be found, special cell types formed and diploid genomes maintained (Lehtonen et al. Reference Lehtonen, Jennions and Kokko2012). Indeed, the cost-benefit ratio may differ radically among species where, for instance, massive population sizes in micro-organisms (and the associated increase in the strength of selection) might obviate the need for sex to avoid Muller's ratchet, the irreversible accumulation of deleterious mutations. Some eukaryotes appear to have lost the ability to reproduce sexually altogether, such as some species of Daphnia and aphids (Innes and Hebert, Reference Innes and Hebert1988; Delmotte et al. Reference Delmotte, Leterme, Bonhomme, Rispe and Simon2001) but ancient asexuality appears to be rare (see Schurko and Logsdon, Reference Schurko and Logsdon2008).
The term ‘protists’, used to encompass numerous groups of species, can be misleading unless we remember just how vast the evolutionary distances between them can be (Baldauf, Reference Baldauf2003). The protists are not monophyletic and genetic distances between the different protist phyla are many orders of magnitude greater than that between fungi and mammals, for example. Of the parasitic protists considered here, the Apicomplexa include the genera Plasmodium, Babesia, Theileria, Toxoplasma, Eimeria, and Cryptosporidium; the Euglenozoa include Trypanosoma and Leishmania; the Fornicata include Giardia; the Parabasalia include Trichomonas; and the Amoebozoa include Entamoeba.
In discussing sex in the parasitic protists, two features should be considered: firstly, their status as protists; and secondly, their status as parasites.
The common feature of protists is their unicellularity, which makes them fundamentally different to multicellular organisms. In terms of their reproduction, both mitotic and meiotic cell divisions are reproduction, in that they both produce new individual cells. It is, therefore, entirely possible that reproduction is achieved only by mitosis. Another ‘unusual’ aspect of protists is that they do not conform to life-patterns familiar from multicellular organisms: diploid cells dividing by mitosis, interspersed with haploid gametes. Even obligate sexual protists, such as Plasmodium, spend the majority of their life cycle as haploid cells with only a brief spell of diploidy prior to meiosis. Therefore, meiosis may not be required for reproduction in some species that are able to complete their life cycles by clonal reproduction. In other species the haploid forms may be integral to the completion of the life cycle.
The second consideration is parasitism, a derived state that has arisen independently in many species. The evolutionary consequences of this are that many adaptations to the particular selective pressures of parasitism may have similar, independently derived results in diverse species. So distantly related parasites in similar niches may show similar adaptations while closely related lineages have evolved very different life cycles. This is shown in Fig. 1, which illustrates the life cycles of several parasites.
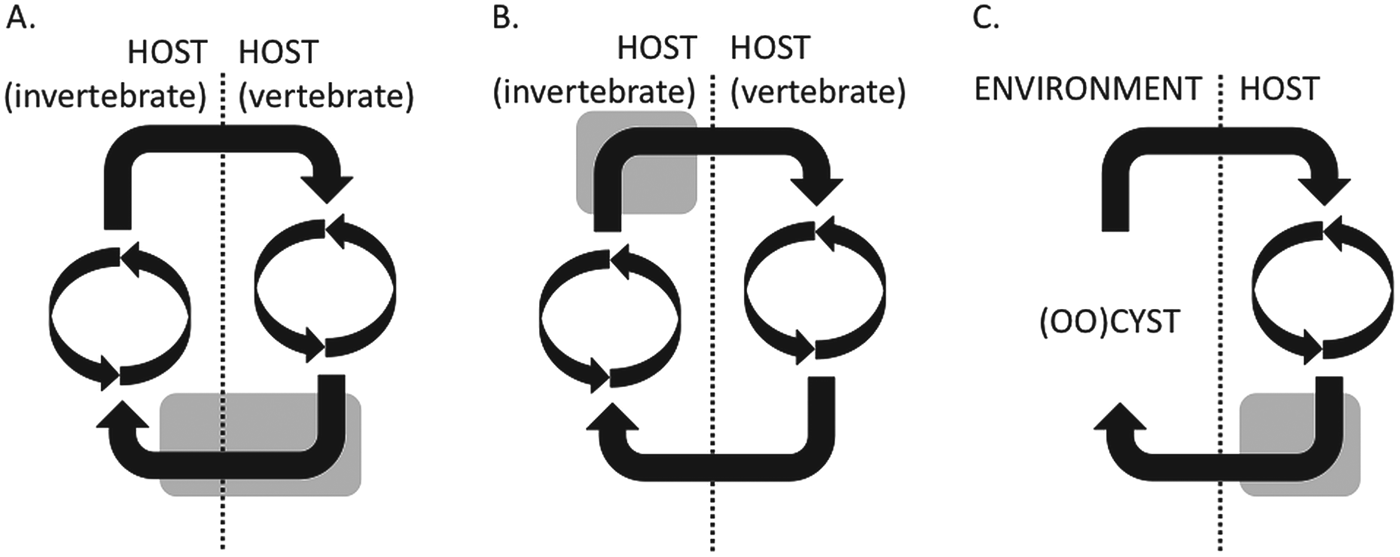
