



Introduction
The Gebel Maghara displays a well-exposed marine Middle to Upper Jurassic succession, which contains, apart from some ammonites, a diverse and well-preserved benthic macrofauna, dominated by bivalves, gastropods, and brachiopods. The most comprehensive study of the Jurassic rocks of Gebel Maghara is that of Al Far (Reference Al Far1966), who focused on sedimentologic and stratigraphic aspects. With respect to paleontology, some fossil groups, particularly bivalves and gastropods, have been neglected from a taxonomic point of view since the pioneer study of Douvillé in the early twentieth century (Douvillé, Reference Douvillé1916). Although limited research has been carried out since then (e.g., brachiopods: Farag and Gatinaud, Reference Farag and Gatinaud1960; Hegab, Reference Hegab1989, Reference Hegab1991a, Reference Hegabb; Feldman et al., Reference Feldman, Owen and Hirsch1991, Reference Feldman, Radulovic, Hegab and Radulovic2012; Ali et al., Reference Ali, Azab and Hussain1997; bivalves and gastropods: Hirsch, Reference Hirsch1980; Abdelhamid, Reference Abdelhamid2002; Khalil, Reference Khalil2003; ammonites: Douvillé, Reference Douvillé1916; Arkell, Reference Arkell1952; Parnes, Reference Parnes1988; echinoids: Fourtau, Reference Fourtau1924), a comprehensive modern taxonomic study of some groups such as the bivalves and gastropods is still lacking. Abdelhady (Reference Abdelhady2014) and Abdelhady and Fürsich (Reference Abdelhady and Fürsich2014, Reference Abdelhady and Fürsich2015a, Reference Abdelhady and Fürsichb, Reference Abdelhady and Fürsichc) concentrated mainly on litho- and biostratigraphy, paleobiogeography, and paleoecology. The latter authors listed 232 benthic and nektonic macrofaunal taxa from Gebel Maghara, which were dominated by bivalves (60 species), followed by gastropods (52), ammonites (34), brachiopods (29), corals (28), echinoids and crinoids (16), sponges (7), serpulids (5), and a single crustacean species (Abdelhady, Reference Abdelhady2014, appendix B), but did not provide a taxonomic analysis. Therefore, their collections were given to the present authors for further processing and revising. Protobranch bivalves are common elements of fossil assemblages throughout the Phanerozoic, restricted to fully marine environments. Due to their detritivorous feeding habits, they only live in organic-rich bottoms as very active shallow burrowers (e.g., McAlester and Rhoads, Reference McAlester and Rhoads1967; Nicol, Reference Nicol1969, Reference Nicol1972; Damborenea, Reference Damborenea1987; Damborenea and Pagani, Reference Damborenea and Pagani2019). The present protobranch bivalve specimens are well preserved, mostly articulated, and their internal characters, such as hinge teeth and muscle scars, can be recognized easily. The present study is the second taxonomic analysis of protobranch bivalves from the area, following the Ayoub-Hannaa et al. (Reference Ayoub-Hannaa, Abdelhady and Fürsich2017) study of Costinuculana magharensis Ayoub-Hannaa, Abdelhady, and Fürsich, Reference Ayoub-Hannaa, Abdelhady and Fürsich2017.
Locality and geological setting
Gebel Maghara is a dome-like structure covering an area of ~400 km2, situated in northern Sinai, 50 km south of the Mediterranean coast (Fig. 1.1). During the Triassic, the Tethys north of Gondwana started to rift and led to the formation of small intracratonic basins (horst-graben systems) with differential subsidence in northern Egypt (Keeley, Reference Keeley1994; Ayyad et al., Reference Ayyad, Darwish and Sehim1998; Garfunkel, Reference Garfunkel1998; Moustafa et al., Reference Moustafa, El-Badrawy and Gibali1998). The opening of these basins started in Late Triassic–Early Jurassic time (Biju-Duval et al., Reference Biju-Duval, Letouzey, Montadert, Watkins, Montadert and Dickerson1979; Garfunkel and Derin, Reference Garfunkel and Derin1984; Mart, Reference Mart1987). Gebel Maghara lies in one of these basins, consisting of an extensional half graben-like structure (Fig. 1.2), and comprises the thickest and most complete Jurassic outcrop in northern Sinai (1800 m; Al Far, Reference Al Far1966; Keeley, Reference Keeley1994). The stratigraphic scheme of Gebel Maghara is based mainly on Al Far (Reference Al Far1966), who provided a complete classification and description of the Jurassic strata. Later, Hirsch (Reference Hirsch1980) and Picard and Hirsch (Reference Picard and Hirsch1987) modified Al Far's classification and compared the succession at Gebel Maghara with that of the adjacent Negev desert. According to Al Far (Reference Al Far1966), the marine sediments are represented by the Rajabiah (Lower Jurassic), Bir Maghara, and Masajid formations, whereas the continental sediments include the Mashabba (Lower Jurassic), Shusha, and Safa formations. Abdelhady (Reference Abdelhady2014) and Abdelhady and Fürsich (Reference Abdelhady and Fürsich2015a, Reference Abdelhady and Fürsichb, Reference Abdelhady and Fürsichc) studied four sections in detail, from west to east Gebel Homayir (1190 m thick), Gebel Arousiah (1177 m thick), Gebel Engabshi (1025 m thick), and Gebel Mowerib (995 m thick) (Fig. 1.2). Three of them (Gebel Homayir, Gebel Arousiah, and Gebel Engabashi) belong to the western flank of the anticline with a dip of the strata varying from 15–30°, while the Gebel Mowerib section is exposed on the eastern flank dipping at an angle >60° in some parts.

Figure 1. (1) Locality map of Gebel Maghara; (2) cross-section and geologic map (modified after Al Far, Reference Al Far1966; Hirsch, Reference Hirsch1980) with position of the investigated sections of the Jurassic succession of Gebel Maghara; (3) the Jurassic subdivision and equivalent formations from older to younger (after Abdelhady and Fürsich, Reference Abdelhady and Fürsich2015a).
The Jurassic succession of these sections has been subdivided into seven formations from older to younger, the Mahl, Bir Maghara, Safa, Kehailia, Arousiah, Tauriat, and Masajid formations (Fig. 1.3). All of these formations were originally introduced by Al Far (Reference Al Far1966), Picard and Hirsch (Reference Picard and Hirsch1987), and recently revised by Abdelhady (Reference Abdelhady2014). Figure 2 shows the lithologs, lateral facies changes, faunal content, and correlation of the sections studied. In addition, the age estimation of the studied section was resolved based on ammonites (Abdelhady and Fürsich, Reference Abdelhady and Fürsich2015a).

Figure 2. Logged sections and their correlations based on ammonites and lateral facies changes (after Abdelhady and Fürsich, Reference Abdelhady and Fürsich2014, Reference Abdelhady and Fürsich2015a, Reference Abdelhady and Fürsichb). Vertical scales marked in 100-m increments.
Materials and methods
Several thousand specimens of the benthic and nektonic macrofauna (~9130) were collected by F.T. Fürsich in 1993 and by A.A. Abdelhady and F.T. Fürsich in 2012 from the Jurassic succession of Gebel Maghara. During that time, four sections, Gebel Homayir, Gebel Arousiah, Gebel Engabshi, and Gebel Mowerib, corresponding to an ~20 km long W-E transect, were studied and measured using a Jacob Staff, hand lens, and 10% concentrated HCl. These studies included detailed documentation of the lithology, bed contacts, sedimentary structures, trace fossils, taphonomic observations, and bed by bed collecting of macrofossils. The collected macrofauna was identified during 2012–2014 as a part of a Ph.D. project by A.A. Abdelhady (Reference Abdelhady2014). All specimens have been deposited in the collections of the Bayerische Staatssammlung für Paläontologie und Geologie in Munich under the prefix BSPG 2014V.
The protobranch bivalve specimens are generally very well preserved, mostly articulated, and preserved with shell. A few specimens are composite molds and laterally deformed due to compaction. Most specimens have been prepared mechanically and cleaned by steel needles under a binocular microscope. Air abrasive was used to clean hinge and teeth. In order to obtain complete hinge structures, the rock matrix that covered the hinge plate was removed using diluted hydrogen peroxide (H2O2). The specimens were photographed after having been coated with ammonium chloride to enhance details of ornamental features and other internal features such as teeth and muscle scars. Identification was greatly supported by the Jurassic bivalve catalogue at the GeoZentrum Nordbayern of the Friedrich-Alexander-Universität, Erlangen. This catalogue contains photocopies of ~95% of all figured Jurassic bivalves, which have been identified from different localities worldwide. In addition, each entry includes information on locality, stratigraphic level, authors, and publication date.
The systematic classification of the bivalves follows that of Carter et al. (Reference Carter, Altaba, Anderson, Araujo, Biakov, Bogan and Campbell2011). The synonymy lists contain only references, which have been carefully checked by the authors, beginning with the first reference of the particular taxon, and followed by literature records from the Jurassic of Egypt. More comprehensive synonymies can be found in the references cited. Abbreviations in synonymy lists and open nomenclature follow the indications given by Matthews (Reference Matthews1973) and Bengtson (Reference Bengtson1988) to indicate the degree of confidence in allocation of each entry. In particular, the period in front of the year means the authors accept the responsibility for attaching this reference to the species under discussion.
Morphological terminology follows the glossary of Cox (Reference Cox, Moore and Teichert1969) in the Treatise on Invertebrate Paleontology. The terminology of hinge teeth follows Babin (Reference Babin1966) and Carter et al. (Reference Carter, Harries, Malchus, Sartori, Anderson, Bieler and Bogan2012; Fig. 3.4). Measured dimensions and terminology of teeth are given in Figure 3.1–3.3. Linear measurements (taken with a Vernier caliper, accurate to 0.1 mm) are in millimeters. Abbreviations for dimensions are: L = length, La = anterior length, Lp = posterior length, H = height, Iav = inflation of articulated valves, and Isv = inflation of a single valve. In the description of taxa, very small refers to a length of 1–5 mm and small to a length of 6–15 mm. A principal component analysis (PCA) using the PAST software, version 2.16 (Hammer et al., Reference Hammer, Harper and Ryan2001), on a variance-covariance matrix of the log-transformed variables was carried out to clarify the morphological relationships of some of the taxa.
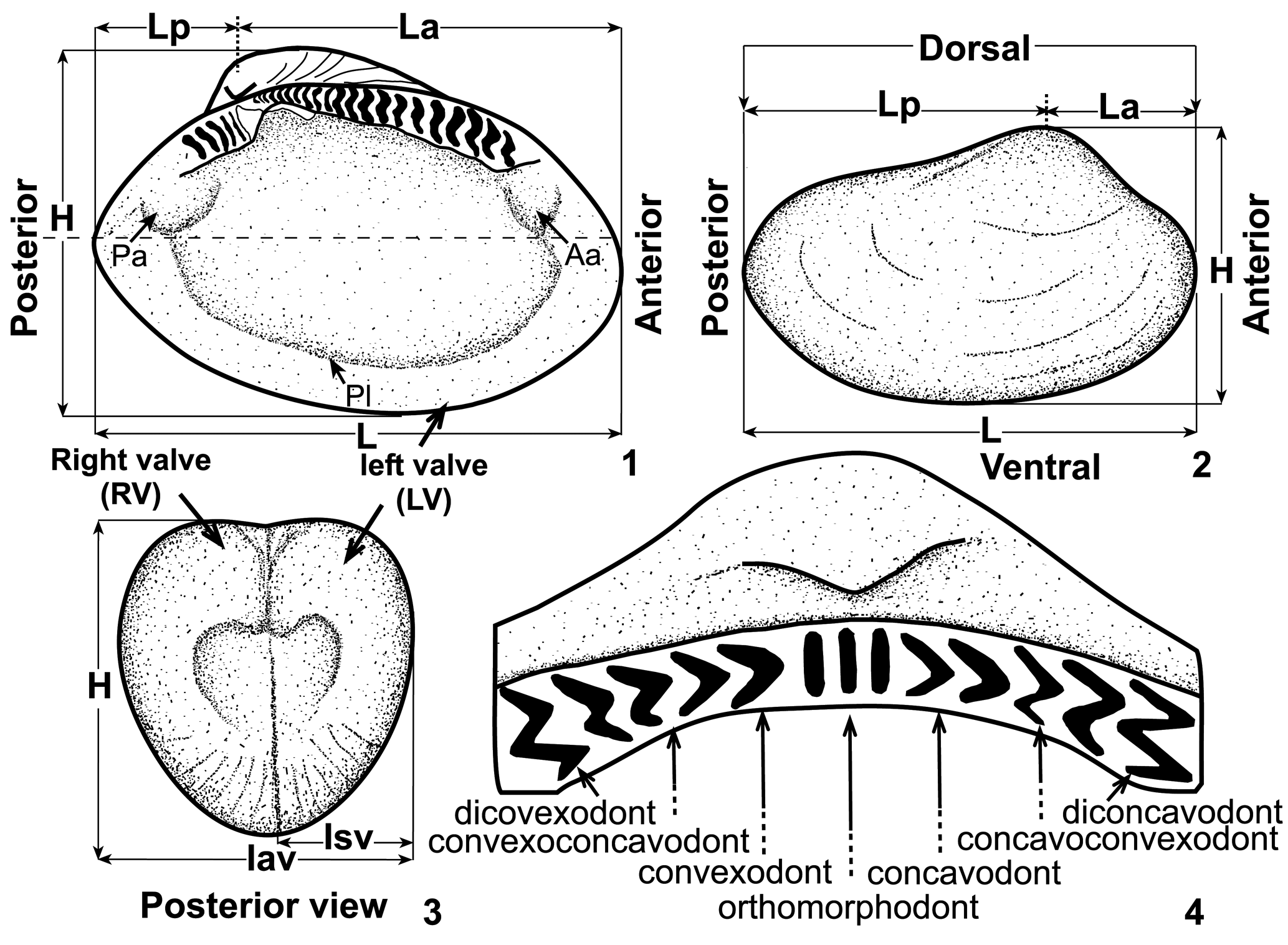
Figure 3. Measured dimensions and orientation of selective protobranch bivalves. (1, 3) Nuculoma, (2) Palaeoneilo, (4) terminology of dentition, based on Babin (Reference Babin1966, fig. 3) and Carter et al. (Reference Carter, Harries, Malchus, Sartori, Anderson, Bieler and Bogan2012, fig. 207). See text for abbreviations.
Repositories and institutional abbreviations
All specimens examined in this study are deposited in the collections of the Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany, under the prefix BSPG 2014V and PIW 1991 III.
Systematic paleontology
Class Bivalvia Linnaeus, Reference Linnaeus1758
Clade Eubivalvia Carter et al., Reference Carter, Altaba, Anderson, Araujo, Biakov, Bogan and Campbell2011
Subclass Protobranchia Pelseneer, Reference Pelseneer1889 (= Palaeotaxodonta Korobkov, Reference Korobkov1954)
Superorder Nuculiformii Dall, Reference Dall1889 (= Foliobranchia Ménégaux, Reference Ménégaux1889)
Order Nuculida Dall, Reference Dall1889
Superfamily Nuculoidea Gray, Reference Gray and Parry1824
Family Nuculidae Gray, Reference Gray and Parry1824
Genus Nuculoma Cossmann, Reference Cossmann1907 (= Habonucula Singh and Kanjilal, Reference Singh and Kanjilal1977)
Type species
Nucula castor d'Orbigny, 1850, from the Callovian of France, monotypy; figured by Cottreau (Reference Cottreau1925, p. 153, pl. 39, figs. 23, 24).
Remarks
Singh and Kanjilal (Reference Singh and Kanjilal1977, p. 189) erected the new genus Habonucula from the lower Callovian rocks of the Kachchh Basin, India, based on the absence of escutcheon and smooth inner shell margins. Jaitly et al. (Reference Jaitly, Fürsich and Heinze1995, p. 155) pointed out that these diagnostic features are not sufficient enough to erect a new genus, and therefore they regarded Habonucula as a junior synonym of Nuculoma. The latter view is followed here. Nuculoma can be distinguished from other Jurassic nuculid genera by its strongly opisthogyrate enrolled umbones, which usually overhang the posterior margin, and its surface, which carries numerous fine commarginal riblets (Jaitly et al., Reference Jaitly, Fürsich and Heinze1995). Nuculoma has a wide stratigraphic range, from the Lower Jurassic to Lower Cretaceous (e.g., Berriasian–lower Hauterivian) of northern Central Siberia (Sanin, Reference Sanin1976) and to Valanginian strata of eastern England (Kelly, Reference Kelly1984).
Nuculoma douvillei new species
Figures 4, 5.1–5.19, 6, 7
- *Reference Douvillé1916
Nucula variabilis? Douvillé, p. 61, pl. 5, figs. 51–55 (non Sowerby, Reference Sowerby1825).
- v.Reference Abdelhady2014
Nuculoma variabilis; Abdelhady, p. 72, fig. 5.4G, H.
- v.Reference Abdelhady and Fürsich2014
Nuculoma variabilis; Abdelhady and Fürsich, p. 181, fig. 6G, H.
- v.Reference Abdelhady and Fürsich2015a
Nuculoma variabilis; Abdelhady and Fürsich, p. 41.

