


Developmental plasticity allows the generation of a number of phenotypes from a single genotype (Gluckman & Hanson, Reference Gluckman and Hanson2004). Epidemiological (Godfrey & Barker, Reference Gluckman and Hanson2001) and experimental (Bertram & Hanson, Reference Bertram and Hanson2002) studies show that aspects of the prenatal environment such as maternal nutrition and stress levels provide cues that modify the phenotype of the offspring without overt reductions in fetal growth. Such nutritional cues operate within the normal range for the human population and contribute to the early-life origins of risk of chronic diseases such as the metabolic syndrome (Godfrey & Barker, 2001).
There is evidence in humans and in experimental models for non-genomic transmission between generations of induced phenotypic traits associated with an impaired capacity to maintain energy balance. The mortality from diabetes was increased in men if the paternal grandfather had been exposed to abundant nutrition during puberty (Pembrey et al. Reference Pembrey, Bygren, Kaati, Edvinsson, Northstone, Sjostrom and Golding2006). The daughters of women exposed to nutrient restriction and environmental stress during pregnancy as a result of the Dutch Hunger Winter showed a decreased birth weight and an increased risk of insulin resistance, and their daughters also were born with a lower birth weight (Stein & Lumey, Reference Stein and Lumey2000; Painter et al. Reference Painter, Roseboom and Bleker2005). In rats, feeding a protein-restricted diet (PRD) to the F0 generation during pregnancy resulted in elevated blood pressure and endothelial dysfunction (Torrens et al. Reference Torrens, Brawley, Dance, Itoh, Poston and Hanson2002) and insulin resistance (Martin et al. Reference Martin, Johnston, Han and Benyshek2000; Zambrano et al. Reference Zambrano, Martinez-Samayoa, Bautista, Deas, Guillen, Rodriguez-Gonzalez, Guzman, Larrea and Nathanielsz2005) in the F1 and F2 generations, despite normal nutrition during pregnancy in the F1 generation. The adverse effects on glucose homeostasis of feeding a PRD during pregnancy in the F0 generation have been found in the offspring up to F3 generation (Benyshek et al. Reference Benyshek, Johnston and Martin2006). The administration of dexamethasone to dams in late pregnancy induced an increased expression of the glucocorticoid receptor (GR) and its target gene phosphoenolpyruvate carboxykinase (PEPCK) in the liver of the F1 and F2 offspring (Drake et al. Reference Drake, Walker and Seckl2005). This effect was transmitted through both the male and female F1 lines. These changes in gene expression were not, however, present in the F3 generation.
The mechanism for the transgenerational transmission of induced phenotypes is not known. Stable changes to gene expression that underlie individual phenotypes are the result of the epigenetic regulation of transcription, which includes DNA methylation and covalent modifications to histones. The methylation of cytosines in CpG dinucleotides in the promoter region of genes permanently suppresses transcription (Bird, Reference Bird2002). Soon after fertilization, the genome undergoes demethylation. This is followed by methylation of the promoters of specific genes in the early embryo (Bird, Reference Bird2002). Such epigenetic gene-silencing is critical for cellular differentiation and is maintained throughout the lifespan. Allele-specific silencing of imprinted genes by DNA methylation is well established (Arnaud & Feil, Reference Arnaud and Feil2005; Lander-Diner & Cedar, Reference Lander-Diner and Cedar2005), and methylation patterns are resistant to demethylation during the early development of the embryo, although the underlying mechanism is unclear (Lane et al. Reference Lane, Dean, Erhardt, Hajkova, Surani, Walter and Reik2003).
We have shown recently that feeding a PRD to pregnant rats resulted in hypomethylation and increased expression of the PPAR-α and GR110 promoters in the liver of the offspring on postnatal day 34 (Lillycrop et al. Reference Lillycrop, Phillips, Jackson, Hanson and Burdge2005). This shows that the induction of different metabolic phenotypes in the offspring by maternal nutrition during pregnancy in non-imprinted genes also involves an altered epigenetic regulation of gene expression. Moreover, in our previous study, this epigenetic change in the PPARα and GR promoters was prevented by supplementing the PRD with folic acid during pregnancy (Lillycrop et al. Reference Lillycrop, Phillips, Jackson, Hanson and Burdge2005), which suggests that altered 1-carbon metabolism is involved in the process of inducing altered DNA methylation.