Fig. 1. Simplified schematic representations of the life cycles of vector- and fecal–oral-transmitted parasites. (A) The Plasmodium falciparum life cycle, which has two cycles of asexual growth, one in each host, and the sexual stages (grey shading) which span transmission from one host to another; (B) the T. brucei life cycle, in which the sexual stages appear to take place in the insect host, after a cycle of asexual cell divisions but before transmission to the mammalian host; (C) the Cryptosporidium and Eimeria life cycle, in which the sexual stages all take place within the single host and cysts are passed into the environment.
METHODS TO DETECT SEX AND GENETIC EXCHANGE IN PROTISTS
Given the appearance of many protists (single celled, no obvious sexual differentiation), it has historically been difficult to detect sexual reproduction. Moreover, since vegetative growth almost always can be seen to occur, it was often assumed to be the only form of reproduction employed by protists. This was also the case for many species of fungi (Dyer and O'Gorman, Reference Dyer and O'Gorman2011). However, these assumptions have, in many cases, proven to be wrong, or certainly to be not as simple as first thought.
How can sexual reproduction be inferred in a species where it is not readily apparent? There are, broadly, three ways to prove, or to infer, that a species is sexual. The clearest method, but often the most difficult, is to directly observe mating in vitro or in vivo. A second, indirect, method is to look for patterns of genetic variation in populations that indicate a sexual population structure, with out-crossing. A third indirect method is to identify genes in the genome that function in meiosis in other organisms.
Direct identification of sex in vivo and in vitro
The clearest way to demonstrate sexual reproduction is to carry out an experimental cross to show genetic exchange. However, this can often be difficult as many parasitic protists are not easy to maintain in vitro. It is possible to identify morphologically distinct sexual stages in some parasites, such as Plasmodium, though many species do not show obvious sexual stages. It is also possible to measure the amount of nuclear DNA, which can indicate changes in ploidy as a result of meiosis. However, gametes may be difficult to recognize due to unstable ploidy in many protists, with different chromosome ploidy being present within a single nucleus (Melville et al. Reference Melville, Gerrard and Blackwell1999; Willhoeft and Tannich, Reference Willhoeft and Tannich1999; Vargas et al. Reference Vargas, Pedroso and Zingales2004; Rogers et al. Reference Rogers, Hilley, Dickens, Wilkes, Bates, Depledge, Harris, Her, Herzyk, Imamura, Otto, Sanders, Seeger, Dujardin, Berriman, Smith, Hertz-Fowler and Mottram2011). If sex is an obligate stage of the parasite life cycle, it may be easier to detect than if it is facultative and occurs only under specific circumstances. Among the fungi, many readily cultured species were for years thought to be exclusively asexual until recently, when sex has been demonstrated under rarely occurring conditions (O'Gorman et al. Reference O'Gorman, Fuller and Dyer2009; Dyer and O'Gorman, Reference Dyer and O'Gorman2011). In some parasitic protists, it is possible that sex is facultative, which may help explain cases where the evidence is somewhat equivocal. If the species has mating incompatibility types, it may not be possible to induce mating if these are not known or included in the experiment.