Figure 4. (1–40) Variation in outline of Nuculoma douvillei n. sp. from the Bathonian–Callovian rocks of the Maghara area. (1–5) BSPG 2014V 309/1, holotype, Kehailia Formation, Gebel Homayir, (1) left valve exterior, (2) posterior view of articulated valves showing the absence of escutcheon, (3) right lateral view, (4) dorsal view, (5) ventral view. (6–9) Paratypes, Kehailia Formation, Gebel Homayir; (6) BSPG 2014V 173/1, left lateral view, (7) BSPG 2014V 173/1, right valve exterior, (8) BSPG 2014V 173/2, dorsal view showing a well-developed anterior umbonal ridge, (9) BSPG 2014V 173/3, right lateral view. (10) BSPG 2014V 167/1 paratype, Kehailia Formation, Gebel Engabashi, right lateral view showing widely spaced commarginal ribs intercalated between faint commarginal growth lines. (11–16) Paratypes, Kehailia Formation, Gebel Homayir; (11) BSPG 2014V 309/2, left valve, (12) BSPG2014V 309/3, right lateral view, (13) BSPG2014V 309/4, left lateral view, (14) BSPG2014V 309/5, right lateral view, (15, 16) BSPG 2014V 309/6 and BSPG 2014V 309/67, respectively, left lateral views. (17–23) Paratypes, Arousiah Formation, Gebel Mowerib; (17) BSPG 2014V 178/1, left lateral view, (18) BSPG 2014V 178/2, right lateral view, (19, 20) BSPG 2014V 178/3, left lateral view and left valve interior, respectively, (21, 22) BSPG 2014V 178/4, exterior and interior of left valve, respectively, (23) BSPG 2014V 178/5, exterior of right valve. (24) Paratype, BSPG 2014V 4/1, Kehailia Formation, Gebel Arousiah, right valve exterior. (25–36) Paratypes, Kehailia Formation, Gebel Homayir; (25, 26) BSPG 2014V 308/1, left lateral view, showing the subrounded outline of the species, and right lateral view, respectively, (27–29) BSPG 2014V 308/2, left lateral view, showing strongly oblique form of species, anterodorsal view, and right lateral view, respectively, (30) BSPG 2014V 308/3, posterior view, (31) BSPG 2014V 308/4, dorsal view, showing a well-developed anterior umbonal ridge, (32) BSPG 2014V 308/5, left valve, (33) BSPG 2014V 308/6, right valve, (34–36) BSPG 2014V 308/7, left lateral view, posterior view, showing the strongly inflated valves, and right lateral view, showing narrow rounded anterior margin; (37) BSPG 2014V 309/8, paratype, Kehailia Formation, Gebel Homayir, right lateral view showing a truncated anterior margin. (38–40) Paratypes, Safa Formation, western Bir Maghara; (38) BSPG 2014V 310/1, right valve interior, (39) BSPG 2014V 310/2, right lateral view, showing a slightly convex anterior margin, (40) BSPG 2014V 310/3, right lateral view. Specimens in (37–40) show increasingly convex anterior margins. Scale bars = 3 mm.

Figure 5. (1–19) Internal characters of Nuculoma douvillei n. sp. from the Bathonian–Callovian rocks of the Maghara area. (1, 2) Paratypes, Arousiah Formation, Gebel Mowerib; (1) BSPG 2014V 178/6, left lateral view, (2) BSPG 2014V 178/7, right lateral view showing subrectangular anterior muscle scar and small rounded posterior muscle scar; (3) BSPG 2014V 310/4, paratype, Safa Formation, western Bir Maghara, incomplete right valve showing the elongated ovate anterior muscle scar; (4) BSPG 2014V 178/8, paratype, Arousiah Formation, Gebel Mowerib, dorsal view of left valve showing abnormally large posterior teeth (arrowed). (5–10) Paratypes, Safa Formation, western Bir Maghara; (5) BSPG 2014V 310/5, posterodorsal view, showing large posterior teeth, (6, 7) BSPG 2014V 310/6, left lateral view and close-up showing shoehorn-shaped chondrophores (arrowed), respectively, (8, 9) BSPG 2014V 310/7, incomplete right valve and close-up showing a short subhorizontal chondrophore, respectively, (10) BSPG 2014V 310/8, right lateral view, with deep umbonal cavity; (11, 12) BSPG 2014V 309/9, paratype, Kehailia Formation, Gebel Homayir, left lateral view, interior and close-up showing concavodont teeth (arrowed) with short chondrophore; (13, 14) BSPG 2014V 178/9, paratype, Arousiah Formation, Gebel Mowerib, interior left lateral view and close-up showing the large strongly concavodont teeth close to beak, respectively; (15) BSPG 2014V 309/10, paratype, Kehailia Formation, Gebel Homayir, left lateral view. (16, 17) BSPG 2014V 178/10, paratype, Arousiah Formation, Gebel Mowerib, interior left lateral view and sketch of hinge showing slightly diconcavodont teeth, respectively; (18, 19) BSPG 2014V 310/9, paratype, Safa Formation, western Bir Maghara, right lateral view and close-up showing convexodont teeth of posterior hinge (arrowed), respectively. (20–38) Nuculoma sinaiensis n. sp. from the upper Lower Jurassic (Toarcian) to Middle–Upper Jurassic (Bajocian–Callovian and lower Kimmeridgian) of Gebel Maghara; (20–24) BSPG 2014V 303/1, holotype, Bir Maghara Formation, Gebel Arousiah; (20) left lateral view, (21) anterodorsal view showing the absence of a lunule, (22) right lateral view, (23) close-up showing fine, dense commarginal ribs, (24) posterior view of articulated valves showing the absence of escutcheon. (25–27) BSPG 2014V 313/1, paratype, Masajid Formation, western Bir Maghara; (25) left lateral view, (26) posterior view of articulated valves, (27) right lateral view; (28–31) paratypes, Masajid Formation, western Bir Maghara; (28, 29) left valve exterior and left valve interior showing terminal umbo (arrowed), (30, 31) BSPG 2014V 312/2, right valve interior and right valve exterior showing position of beak (arrowed); (32–38) paratypes, Masajid Formation, western Bir Maghara; (32–35) right valve exterior, left valve exterior, posterior, and close-up showing the posterior slope (= corselet), respectively, (36–38) BSPG 2014V 313/3, posterior, dorsal, and left lateral views, respectively. Scale bars = 3 mm.

Figure 6. (1) Relative abundance (%) of Nuculoma douvillei n. sp. in the studied sections. (2) Estimated ages of the studied formations based on ammonites (Abdelhady and Fürsich, Reference Abdelhady and Fürsich2015a); P. = Pachyceras; E. = Erymnoceras.

Figure 7. Principal component analysis (PCA) of Nuculoma douvillei n. sp. using the PAST software (Hammer et al., Reference Hammer, Harper and Ryan2001). (1) Scatter plot of PC1 vs. PC2 showing overlap of different morphotypes. (2) Boxplot of the four forms (A–D) based on shell outline (PC2). (3) Percentage of variation explained by PCA of morphological variables. (4) Scatter plot of PC1 vs. PC3 also showing overlap of the different morphotypes based on shell size. (5, 6) Relative abundance of morphotypes collected from the studied sections and boxplot of the forms (A–D) based on shell size (PC1). The large symbols in the scatter plots represent the centroid value for the respective groups (A–D). For data see Appendix A.
Holotype
Articulated specimen BSPG 2014V 309/1 in shell preservation (Fig. 4.1–4.5), from the middle–upper Bathonian Kehailia Formation, Gebel Homayir, Sinai, Egypt.
Paratypes
Three hundred and twelve specimens, in shell preservation, mostly articulated from Bathonian–Callovian rocks of the Maghara area (Fig. 6.1): 121 specimens from the middle–upper Bathonian Kehailia Formation (ammonite Clydomphalites clydocromphalus Zone), Gebel Homayir (BSPG 2014V 173/1–21; 174/1; 175/1–28; 308/1–53; 309/2–19); 19 specimens from the Kehailia Formation, Gebel Arousiah (BSPG 2014V 4/1–19); 17 from the same formation, Gebel Mowerib (BSPG 2014V 311/1–17); a single specimen from the Kehailia Formation, Gebel Engabashi (BSPG 2014V 167/1); 22 specimens from the upper third of the lower Bathonian Safa Formation, western Bir Maghara (BSPG 2014V 310/1–22); and 132 specimens from the Callovian Arousiah Formation (ammonite Pachyceras lalandeanum/Erymnoceras philbyi Zone), Gebel Mowerib (BSPG 2014V 178/1–132).
Diagnosis
Small to very small, strongly oblique Nuculoma, variable in outline, ranging from elongated-ovate to subtrapezoidal, from sub-trigonal to rounded; higher than long or longer than high; strongly inflated; with smooth inner margins, no escutcheon, no lunule, sharply pointed and strongly enrolled beaks, a variable anterior margin (mostly truncated to slightly convex, occasionally narrow and strongly convex), a well-developed anterior umbonal ridge, highly variable anterior and posterior teeth in shape and size; upward-curved, shoehorn-shaped subhorizontal chondrophore in both valves; regular, widely spaced commarginal ribs at mid-flank, occasionally intercalated with faint commarginal growth lines, and smooth anterior and posterior flanks.
Occurrence
Middle Bathonian to upper Callovian, Gebel Maghara, North Sinai (Fig. 6.2).
Description
Shell small to very small, oblique; shape and outline variable, ranging from elongated-ovate (higher than long or longer than high; Table 1) to subtrapezoidal, from sub-trigonal to subrounded, equivalved, strongly inequilateral, and strongly inflated (Iav/L: 0.81–1.19). Maximum inflation slightly below umbo. Posterior end blunt, meeting ventral margin in rounded curve. Anterior margin variable in outline, ranging from truncated to sub-truncated, from slightly to strongly convex (Fig. 4.37–4.40), and meeting ventral margin in blunt angle. Internal margins smooth, umbonal cavity deep. No escutcheon and lunule. Umbones strongly inflated, triangular, elevated above hinge, and extremely enrolled posteriorly. Beak sharply pointed and opisthogyrate. Anterior umbonal ridge well developed dorsally, fading towards anteroventral corner, and separating a narrow, smooth anterior flank from rest of valve (Fig. 4.8, 4.28, 4.31). Anterior adductor muscle scar larger than posterior one, elongated-ovate to subrectangular, and located close to anterior margin (Fig. 5.2, 5.3). Posterior adductor muscle scar small, subrounded and located close to posterior margin below posterior teeth. Pallial line complete, without sinus. Hinge plate broad with strong, unequal teeth and sockets. Anterior part of hinge with 12–20 unequal orthomorphodont to slightly diconcavodont teeth, gradually increasing in size from anterior end to the middle of anterior hinge, where the teeth are exceptionally large and thick, followed by an abrupt decrease in size till umbo (Fig. 5.3–5.5). Posterior hinge much shorter and broader than anterior one with 3–5 very large unequal teeth that are orthomorphodont and perpendicular to posterodorsal margin at posterior end, becoming strongly concavodont towards beak (occasionally convexodont in few specimens; Fig. 5.14, 5.19), gradually increasing in size towards umbo. Anterior and posterior valves with upward-curved, short, shoehorn-shaped subhorizontal chondrophore (Fig. 5.7, 5.9). Ornamentation consisting of regular, fine, widely spaced commarginal riblets on the middle flank of valve, decreasing in strength towards anterior and posterior margins (Fig. 4.3, 4.9, 4.34). Interspaces between riblets, occasionally carrying faint commarginal growth lines.
Table 1. Measurements (in mm) of Nuculoma douvillei n. sp.