In the present study, we have tested the hypothesis that the transmission of phenotypes between the F1 and F2 generations involves an altered epigenetic regulation of specific genes. We report the effect of feeding a PRD during pregnancy in the F0 generation on the methylation status and expression of the GR and PPARα promoters, and on the expression of their respective target genes PEPCK and acyl-CoA oxidase (AOX) in the liver of the F1 and F2 offspring.
Materials and methods
Animal procedures
Female Wistar rats (F0) were mated and then fed throughout pregnancy either a reference diet (RD) containing 180 g/kg (w/w) casein or an isocaloric PRD containing 90 g/kg (w/w) casein as described (Langley & Jackson, 1994; Table 1). At delivery, litters were reduced to eight pups. Dams were fed purified AIN-76A diet throughout lactation (Table 1). The offspring were weaned onto AIN-76A 28 d after birth. Male offspring (F1) were killed on postnatal day 80. The livers were removed immediately, frozen in liquid N and stored at − 80°C. Two female F1 offspring were selected from each litter by random removal from the cage and were mated on postnatal day 125 with males that had received adequate nutrition throughout life. F1 females were fed AIN-76A (Table 1) throughout pregnancy and lactation. Litters were reduced to eight at birth, and the F2 offspring were weaned at postnatal day 28. Male offspring were killed on postnatal day 80. The livers were immediately removed, frozen in liquid N and stored at − 80°C. Livers were selected at random for studies of gene methylation and expression by removal from collections of stored specimens without knowledge of any aspect of the phenotype of the offspring. Liver from one offspring from each litter was studied.
Table 1 Composition of the diets fed
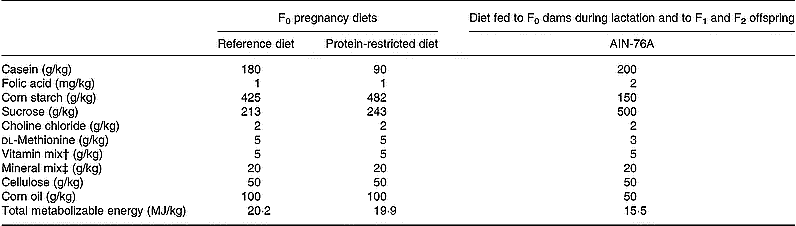
† Vitamin mix: thiamine hydrochloride 2·4 mg/kg, riboflavin 2·4 mg/kg, pyridoxine hydrochloride 2·8 mg/kg, nicotinic acid 12·0 mg/kg, d-calcium pantothenate 6·4 mg/kg, biotin 0·01 mg/kg, cyanocobalbumin 0·003 mg/kg, retinyl palmitate 6·4 mg/kg, dl-tocopherol acetate 79·9 mg/kg, cholecalciferol 1·0 g/kg, menaquinone 0·02 mg/kg.
‡ Mineral mix: calcium phosphate dibasic 11·3 g/kg, NaCl 1·7 g/kg, potassium citrate monohydrate 5·0 g/kg, K2SO4 1·2 g/kg, MgSO4 0·5 g/kg, MgCO3 0·1 g/kg, ferric citrate 0·1 g/kg, ZnCO3 36·2 mg/kg, CuCO3 6·8 mg/kg, KIO3 0·2 mg/kg, sodium selenite 0·2 mg/kg, chromium potassium sulphate 12·5 mg/kg.
Measurement of DNA methylation
The methylation status of the GR110 and PPARα promoters was determined by methylation-sensitive real-time PCR (Lillycrop et al. Reference Lillycrop, Phillips, Jackson, Hanson and Burdge2005). Briefly, genomic DNA (5 μg) was isolated from liver using standard methods and treated with the methylation-sensitive restriction enzymes AciI and HpaII as instructed by the manufacturer (New England Biolabs (UK), Hitchin, Hertfordshire, UK). Purified DNA was then amplified by real-time PCR using the primers listed in Table 2. The reaction was carried out in a total volume of 25 μl with SYBR Green Jumpstart ready mix as described by the manufacturer (Sigma, Poole, Dorset, UK). The promoter region of the rat PPARγ2 gene, which contains no CpG islands and no AciI or HpaII recognition sites, was used as an internal control. There was no effect of maternal diet or generation on the methylation status of the hepatic PPARγ2 promoter. All cycle threshold (Ct) values were normalized to the internal control.