Inferring sex by identification of meiosis genes
This approach has been developed for a set of meiosis genes shown to be present in all major eukaryotic lineages (Ramesh et al. Reference Ramesh, Malik and Logsdon2005; Malik et al. Reference Malik, Pightling, Stefaniak, Schurko and Logsdon2007; Schurko and Logsdon, Reference Schurko and Logsdon2008). These genes are shown in Table 1. The majority of meiosis genes have been detected in Giardia (Ramesh et al. Reference Ramesh, Malik and Logsdon2005), Trichomonas (Malik et al. Reference Malik, Pightling, Stefaniak, Schurko and Logsdon2007), Entamoeba (Loftus et al. Reference Loftus, Anderson, Davies, Alsmark, Samuelson, Amedeo, Roncaglia, Berriman, Hirt, Mann, Nozaki, Suh, Pop, Duchene, Ackers, Tannich, Leippe, Hofer, Bruchhaus, Willhoeft, Bhattacharya, Chillingworth, Churcher, Hance, Harris, Harris, Jagels, Moule, Mungall and Ormond2005; Ehrenkaufer et al. Reference Ehrenkaufer, Weedall, Williams, Lorenzi, Caler, Hall and Singh2013) and Trypanosoma, Plasmodium and Cryptosporidium (Malik et al. Reference Malik, Pightling, Stefaniak, Schurko and Logsdon2007).
Table 1. Genes of the ‘meiosis detection toolkit’ and evidence of their presence in parasitic protozoal lineages. (Adapted from Malik et al. Reference Malik, Pightling, Stefaniak, Schurko and Logsdon2007 and Schurko and Logsdon, Reference Schurko and Logsdon2008)

There are several caveats associated with using this approach to infer a species’ sexuality. Presence of the genes does not necessarily prove that a species is sexual; orthologues can have more than one function or can adapt to perform new functions in asexual species and so be maintained even in the absence of sex (reviewed in Schurko and Logsdon, Reference Schurko and Logsdon2008). The case for inferring sex from possession of meiosis genes is weakened considerably if we note that Drosophila melanogaster appears to lack hop1, hop2, mnd1, dmc1, msh4, msh5 and mer3 (see table in Malik et al. Reference Malik, Pightling, Stefaniak, Schurko and Logsdon2007), while species of uncertain or doubtful sexuality often possess a full or nearly full complement of meiosis genes. Conversely, it is not possible to confirm that a species is asexual based on failure to detect meiosis genes, as the orthologues could be present but highly divergent. However, while this is a problem when using PCR amplification to identify genes in the absence of a sequenced genome, an increasing number of completely sequenced genomes reduce this problem.
Additional support for the involvement of the genes in meiosis can be inferred from studying their expression pattern. In Entamoeba histolytica (Ehrenkaufer et al. Reference Ehrenkaufer, Weedall, Williams, Lorenzi, Caler, Hall and Singh2013) and Trypanosoma brucei (Peacock et al. Reference Peacock, Bailey, Carrington and Gibson2014) the expression of meiosis-specific genes coincides with developmental transitions and other evidence of meiotic events such as assortment in the case of T. brucei or production of tetranucleate cysts in the case of E. histolytica. However, in Giardia, up-regulation of meiosis genes is observed in the absence of meiosis (Carpenter et al. Reference Carpenter, Assaf, Gourguechon and Cande2012).
Inferring sex from population genetics
It is possible to observe the effects of genetic exchange among individuals by analysing the patterns of genetic variation within populations. True sex and mechanistically different parasexual processes of genetic exchange, as seen in bacteria, can have similar effects on patterns of population genetic variation. Clonality maintains linkage disequilibrium among polymorphic sites in the genome because there is no mechanism to break down physical linkage between alleles on the same chromosome or common descent of alleles on different chromosomes. Sex with outcrossing erodes linkage disequilibrium via recombination and re-assortment of chromosomes. However, as both asexual reproduction and sex without outcrossing will maintain linkage disequilibrium, clonal population structures do not rule out sex. The ‘clonal theory’ of parasitic protists stated that populations are primarily clonal, that is, sexual reproduction contributes very little to the population structures of these organisms (Tibayrenc et al. Reference Tibayrenc, Kjellberg and Ayala1990; Tibayrenc and Ayala, Reference Tibayrenc and Ayala2002). The theory is about the relative importance of outcrossing and inbreeding and is only concerned with population structures, not cell division mechanisms (Tibayrenc and Ayala, Reference Tibayrenc and Ayala2013). Therefore, the failure to identify recombination may not be taken as strong evidence for the complete absence of sex. Conversely, although identification of the products of recombination is consistent with sexual reproduction in a species, it is important to remember that, as in bacteria, recombination can occur without true sex, so genetic evidence for recombination is not proof positive of the occurrence of sex.
