


INTRODUCTION
Non-typhoidal salmonellosis is a common foodborne infection. In most cases humans are infected through consumption of raw, undercooked and faecally contaminated foods, mostly of animal origin [Reference Hald1–Reference Pires3]. Over 2500 serovars of Salmonella have been identified and the majority can produce disease in animals and humans. Poultry can carry a significant number of them without showing any clinical signs of infection [Reference Hald1, Reference Poppe, Wray and Wray4]. S. Enteritidis and S. Typhimurium have been the most frequently isolated serovars worldwide [Reference Fashae5–Reference Al-Zenki7], and especially S. Enteritidis, phage type (PT) 4, has emerged with a pandemic occurrence in both poultry and humans since the early 1980s [Reference Liebana8]. However, the occurrence of PTs is a dynamic situation, and the PTs of S. Enteritidis differ between geographical areas [Reference Hasenson9]. Recently, the serovar S. Kentucky has been reported to be more prevalent than S. Enteritidis in poultry around the world [Reference Foley6, Reference Barua10, Reference Ibrahim11].
Salmonella strains resistant to antimicrobial drugs are now widespread due to selection from the use of antimicrobials [Reference Wouafo12, Reference Solghan13]. The use of antimicrobials in animal feed and the indiscriminate use in both humans and animals have created selection pressure that favours increased bacterial resistance. In developed countries, it has been reported that one of the sources of this increased resistance in Salmonella is of zoonotic origin, where bacteria in the food-animal hosts acquire resistance before onward transmission to humans through the food chain [Reference Threlfall14], e.g. S. Kentucky strains isolated from poultry have been identified as being resistant to antimicrobials commonly used in both veterinary and human medicine [Reference Li15]. In the past decade, the emergence of multidrug-resistant non-typhoidal Salmonella strains, including isolates resistant to quinolones, has also been reported with increasing frequency in several African countries [Reference Mandomando16, Reference Wilkens17].
In Ghana, up-to-date information on the occurrence of Salmonella in poultry is not available. The most recent reports originate from 2001 where Salmonella was isolated from the gut content and carcasses of poultry and showed multidrug resistance, leading to the conclusion that locally produced chicken are a potential source of multiple antimicrobial resistant enteropathogenic bacteria [Reference Mandomando16, Reference Sackey18]. Local chicken and broilers are an important food source and form part of most dishes during daily life and festive periods in most parts of the country, but the industry is largely unregulated, with a high use of antimicrobials as a consequence [Reference Donkor, Newman and Yeboah-Manu19].
There is a need for baseline information on the occurrence of Salmonella serovars and their antimicrobial resistance in the poultry production systems in Ghana, in order to enable an assessment of the link between human salmonellosis and consumption of chicken and eggs. The objective of this study was therefore to determine the prevalence of Salmonella serovars in egg-laying hens and broilers and their susceptibility to antimicrobials commonly used in poultry and humans in Ghana. Molecular subtyping was conducted using pulsed-field gel electrophoresis (PFGE) to assess the genetic relatedness of common serovars obtained from different locations in order to estimate whether widespread serovars are associated with clonal spread of bacteria.
MATERIALS AND METHODS
Sample collection and interviews
A cross-sectional study was performed in two main poultry-producing regions with typical poultry production systems of Ghana: Accra, the capital city and Kumasi, the biggest commercial city. Farms were randomly picked from lists of producers in the two regions; however, farmers had to give consent for the sampling to take place. The final sample consisted of 20 poultry farms from Accra and 55 farms from Kumasi.
A local poultry slaughter facility in the central market in Kumasi was also included to increase the number of flocks investigated. The central market has live-bird markets which handle between 300 and 500 birds during the week and about 800–1000 birds either slaughtered upon the customer's request or sold alive during weekends.
A total of 200 samples were collected for Salmonella analysis comprising 75 faecal sock, 75 dust, 10 feed and 10 poultry drinking-water samples from poultry farms and 30 slaughterhouse samples representing birds from flocks (n = 6) not included in the farm sampling. Samples were obtained between August 2011 and November 2012.