Etymology
In honor of Prof. Dr. Joseph Henri Ferdinand Douvillé, French professor of paleontology at the École des Mines around the turn of the twentieth century, who documented the Jurassic fauna of Gebel Maghara for the first time.
Measurements
See Table 1.
Remarks
With respect to shell outline, Nuculoma douvillei n. sp. is a very variable species, ranging from subtrapezoidal to strongly oblique elongated-ovate with intermediate forms. Grouping, which was performed based on shell outline according to their specific H/L ratio, resulted in four groups (termed morphotypes / forms): form A (H/L = 0.80‒0.90), form B (H/L = 0.91‒1.02), form C (H/L = 1.03‒1.10), and form D (H/L >1.10). PCA was applied to determine the clustering of the individuals and whether manual grouping has any statistical ground. Based on the variance-covariance matrix of the log-transformed variables (L, H, Iav, La, H/L, Iva/L), the first Principal Component (PC1) is strongly positively correlated with shell size (e.g., length [L], height [H], and length of anterior area [La]) and accounts for 84.5% of the variation in the data. However, there is no distinction among the four forms with respect to their range on PC2 (Fig. 7.1). In contrast, shell outline (H/L) is strongly positively correlated with Principal Component PC2 ( = 8.427%; Fig. 7.3), where forms with higher H/L values have higher positive values on the PC2-axis (Fig. 7.1). The four forms (A–D) of Nuculoma douvillei n. sp. have significantly different PC2 (shell outline, H/L) values (p < 0.001; Fig. 7.2). PC1 versus PC3 shows high overlapping of the four forms. Moreover, the boxplot of the forms based on PC1 (shell size) shows that the four forms of N. douvillei n. sp. have a very similar size range except form B (Fig. 7.6). In summary, all forms are similar in size range but significantly differ in outline and inflation. The high overlapping of the four forms indicates that they belong to a single species (Fig. 7.1, 7.4).
Nuculoma douvillei n. sp. can be distinguished from other closely related Middle to Upper Jurassic Nuculoma species by having extremely opisthogyrate enrolled beaks, strongly inflated valves (Iav/L: 0.81–1.19), a well-developed anterior umbonal ridge, smooth inner margins, widely spaced commarginal ribs, a subtruncated to slightly convex anterior margin, and in lacking a lunule and escutcheon. The closest species is “Habonucula” agrawali Singh and Kanjilal, Reference Singh and Kanjilal1977, from the Lower Callovian of western India (Singh and Kanjilal, Reference Singh and Kanjilal1977, p. 190, pl. 1, figs. 1–12). The latter species resembles N. douvillei n. sp. in lacking escutcheon and lunule, but differs in having a more elongated-ovate and smooth valve (H/L = 0.67, holotype as opposed to 1.10 on average; Table 1), a less-incurved umbo, irregular fine commarginal ribs, in lacking an anterior umbonal ridge, and in being less inflated (Iav/L = 0.29 as opposed to 0.96 on average). Nucula variabilis J. de C. Sowerby, Reference Sowerby1825 (p. 117, pl. 475, fig. 2) from the Middle Jurassic of England differs from the present species by having a more centrally placed umbo, a less-incurved beak, a well-defined escutcheon, and in being less inflated and smooth. Douvillé (Reference Douvillé1916) doubtfully identified Nuculoma variabilis from the Middle Jurassic rocks of Egypt (Gebel Arousiah, Sinai). Recently, Abdelhady (Reference Abdelhady2014) and Abdelhady and Fürsich (Reference Abdelhady and Fürsich2014, Reference Abdelhady and Fürsich2015a) followed Douvillé and assigned their specimens, which had been collected from the same area (Gebel Arousiah), to N. variabilis. In fact, the specimens figured by Douvillé (Reference Douvillé1916) and Abdelhady and Fürsich (Reference Abdelhady and Fürsich2014, Reference Abdelhady and Fürsich2015a) are identical to N. douvillei n. sp., and therefore, have been included in the latter.
Although N. castor (d'Orbigny, 1850) of Cossmann in Thiéry and Cossmann (Reference Thiéry and Cossmann1907, p. 55, pl. 2, figs. 14, 15) from the Callovian of France also has an oblique valve, it differs from N. douvillei n. sp. by its much more oblique valve, strongly convex posterior and ventral margins, a less-incurved umbo, a smooth valve, and in being less inflated and more elongated than the present species. With respect to shell size and outline, Nucula lacroixi Flamand, Reference Flamand1911, from the Bathonian of Algeria (Flamand, Reference Flamand1911, p. 905, pl. 11, fig. 8a–d) resembles N. douvillei n. sp., but differs in having much more enrolled umbones, a shallow escutcheon, strongly convex posterior and ventral margins, and by details of the ornamentation pattern, especially at the umbonal area, which is smooth.
The other closely related species are N. wynnei Cox, Reference Cox1940 (p. 23, pl. 1, figs. 29–31) and N. blakei Cox, Reference Cox1940, (p. 24, pl. 1, figs. 24–28) from the Upper Bathonian of India. Cox (Reference Cox1940) differentiated N. blakei from N. wynnei by having a sinuous ventral margin and by its elongated form. Later, Kanjilal (Reference Kanjilal1980, p. 333), Agrawal and Kachhara (Reference Agrawal and Kachhara1980, p. 474), Pandey and Agrawal (Reference Pandey and Agrawal1984, p. 180), and Jaitly et al. (Reference Jaitly, Fürsich and Heinze1995, p. 155, text-fig. 5) recorded numerous intermediate forms between N. blakei and N. wynnei and concluded that the two species of Cox are conspecific. Therefore, they regarded N. blakei as a junior synonym of N. wynnei, arguing that some transitional forms exist between the two species. Their view is followed here. The two species differ from N. douvillei n. sp. in having rounded ridges delineating the escutcheon, forming a sunken heart shape between the umbo and posteroventral margin (e.g., Cox, Reference Cox1940, pl. 1, figs. 24c, 30b), fine and dense commarginal ribs, and in lacking anterior umbonal ridges. Nuculoma kathrynae Duff, Reference Duff1978 (p. 21, pl. 1, figs. 2–5) from the Callovian of England resembles the present species in having strongly inflated valves and strongly opisthogyrate enrolled umbones, but it has a small cordate escutcheon, which reaches about halfway to the posteroventral angle, and its surface carries irregular commarginal growth lines. Nucula pollux d'Orbigny, 1850 (p. 339, no. 179) (Cottreau, Reference Cottreau1925, p. 154, pl. 39, figs. 25–27) from the Callovian–Oxfordian of France differs in having a deep escutcheon, less-inflated valves, and more centrally placed umbones.
Nuculoma sinaiensis new species
Figures 5.20–5.38, 8.1–8.35, 9, 10
- non .Reference Sowerby1840
Nucula tenuistriata J. de C. Sowerby (nomen dubium), pl. 22, fig. 3.
- .Reference Douvillé1916
Nucula tenuistriata; Douvillé, p. 61, pl. 5, figs. 46–50 (non J. de C. Sowerby).
- .Reference Hirsch1980
Palaeonucula tenuistriata; Hirsch, p. 130, pl. 1, fig. 7.
- ?Reference Parnes1981
Nuculoma sp. cf. Nucula tenuistriata; Parnes, p. 26, pl. 3, fig. 17.
- ?Reference Holzapfel1998
Nuculoma sp. Holzapfel, p. 94, pl. 3, fig. 1a–c.
- ?Reference Abdelhamid2002
Palaeonucula tenuistriata; Abdelhamid, p. 337, pl. 5, fig. 4.
- v.Reference Abdelhady2014
Palaeonucula tenuistriata; Abdelhady, p. 67.
Holotype
Articulated specimen BSPG 2014V 303/1 with shell preservation from the upper Bajocian Bir Maghara Formation, Gebel Arousiah, Sinai (Fig. 5.20–5.24).
Paratypes
One hundred twenty-eight specimens in shell preservation, mostly articulated, from the upper Lower Jurassic (Toarcian) to Middle–Upper Jurassic (Bajocian–Callovian and Lower Kimmeridgian) of Gebel Maghara: three specimens from the upper part of the lower Toarcian Rajabiah Formation (marl unit), Bir Maghara (BSPG 2014V 317/1–3), 14 specimens from the upper Toarcian Shusha Formation, Bir Maghara (BSPG 2014V 314/1–14); 16 specimens from the upper Bajocian Bir Maghara Formation, Gebel Arousiah (BSPG 2014V 303/2–17); six specimens from the upper third part of the lower Bathonian Safa Formation, Bir Maghara (BSPG 2014V 318/1–6); seven specimens from the middle–upper Bathonian Kehailia Formation, Gebel Engabashi (BSPG 2014V 304/1–2; 315/1–5); 13 specimens from the same formation at Gebel Homayir (BSPG 2014V 305/1–6) and Gebel Mowerib (BSPG 2014V 306/1–7); two specimens from the Callovian Arousiah Formation, Gebel Mowerib (BSPG 2014V 327/1–2); 49 specimens from the lower marl beds of the lower Kimmeridgian Masajid Formation, western Bir Maghara (BSPG 2014V 312/1–22; 316/1–27); and 18 specimens from the upper part of the Masajid Formation, western Bir Maghara (BSPG 2014V 313/1–18).
Diagnosis
Small strongly inflated Nuculoma species, variable in outline, ranging from elongated-ovate to subtrapezoidal, from subrounded to subtrigonal; posterior margin truncated to slightly convex, anterior margin strongly convex; escutcheon lacking (occasionally delimited by a faint ridge, shallow cordate form), no lunule, anisomyarian with large subquadrate anterior adductor muscle scar and small elongated-ovate posterior muscle scar; pallial line entire (located some distance from ventral margin); umbones terminal to subterminal, projecting occasionally beyond posterior margin, causing posterior margin to be strongly oblique; growth lines irregularly spaced, fine.
Occurrence
Toarcian to lower Kimmeridgian, Gebel Maghara, Sinai.
Description
Shell small-sized, variable in outline, elongated-ovate to subtrapezoidal, subrounded to subtrigonal, longer than high (H/L = 0.82 on average; Table 2), equivalved, strongly inequilateral, and strongly inflated. Inflation greatest approximately one-third of total valve height from umbo. Anterior margin strongly rounded, meeting regularly convex ventral margin in rounded curve. Posterior margin truncated to slightly convex, strongly oblique, meeting ventral margin in a continuous rounded curve. Anterodorsal margin long, straight to slightly convex, and higher than posterodorsal one. Umbones strongly inflated, terminal to sub-terminal, strongly enrolled posteriorly, occasionally projecting beyond posterior margin at approximately mid-height of valve, forming a strongly oblique posterior margin (Fig. 5.28–5.31). Beaks sharply pointed and opisthogyrate. Escutcheon absent except in a few specimens where it is indistinct, cordate-shaped (Fig. 8.13). Posterior slope (= corselet) occasionally well developed, extending from umbo to about one-third of posterodorsal margin (Fig. 5.35). No lunule (Figs. 5.21, 8.23). Anterior adductor muscle scar shallowly impressed, large (~41% of total shell height), subrectangular to elongated-ovate, located close to anterior margin. Posterior adductor muscle scar smaller than anterior one, subrounded to elongated-ovate, its dorsal margin flat, located close to posterodorsal margin below posterior teeth (Fig. 8.28, 8.30, 8.31). Pallial line thick, entire, located some distance from ventral margin (approximately one-third of total shell height from ventral margin; Fig. 8.28, 8.31). Hinge plate broad, slightly arched, with strong unequal teeth and sockets. Anterior hinge row with 16–25 (21 on average) convexodont teeth, gradually decreasing in size towards umbo (Fig. 8.33–8.35). Posterior part of hinge short, wide, with strong 3–5 orthomorphodont to slightly concavodont teeth, gradually increasing in size towards umbo (Fig. 8.33). The last one or two posterior teeth (below umbo) occasionally larger and thicker than others, with oblique triangular chondrophore, extending sub-horizontally below the first five smallest anterior teeth. Surface with fine, irregularly spaced, numerous commarginal growth lines of variable strength.
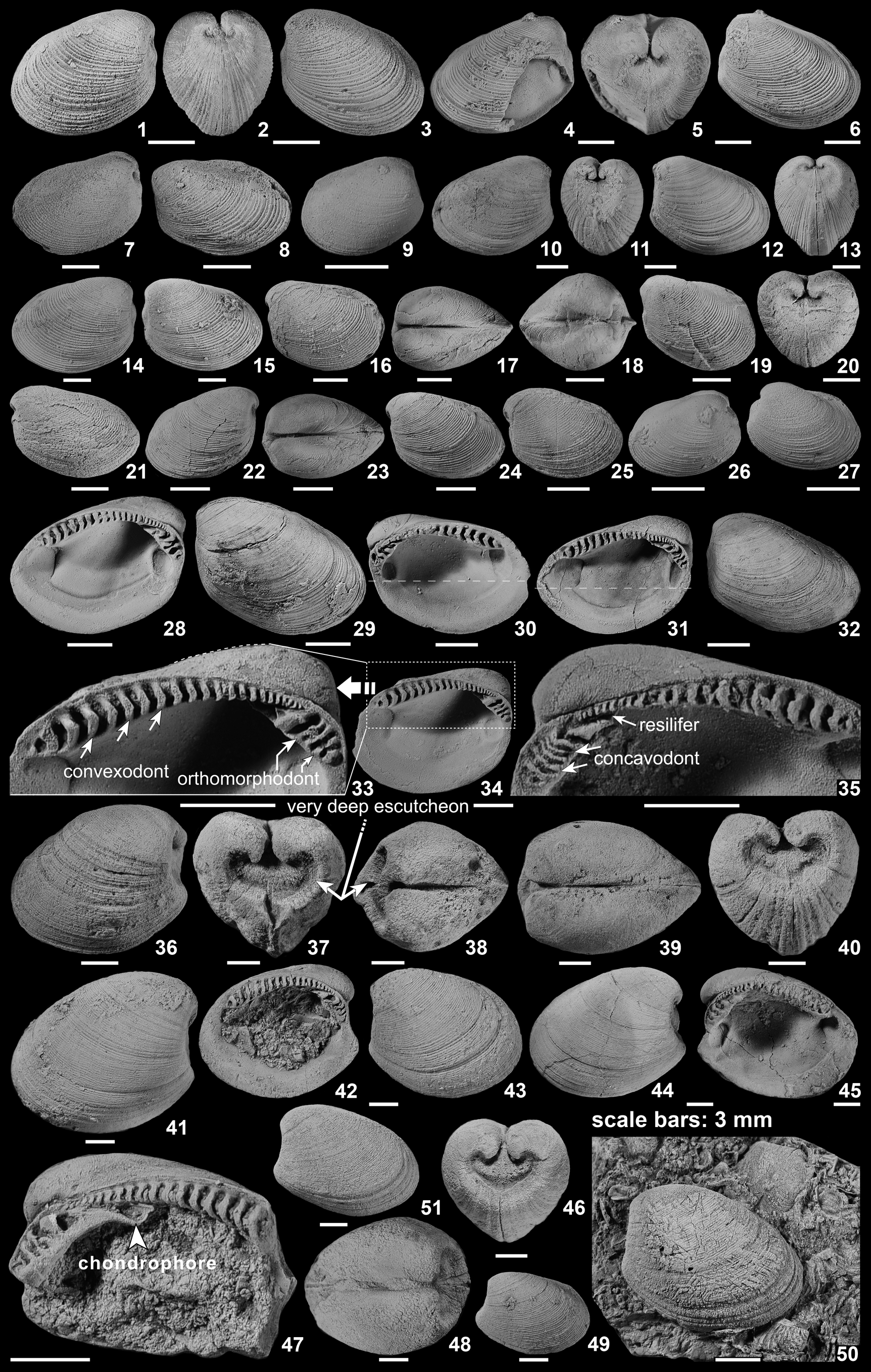
Figure 8. (1–35) Paratypes of Nuculoma sinaiensis n. sp. from the Jurassic of Gebel Maghara, showing variation in outline and internal structures. (1–15) Masajid Formation, western Bir Maghara; (1–3) BSPG 2014V 316/1, left lateral view, posterior view showing the absence of escutcheon, and right lateral view, respectively; (4–6) BSPG 2014V 316/2, left lateral view, posterior view, and right lateral view, respectively; (7) BSPG 2014V 316/3, left lateral view; (8) BSPG 2014V 316/4, right lateral view, sub-trapezoidal form of species; (9) BSPG 2014V 316/5, left lateral view, (10–12) BSPG 2014V 313/4, left lateral view, posterior view of articulated valves showing absence of escutcheon, and right lateral view, respectively; (13) BSPG 2014V 313/5, posterior view, with a faint cordate escutcheon; (14, 15) BSPG 2014V 313/6, left lateral view of sub-rounded form of species and right lateral view, respectively. (16–25) Bir Maghara Formation, Gebel Arousiah; (16, 17) BSPG 2014V 303/2, right lateral view and anterodorsal view showing the absence of lunule, respectively (18, 19) BSPG 2014V 303/3, dorsal and right lateral views respectively, (20, 21) BSPG 2014V 303/4, posterior and right lateral view, respectively, (22–24) BSPG 2014V 303/5, left lateral view of sub-trapezoidal form, anterodorsal view showing lack of a lunule, and right lateral view showing the beak projecting beyond posterior margin, respectively, (25) BSPG 2014V 303/6, right lateral view; (26, 27) BSPG 2014V 315/1, Kehailia Formation, Gebel Engabashi; (26) left lateral and right lateral views, respectively, of elongated-ovate form of species; (28–30) Kehailia Formation, Gebel Homayir; (28, 29) BSPG 2014V 305/1, right valve interior view showing entire pallial line located some distance from ventral margin, and exterior view, (30) BSPG 2014V 305/2, left lateral view showing small rounded posterior adductor muscle scar. (31, 32) Kehailia Formation, Gebel Mowerib; (31) BSPG 2014V 306/1, right valve interior showing large rectangular anterior muscle scar and right valve exterior, respectively. (33–35) Kehailia Formation, Gebel Homayir; (33) BSPG 2014V 305/3, close-up showing anterior and posterior teeth, and right valve interior, respectively, (35) BSPG 2014V 305/4, close-up of hinge. (36–51) Nuculoma wynnei Cox (Reference Cox1940) from the Callovian of Kachchh, India. (36–40) Jumara Dome; (36) PIW 1991 III23/1, left lateral view, (37, 38) PIW 1991 III23/2, posterior view showing a very deep escutcheon, and dorsal view, respectively, (39) PIW 1991 III23/3, anterodorsal view, (40) PIW 1991 III23/4, posterior view showing a rounded ridge delineating the escutcheon. (41–43) Kaladongar, Pachchham Island; (41) PIW 1991 III32/1, left valve, (42, 43) PIW 1991 III32/2, right valve interior showing angulated posterior margin, and right valve, respectively. (44–46) Jhura Dome; (44, 45) PIW 1991 III29/1, left lateral view, and left valve view showing the internal features, respectively, (46) PIW 1991 III29/2, posterior view. (47–49) Jhura Dome; (47) PIW 1991 III46/1, left valve wshowing teeth and chondrophore, PIW 1991 III46/2, dorsal view, and PIW 1991 III46/1, right lateral view, respectively. (50, 51) Jumara Dome; (50) PIW 1991 III23/5, right valve and right lateral view, respectively, showing angulated posterior margin. Scale bars = 3 mm.
Table 2. Measurements (in mm) of Nuculoma sinaiensis n. sp.

Etymology
After Sinai Peninsula, northeastern Egypt.
Measurements
See Table 2.
Remarks
Nuculoma sinaiensis n. sp. can be distinguished from other Jurassic Nuculoma species by its small size (maximum length: 13.7 mm; maximum height: 11.2 mm; maximum inflation: 9.8 mm; Table 2); terminal to subterminal umbones projecting occasionally beyond posterior margin at nearly mid-height of valve, and strongly oblique posterior margin; a thick, entire pallial line, which extends some distance from ventral margin, and in lacking a lunule and escutcheon (occasionally present as very faint, cordate-shaped feature; Fig. 8.13). The specimen figured by Holzapfel (Reference Holzapfel1998) as Nuculoma sp. from the upper Callovian–Oxfordian of southern Tunisia strongly resembles the present species in having strongly enrolled terminal umbones, strongly inflated valves, and in lacking an escutcheon, but its posterior margin is much more rounded than the Nuculoma sinaiensis n. sp. “Habonucula” agrawali Singh and Kanjilal, Reference Singh and Kanjilal1977, (p. 190, pl. 1, figs. 1–12) from the lower Callovian of western India also lacks an escutcheon, but differs in having a rounded posterior margin (sub-straight in the present species), a nearly smooth valve except for faint commarginal growth lines close to the ventral margin, and in being less inflated (holotype: Iav/L = 29.3%; paratypes: 38.5–58.3%; Singh and Kanjilal, Reference Singh and Kanjilal1977, p. 192) and more elongated (H/L: 0.67–0.73 as opposed to 0.69–0.97). In addition, the anterior teeth of “H.” agrawali appear to be straight and stronger than those of the present species (variable in N. sinaiensis n. sp.). Another closely related taxon is Nucula tenuistriata J. de C. Sowerby, which was recorded from the Middle Jurassic (?Callovian) of India by J. de C. Sowerby (Reference Sowerby1840, pl. 22, fig. 3). Cox (Reference Cox1940, p. 23) included Douvillé's specimens of N. tenuistriata (1916, pl. 5, figs. 46–50) from the Middle Jurassic of Sinai (Egypt) in his new species Nuculoma wynnei Cox, Reference Cox1940, (p. 23, pl. 1, figs. 29–31) from the Middle Jurassic of India—a view not accepted here, although Jaitly et al. (Reference Jaitly, Fürsich and Heinze1995, p. 154) included not only the Egyptian material in the synonymy list of N. wynnei as suggested by Cox, but questionably also the Indian holotype of Nucula tenuistriata J. de C. Sowerby. The Egyptian material, however, is regarded herein as a separate species for the following reasons: (1) Douvillé (Reference Douvillé1916) did not illustrate either posterior or dorsal views to define the development of an escutcheon, which distinguishes it from other Nuculoma species; (2) the specimens figured by Douvillé have a sub-truncated posterior margin (angulated in N. wynnei; Fig. 8.42, 8.44, 8.51), terminal umbones, irregularly spaced fine commarginal growth lines; and (3) in N. wynnei, the rounded ridge, delineating the corselet sensu Duff (Reference Duff1978) and others (see Cox, Reference Cox1940, pl. 1, figs. 29b, 30b; Jaitly et al., Reference Jaitly, Fürsich and Heinze1995, pl. 1, fig. 2c; Fig. 8.37, 8.38), forms a concave arch between the umbo and the posteroventral margin enclosing a well-defined, deep heart-shaped escutcheon (absent in the Egyptian material). Figure 9 shows a detailed comparison of N. sinaiensis n. sp. and N. wynnei. In addition, Principal Component Analysis (PCA) was carried out to show the shell size and morphological relationship between N. wynnei and N. sinaiensis n. sp. based on the variance-covariance (VCV) matrix of log-transformed variables such as length (L), height (H), inflation (Iav), height/length ratios (H/L), and inflation/length ratios (Iav/L) (Fig. 10.1, 10.2). The first Principal Component (PC1) accounts for 96.24% of the variation in the data, which positively correlates with shell size (H, L, Iav), whereas PC2 accounts for 2.0%, which positively correlates with shell outline (H/L) and inflation (Iav/L). The plots of PC1 versus PC2 indicate that N. wynnei and N. sinaiensis n. sp. differ in shell size as indicated by the near separation of the two convex hulls of the two species (Fig. 10.1). The boxplots of shell size of both species also show a distinct gap (median of N. wynnei 0.819, median of N. sinaiensis n. sp. −0.862; Fig. 10.3), whereby N. wynnei is distinctly larger than N. sinaiensis n. sp. Although the PC2 values of N. wynnei completely fall within the range of N. sinaiensis n. sp. (PC2), the boxplots based on height/length ratios indicate that some Egyptian specimens are more elongated than the Indian ones (Fig. 10.4). Douvillé (Reference Douvillé1916) doubtfully regarded Nucula venusta Terquem and Jourdy, Reference Terquem and Jourdy1871, from the Bathonian rocks of France as synonym of P. tenuistriata. However, the holotype of Terquem and Jourdy (Reference Terquem and Jourdy1871, pl. 11, figs. 26–28) has a less-incurved, orthogyrate to slightly prosogyrate umbo.

Figure 9. Comparison between Nuculoma sinaiensis n. sp. from the Toarcian–lower Kimmeridgian of Gebel Maghara, Egypt, and N. wynnei Cox, Reference Cox1940, from the Callovian of Kachchh Basin (western India) based on external and internal characters. All scale bars = 5 mm.