Sex in apicomplexan parasites
In apicomplexan parasites, sexual stages are evident and can be produced in in vitro culture. A sexual stage must occur to complete the life cycle. Laboratory strains of Plasmodium falciparum can be crossed in vivo, in the mosquito (Walliker et al. Reference Walliker, Quakyi, Wellems, McCutchan, Szarfman, London, Corcoran, Burkot and Carter1987). Cryptosporidium crosses have been described (Tanriverdi et al. Reference Tanriverdi, Blain, Deng, Ferdig and Widmer2007). A Toxoplasma gondii cross in a cat produced offspring of differing virulence (Herrmann et al. Reference Herrmann, Bärwald, Maksimov, Pantchev, Vrhovec, Conraths and Schares2012). In Babesia and Theileria species, direct fluorimetric measurement of DNA content of parasites through the life cycle showed changes in ploidy consistent with sex in the tick gut (Mackenstedt et al. Reference Mackenstedt, Gauer, Mehlhorn, Schein and Hauschild1990, Reference Mackenstedt, Gauer, Fuchs, Zapf, Schein and Mehlhorn1995; Gauer et al. Reference Gauer, Mackenstedt, Mehlhorn, Schein, Zapf, Njenga, Young and Morzaria1995). Genetic exchange has been demonstrated in Cryptosporidium parvum (Feng et al. Reference Feng, Rich, Tzipori and Widmer2002).
Plasmodium parasites have a dimorphic sexual stage that is closely linked to the transmission cycle (the sexual stage is the transmissible stage). Plasmodium parasites have complex life cycles, passing through two hosts, one of which is an insect vector. The process of gametocytogenesis, in which lineages of blood stage, asexually reproducing cells switch to form male (micro-) or female (macro-) gametocytes, is intimately involved in transmission between hosts. During gametocytogenesis, the parasite changes both morphologically and biochemically (Dixon, Reference Dixon, Thompson, Gardiner and Trenholme2008). Commitment to form gametocytes occurs one cycle before the manifestation of gametocytes (Dixon, Reference Dixon, Thompson, Gardiner and Trenholme2008). All merozoites from a single schizont are committed to sexual/asexual pathway. Also, all merozoites from a committed schizont form only male or only female gametocytes (Silvestrini, Reference Silvestrini, Alano and Williams2000; Smith, Reference Smith, Lourenço, Carter, Walliker and Ranford-Cartwright2000). Gametogenesis is the emergence of micro-/macro-gametocytes from infected red blood cells.
Plasmodium falciparum shows a range of population structures from ‘clonal’ to ‘sexual’, which broadly reflect the local transmission intensity. For instance, African populations with high transmission intensity have a greater chance of outcrossing and show very low levels of linkage disequilibrium (Conway et al. Reference Conway, Roper, Oduola, Arnot, Kremsner, Grobusch, Curtis and Greenwood1999; Anderson et al. Reference Anderson, Haubold, Williams, Estrada-Franco, Richardson, Mollinedo, Bockarie, Mokili, Mharakurwa, French, Whitworth, Velez, Brockman, Nosten, Ferreira and Day2000), while South East Asian populations with lower transmission intensity show less outcrossing and more ‘clonal’ population structures (Volkman et al. Reference Volkman, Sabeti, DeCaprio, Neafsey, Schaffner, Milner, Daily, Sarr, Ndiaye, Ndir, Mboup, Duraisingh, Lukens, Derr, Stange-Thomann, Waggoner, Onofrio, Ziaugra, Mauceli, Gnerre, Jaffe, Zainoun, Wiegand, Birren, Hartl, Galagan, Lander and Wirth2007). Population genetic analysis of the cattle parasite Babesia bovis from Zambia and Turkey indicate a sexual, outcrossing population structure (Simuunza et al. Reference Simuunza, Bilgic, Karagenc, Syakalima, Shiels, Tait and Weir2011). Babesia bovis undergoes sexual development and fertilization in the invertebrate host (Mackenstedt et al. Reference Mackenstedt, Gauer, Fuchs, Zapf, Schein and Mehlhorn1995; Gough et al. Reference Gough, Jorgensen and Kemp1998). Similar observations have been made in the related parasites Theileria parva (Gauer et al. Reference Gauer, Mackenstedt, Mehlhorn, Schein, Zapf, Njenga, Young and Morzaria1995). It is probably unsurprising in tick-borne diseases, where a single host is likely to be bitten by multiple infected ticks, that there is a lot of scope for mixed infections where recombination will occur. The broad host-range mammalian parasite T. gondii, by contrast, displays a highly clonal population structure. It is possible that this reflects the fact that individuals may rarely meet in the host, unlike the situation in Plasmodium or enteric parasites where transmission is high and many unrelated individuals may co-exist.