One faecal sample from each poultry farm (n = 75) was obtained using pairs of socks, an alternative method that has shown high sensitivity for recovering of Salmonella compared to collecting faecal litter samples directly in farmhouses [Reference Gradel, Andersen and Madsen20]. The majority of birds on the farms and market comprised of early layers (~20 weeks); however, in some flocks more mature birds were included in the sampling. At each sampling time, socks (elasticated nurses round cap, Shanghai Channelmed Import and Export Co. Ltd, China) soaked in normal saline (0·90%) was worn over the farmers' boots upon entering each pen. After moving in a ‘figure-of-eight’-like pattern around the pen perimeter, the socks were removed, turned aseptically and placed in labelled sterile plastic bags.
Dust from surfaces of pen fences and cages were sampled in all flocks using saline moistened sterile nurse caps to gather one dust sample from each of the farms. The caps were then placed in individually labelled sterile plastic bags. From 10 randomly selected farms in Kumasi, 10 g feed from feeding troughs were collected using a sterile spatula and placed into sterile bags and 10 ml water from the flock's drinking water troughs was also collected into sterile plastic containers and labelled.
At the slaughter area in Kumasi central market, ~10 g portions of the skin neck region were obtained from birds after plucking of feathers with a pair of sterile scissors. Then 10 ml of chicken carcass rinse water was collected into sterile plastic containers by directly pouring from the aluminium wash basin used for washing the bird before dressing, and the open cut neck region of the slaughtered birds were also swabbed using sterile Cary Blair swabs (Copan Diagnostics Inc., USA). Selection of birds could not be randomized as it was based on consent from buyers of the birds. All sample types were transported to the laboratory at the Kwame Nkrumah University of Science and Technology in Kumasi in a box insulated with ice for Salmonella analysis which was initiated within 3–4 h after sampling.
A structured interview questionnaire was used to obtain information from poultry farm-owners or caretakers. Subjects covered included antimicrobials used for the last month for the particular flock that was sampled, frequency of usage, drug dispensing system, withdrawal periods and knowledge on antimicrobial resistance. The interviews were done face to face by the principal investigator in the local dialect.
Isolation and identification of Salmonella
Isolation and identification of Salmonella were done according to a standard ISO method [21]. All dust, feed and sock (nurse cap) samples were individually placed into sterile plastic containers with a lid after which 225 ml buffered peptone water (BPW) (CM0509; Oxoid Ltd, UK) was added and incubated at 37 °C for 18 h. Flock drinking water, cuts of neck skin, swabs of neck region and carcass rinse water samples were, however, inoculated into 45 ml BPW and incubated at 37 °C for 18 h. Next, 0·1 ml pre-enrichment culture was spread in triplicate onto selective modified semi-solid Rappaport-Vassiliadis (MSRV) (CM0910, Oxoid) supplemented with novobiocin (SR0161E, Oxoid). The plates were incubated at 41·5 °C for 24 h. If the MSRV plates did not show any growth after 24 h, the plates were incubated for further 24 h. After incubation, a loop was dipped into any swarming growth observed on the MRSV plates and inoculated onto the selective solid medium xylose-lysine-deoxycholate agar (XLD) (CM0469, Oxoid) and incubated at 37 °C for 24 h.
All agar plates were examined for typical Salmonella colonies and suspected colonies were confirmed by biochemical tests (Minibact-E, SSI, Denmark). One isolate per sample with a typical biochemical profile of Salmonella was confirmed by slide agglutination test using polyvalent antisera (Poly A-E + Vi, SSI). All agglutinating isolates were serotyped according to White Kauffmann–Le Minor scheme [Reference Grimont and François-Xavier22] at the Istituto Zooprofilattico Sperimentale delle Venezie (IZSVe), Italy.
Phage-typing
Phage-typing of S. Typhimurium (n = 1) and S. Enteritidis (n = 10) isolates was done with a panel of phages according to the PHLS Colindale London scheme [Reference Baggesen23] at the IZSVe. Strains showing a pattern that did not conform to any recognized phage type were designated ‘not conforming to recognized definitive type’ while those that did not show any pattern at all were designated ‘not typable’.
