


Introduction
The Mariana Crow or Aga Corvus kubaryi, is native to the islands of Guam and Rota, Mariana Islands. It was listed as an endangered species in 1984 (U.S. Fish and Wildlife Service 1984) and is currently considered ‘Critically Endangered’ (IUCN 2015). The introduction of the brown tree snake Boiga irregularis on Guam in the 1940s decimated the crow population, along with all other native forest birds and wild crows can no longer be found there (Savidge Reference Savidge1987).
The Mariana Crow population on Rota has also experienced significant declines since the 1980s. A 1982 estimate put the population at around 1,300 individuals (Engbring et al. Reference Engbring, Ramsey and Wildman1986), and a second in 1995 set it at 600 individuals; both estimates used variable circular plot (VCP) methods (Fancy et al. Reference Fancy, Lusk and Grout1999). A count conducted from 1996 to 1998 estimated 117 breeding pairs, based on direct count (85 pairs) and extrapolation (32 pairs) into unsearched, suitable habitat (Plentovich et al. Reference Plentovich, Morton, Bart, Camp, Lusk, Johnson and Vanderwerf2005). Another count in 2007 revealed only 60 pairs, using only direct count methods (Zarones et al. Reference Zarones, Sussman, Morton, Plentovich, Faegre, Aguon, Amar and Ha2015). In 2012, the population was estimated at between 30 and 202 individuals using VCP methods (Camp et al. Reference Camp, Brinck, Gorresen, Amidon, Radley, Berkowitz and Banko2015). In contrast to the situation on Guam, there is not a clear, single reason for the decline of crows on Rota. The cause could include one or more factors including habitat loss and degradation, human disturbance and direct persecution, habitat alteration by typhoon activity and introduced deer, introduced predators such as cats, rats and monitor lizards, as well as the deleterious effects of inbreeding (Morton et al. Reference Morton, Plentovich and Sharp1999, Tarr and Fleischer Reference Tarr and Fleischer1999, U.S. Fish and Wildlife Service 2005). Endemic island species often lack antipredator behaviours and thus suffer from the introduction of non-native predators (Blackburn et al. Reference Blackburn, Casey, Duncan, Evans and Gaston2004), which are one of the most prominent causes of the decline and extinction of island species worldwide (Vitousek et al. Reference Vitousek, Mooney, Lubchenco and Melillo1997, Salo et al. Reference Salo, Korpimäki, Banks, Nordström and Dickman2007).
A recent telemetry study suggests that predation by feral cats may be a significant cause of mortality (Ha et al. Reference Ha, Faegre and Ha2014). In response to these initial findings, a predator control programme was established on Rota in early 2012. In view of this step and other current and proposed population management efforts we aimed to update the island-wide pair count on Rota so that the efficacy of management efforts can be more accurately assessed in the coming years.
Methods
The study was conducted on the island of Rota, Commonwealth of the Northern Mariana Islands, situated in western Micronesia (14°09’N, 145°12’E). Rota is 86 km2 in area and volcanic in origin with terraces of uplifted coral limestone. The climate is tropical with high humidity and warm year-round temperatures.
During the 2013–2014 breeding season we conducted a direct count of all territorial pairs. This method has been successfully used previously to estimate the population of Mariana Crows due to their high nest site fidelity (Plentovich et al. Reference Plentovich, Morton, Bart, Camp, Lusk, Johnson and Vanderwerf2005, Zarones et al. Reference Zarones, Sussman, Morton, Plentovich, Faegre, Aguon, Amar and Ha2015). We searched all territories identified during the September 2007–April 2008 pair survey (Zarones et al. Reference Zarones, Sussman, Morton, Plentovich, Faegre, Aguon, Amar and Ha2015) as well as all new territories identified in the years since (Figure 1). The one exception was the lowest terrace of the Rail Trail area (Figure 1), which had difficult access and a sensitive bat colony, and was determined to have only one pair during the 2007 survey (Zarones et al. Reference Zarones, Sussman, Morton, Plentovich, Faegre, Aguon, Amar and Ha2015). We excluded some areas of the island that were deemed unsuitable habitat for crows; these included the residential and agricultural areas of the Songsong Peninsula, Sinapalo area and much of the Duge region (Figure 1). We did not survey the Sabana plateau because former surveys have established that crows rarely occur there (Fancy et al. Reference Fancy, Lusk and Grout1999), nor did we survey some privately owned lands with restricted access. Finally, areas in which no pairs were found during the 2007 survey, were excluded due to manpower constraints and the low probability of finding pairs. In these unsearched areas we estimated the maximum number of pairs that could have been missed based on the density of pairs found in nearby, searched habitat.