In T. gondii the sexual cycle only occurs in cats, the definitive host. In this case the parasite will differentiate intracellularly into male microgametes and female macrogametes, which undergo fertilization to produce a diploid zygote (Ferguson et al. Reference Ferguson, Hutchison, Dunachie and Siim1974, Reference Ferguson, Hutchison and Siim1975; Ferguson, Reference Ferguson2002). Unlike in Plasmodium, this sexual cycle has not resulted in a panmictic population structure. In fact, what is observed is three clonal lineages found in human populations while a great deal of genetic diversity can be observed in wild animals (Grigg and Sundar, Reference Grigg and Sundar2009). Boyle et al. (Reference Boyle, Rajasekar, Saeij, Ajioka, Berriman, Paulsen, Roos, Sibley, White and Boothroyd2006) have demonstrated that the pattern of polymorphism of Types I and III Toxoplasma are indicative of these being first and second generation offspring of a Type II strain cross with an ancestral line. This observation highlights that a single recombination event can have a major impact on the population structure of a pathogen.
It therefore appears that in Apicomplexa parasites, where haploid stages are often the infective forms of the parasite, sex is common and probably ubiquitous. Clonal population structures such as those seen in Toxoplasma are likely caused by the rapid expansion of epidemic strains that propagate in certain hosts, in this case human.
Sex in kinetoplastid parasites
In kinetoplastid parasites (Trypanosoma and Leishmania), sexual stages are not easily identifiable. However, genetic exchange has been identified from laboratory crosses of T. brucei in the tsetse fly host (MacLeod et al. Reference MacLeod, Tweedie, McLellan, Hope, Taylor, Cooper, Sweeney, Turner and Tait2005) and of Leishmania major in the sand fly host (Akopyants et al. Reference Akopyants, Kimblin, Secundino, Patrick, Peters, Lawyer, Dobson, Beverley and Sacks2009). In T. brucei, cell fusion has been demonstrated between cells containing red or green fluorescent proteins leading to yellow fluorescent hybrids in vivo, in the salivary glands of the tsetse fly host (Gibson et al. Reference Gibson, Peacock, Ferris, Williams and Bailey2008) and in vitro, between cells derived from salivary glands but not between cells derived from the midgut (Peacock et al. Reference Peacock, Bailey, Carrington and Gibson2014). Measurement of nuclear DNA content throughout the life cycle showed a haploid ‘promastigote-like’ cell in the salivary glands of the fly (Peacock et al. Reference Peacock, Bailey, Carrington and Gibson2014) and cells expressing meiosis genes (Mnd1, Dmc1 and Hop1, see below) prior to cell fusion (Peacock et al. Reference Peacock, Ferris, Sharma, Sunter, Bailey, Carrington and Gibson2011, Reference Peacock, Bailey, Carrington and Gibson2014). This, together with the evidence from genetic crosses, indicates that the life cycle of T. brucei does contain a sexual stage and meiosis.