Antimicrobial susceptibility testing
Antimicrobial susceptibility of the Salmonella isolates was determined by the agar disk diffusion method on Mueller–Hinton agar (CM0337, Oxoid) according to the protocol and guidelines of the European Committee on Antibiotic Susceptibility Testing (EUCAST). Testing of all strains was performed twice in two different laboratories, and strains that obtained different classification in the two laboratories were subjected to a third round of testing. The strains were tested for their resistance to the following antimicrobials: ampicillin (AMP, 10 µg), cefotaxime (CTX, 30 µg), cefoxitin (FOX, 10 µg), gentamicin (GEN, 10 µg), ceftazidime (CAZ, 30 µg), tetracycline (TET, 15 µg), chloramphenicol (CHL, 10 µg), trimethoprim (TMP, 5 µg), sulfamethazole (SUL, 5 µg), nalidixic acid (NAL, 30 µg), and ciprofloxacin (CIP, 5 µg) (Rosco Diagnostica Neo-Sensitabs, Denmark). E. coli ATCC 25922 and Pseudomonas aeroginosa ATCC 27 853 were used for quality control. Inhibition zone diameters were recorded and interpreted according to Neo-Sensitabs breakpoints (2013) by Rosco Diagnostica.
The minimum inhibitory concentrations (MICs) of 14 randomly selected strains were determined at the IZSVe, using broth dilution method (Sensititre; EUMVS2 and 1-ESBL). Antimicrobials tested and breakpoint values for resistance were: AMP (>8 µg/ml), CTX (>0·5 µg/ml), FOX (>8 µg/ml), GEN (>2 µg/ml), CAZ (>2 µg/ml), TET (>8 µg/ml), CHL (>16 µg/ml), TMP (>2 µg/ml), SUL (>512 µg/ml), NAL (>16 µg/ml), and CIP (>0·064 µg/ml). Strains were classified as resistant or susceptible according to the epidemiological cut-off by EUCAST.
Comparison of Salmonella strains by PFGE genotyping
PFGE was done essentially following the standardized CDC PulseNet protocol [Reference Ribot24] to establish the genetic diversity and relatedness among 41 isolates belonging to the four dominant Salmonella serovars: Kentucky (n = 14), Enteritidis (n = 8), Muenster (n = 8) and Nima (n = 11). Overnight culture of bacteria grown in Luria–Bertani broth (240 230; Difco, USA) was used to prepare genomic DNA. Plugs were produced using 1% agarose (SeaKem gold agarose; Lonza, USA) and the DNA embedded in the agarose was digested using 20 000 U/ml of restriction endonuclease XbaI (R0145; New England BioLabs, USA) for 2 h at 37 °C. The DNA fragments were isolated by electrophoresis in 0·5× Tris-borate-EDTA (TB) buffer using the CHEF-DR III system (Bio-Rad Laboratories, USA) at 14 °C with initial switch time of 2·2 s, final switch time of 54·4 s, current 6 V/cm, included angle 120° and run time of 19 h. S. Braenderup (H9812) was used as the reference strain and a low range marker (NO350S; New England BioLabs) was used as the size marker. The gel was stained with 1% ethidium bromide (E1510; Sigma-Aldrich, USA) solution for 30 min and destained in deionized water twice with 30-min intervals. The gel image was captured by GelDoc EQ system with Quantity One software v. 4.2.1; Bio-Rad Laboratories). Phylogenetic analysis of band patterns was done using TIFF files analysed with GelCompar II v. 4.6 software (Applied Maths, Belgium). Dice coefficient with a band position tolerance of 1% and 0·5% optimization level were used to determine similarity between fingerprints. The unweighted pair-group method with arithmetic averages (UPGMA) was applied to produce the dendrogram. Any difference in the DNA restriction pattern of the isolates was interpreted as a type.
Ethical statement
The owner of each poultry farm was informed of the study purpose and oral permission was obtained before sampling. Likewise, samples from the Kumasi central market live-bird slaughter area were obtained with oral permission from the seller on condition that farmers and sellers were treated as anonymous units. Participants consent was documented by responding to the questionnaires. The protocol for the study was approved by the University of Ghana ethics review board (Protocol identification number: MS-EI/M.11-P.4.Ll20tL-L2).