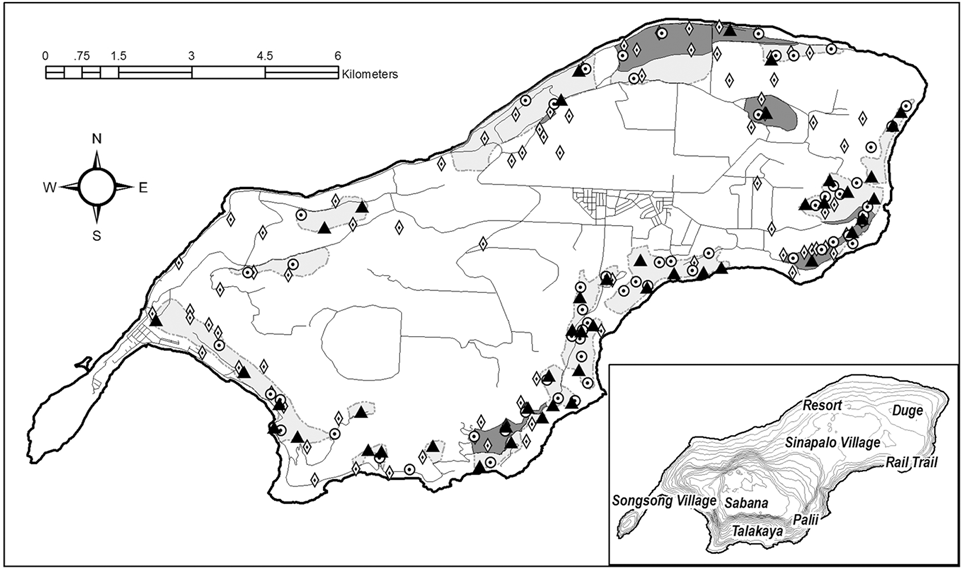
Figure 1. Locations of 2013 pairs represented as black triangles, 2007 pairs as circles and 1998 pairs as diamonds. Areas covered in the search effort are shaded light grey, while areas of overlap with the 1998 and 2007 surveys are dark grey. The location for each pair depicted was either the last nest location from the season, for nests that were found, or a point central to the observed locations of the pair, if no nest was found. Inset: Topographical map of Rota with location names.
We spent 845 person-hours searching and confirming pair locations and pair status from 8 September 2013 to 14 April 2014, not including time spent monitoring active nests. Search effort covered approximately 1,485 ha of forest, of which approximately 329 hectares overlapped with areas searched with similar methods in the 1996–1998 and 2007 surveys (Figure 1) (Plentovich et al. Reference Plentovich, Morton, Bart, Camp, Lusk, Johnson and Vanderwerf2005; Zarones et al. Reference Zarones, Sussman, Morton, Plentovich, Faegre, Aguon, Amar and Ha2015). The most intensive searching coincided with the peak period of the crow’s breeding season from October to February (Zarones et al. Reference Zarones, Sussman, Morton, Plentovich, Faegre, Aguon, Amar and Ha2015). During this time, each former territory was visited at least once every 4–6 weeks regardless of its occupancy status. We worked in a counter-clockwise manner around the island and attempted to visit multiple adjacent territories on the same day, with multiple searchers whenever possible, to reduce the chance that a pair straddling two formerly known territories would be double-counted.