Trypanosoma cruzi and Trypanosoma vivax appear to have all of the genes for meiosis (Malik et al. Reference Malik, Pightling, Stefaniak, Schurko and Logsdon2007), despite very little evidence from the population structure that meiosis is occurring (Westenberger et al. Reference Westenberger, Barnabé, Campbell and Sturm2005; Duffy et al. Reference Duffy, Morrison, Black, Pinchbeck, Christley, Schoenefeld, Tait, Turner and MacLeod2009). Genetic exchange was demonstrated in T. cruzi by observing that two strains carrying different drug-selectable markers produced strains carrying both markers in the mammalian host (Gaunt et al. Reference Gaunt, Yeo, Frame, Stothard, Carrasco, Taylor, Mena, Veazey, Miles, Acosta, de Arias and Miles2003). However, the proposed mechanism of this exchange was a fusion of diploid cells followed by chromosome loss, rather than meiosis; quite different to sex in T. brucei.
The population structure of T. brucei is characteristic of facultative recombination, with clonal population structures in foci of human disease and evidence of rare recombination in the wider population (Macleod et al. Reference MacLeod, Tait and Turner2001). By contrast, T. vivax displays a clonal population structure, consistent with its more limited life cycle in the tsetse fly host where it remains in the mouthparts (Duffy et al. Reference Duffy, Morrison, Black, Pinchbeck, Christley, Schoenefeld, Tait, Turner and MacLeod2009). Due to its more limited life cycle T. vivax is able to utilize various species of biting flies as vectors which is advantageous but may have evolved at the expense of its ability to undergo meiosis and sexual recombination. In T. cruzi, the population structure appears to be clonal, with six major lineages circulating in human population. Recombinant strains have been described (Lewis et al. Reference Lewis, Llewellyn, Yeo, Acosta, Gaunt and Miles2011) but whether or not these were produced by true sex is unknown. Westenberger et al. (Reference Westenberger, Barnabé, Campbell and Sturm2005) proposed a model whereby fusion and loss of heterozygosity between two ancestral strains could give rise to all of the major lineages observed today.
Sex in amoebozoan parasites
Entamoeba histolytica is a parasitic amoeba of humans. Its life cycle has two distinct stages, a motile trophozoite that lives in the colon and a cyst that passes out of the host, can survive in the environment and be transmitted to new hosts.
Entamoeba histolytica was traditionally thought to be asexual, with intestinal trophozoites dividing by mitosis and encystation and excystation being mitotic too. The cysts may be formed by incomplete mitosis, as, in E. histolytica, they contain four nuclei (other species contain different numbers). Indirect evidence indicates sex, including possession of meiosis genes (Loftus et al. Reference Loftus, Anderson, Davies, Alsmark, Samuelson, Amedeo, Roncaglia, Berriman, Hirt, Mann, Nozaki, Suh, Pop, Duchene, Ackers, Tannich, Leippe, Hofer, Bruchhaus, Willhoeft, Bhattacharya, Chillingworth, Churcher, Hance, Harris, Harris, Jagels, Moule, Mungall and Ormond2005) and population genetic evidence (Gilchrist et al. Reference Gilchrist, Ali, Kabir, Alam, Scherbakova, Ferlanti, Weedall, Hall, Haque, Petri and Caler2012; Weedall et al. Reference Weedall, Clark, Koldkjaer, Kay, Bruchhaus, Tannich, Paterson and Hall2012). A study of genomes derived from several geographical origins indicated decay in linkage with increased physical distance between variant sites (Weedall et al. Reference Weedall, Clark, Koldkjaer, Kay, Bruchhaus, Tannich, Paterson and Hall2012) and a set of single nucleotide polymorphism (SNP) markers in field isolates from Bangladesh showed no linkage disequilibrium at all between markers (Gilchrist et al. Reference Gilchrist, Ali, Kabir, Alam, Scherbakova, Ferlanti, Weedall, Hall, Haque, Petri and Caler2012), both indicative of recombination and reassortment.