Table 2 PCR primers for analysis of promoter methylation and mRNA expression
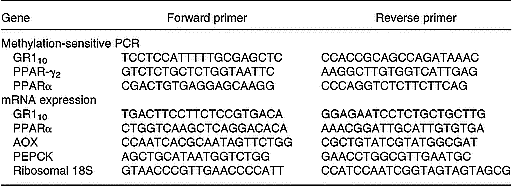
Primers designed by QIAGEN Ltd UK, Crawley, UK.
AOX, acyl-CoA oxidase; GR110, glucocorticoid receptor; PEPCK, phosphoenolpyruvate carboxykinase.
Measurement of mRNA expression
mRNA expression was determined by real-time RTPCR amplification (Harris et al. Reference Harris, White, Phillips and Lillycrop2002). Briefly, total RNA was isolated from cells using TRIZOL reagent (Invitrogen, Paisley, Renfrewshire, UK), and 0·1 μg was used as a template to prepare cDNA using 100 U Moloney murine leukaemia virus reverse transcriptase. The primer sequences are listed in Table 2. The PCR reaction was carried out in a total volume of 25 μl with SYBR Green Jumpstart ready mix as described by the manufacturer (Sigma). mRNA expression was normalized using the housekeeping gene ribosomal 18S RNA using the change in ΔCt method (Bustin, Reference Bustin2000). There was no effect of maternal diet or generation on the mRNA expression of 18S ribosomal RNA.
Statistical comparisons
Normalized Ct values are presented as proportion of the RD group in the F1 generation (mean (sem); n 6 offspring per F0 dietary group, one offspring per litter). Analysis of the covariate by independent variable interaction showed that the homogeneity of the regression slopes could be assumed for each of the genes studied. Therefore, ANOVA was used to assess the effects of diet and generation on promoter methylation status and mRNA expression. The extent of interactions between the diet of the F0 dams and the generation of the offspring was determined by two-way ANOVA. Comparisons of DNA methylation and mRNA expression between F0 dietary groups and generations of offspring were by one-way ANOVA with Dunnett's post hoc test (two-sided) using the RD F1 group as the reference.
Results
DNA methylation
The results of measurements of the methylation status of the GR110 promoter, which is expressed in liver, and of the PPARα promoter are summarised in Fig. 1. Analysis by two-way ANOVA showed that there was no significant effect of generation or interaction between F0 diet and generation on the methylation of the GR110 or PPARα promoters. There was, however, a significant effect of diet (P < 0·001) on the methylation status of both genes. Methylation of the GR110 promoter was significantly lower (P < 0·05) in the liver of the male offspring of the F0 PRD group in the F1 (10·2 %) and F2 (7·9 %) generations compared with the F0 RD group. Methylation of the PPARα promoter was significantly lower in the liver of the male offspring of the F0 PRD group in the F1 (8·2 %) and F2 (10·5 %) generations compared with the F0 RD group. There were no significant differences between the F1 and F2 generations within a F0 maternal dietary group.
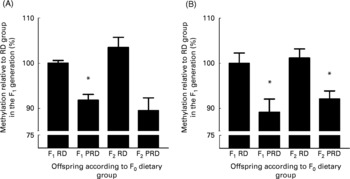
Fig. 1 Effect of maternal protein intake during pregnancy in the F0 generation on hepatic (A) PPARα and (B) glucocorticoid receptor promoter methylation in the F1 and F2 male offspring. Values are means with their standard errors normalized to the offspring of the F1 reference diet (RD) group (six per group). PRD, protein-restricted diet. * Mean values were significantly different between maternal diets within a generation by one-way ANOVA with Dunnett's post hoc test (two-sided) using the RD F1 group as a reference: P < 0·05.