Statistical analysis
All data were entered into a spreadsheet of Microsoft Excel (Microsoft Corp., USA) and transferred to SPSS v. 16 for Windows (SPSS Inc., USA). This was used for the analysis of data collected in the questionnaire interviews. Ninety-five percent confidence intervals (95% CI) for prevalence of Salmonella-positive farms and for strains found to be resistant to ⩾3 antimicrobials were calculated as
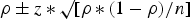 $\rho \pm z\,{\ast}\,\surd {[ {\rho \,{\ast}\,( {1 - \rho })/n}]} $
, where ρ is the estimated prevalence, n is the population size and z = 1·96. In determining the flock prevalence, a flock was considered positive if just one sample from that flock was found positive for Salmonella. χ
2 was used to analyse for differences in flock prevalence between geographical areas, between farms of different size, between farms having different antibiotic use practices and farms that complied with withdrawal periods (for meat selling) and farms that did not. χ
2 was also used to test whether samples that were positive for S. Kentucky were more often sock samples than dust samples. P < 0·05 was considered significant.
$\rho \pm z\,{\ast}\,\surd {[ {\rho \,{\ast}\,( {1 - \rho })/n}]} $
, where ρ is the estimated prevalence, n is the population size and z = 1·96. In determining the flock prevalence, a flock was considered positive if just one sample from that flock was found positive for Salmonella. χ
2 was used to analyse for differences in flock prevalence between geographical areas, between farms of different size, between farms having different antibiotic use practices and farms that complied with withdrawal periods (for meat selling) and farms that did not. χ
2 was also used to test whether samples that were positive for S. Kentucky were more often sock samples than dust samples. P < 0·05 was considered significant.
RESULTS
Salmonella serovars and phage types
Salmonella was isolated from 94 (47%) of the 200 samples collected from the various sampling sites. Of the 20 poultry farms investigated in Accra, five (25%, 95% CI 8·0–42·0) were positive for Salmonella compared to 28/55 (50·9%, 95% CI 37·7–64·1) flocks in Kumasi representing an overall flock prevalence of 44% (95% CI 31·8–56·2). This difference in flock prevalence between the two regions was not significant (χ 2 = 2·235, P = 0·53). When flocks were stratified by flock size, prevalence was found not to be statistically different between the smallest (⩽1000), second smallest (1001–2000), medium (2001–4000) and largest (⩾4001) flocks (Table 1). Sixteen serovars were identified; 13 serovars were identified in faecal samples, nine serovars in dust samples, and one serovar each from slaughterhouse, water and feed samples from the different sampling sites (Table 2). Most (21/33) of the positive poultry farms yielded Salmonella in faecal sock samples. Interestingly, only three farms (3/55) in Kumasi and one farm (1/20) in Accra shared common serovars in both sock and dust samples. The most common serovars identified were S. Nima (nine farms), S. Kentucky (seven farms), S. Muenster (seven farms), S. Enteritidis (six farms), and S. Cairina (six farms). S. Kentucky, was found only in Kumasi and more often (16/17) in sock samples than any other sample type (Table 2) (P < 0·0001 for comparison between fecal and dust samples). Positive poultry farms (n = 5) in Accra showed six serovars: S. Enteritidis (2), S. Virchow (3), S. Haifa (2), S. Bochum (1), S. Poona (1) and S. Ituri (1).
Table 1. Prevalence of Salmonella in poultry farms in Ghana and antimicrobial use practices by farmers

* χ 2 = 2·235, P = 0·525.
† χ 2 = 14·365, P = 0·002.
‡ χ 2 = 0·014, P = 0·906.
Table 2. No. of Salmonella-positive samples in poultry farms from Ghana

Phage-typing differentiated the 10 S. Enteritidis isolates into two lysotypes; nine strains belonged to PT1 and one belonged to PT21.
Antimicrobial resistance
The prevalence of resistance was calculated at the strain level. Multiple strains were included from 15 farms because they were clearly different (defined as showing a different serovar, phage type or resistance pattern). Of the 94 Salmonella strains tested, 60·6% (95% CI 50·7–70·5) corresponding to 57/94 strains were resistant to ⩾1 antimicrobials. Of these 23 (40·35%) showed multi-resistance (resistant to ⩾3 classes of antimicrobials), corresponding to a prevalence of multi-resistance in strains of Salmonella from Ghana of 24·5% (95% CI 15·9–33·1). In all, seven different multi-resistance patterns were obtained. The most significant resistances obtained were to NAL (89·5% of resistant strains), TET (80·7%), CIP (64·9%), SUL (42·1%), TMP (29·8%) and AMP (26·3%). Table 3 shows the serovars and their resistance to the antimicrobials tested and Table 4 lists the multi-resistance patterns observed.