If the species is sexual, when might meiosis occur? Either it must occur at some point in the transition between trophozoite or during a facultative sexual cycle under specific circumstances. During the transition from trophozoite to cyst, several meiosis genes are up-regulated (Ehrenkaufer et al. Reference Ehrenkaufer, Weedall, Williams, Lorenzi, Caler, Hall and Singh2013; Singh et al. Reference Singh, Bhattacharya and Bhattacharya2013) and homologous recombination occurs (Singh et al. Reference Singh, Bhattacharya and Bhattacharya2013). This may indicate meiosis, as appears to be the case in T. brucei (Peacock et al. Reference Peacock, Ferris, Sharma, Sunter, Bailey, Carrington and Gibson2011, Reference Peacock, Bailey, Carrington and Gibson2014), but could also indicate a non-meiotic process, as appears to be the case in Giardia lamblia (Carpenter et al. Reference Carpenter, Assaf, Gourguechon and Cande2012).
Another (albeit distantly related) amoebazoan, Dictyostelium discoideum, has a sexual cycle and appears to mate frequently in wild populations (Flowers et al. Reference Flowers, Li, Stathos, Saxer, Ostrowski, Queller, Strassmann and Purugganan2010). Sex appears to be facultative and to occur under stress conditions (Saga et al. Reference Saga, Okada and Yanagisawa1983). Dictyostelium discoideum has three sexes, determined by a mating type locus (Bloomfield et al. Reference Bloomfield, Skelton, Ivens, Tanaka and Kay2010). Haploid amoebae of different sexes fuse, attract surrounding cells and form a macrocyst in which meiosis and mitosis (presumably) occur to produce haploid cells (Saga et al. Reference Saga, Okada and Yanagisawa1983). However, D. discoideum does appear to lack five meiosis genes.
Sex in Giardia and Trichomonas parasites
Direct observation in vitro or in vivo can confirm sex, but not observing it does not provide definitive evidence that a species is asexual. For instance, in G. lamblia population genetic evidence has been used to infer sex (Cooper et al. Reference Cooper, Adam, Worobey and Sterling2007), and if sex occurs, it is thought to happen during encystation or excystation. Laboratory studies have demonstrated that nuclear division during encystation is mitotic, not meiotic (Carpenter et al. Reference Carpenter, Assaf, Gourguechon and Cande2012; Jirakova et al. Reference Jiráková, Kulda and Nohýnková2012). Using the distribution of different markers integrated into the genomes of two cell lines, Carpenter et al. (Reference Carpenter, Assaf, Gourguechon and Cande2012) were able to track the inheritance of nuclei during encystation and excystation. They found no evidence of cell fusion and also that pairs of parental nuclei were co-inherited (i.e. nuclear sorting did not occur). Several meiotic gene homologues are up-regulated during encystation, indicating that homologous recombination might occur during encystation/excystation (Carpenter et al. Reference Carpenter, Assaf, Gourguechon and Cande2012). The study found evidence for ‘diplomixis’, or chromosomal genetic exchange between nuclei, and the authors argued that this, along with homologous recombination, might be sufficient to maintain the low levels of allelic heterozygosity seen in sequenced genomes (Morrison et al. Reference Morrison, McArthur, Gillin, Aley, Adam, Olsen, Best, Cande, Chen, Cipriano, Davids, Dawson, Elmendorf, Hehl, Holder, Huse, Kim, Lasek-Nesselquist, Manning, Nigam, Nixon, Palm, Passamaneck, Prabhu, Reich, Reiner, Samuelson, Svard and Sogin2007; Jerlstrom-Hultqvist et al. Reference Jerlström-Hultqvist, Franzén, Ankarklev, Xu, Nohýnková, Andersson, Svärd and Andersson2010). However, this model of genetic exchange cannot explain population genetic evidence for recombination within and between assemblages of G. lamblia (Cooper et al. Reference Cooper, Adam, Worobey and Sterling2007; Lasek-Nesselquist et al. Reference Lasek-Nesselquist, Welch, Thompson, Steuart and Sogin2009), which would require cell fusion events and some form of meiosis to occur.
The discrepancy between the evidence from in vitro experiments and population genetic observations of G. lamblia (Cooper et al. Reference Cooper, Adam, Worobey and Sterling2007; Carpenter et al. Reference Carpenter, Assaf, Gourguechon and Cande2012) could be resolved if sex is facultative and occurs, even rarely, in natural populations. Also, if mating compatibility is determined by mating type loci, as in D. discoideum, the in vitro experiments may not show any mating if a single strain is used.