Figure 10. Principal component analysis (PCA) of Nuculoma sinaiensis n. sp. from the Jurassic rocks (Toarcian–lower Kimmeridgian) of Gebel Maghara and N. wynnei Cox, Reference Cox1940, from the Callovian of the Kachchh Basin (western India) using the PAST software (Hammer et al., Reference Hammer, Harper and Ryan2001). (1) Scatter plot of PC1 vs. PC2 showing nearly complete separation between the species from Egypt and India. (2) Percentage of variation explained by PCA of morphological variables. (3) Boxplot of the two species based on shell size (PC1). (4) Boxplot of the two species based on shell outline (H/L). (5) Scatter plot of log-transformed height versus length of N. sinaiensis n. sp. The specimens from Kachchh are from the Fürsich collection in the Bayerische Staatssammlung für Paläontologie und Geologie, Munich. nind = number of individuals. See Appendix B for data.
Cox (Reference Cox1940, p. 24, pl. 1, figs. 24–28) erected Nuculoma blakei from the upper Bathonian–Callovian of Kachchh and differentiated it from N. wynnei by its elongated shape, an obtuse angle between posterior and ventral margins, and by its more sinuous ventral margin. Actually, N. blakei carries some similarities to N. sinaiensis n. sp., particularly the terminal umbones, which extend beyond the posterior margin (e.g., Cox, Reference Cox1940, pl. 1, fig. 24a), but differs in having a well-defined, deep heart-shaped escutcheon, an angulated posterior margin (Cox, Reference Cox1940, pl. 1, fig. 27a, c), in lacking a lunule, and in being larger and more elongated than N. sinaiensis n. sp. (L = 26.0 mm, H = 16.0 as opposed to L = 10.40, H = 8.53 mm on average; H/L = 0.61 as opposed to 0.82 on average; Table 2). Jaitly et al. (Reference Jaitly, Fürsich and Heinze1995) regarded N. blakei and N. wynnei as conspecific, with N. blakei being a junior synonym, based on numerous intermediate morphotypes recorded by some authors (e.g., Agrawal, Reference Agrawal1956, p. 54; Agrawal and Kachhara, Reference Agrawal and Kachhara1980, p. 476–477; Kanjilal, Reference Kanjilal1980, p. 333; Pandey and Agrawal, Reference Pandey and Agrawal1984, p. 180). Nucula (Nuculoma) castor d'Orbigny, 1850, described and figured by Thiéry and Cossmann (Reference Thiéry and Cossmann1907, p. 55, pl. 2, figs. 14, 15) and Cossmann (Reference Cossmann1924, p. 42, pl. 6, figs. 32–35) from the Callovian of France differs in having a strongly oblique subtriangular valve, a subterminal umbo, a rounded anterior muscle scar, and in being less elongated and more strongly inflated than the present species.
Subfamily Palaeonuculinae Carter, Reference Carter2001
Genus Palaeonucula Quenstedt, Reference Quenstedt1930
Type species
Nucula hammeri Defrance, Reference Defrance1825, from the Upper Lias of France; original designation by Quenstedt (Reference Quenstedt1930).
Diagnosis
An updated diagnosis has been provided by Hodges (Reference Hodges2000, p. 13).
Remarks
There is some confusion regarding the generic/subgeneric relationships among Palaeonucula Quenstedt, Reference Quenstedt1930, Nuculopsis Girty, Reference Girty1911, and Nuculoma Cossmann, Reference Cossmann1907. For instance, Quenstedt (Reference Quenstedt1930) regarded Palaeonucula as a subgenus of the Paleozoic genus Nuculopsis. Cox (Reference Cox1940, p. 12) noted that the latter genus has a more primitive type of chondrophore than Palaeonucula, in which the chondrophore is wider and shorter, and not projecting into the shell cavity. Therefore, he placed Palaeonucula as a subgenus in Nucula, and accepted Nuculoma as a genus. The general shell morphology of Palaeonucula is similar to Nucula Lamarck, but the shell is non-nacreous and has smooth shell margins (Damborenea and Pagani, Reference Damborenea and Pagani2019, p. 929). In addition, species of Nucula should be restricted to forms with radial ribbing on the shell surface, which clearly distinguishes them from both Palaeonucula and Nuculoma (Duff, Reference Duff1978, p. 23). Recently, Hodges (Reference Hodges2000, p. 13) stated that the main differences between Palaeonucula and Nuculoma are that the latter has more strongly enrolled opisthogyrate umbones and fine, regular commarginal growth lines on the shell surface. For more details, see Duff (Reference Duff1978), Hodges (Reference Hodges2000), and Damborenea and Pagani (Reference Damborenea and Pagani2019, p. 929). The paleogeographic distribution of Palaeonucula during Middle–Late Triassic and Early Jurassic has been summarized by Ros et al. (Reference Ros, Márquez-Aliaga and Damborenea2014, p. 10, fig. 1).
Palaeonucula cuneiformis (J. de C. Sowerby, Reference Sowerby1840)
Figures 11.1–11.32, 12, 13
- *Reference Sowerby1840
Nucula? cuneiformis J. de C. Sowerby, pl. 22, fig. 4 and explanation.
- Reference Holdhaus1913
Nucula spitiensis Holdhaus, p. 428, pl. 95, figs. 11–13.
- Reference Holdhaus1913
Nucula hyomorpha Holdhaus, p. 430, pl. 95, figs. 14–17.
- .Reference Douvillé1916
Nucula lateralis; Douvillé, p. 60, pl. 5, fig. 45.
- non Reference Stefanini1939
Nucula cuneiformis; Stefanini, p. 219, pl. 24 (25), fig. 3.
- .Reference Cox1940
Nucula (Palaeonucula) cuneiformis; Cox, p. 13, pl. 1, figs. 5–10.
- .Reference Cox1940
Nucula (Palaeonucula) kaoraensis Cox, p. 15, pl. 1, figs. 11–14.
- .Reference Agrawal1956
Nucula (Palaeonucula) kaoraensis; Agrawal, p. 51, pl. 7, fig. 3a.
- .Reference Jaboli1959
Nucula cuneiformis; Jaboli, p. 46, pl. 6, fig. 3.
- .Reference Hirsch1980
Palaeonucula lateralis; Hirsch, p. 130, pl. 1, figs. 5, 6.
- .Reference Kanjilal1980
Palaeonucula kaoraensis; Kanjilal, p. 335, pl. 1, figs. 8–10.
- .Reference Kanjilal1980
Palaeonucula cuneiformis; Kanjilal, p. 334, pl. 1, figs. 4–7.
- .Reference Jaitly, Fürsich and Heinze1995
Palaeonucula cuneiformis; Jaitly et al., p. 155, pl. 1, figs. 8–17, pl. 2, figs. 1, 2, text-figs. 6–9.
- .Reference Kanjilal and Pathak1998
Palaeonucula cuneiformis; Kanjilal and Pathak, p. 30, pl. 1, fig. 1.
- .Reference Abdelhamid2002
Palaeonucula lateralis; Abdelhamid, p. 337, pl. 5, figs. 1, 2.
- .Reference Khalil2003
Palaeonucula lateralis; Khalil, p. 308, pl. 1, figs. 9, 10.
- v.Reference Abdelhady2014
Palaeonucula lateralis; Abdelhady, p. 72, fig. 5.4I, J.
- v.Reference Abdelhady and Fürsich2014
Palaeonucula lateralis; Abdelhady and Fürsich, p. 181, fig. 6I, J.
- v.Reference Fürsich, Heinze and Ayoub-Hannaa2019
Palaeonucula cuneiformis; Fürsich et al., p. 146, pl. 1, figs. 1–3.
Holotype
Nucula cuneiformis J. de C. Sowerby, Reference Sowerby1840, pl. 22, fig. 4, from the “Upper Secondary Formation” (Callovian) of Khera Hill (Keera Dome), Kachchh, western India.
Occurrence
The species has a wide stratigraphic range, from Bajocian to Kimmeridgian strata of Kachchh, western India, upper Bathonian–Callovian strata of Madagascar, Middle–Upper Jurassic of Ethiopia, and from the Lower Jurassic (Toarcian) to early Kimmeridgian successions of Gebel Maghara, Sinai (present study, first record).
Description
Shell small to medium-sized, highly variable in shape, ranging from elongated ovate to subtriangular, from sub-trapezoidal to strongly elongated (Fig. 12), inequilateral, equivalved, moderately inflated. Anterior area much longer than posterior area (Al/L: 0.67–0.80; Table 3). Anterodorsal margin slightly convex, meeting anterior margin in rounded curve. Posterodorsal margin short, weakly convex. Posterior margin narrow, angulated. Anterior end tapering, rounded, meeting ventral margin in rounded curve. Ventral margin irregularly and strongly convex (occasionally regularly convex). Umbo broad, slightly depressed, inflated, located approximately one-third of total valve length from posterior end (occasionally subterminal). Beaks pointed, slightly opisthogyrate. Anterior and posterior muscle scars oval, subequal, located close to anterior and posterior margins, respectively, and more strongly impressed towards dorsal margin (Fig. 11.12). Pallial line well-impressed, continuous, parallel to ventral margin. Anterior pedal retractor muscle scars (Apr) small, rounded, and located close to anterodorsal margin, below anterior hinge (Fig. 11.29–11.32). Escutcheon well developed, sub-ovate to cordiform, deep. Lunule generally absent, but occasionally faint, very shallow, lanceolate, narrow in few specimens. A well-developed rounded umbonal ridge running to posteroventral angle of shell with a slightly concave area located close to posterior margin (Figs. 11.6, 11.11, 13.1, 13.2). Hinge taxodont with numerous unequal teeth and sockets. Anterior part of hinge longer than posterior one with ~22 small convexodont teeth, decreasing in size towards umbo (Fig. 11.28, 11.32). Posterior hinge with 8–10 convexodont teeth and sockets, gradually decreasing in size towards umbo. Chondrophore well developed, spoon-shaped, projecting sub-horizontally below the smallest teeth of anterior hinge close to beak (Fig. 11.32). Shell surface covered with irregularly spaced fine commarginal growth lines, variable in strength.

Figure 11. (1–32) Variation and internal characters of Palaeonucula cuneiformis (J. de C. Sowerby, Reference Sowerby1840) from the Lower–Middle Jurassic of Gebel Maghara. (1, 2) BSPG 2014V 328/1, Rajabiah Formation, western Bir Maghara; (1) left lateral view, elongated-ovate form, and left valve, interior, respectively. (3–5) BSPG 2014V 196/1, Kehailia Formation, Gebel Homayir; (3) left lateral, right lateral, and dorsal views of articulated valves; (6, 7) BSPG 2014V 188/1, Bir Maghara Formation, Gebel Arousiah; (6) left lateral view showing narrowly rounded posterior margin and shallow posterior sulcus,and right lateral view, respectively; (8, 9) BSPG 2014V 329/1, Masajid Formation, western Bir Maghara, left lateral and dorsal views, respectively; (10) BSPG 2014V 300/1, Kehailia Formation, Gebel Homayir, right lateral view; (11–18) Kehailia Formation, Gebel Homayir; (11, 12) BSPG 2014V 195/1, left lateral view and left valve, interior view, respectively, (13, 14) BSPG 2014V 195/2, exterior right lateral view and right valve interior view, respectively (15, 16) BSPG 2014V 195/3, left lateral and right lateral views, respectively, of elongated-ovate form, (17, 18) BSPG 2014V 195/4, subtriangular form of species, left lateral view of exterior and interior left valve view, respectively, showing hinge teeth and muscle scars; (19–21) BSPG 2014V 330/1, Masajid Formation, western Bir Maghara, (19) dorsal view, (20) left lateral view showing a shallow umbonal posterior sulcus, (21) right valve; (22) BSPG 2014V 199/1, Bir Maghara Formation, Gebel Arousiah, left lateral view, shell partly preserved; (23) BSPG 2014V 198/1, Kehailia Formation, Gebel Homayir, right lateral view, composite mold, subtriangular form; (24) BSPG 2014V 185/1, Kehailia Formation, Gebel, Engabashi, left valve, elongated form; (25) BSPG 2014V 194/1, Kehailia Formation, Gebel, Arousiah, left valve. (26–30) Internal characters of P. cuneiformis from the Kehailia Formation, Gebel Engabashi; (26) BSPG 2014V 186/1, right lateral view, interior, (27, 28) BSPG 2014V 186/2, left lateral and close-up views, respectively, showing anterior and posterior convexodont teeth, (29) BSPG 2014V 186/3, left lateral view showing well-developed anterior pedal retractor muscle scars (Apr), (30) BSPG 2014V 186/4, left lateral view showing entire pallial line, Apr (arrowed), and muscle scars; (31, 32) BSPG 2014V 200/1, Kehailia Formation, Gebel Engabashi, left valve and close-up, respectively, showing sub-horizontally projecting spoon-shaped chondrophore, Apr (arrowed), and teeth. (33–44) Palaeonucula fraasi (Noetling, Reference Noetling1887) from the Middle–Upper Jurassic of Gebel Maghara. (33–35) BSPG 2014V 179/1, Kehailia Formation, Gebel Mowerib, left lateral, right valve, and dorsal views, respectively. (36–40) Masajid Formation, western Bir Maghara: (36, 37) BSPG 2014V 334/1, right lateral and dorsal views, respectively, (38–40) BSPG 2014V 334/2, left lateral view, dorsal view with deep escutcheon (arrowed), and right lateral view, respectively; (41–44) Bir Maghara Formation, Gebel Arousiah; (41–43) BSPG 2014V 301/1, right valve, dorsal view, and close-up, respectively, showing taxodont teeth and sockets, (44) BSPG 2014V 301/2, dorsal view showing deep escutcheon (arrowed). Scale bars = 5 mm.

Figure 12. (1–12) Variation in outline of Palaeonucula cuneiformis (J. de C. Sowerby, Reference Sowerby1840) from the Lower–Middle Jurassic of the Maghara area. Specimens from several populations. Aa, anterior adductor muscle scar, Pa, posterior adductor muscle scar, Pl, pallial line. Scale bars = 5 mm.

Figure 13. Effect of preservation quality on the shell outline of Palaeonucula cuneiformis (J. de C. Sowerby, Reference Sowerby1840) from the Lower–Middle Jurassic of the Maghara area, Sinai. (1, 2) BSPG 2014V 300/1, Kehailia Formation, Gebel Homayir, Maghara area. (3–6) Bir Maghara Formation, Gebel Arousiah; (3, 4) BSPG2014V 199/1, (5, 6) BSPG 2014V 199/2; (7, 8) BSPG 2014V 198/1, Kehailia Formation, Gebel Homayir, Maghara area. Scale bars = 5 mm.
Table 3. Measurements (in mm) of Palaeonucula cuneiformis (J. de C. Sowerby, Reference Sowerby1840). Sv = inflation of the single valve.