Table 3. Antimicrobial resistance patterns of Salmonella serovars isolated from poultry in Ghana

AMP, Ampicillin; AMX + CL, amoxicillin + clavulanic acid; CHL, chloramphenicol; CIP, ciprofloxacin; GEN, gentamicin; NAL, nalidixic acid; SUL, sulfamethazole; TET, tetracycline; TMP, trimethoprim.
* All serovars were susceptible to cefotaxime, ceftazidime and cefoxitin.
† 0, Susceptible to all tested antimicrobials; 1, resistant to 1 antimicrobial; 2–3, resistant to 2–3 antimicrobials; >3, multiresistant to >3 antimicrobials.
‡ Numbers under the different antimicrobials indicate the number of resistant isolates.
Table 4. Multi antimicrobial resistance patterns of (⩾3 classes of antimicrobials) Salmonella serovars

AMP, Ampicillin; AMP + CL, ampicillin + clavulanic acid; CIP/NAL, ciprofloxacin/nalidixic acid; CHL, chloramphenicol; GEN, gentamicin; NAL, nalidixic acid; SUL, sulfamethazole; TET, tetracycline; TMP, trimethoprim.
All strains of the 16 serovars were susceptible to CTX, FOX and CAZ. Thirty-seven isolates from 12 serovars did not show resistance to any of the antimicrobials tested: S. Muenster (7), S. Virchow (5), S. Haifa (4), S. Enteritidis (2), S. Cairina (5), S. Nima (3), S. Duisburg (3), S. Poona (2), S. Rubislaw (2), S. Bochum (2), S. Typhimurium (1), and S. Stanleyville (1). Multi-resistant strains belonged to eight serovars: S. Kentucky (6), S. Nima (5), S. Virchow (4), S. Enteritidis (3), S. Cairina (2) and S. Muenster (2) (Table 4). All S. Kentucky strains shared a common resistance to CIP/NAL and TET in addition to other antimicrobials (Table 4).
As a quality control of antimicrobial resistance testing, 14 strains were randomly selected and subjected to antimicrobial testing by microdilution broth. This testing confirmed the classification of strains into resistant and sensitive groups, as obtained previously by disc diffusion method (data not shown).
Antimicrobial use practices by poultry farmers
The questionnaire interviews revealed that all 75 poultry keepers provided antimicrobials to their poultry. These antimicrobials were predominantly (98·7%) obtained because of initial advice of veterinarians. Drugs were administered by farmers through drinking water and not in feed. With regard to choice of drug, 45·3% of poultry keepers acknowledged choosing the antimicrobials on the type of illness combined with the advice of veterinarians, 41·3% used drugs suggested to them by sales persons and 13·3% of the farmers decided which antimicrobials to use based on experience and recommendation by sales persons. The most common drugs used (per number of farms) included oxytetracycline (60%), penicillin (40%), tylosin (40%), ciprofloxacin (20%), erythromycin (18·7%), enrofloxacin (13·3%), streptomycin (9·3%), doxycycline (9·3%), trimethoprim sulfadozine (8·0%) and neomycin (6·7%). Most of the farmers (42/75, 56%) used these antimicrobials routinely for the dual purpose of prevention and treatment of diseases. Twenty-one farmers (28%) used antimicrobials for only treatment of infections while eight (10·7%) used them solely for prevention of diseases. A small percentage (5·3%) used the antimicrobials for prevention, treatment and growth promotion. None of the 75 poultry keepers complied with the withdrawal period indicated on the product label as they sold eggs while the birds were on drug treatment, but 70/75 did not slaughter the birds for meat while on treatment (Table 1).