Searchers covered each area on foot; the standard search was 1.5–2 hours or until crows were detected, whichever was shorter. Searches were not conducted during heavy rain or wind. Playbacks of crow location calls were used when there were no recent detections in an area. Searchers attempted to determine the status of each pair located during a visit (nesting, with fledgling, etc.), but if the status could not be determined, repeat visits were made to the territory to confirm status. If a pair behaved territorially (agitation, defensive calls, bill pounding, stick throwing, etc.), but was not actively nesting, the territory was visited again after 4–6 weeks. If a pair was found to be nesting, a nest monitoring protocol was used and that territory was not searched for additional occupancy until the nest was determined to have fledged or failed. After a nest attempt terminated, the area was placed back on the regular schedule of searches every 4–6 weeks, to reduce the chance of double counting if that pair later moved to a vacant neighbouring territory.
In the absence of detected nests or young fledglings, individual pairs were identified on the basis of multiple factors: the presence of colour-banded crows, family groups with older fledglings, consistency of sighting locations, observed territorial (defensive calls, bill pounding, stick throwing, etc.) or pair-like behaviour (mate feedings, carrying sticks, etc.) and timing of crow observations in neighbouring areas (Zarones et al. Reference Zarones, Sussman, Morton, Plentovich, Faegre, Aguon, Amar and Ha2015).
While both the 1996–1998 and the 2007 counts included pairs located outside the former study plots, uncertainty about the comparability of search methods used in these outer areas spurred us to compare areas that were known, based on the available literature, to be searched using the most similar methods. To compare pair estimates across years we overlaid the study plots searched in the two former pair surveys (Plentovich et al. 2005, Zarones et al. Reference Zarones, Sussman, Morton, Plentovich, Faegre, Aguon, Amar and Ha2015) with the area searched during this survey, then counted pairs occurring in the areas of overlap (Figure 1). Because the Plentovich et al. (Reference Plentovich, Morton, Bart, Camp, Lusk, Johnson and Vanderwerf2005) study included pairs found over the course of three years within their study plots, we choose to use the number of pairs found only during the latest year, 1998, as a snapshot to compare both this survey and the 2007 survey which were each conducted in one season. Pair density was calculated across years by dividing the number of pairs that occurred in a region (north or south) by the hectares searched in that region that year. For these purposes the north is considered the entire north or northwest-facing shoreline from Songsong Village to the easternmost point of the island, including the Duge homestead area.
To provide an estimate of the population size of Mariana Crows on Rota that was inclusive of non-breeding subadult and adult birds, we recorded all subadult and adult banded and unbanded birds seen throughout the island during the 845 person-hour search. We used the Chapman estimator analysis on these data to acquire a total population size that was unbiased due to small sample size (Chapman Reference Chapman1951).
Results
We identified a total of 46 Mariana Crow pairs on Rota during the 2013–2014 breeding season (Figure 1). In addition, there were nine unpaired banded crows (six adults, three subadults) resighted on separate occasions, at least once during the season. Thirty-three pairs were detected actively engaged in a nesting attempt (any nest stage from building through fledge) and four pairs were found with young fledglings although the actual nest had not been found in time. A total of 20 fledglings from 16 pairs were confirmed (Table 1).
Table 1. Summary of 46 crow pairs and their status during the period September 2013-April 2014. Shows the highest achieved nesting status of all crow pairs found (i.e. non-breeding pairs, pairs with failed breeding attempts, or pairs that fledged young). *Two fledglings taken into captivity due to poor condition at time of fledging.