Materials
One hundred and two specimens, mostly articulated, in shell preservation, from the Lower–Middle Jurassic of the Maghara area, Sinai: a single specimen from the Toarcian Rajabiah Formation, western Bir Maghara (BSPG 2014V 328/1); two specimens from the upper Bajocian Bir Maghara Formation, Gebel Engabashi (BSPG 2014V 183/1–2); 14 specimens from the same formation, Gebel Arousiah (BSPG 2014V 188/1–10; 189/1; 190/1–3); two specimens from the Lower Bathonian Safa Formation, Gebel Arousiah (BSPG 2014V 191/1–2); 20 specimens from the middle–upper Bathonian Kehailia Formation, Gebel Engabashi (BSPG 2014V 185/1; 186/1–11; 199/1–3; 200/1–4; 302/1); 13 specimens from the same formation, Gebel Homayir (BSPG 2014V 193/1–5; 195/1–5; 198/1; 300/1–2); 17 specimens from the Kehailia Formation, Gebel Arousiah (BSPG 2014V 194/1; 196/1–3; 197/1–13); 20 specimens from the same formation, Gebel Mowerib (BSPG 2014V 181/1; 202/1–19); a single specimen from the Callovian Arousiah Formation, Gebel Mowerib (BSPG 2014V 182/1); a single specimen from the same formation at Gebel Engabashi (BSPG 2014V 5/1); and 11 specimens from the basal part of the lower Kimmeridgian Masajid Formation, western Bir Maghara (BSPG 2014V 323/1–3; 329/1; 330/1; 383/1; 386/1–4; 388/1).
Measurements
See Table 3.
Remarks
It is interesting to note that Palaeonucula cuneiformis (J. de C. Sowerby, Reference Sowerby1840) preserved with shell is elongated-ovate (Fig. 13.1, 13.2), whereas specimens preserved as internal molds have completely different shapes (e.g., sub-triangular or sub-trapezoidal; Fig. 13.3–13.8), which led many authors to erect new genera and species, which are regarded as nomina dubia. With respect to general outline and length/height ratios, P. cuneiformis is, therefore, a highly variable species (Fig. 12). The present specimens fall well within the range of variation of the species as documented by Cox (Reference Cox1940, figs. 5–14) and Jaitly et al. (Reference Jaitly, Fürsich and Heinze1995, text-fig. 9) from the upper Bajocian–Tithonian of Kachchh, western India, and by Fürsich et al. (Reference Fürsich, Heinze and Ayoub-Hannaa2019, figs. 1–3) from the Callovian–Oxfordian of Madagascar. Palaeonucula cuneiformis can be distinguished from other Jurassic Palaeonucula species by having a deep, ovate escutcheon, an asymmetrical or regularly convex ventral margin, a posteriorly directed umbo, and in lacking a lunule (occasionally poorly developed, e.g., Jaitly et al., Reference Jaitly, Fürsich and Heinze1995. pl. 1, fig. 14b). Although the preservation of Nucula spitiensis and N. hyomorpha, which were erected by Holdhaus (Reference Holdhaus1913) from the Middle Jurassic of India, is poor, they fall within the range of variation of P. cuneiformis and are considered herein as synonyms of the latter. Nucula lateralis Terquem and Jourdy, Reference Terquem and Jourdy1871, has been recorded from the Middle–Upper Jurassic of Gebel Maghara (Sinai) by some authors, including Douvillé (Reference Douvillé1916), Abdelhamid (Reference Abdelhamid2002), Khalil (Reference Khalil2003), Abdelhady (Reference Abdelhady2014), and Abdelhady and Fürsich (Reference Abdelhady and Fürsich2014). Fürsich and Werner (Reference Fürsich and Werner1987, p. 108) regarded Nucula lateralis as a synonym of Palaeonucula menkii (Roemer, 1836), which has been recorded from the lower–middle Kimmeridgian of the Lusitanian Basin (Portugal). According to Fürsich and Werner (Reference Fürsich and Werner1987), the length/height ratio and the degree of regularity of the convexity of ventral margin of the latter species are highly variable, and it is, therefore, difficult to separate the various Middle and Upper Jurassic species such as Nucula lateralis Terquem and Jourdy, Reference Terquem and Jourdy1871 (p. 107, pl. 11, figs. 19, 20), N. amata d'Orbigny, 1850 (p. 310, no. 255) (Thevenin, Reference Thevenin1913, p. 150, pl. 27, figs. 26, 27), N. waltoni Morris and Lycett, 1853 (p. 52, pl. 5, fig. 14), and N. pseudo-menkii de Loriol, Reference de Loriol1901 (p. 89, pl. 5, figs. 22, 23) from P. menkii. Consequently, Fürsich and Werner (Reference Fürsich and Werner1987) regarded the latter species as junior synonyms of P. menkii, arguing that numerous transitional forms exist between two or more of these species. Actually, P. menkii (Roemer, 1836) carries some similarities to P. cuneiformis, but differs in having a sharply demarcated, narrow, and deep lunule (see the holotype of Roemer, 1836, p. 98, pl. 6, fig. 10b), and its surface is covered with widely spaced and thick commarginal ribs. Agrawal and Kachhara (Reference Agrawal and Kachhara1977), Pandey and Agrawal (Reference Pandey and Agrawal1984), and Jaitly et al. (Reference Jaitly, Fürsich and Heinze1995) regarded Nucula (Palaeonucula) kaoraensis Cox, Reference Cox1940, and N. (P.) blanfordi Cox, Reference Cox1940, from the Middle–Upper Jurassic of India as junior synonyms of P. cuneiformis, arguing that numerous transitional forms exist between the latter species (but see Fürsich et al., Reference Fürsich, Alberti, Pandey and Ayoub-Hannaa2022).
Another very close species appears to be P. hausmanni (Roemer, 1836) (p. 98, pl. 6, fig. 12a–d) from the Jurassic of North Germany. The latter species has an irregular convex ventral margin, a more prominent beak, a distinct shallow lunule, a nearly straight anterodorsal margin, and widely spaced strong commarginal ribs. According to Damborenea and Pagani (Reference Damborenea and Pagani2019, p. 930–931), the type species of the genus, P. hammeri (Defrance, Reference Defrance1825) (Quenstedt, 1851, p. 527, pl. 44, figs. 4–5; 1856, p. 313, pl. 43, figs. 7–12; Schenck, Reference Schenck1934, p. 35, pl. 2, fig. 20, pl. 4, fig. 1), from the Toarcian–Bajocian of Europe, has a more posteriorly placed umbo and a subrectangular inflated shell. The same applies to its possible synonym (Schenck, Reference Schenck1934, p. 37; Aberhan, Reference Aberhan1998, p. 64), P. hausmanni (Roemer, 1836, p. 98, pl. 6, fig. 12a–d; Kuhn, Reference Kuhn1935, p. 120, pl. 8, figs. 33a, b, 41a–d). In addition, P. hammeri has a more inflated valve, sharply pointed beaks, an asymmetrically convex ventral margin, and a distinct lunule (e.g., Quenstedt, 1856, p. 313, pl. 43, figs. 7–12). Nucula calliope d'Orbigny, 1850, figured by Cossmann (Reference Cossmann1924, p. 41, pl. 6, figs. 38–40) and Cottreau (Reference Cottreau1925, p. 153, pl. 18, figs. 21, 22) from the Callovian of France, by Makowski (Reference Makowski1952, p. 5, pl. 5, fig. 2) from the Callovian of Poland, and by Duff (Reference Duff1978, p. 25, pl. 1, figs. 14–16, 18–21) from the Callovian of England, has a more prominent beak, well-demarcated, wide, and deep lunule and escutcheon, and is less elongated and more inflated than P. cuneiformis.
Palaeonucula fraasi (Noetling, Reference Noetling1887)
Figures 11.33–11.44, 14.1–14.5
- .Reference Fraas1877
Nucula variabilis; Fraas, fig. 24 (non Sowerby).
- *Reference Noetling1887
Nucula Fraasi Noetling, p. 34, pl. 5, figs. 12, 12a, b.
- .Reference Hirsch1980
Palaeonucula fraasi; Hirsch, p. 132, pl. 1, figs. 1–4.
- .Reference Abdel-Gawad and Gameil1999
Palaeonucula fraasi; Abdel-Gawad and Gameil, p. 777, pl. 2, fig. 1.
- .Reference Abdelhamid2002
Palaeonucula fraasi; Abdelhamid, pl. 1, fig. 1.
- .Reference Khalil2003
Palaeonucula fraasi; Khalil, p. 307, pl. 1, figs. 11–16.
Holotype
Nucula fraasi Noetling, Reference Noetling1887, pl. 5, fig. 12 from the Jurassic of Hermon, Syria.
Occurrence
Lower Jurassic of Syria and Middle–Upper Jurassic (upper Bajocian–lower Oxfordian) of El Minshera area (Abdel-Gawad and Gameil, Reference Abdel-Gawad and Gameil1999) and Gebel Maghara, Sinai (Hirsch, Reference Hirsch1980; Khalil, Reference Khalil2003; present study).
Description
Shell small, outline elongated-ovate, inequilateral, equivalved, longer than high (H/L: 0.62–0.71), moderately inflated (Iav/L: 0.39–0.49; Table 4). Inflation greatest close to umbonal area. Anterior area larger than posterior one (Al/L: 0.68–0.77). Anterodorsal margin straight to slightly convex dorsally, meeting anterior margin in rounded angle. Posterodorsal margin short, slightly convex dorsally. Anterior and posterior margins convex, meeting ventral margin in rounded curves. Ventral margin regularly convex. Umbo wide, inflated, located one-third of total valve length from posterior end. Beaks small, sharply pointed, opisthogyrate. Escutcheon ovate to cordate, deep (Figs. 11.39, 11.44, 14.2). No lunule. Hinge taxodont with numerous teeth and sockets, decreasing in size towards umbo (Figs. 11.43, 14.4). Anterior part of hinge longer than posterior one with 15–20 chevron-shaped teeth and sockets. Posterior part of hinge with ~5–7 teeth and sockets. Ornamentation consisting of numerous fine commarginal ribs (Fig. 11.33, 11.34).

Figure 14. (1–5) Palaeonucula fraasi (Noetling, Reference Noetling1887) from the middle–upper Bathonian Kehailia Formation, Gebel Homayir. (1–3) BSPG 2014V 331/1, left lateral view of composite mold, dorsal view, and right valve, respectively; (4, 5) BSPG 2014V 331/2, dorsal view showing traces of taxodont hinge, and internal mold of right valve, respectively. (6–19) Palaeonucula variabilis (J. de C. Sowerby, Reference Sowerby1825) from the Middle–Upper Jurassic of Gebel Maghara; (6–8) BSPG 2014V 335/1, Masajid Formation, western Bir Maghara, left lateral view of composite mold, dorsal view, and right lateral view, respectively. (9–15) Kehailia Formation, Gebel Mowerib; (9–12) BSPG 2014V 319/1, right valve, left valve, dorsal view, and close-up, respectively, showing poorly preserved taxodont hinge, (13–15) BSPG 2014V 319/2, left lateral view showing slightly oblique sub-trigonal valve, dorsal view, and right lateral view, respectively; (16–19) BSPG 2014V 335/2, Masajid Formation, western Bir Maghara, left valve, dorsal view, right lateral view, and close-up showing fine, commarginal growth lines, respectively. (20–27) Palaeonucula sp. A. from the Middle–Upper Jurassic of Gebel Maghara; (20–23) BSPG 2014V 336/1, Kehailia Formation, Gebel Homayir, (20) dorsal view of articulated valves, (21) close-up showing traces of taxodont hinge (arrowed), (22) left lateral view, (23) right lateral view; (24–27) Masajid Formation, western Bir Maghara; (24, 25) BSPG 2014V 337/1, left lateral view and dorsal view showing deep escutcheon, respectively, (26, 27) BSPG 2014V 337/2, left lateral and dorsal views showing taxodont hinge. (28–46) Palaeoneilo aegyptiaca n. sp. from the Middle Jurassic (Bathonian‒Callovian) of Gebel Maghara. (28–30) BSPG 2014V 249/1, holotype, Kehailia Formation, Gebel Homayir, left lateral view, dorsal view showing pointed and slightly prosogyrate beaks, and right valve, respectively; (31–46) paratypes, Bir Maghara Formation, Gebel Arousiah: (31, 32) interior view of left valve and exterior view of right valve, respectively, (33, 34) BSPG 2014V 144/2, left lateral view and interior view of left valve, respectively, (35–37) BSPG 2014V 144/3, interior view of left valve, close-up showing slightly convexodont to concavo-convexodont teeth, and left valve exterior, respectively, (38, 39) BSPG 2014V 144/4 left valve and left valve interior, respectively, (40) BSPG 2014V 144/5, right valve of elongated-ovate form of species, (41–43) BSPG 2014V 144/6, left valve, internal view of left valve, and close-up, respectively, showing slight overlap of posterior teeth above anterior row of teeth below umbo and no resilifer, (44–46) BSPG 2014V 144/7, left valve, interior and dorsal views of left valve, and close-up, respectively, showing the absence of resilifer and slight overlap of posterior teeth above anterior row of teeth. Scale bars = 5 mm.
Table 4. Measurements (in mm) of Palaeonucula fraasi (Noetling, Reference Noetling1887).

Materials
Twenty-three specimens, mostly preserved as composite molds, occasionally in shell preservation, from the Middle–Upper Jurassic of Gebel Maghara: four specimens from the Upper Bajocian Bir Maghara Formation, Gebel Arousiah (BSPG 2014V 301/1–4); a single specimen from the middle–upper Bathonian Kehailia Formation, Gebel Engabashi (BSPG 2014V 333/1); 16 specimens from the same formation at Gebel Homayir (BSPG 2014V 331/1–5; 332/1), Gebel Mowerib (BSPG 2014V 179/1–5), and Gebel Arousiah (BSPG 2014V 307/1–5); and two specimens from the lower Kimmeridgian Masajid Formation, western Bir Maghara (BSPG 2014V 334/1–2).
Measurements
See Table 4.
Remarks
The present specimens correspond well to the material figured and described by Noetling (Reference Noetling1887, fig. 5, fig. 12, 12a, b) from the Lower Jurassic of Syria as Nucula fraasi. The latter species can be distinguished from other Palaeonucula species identified herein by having a deep cordate escutcheon, a regularly rounded ventral margin, numerous fine commarginal ribs, sharply pointed and slightly opisthogyrate beaks, in lacking a lunule, and in being less inflated. Noetling (Reference Noetling1887) included Nucula variabilis J. de C. Sowerby of Fraas (Reference Fraas1877) from Lower Jurassic strata of Syria in the synonymy list of his new species. This view is accepted here, because N. variabilis has a rounded-subtrigonal valve, a shallow lunule, and its umbo is located slightly posterior of mid-length of valve. Similarly, Palaeonucula cuevitana Aberhan, Reference Aberhan1994 (p. 9, pl. 1, figs. 1–4, text-fig. 2) from the Lower Jurassic (upper Hettangian) of northern Chile and P. triangularis Duff, Reference Duff1978 (p. 23, pl. 1, figs. 6–13, 17, 23, text-fig. 6) from the Middle Jurassic (Callovian) of England have rounded-triangular valves. In addition, P. cuevitana has no escutcheon and its valve is smaller than the present species (L = 7.6–9 mm, H = 5.9–7.0 mm). With respect to shell outline, another close species appears to be P. cuneiformis (J. de C. Sowerby, Reference Sowerby1840) from the Middle Jurassic of India (J. de C. Sowerby, Reference Sowerby1840, pl. 22, fig. 4), but the latter differs in being more inflated and larger in size, and in having posteriorly directed umbones, which are terminal to sub-terminal in some varieties (see Jaitly et al., Reference Jaitly, Fürsich and Heinze1995, pl. 1, figs. 8–17, pl. 2, figs. 1, 2; present study, Fig. 12).
Palaeonucula variabilis (J. de C. Sowerby, Reference Sowerby1825)
Figure 14.6–14.19
- *Reference Sowerby1825
Nucula variabilis J. de C. Sowerby, p. 117, pl. 475, fig. 2.
- .Reference Phillips1829
Nucula variabilis; Phillips, p. 151, pl. 9, fig. 11, pl. 11, fig. 19.
- .Reference Zieten1833
Nucula variabilis; Zieten, p. 77, pl. 57, fig. 9a–c.
- .Reference Morris and Lycett1853
Nucula variabilis; Morris and Lycett, p. 51, pl. 5, figs. 13, 13a, pl. 9, fig. 5.
- .Reference Quenstedt1856
Nucula variabilis; Quenstedt, p. 188, pl. 23, fig. 28.
- .Reference Quenstedt1857
Nucula variabilis; Quenstedt, p. 765, pl. 93, fig. 15.
- ?Reference Greppin1899
Nucula variabilis; Greppin, p. 98, pl. 9, figs. 2, 2a, b.
- non .Reference Douvillé1916
Nucula variabilis?; Douvillé, p. 61, pl. 5, figs. 51–55 (= Nuculoma douvillei n. sp.).
- Reference Kuhn1938
Nucula variabilis; Kuhn, p. 132, pl. 2, figs. 16a, b.
- ?Reference Romanov1973
Nucula variabilis; Romanov, p. 29, pl. 1., figs. 28, 28a.
- .Reference Pugaczewska1986
Palaeonucula variabilis; Pugaczewska, p. 50, pl. 15.4–15.7.
- non .Reference Abdelhady and Fürsich2014
Nuculoma variabilis; Abdelhady and Fürsich, p. 181, fig. 6G, H (= Nuculoma douvillei n. sp.).
Holotype
Nucula variabilis J. de C. Sowerby, Reference Sowerby1825, pl. 475, fig. 2, from the “Great Oolite” (Bathonian) of Ancliff, southern England.
Occurrence
Upper Bajocian–lower Bathonian of Poland, Bajocian of Switzerland, Aalenian–Bathonian of southeastern England, Middle Jurassic (Bajocian) of Germany, upper Bajocian of central Asia (Moldavia and Turkmenia), and middle Bathonian–lower Kimmeridgian of the Maghara area, Sinai (present study).
Description
Shell small, outline sub-trigonal, slightly oblique, inequilateral, equivalve, slightly longer than high (H/L: 0.83–0.95; Table 5), strongly inflated, and anterior length greater than posterior one. Maximum inflation at umbonal area. Anterodorsal margin slightly convex, meeting anterior margin in a nearly right angle. Posterodorsal margin short, slightly concave, meeting posterior margin in rounded angle. Anterior and posterior margins convex and meeting ventral margin in rounded curves. Ventral margin slightly convex. Umbo wide, inflated, not enrolled, located posteriorly (approximately one-third of total valve length from posterior end). Beaks sharply pointed, slightly opisthogyrate. Escutcheon well excavated, moderately wide, cordate-shaped, moderately deep. No lunule. Specimens are articulated, but the anterodorsal margin shows a taxodont hinge (Fig. 14.12). Shell thin, covered with distinct, fine, commarginal growth lines (Fig. 14.19).
Table 5. Measurements (in mm) of Palaeonucula variabilis (J. de C. Sowerby, Reference Sowerby1825).

Materials
Six specimens, in shell preservation, from the middle Bathonian–lower Kimmeridgian of Gebel Maghara: four specimens from the middle–upper Bathonian Kehailia Formation, Gebel, Mowerib (BSPG 2014V 319/1–4) and two specimens from the higher part of the Masajid Formation (lower Kimmeridgian), western Bir Maghara (BSPG 2014V 335/1–2).
Measurements
See Table 5.
Remarks
Palaeonucula variabilis (J. de C. Sowerby, Reference Sowerby1825) can be distinguished from other Jurassic Palaeonucula species such as P. triangularis Duff (Reference Duff1978, p. 23, pl. 1, figs. 6–13, 17, 23, text-fig. 6) from the Callovian of England and P. cuevitana Aberhan (Reference Aberhan1994, p. 9, pl. 1, figs. 1–4, text-fig. 2) from the Hettangian of northern Chile by having a slightly oblique sub-trigonal valve, a slightly convex ventral margin, a strongly inflated umbo, and in lacking a lunule. Moreover, its beak is located about one-third of the total valve length from the posterior end (sub-submesial in other taxa). The present specimens are closest in size, outline, and ornamentation to Nucula (Palaeonucula) stoliczkai Cox, Reference Cox1940) (p. 20, pl. 1, figs. 21–23) from the Bathonian–Callovian rocks of India, but the latter has a well-rounded ventral margin, a sharply demarcated escutcheon (Cox, Reference Cox1940, pl. 1, fig. 21b), and is much more trigonal than the present species. Nucula variabilis J. de C. Sowerby, as figured by Greppin (Reference Greppin1899) and Romanov (Reference Romanov1973) from the Bajocian of Switzerland and Russia, respectively, differ in being much more elongated than the holotype of J. de C. Sowerby (H/L = 0.83, J. de C. Sowerby, Reference Sowerby1825, pl. 475, as opposed to 0.65, Romanov, Reference Romanov1973, pl. 1.28) and in having sub-terminal umbones. With respect to shell size and outline, the specimens from Switzerland and Russia are, in fact, much closer to P. cuneiformis (J. de C. Sowerby, Reference Sowerby1840) (pl. 22, fig. 4) than to N. variabilis, and therefore, they are questionably regarded as junior synonyms of the latter. Palaeonucula navis (Piette, Reference Piette1856) of Hodges (Reference Hodges2000, p. 14, pl. 1, figs. 1–29, text-figs. 7–14) from the Lower Jurassic of England resembles Palaeonucula variabilis in having a subtrigonal shell, but differs in having a convex ventral margin, distinct escutcheon and corselet (Hodges, Reference Hodges2000, text-fig. 12b), and in being more inflated than the present species. In addition, the British species is stratigraphically older than P. variabilis (Lower Jurassic; Hodges, Reference Hodges2000, p. 21, text-fig. 14). The strongly inflated valves (maximum inflation close to ventral margin), sub-mesial umbo, and strongly convex ventral margin distinguish Palaeonucula monnandi (Chavan, Reference Chavan1952) figured by Delvene (Reference Delvene2001, p. 50, pl. 1, fig. 5) from the upper Oxfordian–lower Kimmeridgian of Spain from P. variabilis.
Palaeonucula sp. A
Figure 14.20–14.27
Description
Very small, nuculiform elongated-ovate, longer than high (H/L: 0.65–0.71), inequilateral, equivalved, moderately inflated. Anterior area greater than posterior one (Al/L: 0.76–0.81; Table 6). Anterodorsal margin slightly convex, meeting anterior one at nearly right angle. Posterodorsal margin short, sub-straight. Posterior margin narrow, blunt. Anterior margin strongly convex, narrow, meeting the regularly convex ventral margin in rounded curve. Umbo broad, inflated, located approximately one-third of total valve length from posterior end. Beaks sharply pointed, slightly opisthogyrate. Escutcheon narrow, deep, ovate (Fig. 14.25). Lunule very shallow, lanceolate, narrow. Hinge taxodont with numerous unequal teeth and sockets (Fig. 14.21, 14.27). Shell surface with fine commarginal threads.
Table 6. Measurements (in mm) of Palaeonucula sp. A.