PFGE genotyping
PFGE typing of 41 isolates belonging to the most frequent Salmonella serovars (Kentucky, Nima, Enteritidis, Muenster) demonstrated that the strains belonging to the same serovar were typically closely related with common band patterns (Fig. 1). The majority (10/14) of S. Kentucky strains showed the same PFGE type and one additional strain differed by only one band from this pattern. However, two Kentucky strains had different band patterns. All strains of S. Kentucky were from Kumasi and all but one strain were isolated from faeces. The dominant clone of this serovar, identified in the current study, corresponds to the European PFGE database profile XbaI.0916, originally submitted from a human patient by an English laboratory. This type was demonstrated in both dust and faecal samples and was isolated from five different farms from a total of 11 flocks. S. Nima was also only isolated from farms in the Kumasi area. Analysis of S. Nima strains revealed greater diversity in band patterns with seven clearly different subtypes. Four strains had the same pattern. They originated in three farms from four different flocks. Moreover, strains of S. Enteritidis differed as they showed five subtype patterns; however, all but two strains showed similarities >91·9%. Evidence of infection with two different strains was obtained in one farm from Accra. Besides that, patterns were unique to one farm. S. Muenster (four subtypes) on the other hand clustered in two separate groups, indicating that two major clones of this serovar were present in the positive flocks. However, each pattern demonstrated was unique to one farm in the study.

Fig. 1. Dendogram showing clusters of XbaI PFGE of (a) Salmonella Kentucky (n = 14). Strain BUG1 and BUG2 are from the same farm but different flocks, ED3, ED4 and ED5 similarly from one farm but different flocks and MB2 and MB1 from the same farm but different flocks. The remaining strains were from different flocks on different farms. (b) S. Nima (n = 11). Strain D11 and D13 are from the same farm but isolated from different flocks. The remaining strains originate from different flocks at different farms. (c) S. Enteritidis (n = 8). PB05-1, PB05-2 and PD05-2 are from the same farm in the Accra region, but from different flocks. The remaining strains were obtained from different flocks and farms. (d) S. Muenster (n = 8) isolates from poultry farms in Ghana. All strains were from different flocks at different farms. Dice coefficient was used for the similarity analysis and clustering was done using unweighted pair-group method with arithmetic means with 1% band position tolerance and 0·50% optimization parameter. KB, PB and B, Salmonella from faecal sources; PD and D, Salmonella from dust; AYFe, Salmonella from feed.
DISCUSSION
This cross-sectional study investigated the prevalence of Salmonella serovars in egg-laying hens and broilers and their resistance to antimicrobials generally used in poultry production and human medicine in Ghana. We found an overall Salmonella farm flock prevalence of 44·0% with regional prevalence of 25% for Accra and 50·9% for Kumasi. This difference in regional prevalence was not significantly different, and it should be noted that the overall estimate was based on an under-representation of the farms located in the Accra region compared to the farms included in Kumasi. There are about 300 small- to large-scale poultry farms involved in both layer and broiler production with the majority located in the peri-urban centres of Accra (Greater Accra Poultry Farmers association), compared to about 480 in Kumasi (Ashanti Region Poultry Farmers Association). The prevalence we report is above what has recently been reported as average from laying hens in Europe [25] demonstrating an overall Salmonella prevalence of 30·8% and between 0% and 79·5% for individual member states. The prevalence found in Ghana is also considerably higher than the 18% prevalence reported in Bangladesh [Reference Barua10], a country with a comparable structure of the poultry industry. There is a need to conduct further investigations into what are the main sources and factors associated with the occurrence of Salmonella, e.g. infection status of 1-day-old chickens, and efficiency of cleaning and other hygiene and biosecurity measures in general in the poultry industry in Ghana. We found a relatively high proportion of dust samples from farms to be Salmonella positive, suggesting that a farm environment once contaminated with Salmonella may contribute to the persistence of Salmonella infections within the farm. Generally cleaning procedures were found to be suboptimal. The production was run as all in/all out, but with insufficient cleaning between flocks. Litter was removed from the ground and washing of floors was performed, and fresh litter was added. No disinfection was used and no empty period was implemented between flocks. This may contribute to the relatively high flock prevalence found in this study. Serovar distribution did not differ much between sock samples, but some serovars were more common to sock than dust samples, and vice versa. However, in contrast to reports from Europe, where dust samples were twice more likely to be positive for Salmonella than faecal samples [25], sock samples with faeces were the most common positive type of sample in our investigation, possibly due to climatic conditions, with a higher temperature in the dust.