mRNA expression
The results of measurements of mRNA expression are summarised in Table 3. Analysis by two-way ANOVA showed that there was no significant interaction effect between generation and F0 dietary group on the expression of any of the genes measured. Analysis by one-way ANOVA showed that there were no significant differences in the expression of any of these genes between the F1 and F2 generations with an F0 maternal dietary group. There was no significant effect of F0 maternal diet on the expression of PPARα or GR110 in the F1 or F2 generations, although there were trends (P < 0·1) towards higher mRNA expression in the PRD group in the F1 (29 % and 15 %, respectively) and F2 (44 % and 31 %, respectively) compared with the F1 RD group. Two-way ANOVA showed that there was a significant effect of the F0 maternal diet on the expression of AOX (P = 0·016) and PEPCK (P = 0·006). For AOX, one-way ANOVA showed a significant difference between groups, although this did not reach statistical significance in pair-wise comparisons. There were, however, trends (ANOVA P < 0·05) towards higher AOX mRNA expression in the PRD group in the F1 (65 %) and F2 (105 %) generations, although these did not reach statistical significance in pair-wise comparisons (Table 3). PEPCK expression was significantly (P < 0·05) greater in the PRD group in the F1 (59 %) and F2 (73 %) generations (Table 3).
Table 3 Measurements of hepatic mRNA expression (Mean values and their standard errors for six male offspring in each F0 dietary group in each generation)
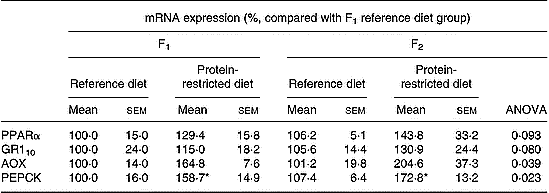
AOX, acyl-CoA oxidase; GR110, glucocorticoid receptor; PEPCK, phosphoenolpyruvate carboxykinase.
* Mean values were significantly different between maternal diets within a generation by one-way ANOVA with Dunnett's post hoc test (two-sided) using the reference diet F1 group as a reference: P < 0·05.
Discussion
The results of this study show for the first time that the altered methylation of gene promoters induced in the F1 offspring by maternal protein restriction during pregnancy is transmitted to the F2 offspring.
The majority of studies on the induction of an altered metabolic phenotype by maternal dietary restriction in humans and in experimental models have focused on the first-generation offspring. There is, however, evidence from epidemiological studies (Stein & Lumey, Reference Stein and Lumey2000; Painter et al. Reference Painter, Roseboom and Bleker2005), and in particular from animal models (Martin et al. Reference Martin, Johnston, Han and Benyshek2000; Torrens et al. Reference Torrens, Brawley, Dance, Itoh, Poston and Hanson2002; Zambrano et al. Reference Zambrano, Martinez-Samayoa, Bautista, Deas, Guillen, Rodriguez-Gonzalez, Guzman, Larrea and Nathanielsz2005; Benyshek et al. Reference Benyshek, Johnston and Martin2006), that the phenotype induced in the offspring of the F1 generation can be transmitted to subsequent generations.
We have previously shown that an increased expression of GR110 and PPARα in the liver of the offspring on postnatal day 34 as a result of feeding a PRD in pregnancy is due to hypomethylation of the respective gene promoters (Lillycrop et al. Reference Lillycrop, Phillips, Jackson, Hanson and Burdge2005). Our current findings show that the methylation status of the GR110 and PPARα promoters was reduced in the F1 and F2 offspring of the F0 PRD dams. One possible explanation is that the level of methylation of the GR110 and PPARα promoters was set during the development of the F1 generation and that this is maintained through gamete production, through demethylation of the maternal and paternal genomes after fertilization and during gene specific remethylation in the early embryo. If so, this implies that the process which results in the hypomethylation of GR110 and PPARα in the liver also induces a stable reduction of methylation of these genes in the germ cells. As the F1 females, but not the males with which they were mated, had been exposed to nutritional constraint during pregnancy, our findings suggest that the transmission of GR110 and PPARα hypomethylation must have occurred via the female genome and that this was sufficient to alter the methylation of the promoters of these genes in the livers of the F2 males. Studies of the expression of intracisternal A-type particles show that the methylation of these repetitive sequences is resistant to demethylation during preimplantation development, and it has been suggested that such resistance may explain the inheritance of patterns of gene imprinting (Lane et al. Reference Lane, Dean, Erhardt, Hajkova, Surani, Walter and Reik2003). If the level of methylation of non-imprinted genes was also ‘protected’ during post-fertilization demethylation, this might explain how patterns of GR110 and PPARα methylation induced in the F1 generation may be transmitted to the F2 generation.