Materials
Five specimens in shell preservation from the Middle–Upper Jurassic of Gebel Maghara: a single specimen from the middle–upper Bathonian Kehailia Formation, Gebel Homayir (BSPG 2014V 336/1) and four specimens from the basal part of the Lower Kimmeridgian Masajid Formation, western Bir Maghara (BSPG 2014V 337/1–3; BSPG 2014V 420/1).
Measurements
See Table 6.
Remarks
With regard to overall shape and shell margins, the present specimens correspond well to Palaeonucula menkii (Roemer, 1836) as figured, for example, by Roemer (1836, p. 98, pl. 6, fig. 10) from the Middle Jurassic of North Germany, by Fürsich and Werner (Reference Fürsich and Werner1987, p. 108, pl. 1, figs. 1–3) from the Upper Jurassic of Portugal, and by Jaitly et al. (Reference Jaitly, Fürsich and Heinze1995, p. 158, pl. 2, figs. 3–6) from the Middle–Upper Jurassic of western India. According to Delvene (Reference Delvene2001, p. 50), P. menkii is a medium-sized nuculid with an average length of 14.0 mm. Because the holotype of Roemer (1836, p. 98, pl. 6, fig. 10) is much larger than the present specimens (L = 19.5 mm as opposed to L = 6.0 mm on average, Table 6), it is better to keep them in open nomenclature. The present specimens resemble P. fraasi (Noetling, Reference Noetling1887) from the Lower Jurassic of Syria (Noetling, Reference Noetling1887, fig. 24) in general outline, but the latter differs in having a less-inflated valve and in being larger than the present material.
Superorder Nuculaniformii Carter, Campbell, and Campbell, Reference Carter, Campbell, Campbell, Harper, Taylor and Crame2000
Order Nuculanida Carter, Campbell, and Campbell, Reference Carter, Campbell, Campbell, Harper, Taylor and Crame2000
Superfamily Malletioidea Adams and Adams, Reference Adams and Adams1858
Family Cucullellidae Fischer, Reference Fischer1886
Subfamily Palaeoneilinae Babin, Reference Babin1966
Genus Palaeoneilo Hall and Whitfield, Reference Hall and Whitfield1869
Type species
Nuculites constricta Conrad, Reference Conrad1842, from the Middle Devonian of New York State; subsequent designation by Hall (Reference Hall1885).
Remarks
Some earlier workers (e.g., Hallam, Reference Hallam1976, Reference Hallam1977; Duff, Reference Duff1978) did not accept Palaeoneilo as a Jurassic genus, because originally it has been recorded from Paleozoic rocks (e.g., Devonian of North America; Hall and Whitfield, Reference Hall and Whitfield1869). The genus is also known from the Triassic of Europe, Mexico, Chile, New Zealand, China, and Japan and from the Jurassic of Switzerland, England, Sweden, Russia, India, and Japan (Damborenea, Reference Damborenea1987, p. 54). Duff (Reference Duff1978) included some Jurassic species of Palaeoneilo in Mesosaccella Chavan, Reference Chavan1946, due to the lack of a resilifer and presence of a shallow radial sulcus close to the posterodorsal side. Mesosaccella originally had been erected on specimens from Campanian–Maastrichtian rocks (Upper Cretaceous) of Germany and had been placed in the family Nuculanidae by Müller (Reference Müller1847, p. 17, pl. 2, fig. 1).
Recently, Hryniewicz et al. (Reference Hryniewicz, Little and Nakrem2014, p. 16) followed Duff's (Reference Duff1978) view and assigned some Jurassic Palaeoneilo species, such as P. elliptica (Goldfuss, Reference Goldfuss1837) from the Rhaetian–Pliensbachian of Europe (Palmer, Reference Palmer1973; Hodges, Reference Hodges2000) and P. morrisi (Deshayes, Reference Deshayes1853) from the Callovian of England and France (Duff, Reference Duff1978), to Mesosaccella, but on the basis of shell outline and ornamentation. Therefore, there is some confusion in the literature about the assignment of Mesozoic species to Palaeoneilo or Mesosaccella. Cox (Reference Cox1937) and Cox et al. (Reference Cox, Newell, Branson, Casey, Chavan, Coogan, Dechaseaux, Moore and Teichert1969) stated that Palaeoneilo ranged from the Ordovician to the end of the Mesozoic and that there is no reason to separate the Paleozoic and Mesozoic species in different genera—a view that is not followed here. In fact, the distinction between the two genera is very difficult, especially because the type species of Palaeoneilo (Nuculites constricta Conrad, Reference Conrad1842, p. 249) from Middle Devonian strata of New York State is poorly known. According to Cox et al. (Reference Cox, Newell, Branson, Casey, Chavan, Coogan, Dechaseaux, Moore and Teichert1969, p. N237) and Sha and Fürsich (Reference Sha and Fürsich1994, p. 40), the presence of a faint depression between anterior and posterior teeth below umbo, foreshadowing a resilifer, is typical of Mesosaccella. However, Carter (Reference Carter and Carter1990) observed a very small resilifer in the right valve of Palaeoneilo elliptica (Goldfuss, 1837) from the Lower Jurassic of Germany. In addition, some Devonian Palaeoneilo species also have a break in the alignment of the anterior and posterior hinge teeth (Hodges, Reference Hodges2000, p. 28). According to Aberhan (Reference Aberhan1998, p. 66), a feature common to both species-groups of Palaeoneilo (see Cox, Reference Cox1937) is the tendency of the posterior teeth to pass above the anterior row of teeth in the area below the umbo. In the present study, the arrangement of hinge teeth is also variable; some specimens have continuous series of anterior and posterior teeth without resilifer, whereas the smallest posterior teeth of other specimens slightly extend above the anterior teeth below the umbo (Fig. 14.36, 14.43, 14.46). It is obvious that the absence or presence of a small depression between the posterior and anterior teeth is not a distinct enough feature to differentiate between the two genera.
Nuculites constricta Conrad, Reference Conrad1842, the type species of Palaeoneilo from the Middle Devonian of New York State, has a sulcus extending from the umbo to posteroventral corner. Referring to the extensive literature on this aspect, the majority of the Triassic and Jurassic Palaeoneilo species (as well as the present material) lacks a posteroventral sulcus. The latter is confirmed by Wasmer and Hautmann in Wasmer et al. (Reference Wasmer, Hautmann, Hermann, Ware, Roohi, Ur-Rehman, Yaseen and Bucher2012, p. 1048), who stated that the posterior flank of Triassic Palaeoneilo species lacks a shallow radial posteroventral sulcus. Cox (Reference Cox1937, p. 192–193), therefore, identified two groups within the genus Palaeoneilo ranging through the Paleozoic and Mesozoic. The first group can be distinguished by having a shallow posteroventral sulcus, as in the type species of Palaeoneilo, whereas the second group lacks this feature. Although the type species of Mesosaccella (Nucula foersteri Müller, Reference Müller1847) from the Upper Cretaceous of Germany lacks the posteroventral sulcus, it exhibits a few other diagnostic features that are not present in the type of Palaeoneilo, including (1) a deeply excavated, comparatively narrow, well-demarcated lunule (absent or shallow in Palaeoneilo); (2) a long, narrow, and deep escutcheon (lacking in the type species of Palaeoneilo); (3) sharply pointed orthogyrate beaks; (4) angulated anterior and posterior ends (variable in Palaeoneilo); and (5) a relatively well-ornamented shell surface (smooth in Palaeoneilo).
Fürsich and Werner (Reference Fürsich and Werner1987, p. 111) and Sha and Fürsich (Reference Sha and Fürsich1994, p. 40) tried to amend the generic characteristics of Mesosaccella to include some additional diagnostic characters such as: (1) an elongated to slightly rostrate shell (less elongated than Palaeoneilo), (2) an entire pallial line or a shallow pallial sinus, (3) lack of a posterior sulcus, (4) straight or slightly concave anterior and posterior rows of teeth, and (5) an external amphidetic ligament. It is interesting to note that these features fit well some Cretaceous nuculanid taxa such as Nuculana speetoniensis Woods, Reference Woods1899 (p. 3, pl. 1, figs. 6, 7) and N. lineata (J. de C. Sowerby, Reference Sowerby1835) of Woods (Reference Woods1899, p. 7, pl. 1, figs. 28–32) from the Aptian–Albian of England; Nuculana? mutuata Stephenson, Reference Stephenson1953 (p. 57, pl. 10, figs. 10–12) from the Cenomanian of Texas, Mesosaccella donganensis Sha and Fürsich, Reference Sha and Fürsich1994) (p. 40, pl. 1, figs. 8–10, text-figs. 28, 29), and M.? wunsuliensis Sha and Fürsich, Reference Sha and Fürsich1994 (p. 41, pl. 1, figs. 3–7, text-figs. 30, 31) from the Lower Cretaceous (Berriasian–Valanginian) of northeastern China.
For the reasons stated above, it is better to assign these Cretaceous species to Mesosaccella (as a Cretaceous genus). In agreement with Hodges (Reference Hodges2000), Palaeoneilo is thought to be a conservative genus morphologically that changed very little in general shape from the Ordovician to Early Jurassic time interval. Ros et al. (Reference Ros, Márquez-Aliaga and Damborenea2014, p. 16) pointed out that the last occurrence of Palaeoneilo is in the upper Lower Jurassic (Toarcian) and did not accept any younger records, “since almost all have descriptive problems.” As is demonstrated here, the genus certainly also occurs in the Middle Jurassic (Bajocian‒Bathonian) of Gebel Maghara. This is its first record from Middle Jurassic rocks, not only in Egypt but also in North Africa (see Ros et al., Reference Ros, Márquez-Aliaga and Damborenea2014, fig. 3; Fig. 15).

Figure 15. Paleogeographic distribution of Palaeoneilo Hall and Whitfield, Reference Hall and Whitfield1869, and Dacryomya Agassiz, Reference Agassiz1840, during Upper Triassic and Jurassic time (after Scotese, Reference Scotese2001). Data taken from Hayami (Reference Hayami1961), Cox (Reference Cox1965), Choubert and Faure-Muret (Reference Choubert and Faure-Muret1967), Kanjilal and Singh (Reference Kanjilal and Singh1973), Kanjilal (Reference Kanjilal1985), Aberhan (Reference Aberhan1994, Reference Aberhan1998), Holzapfel (Reference Holzapfel1998), Delvene (Reference Delvene2001), Aberhan et al. (Reference Aberhan, Scholz and Schubert2011), Ros et al. (Reference Ros, Márquez-Aliaga and Damborenea2014), and Jaitly (Reference Jaitly2017).
Palaeoneilo aegyptiaca new species
Figures 14.28–14.46, 16.1–16.16, 17
Holotype
Articulated specimen in shell preservation from the middle–upper Bathonian Kehailia Formation, Gebel Homayir (BSPG 2014V 249/1) (Fig. 14.28–14.30).
Paratypes
Twenty-nine specimens, mostly disarticulated, in shell preservation: 23 specimens from the lower–upper Bajocian Bir Maghara Formation, Gebel Arousiah, Sinai (BSPG 2014V 143/1; 144/1–22), two specimens from the lower–upper Callovian Arousiah Formation, Gebel Mowerib (BSPG 2014V 136/1–2), and four specimens from the middle–upper Bathonian Kehailia Formation, Gebel Homayir (BSPG 2014V 241/1; 249/2–4).
Diagnosis
Small Palaeoneilinae, variable in shape and outline, ranging from subtrapezoidal to subpentagonal, from elongated-ovate to subtriangular, strongly inequilateral, moderately to strongly inflated dorsally, compressed posteriorly, with an angulated anterior margin, a subtruncated posterior margin, a straight to slightly convex ventral margin, no posterior umbonal ridge or escutcheon, a shallow elongated-ovate lunule, sharply pointed and slightly prosogyrate beaks, a wide hinge with continuous series of gradidentate anterior and posterior teeth without resilifer, forming an obtuse angle of ~130–145°; the smallest posterior teeth occasionally overlapping the anterior row of teeth below umbo; shell surface smooth.
Occurrence
Lower Bajocian to upper Bathonian, Gebel Maghara.
Description
Shell small, variable in outline ranging from subtrapezoidal to subpentagonal (distinctly longer than high), from elongated-ovate to subtriangular (Fig. 17), longer than high (H/L ratio ~0.60, on average; Table 7), inequilateral, equivalved, moderately to strongly inflated. Inflation greatest approximately one-third of total shell height from umbo, curvature flattening to slightly concave close to posterior margin, but no radial sulcus. Posterodorsal margin straight with narrow groove extending up to 3 mm from umbo towards posterior margin and referring to position of external amphidetic ligament (Fig. 14.46). Anterodorsal margin straight to slightly convex, forming a nearly right angle with anterior margin. Anterior margin narrowly angulated (Figs. 14.41, 16.2, 16.14, 17.1), meeting ventral margin in rounded curve. Posterior margin narrow, slightly convex to subtruncated and occasionally pointed. Ventral margin slightly convex to straight, occasionally sinuate close to posteroventral margin (Fig. 14.34). Umbones broad, little inflated, situated from one-third to one-fourth of total valve length from anterior end. Beaks sharply pointed, small, orthogyrate to slightly prosogyrate. No escutcheon or posterior umbonal ridge. Lunule shallow, narrow, elongated-ovate, deeply sunken towards beaks (Fig. 16.7, 16.8, 16.13). Posterior adductor muscle scar small, rounded. Hinge taxodont, gradidentate, arranged in two rows meeting below beak without resilifer (Fig. 14.46). Posterior hinge with 21–30 straight to slightly convexodont teeth, decreasing in size gradually towards umbo. Anterior hinge broad with 6–10 slightly convexodont to concavo-convexodont teeth, forming an obtuse angle of ~130–145° with posterior hinge. Rows of teeth uninterrupted below umbo (Fig. 16.3, 16.4). Smallest posterior teeth occasionally extending above anterior row of teeth below umbo (Fig. 14.36, 14.43, 14.46). Shell surface smooth except for faint commarginal growth lines close to ventral margin.

Figure 16. (1–16) Variation in outline of Palaeoneilo aegyptiaca n. sp. paratypes from the Middle Jurassic of Gebel Maghara. (1) BSPG 2014V 136/1, Arousiah Formation, Gebel Mowerib, left valve, elongated-ovate form with oyster encrustations; (2, 3) BSPG 2014V 144/8, Bir Maghara Formation, Gebel Arousiah, left lateral view showing angulated anterior margin and left valve interior, respectively; (4) BSPG 2014V 249/2, Kehailia Formation, Gebel Homayir, right valve interior showing a continuous series of gradidentate anterior and posterior teeth without resilifer; (5–16) Bir Maghara Formation, Gebel Arousiah; (5, 6) BSPG 2014V 144/9, left lateral view of elongated-ovate form with straight ventral margin and right lateral view, respectively, (7) BSPG 2014V 144/10, dorsal view, (8) BSPG 2014V 144/11, dorsal view, (9–11) BSPG 2014V 144/12, right lateral view, right valve, and left valve, respectively, of less elongated-ovate form of species, (12–14) BSPG 2014V 144/13, right valve, dorsal view with narrow lunule, and left lateral view of sub-pentagonal form of species, (15) BSPG 2014V 144/14, left lateral view, (16) BSPG 2014V 144/15, left lateral view of sub-triangular form of species. (17–30) Variation in outline of Palaeoneilo muensteri (Goldfuss, 1841) from the Lower Jurassic (Toarcian) Rajabiah Formation, western Bir Maghara. (17, 18) BSPG 2014V 338/1, right lateral view and left valve respectively, (19–21) BSPG 2014V 338/2, right valve, (20) dorsal view showing the deep lunule, and left valve, respectively, (22–24) BSPG 2014V 338/3, right lateral view, left valve, and dorsal view, respectively, (25) BSPG 2014V 338/4, dorsal view showing traces of taxodont hinge (arrowed), (26) BSPG 2014V 338/5, close-up showing the poorly preserved convexodont teeth (arrowed), (27–29) right valve, dorsal view, and left valve, respectively, (30) BSPG 2014V 338/7, right valve. (31–38) Dacryomya diana (d'Orbigny, 1850) from the middle‒late Bathonian Kehailia Formation, Gebel Mowerib. (31–34) BSPG 2014V240/1; (31) left lateral view showing narrow posterior end with deep sulcus, (32) interior view of left valve, (33) close-up showing few slightly convexodont teeth and small, sub-rounded anterior and posterior adductor muscle scars, (34) dorsal view showing a well-developed posterior umbonal ridge with deep sulcus (arrowed), (35–38) BSPG 2014V240/2, left valve, dorsal view, right valve, and posterodorsal view, respectively. (39–43) Dacryomya lacryma (J. de C. Sowerby, 1824) from the Lower‒Middle Jurassic of Gebel Maghara. (39, 40) BSPG 2014V 339/1, Bir Maghara Formation, Gebel Arousiah, left lateral and dorsal views, respectively, (41–43) BSPG 2014V BSPG 2014V 340/1, Kehailia Formation, Gebel Mowerib, (41) left valve, (42) left valve interior with traces of taxodont hinge, (43) dorsal view. Scale bars = 5 mm.

Figure 17. (1–12) Variation in outline of Palaeoneilo aegyptiaca n. sp. from the Middle Jurassic of Gebel Maghara. Specimens from several populations collected from the studied sections. Scale bars = 5 mm.
Table 7. Measurements (in mm) of Palaeoneilo aegyptiaca n. sp.