In a comparison of pair loss across the three surveys (1998, 2007 and 2013) and between north and south regions of the island we found that the three surveys all searched somewhat different, but overlapping areas (Figure 1). If we limit comparisons to direct counts in those areas searched by the same methods, the number of pairs in the North decreased across the survey years from 16 to eight (Table 2). Given that 77% more hectares were surveyed in 2013, the decline in density of pairs is even more dramatic. In the south, the number of pairs counted increased between 1998 and 2013, but, because the number of hectares searched was four times greater, the density still declined (Table 2). Furthermore, in those areas of overlap that were searched in all three surveys using the same methods, the density of pairs has been consistently higher in the south than in the north (Table 3; Figure 1). The sample sizes are too small to conclude that losses from 1998 to 2013 were any greater in the north than in the south (Table 3).
Table 2. Pair counts and density of pairs found in 1998 and 2007 within the former study plots (see Plentovich et al. Reference Plentovich, Morton, Bart, Camp, Lusk, Johnson and Vanderwerf2005) and all area intensively searched in 2013.

Table 3. Pair counts and densities limited to geographically overlapping areas searched in all surveys 1998–2013.

We saw 129 total subadult and adult banded (n = 23) and unbanded crows (n = 106) during the 845 person-hour search. Based on banding records and resightings from May 2012–May 2015 of birds banded prior to 8 September 2013, we determined that there were 32 banded subadult and adult birds that were available to be seen from 8 September 2013 to 14 April 2014, of which we detected 23/32, or 71.86%. Based on the Chapman estimator analysis, the total population size of subadult and adult Mariana Crows on Rota is thus 178 (Chapman Reference Chapman1951).
Discussion
A number of different methods have been used to estimate the Mariana Crow population on Rota. Estimates in 1982, 1995, 1998 and 2012 used variable circular plot (VCP) surveys island-wide to estimate the total number of individuals (Engbring et al. Reference Engbring, Ramsey and Wildman1986, Fancy et al. Reference Fancy, Lusk and Grout1999, U.S. Fish and Wildlife Service 2005, Camp et al. Reference Camp, Brinck, Gorresen, Amidon, Radley, Berkowitz and Banko2015). Comparisons of VCP surveys have shown an estimated population decline of 84–97% from 1982 to 2012 (Enbring et al. Reference Engbring, Ramsey and Wildman1986, Camp et al. 2012). A comparison of the population estimates based on direct count methods shows a decline of approximately 49% from 1998 to 2007, and an additional reduction of around 10–23% from 60 pairs in 2007 to 46–54 pairs in 2013 (Zarones et al. Reference Zarones, Sussman, Morton, Plentovich, Faegre, Aguon, Amar and Ha2015). When all three direct count surveys are compared there is an approximately 46–53% decline from 1998 to 2013.
The 2007 survey observed substantial pair losses in the northern study areas when compared to the 1998 study, but relatively stable pair numbers in the Rail Trail and Palii study areas (Zarones et al. Reference Zarones, Sussman, Morton, Plentovich, Faegre, Aguon, Amar and Ha2015). Mariana Crow studies on Rota in the 1990s restricted most search effort to six study plots, four of which were on the north side of Rota, and pairs were located outside these plots somewhat opportunistically over the course of 10 years (Plentovich et al. Reference Plentovich, Morton, Bart, Camp, Lusk, Johnson and Vanderwerf2005). Searches in more recent years have included much more of the south side of the island (Table 2). Our comparisons of pair densities across the three direct-count surveys suggests that the crow population has typically been denser in the south than the north. However, even the 1998 survey occurred after Rota population declines had been detected, and may not fully represent the natural density and distribution of the population across the island prior to the decline. The apparently greater decline in the north between 1998 and 2007, was not seen between 2007 and 2013. In fact, the declines between the north and south are very similar for 2007–2013.
These density differences could be due to a number of factors, including habitat loss from development or degradation by typhoons, reduced habitat connectivity, and persecution by humans in the more accessible northern parts of the island. The north and north-eastern portions of the island consist largely of privately owned land, or have been slated for homestead development. Although there has been limited direct evidence of persecution, when the Mariana Crow was listed as endangered in 1984 much of the homestead development was halted, and this is still a contentious issue with some residents (Morton et al. Reference Morton, Plentovich and Sharp1999, Sussman et al. Reference Sussman, Ha and Henry2015). Further, recent estimates of forest loss only account for broad scale changes for the island as a whole and no studies have investigated differences in predator abundance across the island (U.S. Fish and Wildlife Service 2005, Camp et al. Reference Camp, Brinck, Gorresen, Amidon, Radley, Berkowitz and Banko2015).