S. Kentucky was one of the most common serovars identified, but only in the Kumasi study area, whiles S. Haifa and S. Virchow were common in the poultry farms located around Accra. The serovar diversity found in poultry farms in the two regions of Ghana is lower than reported for Senegal [Reference Bada-Alambedji26]. The majority of S. Kentucky strains were demonstrated to be of clonal origin as shown in the PFGE analysis, suggesting a common source of infection. Up to three flocks belonging to the same farm were positive by this serovar, but the sampling strategy does not allow us to determine whether they share a common external source or whether the rather poor biosecurity within farms contributes to this observation. Studies by Le Hello et al. [Reference Le27, Reference Le28] documents that a strain of S. Kentucky seems to have spread throughout several developing countries and infected both human and non-human sources. Although the CIPR S. Kentucky strain belonging to ST198-X1 reported by Le Hello et al. was similar in resistance pattern (amoxicillin, streptomycin, spectinomycin, gentamicin, sulfamethoxazole, tetracycline, nalidixic acid, ciprofloxacin) to our Kentucky strains (ampicillin, gentamicin, sulfamethoxazole, tetracycline, nalidixic acid, ciprofloxacin), the PFGE type we demonstrated was of a different type. This serovar is regularly isolated from humans with symptoms consistent with salmonellosis. Although the source(s) is currently is unknown, research has demonstrated poultry eggs and meat as major vehicles for infection, while the spread across continents can be attributed to possible human travel and egg imports [Reference Le27, Reference Le28]. Strains of S. Nima and S. Enteritidis also showed clonal relationships as they clustered in groups with high similarity. Contrary to this, strains of S. Muenster were found to be more diverse. This finding is similar to work by Ayala [Reference Ayala29] where S. Muenster was the most diverse with 11 different subtypes. For the three last serovars, patterns demonstrated were found to be unique for each positive farm, suggesting that several sources exist for spread of these serovars in poultry in Ghana.
The current study is the first report on specific Salmonella serovars in egg-laying hens and broilers in Ghana. Only limited information is available about serovars associated with human gastroenteritis [Reference Saba30], and there is a pressing need to perform similar characterization of strains from humans in order to be able to assess the importance of the serovars detected in the poultry industry in the current study. S. Kentucky has been reported in other West African countries such as Nigeria [Reference Ogunleye and Carlson31] and Senegal [Reference Bada-Alambedji26] and it would be interesting to determine if the genotypes found in these countries are identical to the strains common in the poultry industry in Ghana. Phage-typing of S. Enteritidis strains revealed the dominance of PT1 which has been known to have originated from the Baltic region, but has over time been identified in other regions and which has caused several human diarrhoeal outbreaks around the world [Reference Fisher32–Reference Lukinmaa34].
After repeated disc diffusion tests in two different laboratories, results showed that all Salmonella strains were susceptible to cephalosporins (third generation) and most to amoxicillin in combination with clavulanic acid. The observation of lack of cephalosporin resistance was surprising, but was confirmed by analysis in two different laboratories and with two different methods. Breakpoints in the current study were defined according to Neo-Sentitab recommendations; however, had inhibition zone diameters been interpreted according to the EUCAST epidemiological cut-off, still no strains would have been characterized as resistant to third-generation cephalosporins. ESBL Salmonella has been reported in several clinical diagnoses and from researchers around Africa. However, information on its occurrence in poultry is very scarce. In Ghana, ESBL has been reported in Enterobacteriaceae from human clinical specimens, but this has not been linked to poultry. Most strains (60·6%) showed resistance to one or more antimicrobials tested and almost 1/4 strains were resistant to more than three of the antimicrobial classes tested. Although no cephalosporin resistance was reported in our study, resistance to a third-generation cephalosporin, cefuroxime, has been reported earlier in Ghana for E. coli isolated from poultry and humans [Reference Donkor, Newman and Yeboah-Manu19].