Etymology
After the type area of the species (Egypt).
Measurements
See Table 7.
Remarks
Palaeoneilo aegyptiaca n. sp. is a very variable species with respect to shape and shell outline (Fig. 17). It can be distinguished from other Jurassic Palaeoneilo species by having a smooth surface, a subtruncated posterior margin, an angulated anterior margin, a sharply pointed and slightly prosogyrate beak, a narrow elongated-ovate lunule (more sunken towards beaks), and by lacking an escutcheon and posterior umbonal ridge. The closest species appears to be Palaeoneilo galatea (d'Orbigny, 1850), which has been recorded from Lower Jurassic strata of central and western Europe. Palaeoneilo galatea differs, however, in having broad and depressed umbones, a rounded anterior margin, and a posterior umbonal ridge forming a flattened to weakly concave area between it and the posterodorsal margin (e.g., Aberhan et al., Reference Aberhan, Scholz and Schubert2011, pl. 1, figs. 3, 4; Karapunar et al., Reference Karapunar, Werner, Fürsich and Nützel2020, pl. 2, figs. 4, 6). Moreover, it is less elongated and less inflated (H/L = 0.58, Iav/L = 0.33, respectively; Dumortier, Reference Dumortier1869). In addition, the posterior and anterior teeth of P. galatea are straight to slightly concave, and meet in a more obtuse angle than P. aegyptiaca n. sp. (e.g., 156°, Damborenea, Reference Damborenea1987, as opposed to 130–145°). Palaeoneilo aegyptiaca n. sp. also shares some similarities with Leda trapezoidalis Monke, Reference Monke1888 (p. 216, pl. 2.3, figs. 8–8a) from the Lower Jurassic of Germany, which has been regarded as a synonym of P. galatea by Hodges (Reference Hodges2000, p. 28) and Aberhan et al. (Reference Aberhan, Scholz and Schubert2011, p. 69). Leda alpina d'Orbigny, 1850 (p. 336, no. 136) from the Callovian of France figured by Cottreau (Reference Cottreau1925, p. 143, pl. 17 [38], fig. 3) differs in having a narrow and strongly convex posterior margin, a regularly rounded ventral margin, a concave posterodorsal margin, a straight anterodorsal margin, compressed umbones, and is more elongated than the present species (H/L = 0.46 as opposed to 0.60 on average; Table 7), and is much more inflated dorsally.
Mesosaccella morrisi (Deshayes, 1853) of Duff (Reference Duff1978, p. 28, pl. 1, figs. 22, 23–33, text-fig. 7) and Fürsich and Pan (Reference Fürsich and Pan2014, p. 6, pl. 1, figs. 1–3) from the Callovian–Oxfordian of England and Iran, respectively, resemble P. aegyptiaca n. sp. in having an angulated anterior margin and an elongated and inflated valve, but differ in having submesial umbones, well-developed commarginal ribs, small resilifers, regularly rounded ventral margins, and less-prominent beaks. Palaeoneilo? patagonidica (Leanza, Reference Leanza1942) figured and described by Damborenea and Pagani (Reference Damborenea and Pagani2019, p. 939, figs. 6.10–16, 7.6) from the Lower Jurassic of Argentina has strongly rounded shell margins, less-prominent beaks, less-inflated valves, and a higher number of anterior teeth (up to 13 as opposed to 6–10 in P. aegyptiaca n. sp.). Leda palmae (J. de C. Sowerby, Reference Sowerby1825) figured by Dumortier (Reference Dumortier1869, p. 120, pl. 19, figs. 3, 4) from the Lower Jurassic of France differs from the present species by subcentral umbones and rounded margins. Cox (Reference Cox1965, p. 25, pl. 1, fig. 1) erected P. asaharbitensis from Bathonian–?Callovian beds of Kenya based on a single internal mold. The latter species probably falls within the range of variation of known Jurassic Palaeoneilo species. However, P. asaharbitensis differs from P. aegyptiaca n. sp. in having a strongly rounded anterior margin, and its posterodorsal margin is straight, oblique, and forms an acute angle with the posterior margin. Palaeoneilo bornholmiensis (von Seebach, Reference von Seebach1865) figured by Troedsson (Reference Troedsson1951, p. 150, pl. 16, fig. 3, text-fig. 39) from the Lower Jurassic of Sweden resembles P. aegyptiaca n. sp. in having an elongated valve, anteriorly placed umbones, smooth shell surface, but differs in having a convex ventral margin, a rounded anterior margin, slightly concave antero- and posterodorsal margins, and in being smaller (L = 7.5, H = 4.0 mm) and more elongated than the present species. The latter diagnostic features clearly fall into the range of variation of Palaeoneilo elliptica (Goldfuss, 1837) and, therefore, has been regarded as a junior synonym of the latter by Hodges (Reference Hodges2000, p. 28). de Loriol (Reference de Loriol1899) described and figured Nucula longiuscula Mérian from the Oxfordian of Switzerland (de Loriol, Reference de Loriol1899, p. 159, pl. 10, figs. 23–25) (Mérian only used the species name on a museum label; de Loriol subsequently established the species with Mérian as author). Based on the hinge structure, his specimens likely belong to Palaeoneilo, but differ from P. aegyptiaca n. sp. in having a strongly concave anterodorsal margin, well-developed lunule and escutcheon, less prominent beaks, and in being more inflated.
Palaeoneilo muensteri (Goldfuss, 1841)
Figure 16.17–16.30
- .Reference Goldfuss1837
Nucula elliptica Goldfuss, p. 153, pl. 124, fig. 16a–e [non Nucula elliptica Phillips, 1829].
- *Reference Goldfuss1841
Nucula Münsteri Goldfuss, p. 304. [nom. nov. for Nucula elliptica Goldfuss, 1837].
- .Reference Quenstedt1852
Nucula tunicata Quenstedt, p. 529, pl. 44, fig. 9.
- .Reference Monke1888
Leda trapezoidalis Monke, p. 216, pl. 2.3, figs. 8, 8a.
- p.Reference Kuhn1936
Leda galathea; Kuhn, p. 259, pl. 9, fig. 9a, b; non fig. 5a–c [ = Palaeoneilo galatea (d'Orbigny, 1850)].
- p.Reference Cox1937
Palaeoneilo galatea; Cox, p. 191, pl. 5, figs 5–7, non fig. 4.
- .Reference Troedsson1951
Palaeoneilo oviformis Troedsson, p. 151, pl. 16, figs. 7, 8, 12, 17.
- .Reference Troedsson1951
Palaeoneilo galatea; Troedsson, p. 149, pl. 16, figs. 1, 2, 6, (9–11?).
- .Reference Troedsson1951
Palaeoneilo bornholmiensis; Troedsson, p. 150, pl. 16, fig. 3.
- .Reference Carter and Carter1990
Palaeoneilo elliptica; Carter, p. 159, fig. 12A–E.
- .Reference Hodges2000
Palaeoneilo elliptica; Hodges, p. 28, pl. 2, figs. 1–30, text-figs. 25–33 (with extensive synonymy).
- non .Reference Scholz, Aberhan and González-León2008
Palaeoneilo elliptica; Scholz et al., p. 274, fig. 4A, B.
- .Reference Aberhan, Scholz and Schubert2011
Palaeoneilo elliptica; Aberhan et al., p. 68, pl. 1, figs. 1, 2.
- .Reference Karapunar, Werner, Fürsich and Nützel2020
Palaeoneilo muensteri; Karapunar et al. p. 11, pl. 2, figs. 1–3, pl. 10, fig. 9, text-figs. 4–6.
Holotype
Nucula elliptica Goldfuss, 1837, pl. 124, fig. 16a–e from the Lower Jurassic (Pliensbachian) of Germany.
Occurrence
Lower Jurassic of Germany (Pliensbachian), southwestern England (Sinemurian), France, Upper Triassic of Italy, New Zealand, South America, and from Lower Jurassic (Toarcian) of Sinai (present study, first record).
Description
Shell small, outline subelliptical, inequilateral, longer than high (H/L = 0.63 on average; Table 8), equivalved, and moderately inflated. Maximum inflation at about anterior third of total valve length. Posterior area approximately twice as long as anterior one. Posterodorsal margin straight, slightly convex, meeting posterior margin in rounded angle. Anterodorsal margin short, slightly concave, forming a blunt angle with anterior margin. Posterior margin rounded, narrow, occasionally sharply rounded, meeting ventral margin in continuous rounded curve. Ventral margin gently convex. Umbo low, moderately inflated, located about one-third of total valve length from anterior end. Beak hardly prominent and slightly prosogyrate. Hinge teeth not well preserved, but clearly taxodont (Fig. 16.25, 16.26). Posterior teeth partially well preserved, convexodont, and gradually decreasing in size towards beak (Fig. 16.26). No escutcheon. Lunule well defined, cordate, moderately deep. Shell surface smooth except for fine commarginal ribs close to ventral margin (Fig. 16.17, 16.18).
Table 8. Measurements (in mm) of Palaeoneilo muensteri (Goldfuss, 1841).

Materials
Ten composite molds, occasionally in shell preservation (BSPG 2014V 338/1–10), from the Lower Jurassic (Toarcian) Rajabiah Formation, western Bir Maghara (BSPG 2014V 317/1–10), Gebel Maghara.
Measurements
See Table 8.
Remarks
Hodges (Reference Hodges2000) carefully examined Palaeoneilo muensteri [as Palaeoneilo elliptica (Goldfuss, 1837)] and analyzed a large number of specimens from the Lower Jurassic (Lower Lias) of England. He noted that the species is highly variable with respect to height/length ratio, shell inflation, and length of anterior area (see biometric analyses of Hodges, Reference Hodges2000, p. 29–31, text-figs. 25–30, and shell forms, pl. 2, figs. 1–30). The present material clearly falls into the range of variation given by Hodges (Reference Hodges2000). Karapunar et al. (Reference Karapunar, Werner, Fürsich and Nützel2020) discussed the problematic aspects of the species name, which was erected by Goldfuss (1837) as Nucula elliptica from the Lias of northern Germany. After a detailed discussion, they corrected the name of the species to Palaeoneilo muensteri—a correction that is accepted here (see Karapunar et al., Reference Karapunar, Werner, Fürsich and Nützel2020, p. 12–13 for details). Palaeoneilo muensteri can be distinguished from other Lower Jurassic Palaeoneilo species by having an elliptical to subelliptical valve, a narrowly rounded posterior margin (occasionally angulated), a faintly convex ventral margin, a slightly prominent, slightly prosogyrate beak, a deep lunule, and by lacking an umbonal posterior ridge. In addition, Hodges (Reference Hodges2000) noted that the flank of P. elliptica carries fine commarginal striae obliquely crossed by straight striae. He considered the latter ornament as a diagnostic feature of P. elliptica. Recently, Karapunar et al. (Reference Karapunar, Werner, Fürsich and Nützel2020) observed the same ornamentation in other Lower Jurassic Palaeoneilo species, such as P. galatea. It generally seems to be visible in well-preserved preserved specimens of the genus Palaeoneilo. The most closely related species is P. galatea (d'Orbigny, 1850), which has been recorded from the Lower Jurassic at different localities in Europe, South America, and China. According to Hodges (Reference Hodges2000), P. galatea can be distinguished from P. elliptica by having an angulate posterior margin. In addition, P. galatea has a well-developed posterior umbonal ridge and a flattened to feebly concave area between the umbonal ridge and posterodorsal margin (see Kuhn, Reference Kuhn1935, pl. 18, fig. 30a–c; Aberhan et al., Reference Aberhan, Scholz and Schubert2011, pl. 1, figs. 3, 4; Karapunar et al., Reference Karapunar, Werner, Fürsich and Nützel2020, pl. 2, figs. 4–6).
The high variability of P. muensteri prompted some authors to erect new species such as Nucula tunicata Quenstedt, Reference Quenstedt1852, and Leda trapezoidalis Monke, Reference Monke1888, from Germany, and Palaeoneilo oviformis Troedsson, Reference Troedsson1951, from Sweden. Hodges (Reference Hodges2000) revised these latter species and noted that most of them lie well within the range of variation of P. muensteri (as P. elliptica)—a view that is followed here.
“Nucula” patagonidica from the Lower Jurassic of Argentina was erected by A. Leanza (Reference Leanza1942, p. 151, pl. 1, figs. 1, 2, 4, non fig. 3). The latter species strongly resembles P. muensteri in having an elongated shell, little prominent beaks, and low umbones, but differs in having strongly rounded shell margins. In addition, the hinge plate of “N.” patagonidica is gently convex throughout without posterodorsal angle and its external surface is less inflated with faint commarginal growth lines that become stronger ventrally (Damborenea, Reference Damborenea1987, p. 54–55; Damborenea and Pagani, Reference Damborenea and Pagani2019, p. 939, fig. 6.10–6.16). Hodges (Reference Hodges2000, p. 28) and Scholz et al. (Reference Scholz, Aberhan and González-León2008, p. 274) regarded P. patagonidica (Leanza, Reference Leanza1942) as a junior synonym of P. elliptica (Goldfuss), arguing that some transitional forms exist between the two species. Damborenea and Pagani (Reference Damborenea and Pagani2019) did not accept the latter view and kept it as a separate species. Palaeoneilo aegyptiaca n. sp. from the early–late Bajocian Bir Maghara Formation (described above) has an elongated subtrapezoidal valve, a smooth surface, an angulated anterior margin, a sharply pointed orthogyrate to slightly prosogyrate beak, and no lunule. With respect to general outline, height/length ratio, and ornamentation, Leda alpina d'Orbigny (1850, p. 336, no. 136) from the Callovian of France figured by Cottreau (Reference Cottreau1925, p. 143, pl. 17 [38], fig. 3) resembles the present species, but differs in having a straight anterodorsal margin (forming an obtuse angle with the posterodorsal margin), a strongly rounded posterior margin, a strongly concave posterodorsal margin, and more-inflated umbones. For more details concerning P. muensteri and other closely related taxa, see Hodges (Reference Hodges2000, p. 34), Aberhan et al. (Reference Aberhan, Scholz and Schubert2011, p. 68–69), and Karapunar et al. (Reference Karapunar, Werner, Fürsich and Nützel2020, p. 12–13).
Superfamily Nuculanoidea Adams and Adams, Reference Adams and Adams1858
Family Polidevciidae Kumpera, Prantl, and Růžička, Reference Kumpera, Prantl and Růžička1960
Genus Dacryomya Agassiz, Reference Agassiz1840
Type species
Nucula lacryma J. de C. Sowerby, Reference Sowerby1824, from the “Great Oolite” (Bathonian) of Ancliff, southern England; subsequent designation by Herrmannsen (Reference Herrmannsen1846, p. 368).
Remarks
According to Ros et al. (Reference Ros, Márquez-Aliaga and Damborenea2014, p. 20), there is no consensus about the family affiliation of the genus Dacryomya. It was assigned to Nuculanidae (e.g., Cox et al., Reference Cox, Newell, Branson, Casey, Chavan, Coogan, Dechaseaux, Moore and Teichert1969; Hayami, Reference Hayami1975), Nuculidae (e.g., Hodges, Reference Hodges2000; Karapunar et al., Reference Karapunar, Werner, Fürsich and Nützel2020) or to Polidevciidae (Carter, Reference Carter and Carter1990; Jaitly et al., Reference Jaitly, Fürsich and Heinze1995; Delvene, Reference Delvene2001). The Ordovician–Upper Jurassic subfamily Polidevciinae was erected by Kumpera et al. (Reference Kumpera, Prantl and Růžička1960, p. 33–34) during their revision of the Nuculanidae from the Ostrava-Karviná District based on a strong posterior umbonal ridge, a short rostrum, and an internal ligament. Later, Carter (Reference Carter and Carter1990) raised the subfamily Polidevciinae to family rank (Polidevciidae Kumpera et al., Reference Kumpera, Prantl and Růžička1960) based on ligament structure and shell microstructure. Recently, Carter et al. (Reference Carter, Altaba, Anderson, Campbell, Fang, Harries and Skelton2015, p. 3) added other diagnostic features for this family, such as (1) nacreous, not porcelaneous shells; (2) a shallow to deep submarginal resilium; (3) an external weakly mineralized ligament; and (4) paleotaxodont hinge teeth.
In agreement with Ros et al. (Reference Ros, Márquez-Aliaga and Damborenea2014, p. 20), Dacryomya is provisionally assigned herein to the family Polidevciidae until a larger data base is available to investigate carefully the ligamental structure. For more details concerning the diagnostic features of Polidevciidae and comparison with closely related families, see Ros et al. (Reference Ros, Márquez-Aliaga and Damborenea2014) and Carter et al. (Reference Carter, Altaba, Anderson, Campbell, Fang, Harries and Skelton2015, p. 3, 6). Apart from ligament structure, the genera that belong to Polidevciidae can be distinguished from other nuculanid genera such as Nuculana and Ryderia by having a relatively short rostrum, strong posterior umbonal ridge (folded in cross-section), deep umbonal posterior sulcus, well-developed shallow escutcheon (without distinct ridge), a smaller number of anterior and posterior teeth, and a greater inflation. According to Ros et al. (Reference Ros, Márquez-Aliaga and Damborenea2014, p. 20, fig. 4), Dacryomya has a long stratigraphic range (from Norian to Kimmeridgian) and probably also occurs in the Lower Triassic. It has been recorded from Upper Triassic–Lower Jurassic rocks of England and Lower Jurassic rocks of Portugal, Germany, Switzerland, France, China, Japan, Siberia, and the Arctic region. The youngest record of the genus is from the Upper Oxfordian Sot de Chera Formation of Spain (Delvene, Reference Delvene2001). Mongin (Reference Mongin1967, p. 41, pl. 1, figs. 15, 16) recorded Nuculana (Dacryomya) cf. D. lacryma (Sowerby) from the Bathonian of Morocco, and Holzapfel (Reference Holzapfel1998, p. 95, pl. 3, fig. 5a, b) recorded Dacryomya acuta Mérian (of de Loriol, Reference de Loriol1899, p. 164, pl. 10, figs. 29–32) from the Callovian–Oxfordian of southern Tunisia. Because Holzapfel's specimens are internal molds, her placement of the specimen in Dacryomya is somewhat doubtful. Based on Figure 13, Holzapfel's specimen is much closer to Palaeonucula than Dacryomya. In the present study, Dacryomya is recorded from the Lower–Middle Jurassic of Egypt for the first time (Fig. 15).
Dacryomya diana (d'Orbigny, Reference d'Orbigny1850)
Figure 16.31–16.38
- .Reference Goldfuss1837
Nucula mucronata; Goldfuss, p. 155, pl. 125, fig. 9a–d (non Sowerby, Reference Sowerby1825).
- *Reference d'Orbigny1850
Leda Diana d'Orbigny, p. 253.
- p.Reference Quenstedt1856
Nucula claviformis; Quenstedt, p. 313, pl. 43, fig. 4, non figs. 5, 6.
- .Reference Oppel1856
Leda Diana; Oppel, p. 398.
- .Reference Dumortier1874
Leda diana; Dumortier, p. 298, pl. 6, figs. 14, 15.
- p.Reference Quenstedt1884
Nucula claviformis; Quenstedt, p. 804, pl. 63, fig. 22, non fig. 23.
- .Reference Borissjak1904
Leda diana; Borissjak, p. 42, pl. 3, fig. 1-b.
- .Reference Frebold1929
Leda cf. diana; Frebold, p. 265, pl. 3 (35), fig. 7.
- .Reference Klöcker1966
Nuculana diana; Klöcker, p. 222, fig. 2.
- .Reference Etter1990
Nuculana diana; Etter, pl. 3, fig. 8 (right valve).
Holotype
Nucula mucronata Goldfuss, 1837, pl. 125, fig. 9, from “E stratis Lias dictis Franconiae et Württembergiae” (Lower Jurassic of southern Germany).
Occurrence
Lower–Middle Jurassic strata of southwestern Spitsbergen, France, Germany, Russia, and from the Middle Jurassic of Sinai (present study, first record).
Description
Small nuculaniform shell, subtriangular in outline, inequilateral, equivalved, longer than high (H/L = 0.73; Table 9), with short posterior rostrum, and moderate inflation. Anterior margin strongly convex, meeting ventral margin in rounded curve. Posterodorsal margin slightly concave. Anterodorsal margin slightly convex. Posterior end of rostrum narrow, short, and angular. Umbo inflated, triangular, and located slightly anterior of the mid-length of valve. Beak sharply pointed and slightly opisthogyrate. Posterior umbonal ridge blunt, extending from umbo to posteroventral corner, and separating strongly concave posterodorsal flank from the rest of valve (Fig. 16.34). Deep sulcus anterior of the posterior umbonal ridge (max. width: 2.2 mm) widening posteroventrally (Fig. 16.31, 16.35). Escutcheon shallow, elongated, lanceolate, and ornamented with faint riblets. Lunule narrow and shallowly depressed. Anterior and posterior adductor muscle scars subrounded, isomyarian, strongly impressed, and located close to anterior and posterodorsal margin, respectively (Fig. 16.32). Pallial line distinct, entire. Inner shell margin smooth. Hinge plate wide, with rows of slightly convexodont teeth (Fig. 16.33). Shell surface smooth except for well-developed commarginal ribs close to ventral margin.
Table 9. Measurements (in mm) of Dacryomya diana (d'Orbigny, 1850).