Despite efforts to locate all pairs, crows are not always detectable, particularly when they are not calling, and they are not always responsive to playbacks. An established pair with a known territory can wander up to 1 km away even during the breeding season, meaning that pair would have gone undetected if the area was searched only once (pers. obs.). This is why we conducted multiple searches in each area in order to consider it sufficiently searched. In addition, due to limitations in manpower, some areas where pairs were identified in 1998 were not searched sufficiently, or at all, during this survey. Some areas were not searched due to the presence of sensitive fruit bat colonies, such as the upper reaches of the Palii area (Figure 1). Others had minimal or no detections during the 2007 survey, such as the Resort area and the eastern half of the Duge area, and still others were not searched at all in 2007 with the exception of the coverage provided by the quarterly road survey (Berry et al. Reference Berry, Ha and Ha2008). Given limited manpower, exploratory searches into these areas were left at a minimum. Therefore, it is possible that additional pairs occur there. Based on the densities found in the surrounding areas, no more than an additional eight pairs could be added to the total of 46 detected pairs. This estimate is consistent with a follow-up survey done during the 2015 breeding season in which 50 pairs were found in an expanded search effort with a 55% increase in person-hours and 425 more hectares covered. Forty-seven of the 50 pairs were found in areas that had also been searched in 2013 (Faegre et al. Reference Faegre, Ha, Hubl, Ware and Wiitala2016).
The 2013 pair survey estimated a population of up to 54 pairs of crows, or 108 adults. Our mark-resight population estimate using the Chapman index is 178 birds. Given the low end of the pair estimate and not counting sub-adult or fledgling birds, the adult population is between 92 and 108 individuals based on the pair survey. Subtracting the median of that range, or 100, from the Chapman estimate (178), gives an additional 78 individuals that may not have been directly detected in the pair survey. Many of these are likely to be subadult individuals (floaters), as well as some unobserved pairs.
Conclusions
Even at the upper limit of this range of 46–54 pairs, the pair count is still much lower than the 75 pairs needed to maintain a viable population on Rota (U.S. Fish and Wildlife 2005), and the total population size is below 180. We therefore recommend that predator control continue on the island of Rota and that a pair count be repeated with greater frequency to better determine the effectiveness of these management efforts. An increase in colour banding efforts, so that pairs can be more individually identifiable, would also aid in monitoring demographics of the population as a whole. Those conducting pair counts should keep track of search effort in terms of person-hours, hectares searched and frequency of visits to allow better comparisons between years.
Acknowledgements
Work was conducted in accordance with Federal Fish and Wildlife Permit TE09155B-1, Federal Bird Banding Permit number 22802 and University of Washington Institutional Animal Care and Use Committee protocol number 2858-04. We thank Laura Bussolini, Sarah Faegre, Henry Fandel, Phil Hannon, Mike Hitchcock and Dacia Wiitala for their assistance in the field, and Manny Pangelinan, Director, Commonwealth of the Northern Mariana Islands, Department of Lands and Natural Resources (DLNR), Division of Fish and Wildlife, and DLNR Secretary, Richard B. Seman, for their support of our work in the Marianas. We thank Anne B. Clark and an anonymous reviewer for their helpful critiques on earlier drafts. RRH thanks the Whiteley Center at Friday Harbor Laboratories, University of Washington, which provided support for working on this manuscript. This work was supported by the Commonwealth of the Northern Mariana Islands Department of Lands and Natural Resources-Division of Fish and Wildlife from U.S. Fish and Wildlife Section 6 Endangered Species funds, grant F12AP00963.