Drug resistance has been attributed to use, overuse and misuse of antimicrobials which favour emergence of resistant strains [Reference Newman35]. The high level of antimicrobial resistance in the current study can be attributed to the high rate of antimicrobial usage by poultry keepers, as 56% used antimicrobials routinely for the dual purpose of prevention and treatment of diseases. The drugs commonly used in the Ghanaian poultry industry correlated with the observed drug resistance with a high level of resistance seen against tetracycline and ciprofloxacin. Recently, a study in Ghana showed a high level of exposure of production animals to feed commonly containing antimicrobials [Reference Donkor, Newman and Yeboah-Manu19]. The study also revealed that antimicrobials were applied in therapeutic doses by livestock keepers primarily to prevent diseases.
One-day-old chickens and hatching eggs for both layer and broiler production in Ghana are imported from other countries, mainly the European Union, United States and Brazil (http://ghanatrade.org/agricultural-import-ghana/). It remains to be determined to what extent the antimicrobial resistance observed in this study is due to the high usage of antimicrobials in animals in Ghana, as reported previously for the human and animal sector [Reference Wilkens17, Reference Donkor, Newman and Yeboah-Manu19, Reference Newman35], or introduction of resistant Salmonella from other countries, which could, for example, be the case for S. Kentucky. Our study revealed that antibiotics were commonly used in the poultry industry, often in absence of disease and based on advice given by persons with no particular insight into disease problems in poultry. This is parallel to the situation in other developing countries such as Tanzania [Reference Katakweba36], and calls for action to implement more strict bylaws for use of antibiotics in livestock. Salmonella were once susceptible to a broad range of affordable and effective antimicrobial drugs [Reference Feasey37] but multidrug-resistant strains have emerged throughout Africa [Reference Wouafo12, Reference Bada-Alambedji26, Reference Saba30, Reference Ammari38–Reference Zewdu and Cornelius42].
Salmonella serovars isolated during our study showed in common resistance to nalidixic acid and tetracycline (both very high) and to ciprofloxacin, ampicillin, sulfamethazole and trimethoprim and some further showed reduced susceptibility to amoxicillin/clavulanic acid. The high resistance to nalidixic acid is indicative of a high risk of development of ciprofloxacin resistance. These antimicrobials are also commonly used for treatment of other animal and human bacterial infections in Ghana [Reference Wilkens17, Reference Donkor, Newman and Yeboah-Manu19, Reference Newman35]. The antimicrobial percentage and resistance patterns of Salmonella strains (overall 60·6% and 19·8% multi-resistance) in this study are lower compared with Salmonella isolated in Senegal [Reference Bada-Alambedji26] which showed an overall 78·9% resistance and 45·6% multi-resistance and that in Ethiopia where 62·1% of strains were multi-resistant [Reference Zewdu and Cornelius42]. All 17 S. Kentucky strains showed resistance to more than one and up to nine of the antimicrobials tested with high multiple drug resistance. These corroborate reports that the S. Kentucky found worldwide is multidrug-resistant [Reference Le27, Reference Ogunleye and Carlson31, Reference Zewdu and Cornelius42]. However, it should be noted that all Salmonella isolated in this study were susceptible to the third-generation cephalosporin, cefotaxime, which was also the case for E. coli strains isolated in a recent study of poultry in the Accra region [Reference Donkor, Newman and Yeboah-Manu19].
In conclusion, the Ghanaian broiler and egg-laying industry is showing some moderate prevalence of multi-resistant Salmonella and the common finding of clonal S. Kentucky in the Kumasi area warrants further investigations into the epidemiology of this serovar. There is an urgent need to establish a surveillance and control programme on Salmonella and use of antimicrobials not just in the Ghanaian poultry industry, but also in other production animals, foods and humans to ensure food safety and human health.
ACKNOWLEDGEMENTS
We are grateful to Danida (Danish International Development Assistance) for supporting this research through the Antimicrobial Drug Monitoring and Evaluation of Resistance (ADMER) project. We thank the Poultry Farmers Association of Ghana, Gitte Petersen (University of Copenhagen, Denmark); Eric Acheampong and Kweku Peprah (TAB, Kwame Nkrumah University of Science and Technology) and Michael Oltaiwo (University of Ghana Medical School) for field and technical support.
DECLARATION OF INTEREST
None.