Trichomonas vaginalis is a parabasalid parasite of humans that is sexually transmitted. Two major lineages, both with a global distribution, can be defined by microsatellite patterns, yet within populations there is little genome wide linkage disequilibrium, suggesting that recombination is breaking linkage (Conrad et al. Reference Conrad, Gorman, Schillinger, Fiori, Arroyo, Malla, Dubey, Gonzalez, Blank, Secor and Carlton2012).
SEXUAL STAGES AND TRANSMISSION BETWEEN HOSTS
In many different species, meiosis and recombination are linked to dispersal. Either gametes or dormant zygotes (or seeds) are dispersed to find new environmental niches. Dispersal (transmission) is central to the parasitic lifestyle. Cysts, often the transmissible stages in parasitic species, may have occurred in early eukaryotes and may be linked to the evolution of sex (Cavalier-Smith, Reference Cavalier-Smith2002). However, cysts and cyst-like stages may be examples of convergent evolution, as the materials forming cyst walls differ among species and the encystation process may also be very different (Lauwaet et al. Reference Lauwaet, Davids, Reiner and Gillin2007; Ehrenkaufer et al. Reference Ehrenkaufer, Weedall, Williams, Lorenzi, Caler, Hall and Singh2013; Samuelson et al. Reference Samuelson, Bushkin, Chatterjee and Robbins2013). We may identify a link between sex and transmission in parasites.
To illustrate how the sexual stages of parasites can be linked to their life cycle and adapt to their differing lifestyles, consider two greatly different modes of transmission in parasites of the Apicomplexa: transmission via a vector species and direct transmission via the fecal–oral route. Some apicomplexans, including Plasmodium and Babesia species, are transmitted from a vertebrate host to an insect or tick host and to another vertebrate host. Others, including Cryptosporidium, Eimeria and Toxoplasma species, are transmitted among hosts via an environmental stage (the oocyst). The requirements of these modes of transmission are different and the sexual stages appear to be adapted accordingly. In the environmentally transmitted parasites (e.g. Eimeria), oocysts are tough structures allowing survival outside of a host. Gametocytogenesis and gametogenesis form a continuous process that takes place inside one infected host cell, and appears to be programmed to occur after approximately three asexual cycles. In contrast, vector borne parasites must negotiate a novel host. Plasmodium and Babesia parasites display a pause between gametocytogenesis and gametogenesis so that gametogenesis occurs after transmission, within the gut of the mosquito/tick host, rather than in the vertebrate host. Plasmodium parasites have a stage not seen in fecal–oral transmitted parasites: the ookinete is formed after gamete fusion, it crosses the insect gut wall before forming the oocyst. In contrast to the apparently timed triggering of sexual commitment in Eimeria, in Plasmodium, commitment to sexual development appears to be triggered by environment and to be ‘staggered’ to facilitate transmission. Producing transmissible stages over a longer time makes sense for vector borne parasites to increase the chances of encountering a vector. In fecal–oral transmitted parasites, transmissible stages will be passed into the environment in any case and controlling of the timing of their production is likely to be less important than their ability to survive once there.
Concluding remarks
Understanding the sexual biology of parasitic protists is not only of intrinsic scientific interest but may also be biomedically relevant, informing potential treatments to target sexual stages of the parasites. For instance, knowing that sex is necessary for apicomplexan parasites to complete their life cycles makes sexual stages potential targets for intervention. By contrast, facultative sexual stages may not be useful targets. Biological knowledge of the sexual stages, in particular the comparative biology of sex, may allow for the identification of conserved target processes, molecules or structures (e.g. proteins mediating cell binding or flagella in microgametes (Wallach et al. Reference Wallach, Smith, Petracca, Miller, Eckert and Braun1995, Reference Wallach, Ashash, Michael and Smith2008). Therefore, improving our understanding of sex in parasitic protists is an important goal for future parasitological study.
ACKNOWLEDGEMENTS
We acknowledge two anonymous reviewers for their careful reading of the manuscript and constructive comments.
FINANCIAL SUPPORT
Both authors have been supported by the Medical Research Council and The Wellcome Trust.



 Open access
Open access