One alternative explanation is that prenatal undernutrition induced changes in the F1 females that constrained the intrauterine environment experienced by the F2 male offspring, and that hypomethylation of the PPARα and GR110 promoters was thus induced de novo in the male offspring in each generation. This seems unlikely because of the similarity in the degree of hypomethylation induced in the F1 and F2 generations. It might be anticipated that if promoter hypomethylation were induced de novo in each generation, it would result in different levels of methylation because of differences in the degree of environmental constraint. However, a single environmental challenge in the F0 generation might be expected to induce a similar level of promoter methylation in both generations if the effect on the F1 generation were transmitted to the F2 generation.
Feeding a PRD during pregnancy in the F0 generation induced a trend towards an increased expression of PPARα and GR110, and their respective target genes AOX and PEPCK, in the liver of the F1 and F2 offspring at day 80, although only the increase in PEPCK expression reached statistical significance. The trend in the expression is consistent with reduced methylation of the GR110 and PPARα promoters. We have previously shown that feeding a PRD during pregnancy significantly increased the expression of PPARα, GR110 and AOX in the liver of the F1 offspring on postnatal day 34 due to hypomethylation of the GR110 and PPARα promoters (Lillycrop et al. Reference Lillycrop, Phillips, Jackson, Hanson and Burdge2005), and others have shown increased gluconeogenesis in this model (Burns et al. Reference Burns, Desai, Cohen, Hales, Iles, Germain, Going and Bailey1997).
One possible explanation for the difference between our previous report (Lillycrop et al. Reference Lillycrop, Phillips, Jackson, Hanson and Burdge2005) and the present study in the extent to which hypomethylation of the GR and PPARα promoters altered the expression of GR110 and PPARα, and of their target genes AOX and PEPCK, is that the transcription of PPARα and GR is responsive to environmental stimuli such as dietary fat intake and stress, respectively. In the absence of a dietary or stress challenge, the elevated levels of transcription found in recently weaned animals (Lillycrop et al. Reference Lillycrop, Phillips, Jackson, Hanson and Burdge2005) may have diminished by day 80. For example, in the rat, hepatic PPARα expression decreases after weaning due to the reduction in fat intake (Panadero et al. Reference Panadero, Herrera and Bocos2000). In addition, PPARα expression is less sensitive to dietary fat intake in adult liver than neonates (Panadero et al. Reference Panadero, Herrera and Bocos2005). Nevertheless, feeding the PRD diet to F0 dams induced in the F1 and F2 offspring the potential for an exaggerated response to stress or dietary fat.
These findings suggest that the transmission of an altered metabolic phenotype as a result of prenatal nutritional constraint to at least one subsequent generation is the result of an induction of altered epigenetic regulation of gene expression in both the F1 and F2 generations. This may also explain the transmission of induced phenotypes from the F1 to F2 generation in other experimental systems, such as increased hepatic GR expression and PEPCK activity as a result of exposure of the F0 dams to dexamethasone in late gestation (Drake et al. Reference Drake, Walker and Seckl2005). If this occurs in humans, as indicated by epidemiological studies (Stein & Lumey, Reference Stein and Lumey2000; Painter et al. Reference Painter, Roseboom and Bleker2005), the findings would suggest that the nutrition of pregnant women has a critical impact not only on the health of their children, but also on subsequent generations.
Acknowledgements
G. C. B. and M. A. H. are supported by the British Heart Foundation, which also funded part of this study.