Materials
Three specimens, one in shell preservation and two composite molds (BSPG 2014V 240/1–3) from the middle–late Bathonian Kehailia Formation of Gebel Mowerib, Sinai.
Measurements
See Table 9.
Remarks
Dacryomya diana (d'Orbigny, 1850) can be distinguished from other Jurassic nuculanids by having a triangular valve, strongly rounded anterior and ventral margins, a well-defined and ornamented escutcheon, a slightly rostrate and narrow posterior end, and a well-developed posterior umbonal ridge with a deep sulcus located anterior of it (Fig. 16.34, 16.35). d'Orbigny (1850) had regarded Nucula mucronata as figured and described by Goldfuss (1837, p. 156, pl. 126, figs. 9a–d; non J. Sowerby, Reference Sowerby1825) from the Lower Jurassic of Germany as the holotype of his new species Leda diana from the Lower Jurassic (Toarcian) of France [lectotype and lectostratotype: Goldfuss’ material, E stratis Lias dictis Franconiae et Württembergiae, Germany]. Nucula mucronata J. de. C. Sowerby (Reference Sowerby1825, p. 120, pl. 476, fig. 4) from the Bajocian of England has a sub-rhombohedral valve, a narrower rostrum, a strongly convex ventral margin (sinuate posteriorly), well-developed commarginal ribs between posterior umbonal ridge and anterior margin, and is less elongated and less inflated than the present material. Quenstedt (Reference Quenstedt1884) suggested that D. diana is a juvenile form of N. rostralis (Lamarck, Reference Lamarck1819). As long as no intermediate forms are recorded from Jurassic strata, the suggestion of Quenstedt is not acceptable. In addition, N. rostralis has a much longer rostrum than D. diana, well-developed ridges bordering the deep escutcheon (see Goldfuss, 1837, pl. 125, fig. 8c), and an elongated shallow lanceolate lunule. Klöcker (Reference Klöcker1966, p. 220) regarded Nucula claviformis J. de C. Sowerby, Reference Sowerby1825 (p. 119, pl. 476, fig. 2) from the Upper Jurassic of England as a synonym of N. rostralis. Although the two species are very similar, N. claviformis has a shorter rostrum, a deeper posterior sulcus, lacks a lunule, and its posterior umbonal ridge is less developed and concave posterodorsally (straight in N. rostralis). Another very similar species is Leda medusa Borissjak, Reference Borissjak1904, from the Callovian–Oxfordian of Russia (Borissjak, Reference Borissjak1904, p. 43, pl. 3, figs. 2a–e, 3a–e, 4a–e). The latter species can be distinguished by its large and thick valve, smooth escutcheon and lunule, a strongly inflated umbo with well-developed beak, and irregular commarginal growth lines. Nuculana (Dacryomya) lacryma (J. de. C. Sowerby, Reference Sowerby1825) figured by Cox (Reference Cox1940, p. 29, pl. 2, figs. 2–5) and Jaitly et al. (Reference Jaitly, Fürsich and Heinze1995, p. 161, pl. 2, figs. 21–23) from the Middle Jurassic of western India resembles the present species in having a well-developed posterior umbonal ridge, a posterior sulcus, and fine commarginal ribs, but differs in having a more rostrate posterior elongation, a deep escutcheon (delimited by a blunt ridge; e.g., Cox, Reference Cox1940, pl. 2, fig. 2b; Jaitly et al., Reference Jaitly, Fürsich and Heinze1995, pl. 2, figs. 21b, 23b), and a strongly convex anterodorsal margin.
Dacryomya lacryma (J. de C. Sowerby, Reference Sowerby1824)
Figures 16.39–16.43, 18.1–18.7
- *Reference Sowerby1824
Nucula lacryma J. de. C. Sowerby, p. 119, pl. 476, fig. 3.
- .Reference Quenstedt1857
Nucula lacryma; Quenstedt, p. 505, pl. 67, figs. 18–21.
- .Reference Ogérien1867
Nucula lacryma; Ogérien, p. 775, fig. 426.
- .Reference Greppin1899
Leda lacryma; Greppin, p. 97, pl. 9, figs. 1, 1a.
- .Reference Cossmann1912
Leda lacryma; Cossmann, p. 7, pl. 3, figs. 11–13.
- .Reference Marzloff, Dareste De La Chavanne and Moret1936
Leda lacryma; Marzloff et al., p. 108, pl. 12, figs. 9, 10a, b.
- .Reference Kuhn1938
Leda lacryma; Kuhn, p. 132, pl. 5, fig. 6.
- .Reference Cox1940
Nuculana (Dacryomya) lacryma; Cox, p. 29, pl. 2, figs. 2–5.
- .Reference Agrawal1956
Nuculana (Dacryomya) lacryma; Agrawal, p. 55, pl. 7, fig. 3c.
- ?Reference Mongin1967
Nuculana (Dacryomya) cf. lacryma; Mongin, p. 41, pl. 1, figs. 15, 16.
- .Reference Romanov1973
Leda lacryma; Romanov, p. 35, pl. 2, figs. 8–12.
- .Reference Jaitly, Fürsich and Heinze1995
Dacryomya lacryma; Jaitly et al., p. 161, pl. 2, figs. 21–23, text-fig. 13.
- vReference Ali1989
Nuculana rostrata Ali, p. 89, pl. 24, figs. 1–7. [Homonym of the type species of Nuculana, viz. N. rostrata (Gmelin, Reference Gmelin1791)]
- vReference Abdelhamid2002
Nuculana rostrata; Abdelhamid, pl. 5, figs. 7, 8. [= Dacryomya lacryma (J. de C. Sowerby, Reference Sowerby1824)]
- vReference Khalil2003
Cuspidaria sp., Khalil, pl. 1, figs. 18, 19. [= D. lacryma (J. de C. Sowerby, Reference Sowerby1824)]
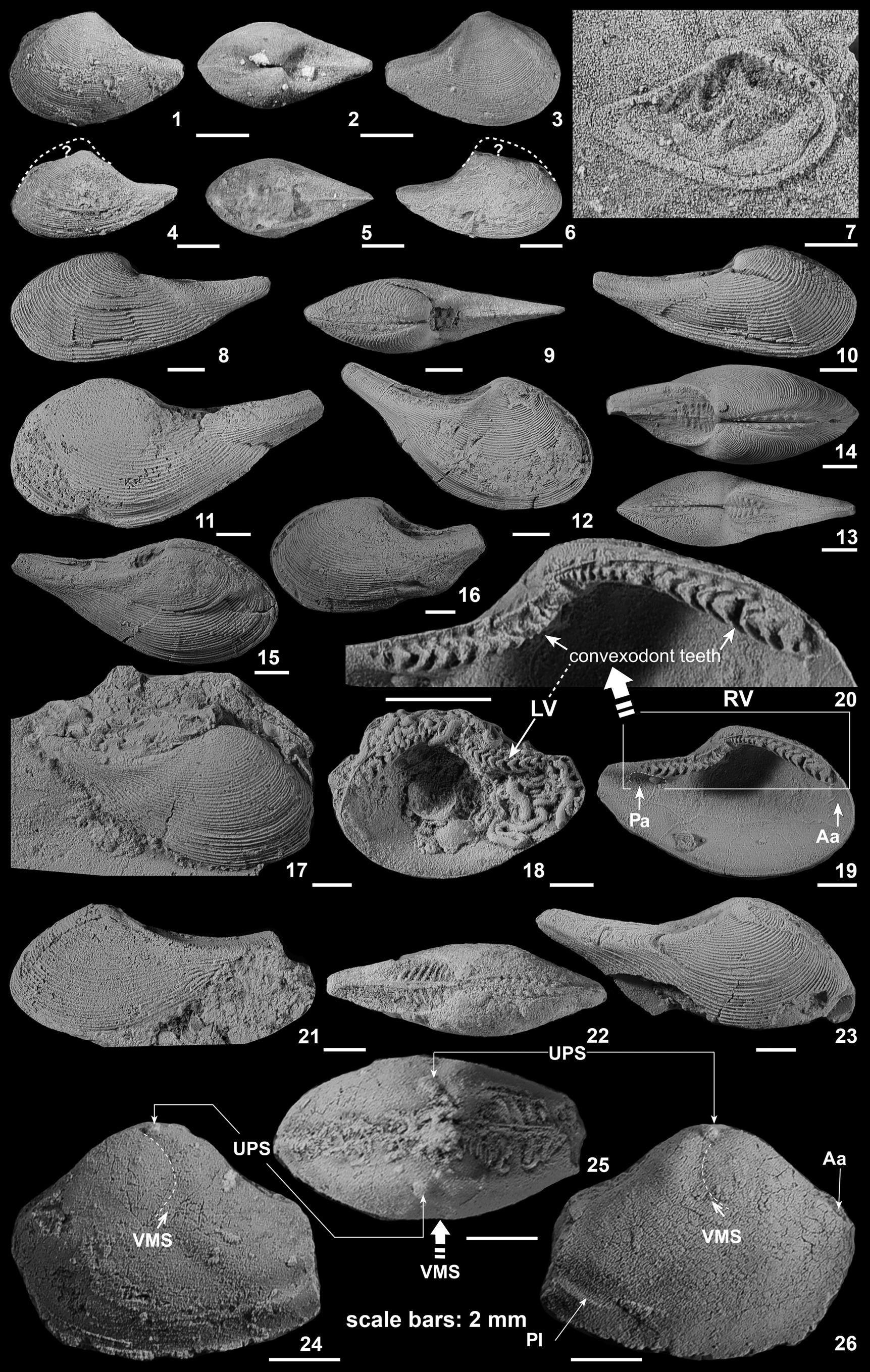
Figure 18. (1–7) Dacryomya lacryma (J. de C. Sowerby, Reference Sowerby1824) from the Lower‒Middle Jurassic of Gebel Maghara. (1–6) Shusha Formation, western Bir Maghara; (1–3) BSPG 2014V 417/1, left valve, dorsal view showing deep escutcheon, and right lateral view, respectively, (4–6) BSPG 2014V 417/2, left lateral, dorsal, and right lateral views, respectively, (7) BSPG 2014V 245/1, Kehailia Formation, Gebel Arousiah, left valve interior showing the gradidentate convexodont teeth. (8–26) Ryderia decorata (Douvillé, Reference Douvillé1916) from the Jurassic (Toarcian–lower Kimmeridgian) rocks of Gebel Maghara; (8–10) BSPG 2014V 416/1, Masajid Formation, western Bir Maghara, left lateral, dorsal, and right lateral views, respectively, (11–14) Kehailia Formation, Gebel Homayir; (11) BSPG 2014V 243/1, left lateral view, (12) BSPG 2014V 243/2, right lateral view, (13) BSPG 2014V 243/3, dorsal view; (14) BSPG 2014V 342/1, dorsal view showing a wide, cordate-shaped, ornamented escutcheon; (15) BSPG 2014V 343/1, Kehailia Formation, Gebel Mowerib, right lateral view; (16) BSPG 2014V 345/1, Masajid Formation, western Bir Maghara, left lateral view; (17, 18) Kehailia Formation, Gebel Engabashi; (17) BSPG 2014V 3/1, right lateral view, (18) BSPG 2014V 3/2, interior of right valve showing the convexodont teeth and sockets with serpulid encrustations; (19, 20) BSPG 2014V 342/2, Kehailia Formation, Gebel Homayir; (19) left valve interior showing anterior (Aa) and posterior muscle scars (Pa), (20) close-up showing slightly convex anterior hinge with convexodont teeth and slightly concave posterior hinge; (21) BSPG 2014V 341/1, Kehailia Formation, Gebel Homayir, left lateral view; (22) BSPG 2014V 416/2, Masajid Formation, western Bir Maghara, dorsal view; (23) BSPG 2014V 342/3, Kehailia Formation, Gebel Homayir, right valve; (24–26) BSPG 2014V 242/1, Kehailia Formation, Gebel Homayir; (24) left valve, internal mold showing well-developed visceral muscle scar (VMS) and umbonal pedal muscle scars (UPS), (25) dorsal view showing the well-preserved (UPS), (26) right lateral view showing anterior muscle scar (Aa), part of the pallial line (Pl), (UPS), and VMS (arrowed). Scale bars = 2 mm.
Holotype
Nucula lacryma J. de. C. Sowerby, Reference Sowerby1824, pl. 476, fig. 3, from the “Great Oolite” (Bathonian) of Ancliff, southern England.
Occurrence
Bajocian–Bathonian of England, Germany, France, USSR, Bajocian of Switzerland, upper Bajocian–lower Kimmeridgian of western India, ?Bathonian of Morocco, and Bajocian–Callovian of Gebel Maghara (first record).
Description
Small nuculaniform shell, elongated-ovate, very inequilateral, longer than high (H/L: 0.62–?0.67; Table 10), moderately inflated, slightly rostrate. Anterior margin strongly convex, meeting ventral margin in rounded curve. Posterodorsal margin slightly concave. Ventral margin moderately convex anteriorly, becoming straight towards posterior end. Umbo inflated, triangular, and located slightly anterior of mid-length of valve. Beak prominent, slightly opisthogyrate, elevated above dorsal margin. Escutcheon well impressed, smooth, and bordered by a well-defined rounded ridge. Hinge not well preserved, gradidentate with convexodont teeth (Figs. 16.42, 18.7). Anterior and posterior rows of teeth decreasing in size towards umbo. Ornamentation consisting of fine irregular commarginal growth lines.
Table 10. Measurements (in mm) of Dacryomya lacryma (J. de C. Sowerby, Reference Sowerby1824).

Materials
Eight specimens, in shell preservation, from upper Toarcian to upper Bathonian strata of Gebel Maghara; four specimens from the upper Toarcian Shusha Formation (BSPG 2014V 417/1–4), western Bir Maghara; two incomplete specimens from the upper Bajocian Bir Maghara Formation, Gebel Arousiah (BSPG 2014V 339/1–2); a single specimen from the middle–upper Bathonian Kehailia Formation, Gebel Mowerib (BSPG 2014V 340/1); and a single specimen from the same formation of Gebel Arousiah (BSPG 2014V 245/1).
Measurements
See Table 10.
Remarks
Ali (Reference Ali1989) erected Nuculana rostrata from the Lower Bathonian Safa Formation of Gebel Maghara. Later on, Abdelhamid (Reference Abdelhamid2002) recorded that species from the same area, but from a younger stratigraphic level (upper Bathonian–Callovian Kehailia member, Masajid Formation). According to the ICZN (1999), Ali's N. rostrata is invalid, because the species name is preoccupied by the type species of genus Nuculana—N. rostrata (Gmelin, Reference Gmelin1791). Moreover with respect to shell outline and size and length of rostrum, the Egyptian material is identical to Dacryomya lacryma (J. de C. Sowerby, Reference Sowerby1824; Fig. 19.1–19.3, 19.9) as figured and described by other authors (e.g., J. de C. Sowerby, Reference Sowerby1824, pl. 476, fig. 3, from the Middle Jurassic Great Oolite of England; Quenstedt, 1857, pl. 67, figs. 18–21, from the Middle Jurassic of Germany; Cox, Reference Cox1940, pl. 2, figs. 2–5, from the Middle Jurassic [Bathonian–Oxfordian] of western India; and Jaitly et al., Reference Jaitly, Fürsich and Heinze1995, pl. 2, figs. 21–23, from the Middle Jurassic [Bathonian–Oxfordian] of western India). The specimens figured by Khalil (Reference Khalil2003, pl. 1, figs. 18, 19) as Cuspidaria sp. from the upper Bathonian rocks of Gebel Maghara show a taxodont hinge, a shallow posterior umbonal sulcus, a narrow rostrate posterior end, and a strongly convex anterodorsal and anterior margin (Fig. 19.5–19.7). These features closely fit Dacryomya and there is no doubt that the specimens of Khalil (Reference Khalil2003) clearly fall into the range of variation of D. lacryma (J. de C. Sowerby, Reference Sowerby1824) as documented by Cox (Reference Cox1940) and Jaitly et al. (Reference Jaitly, Fürsich and Heinze1995). Therefore, the specimens of Ali (Reference Ali1989), Abdelhamid (Reference Abdelhamid2002), and Khalil (Reference Khalil2003) have been included as junior synonyms of D. lacryma.
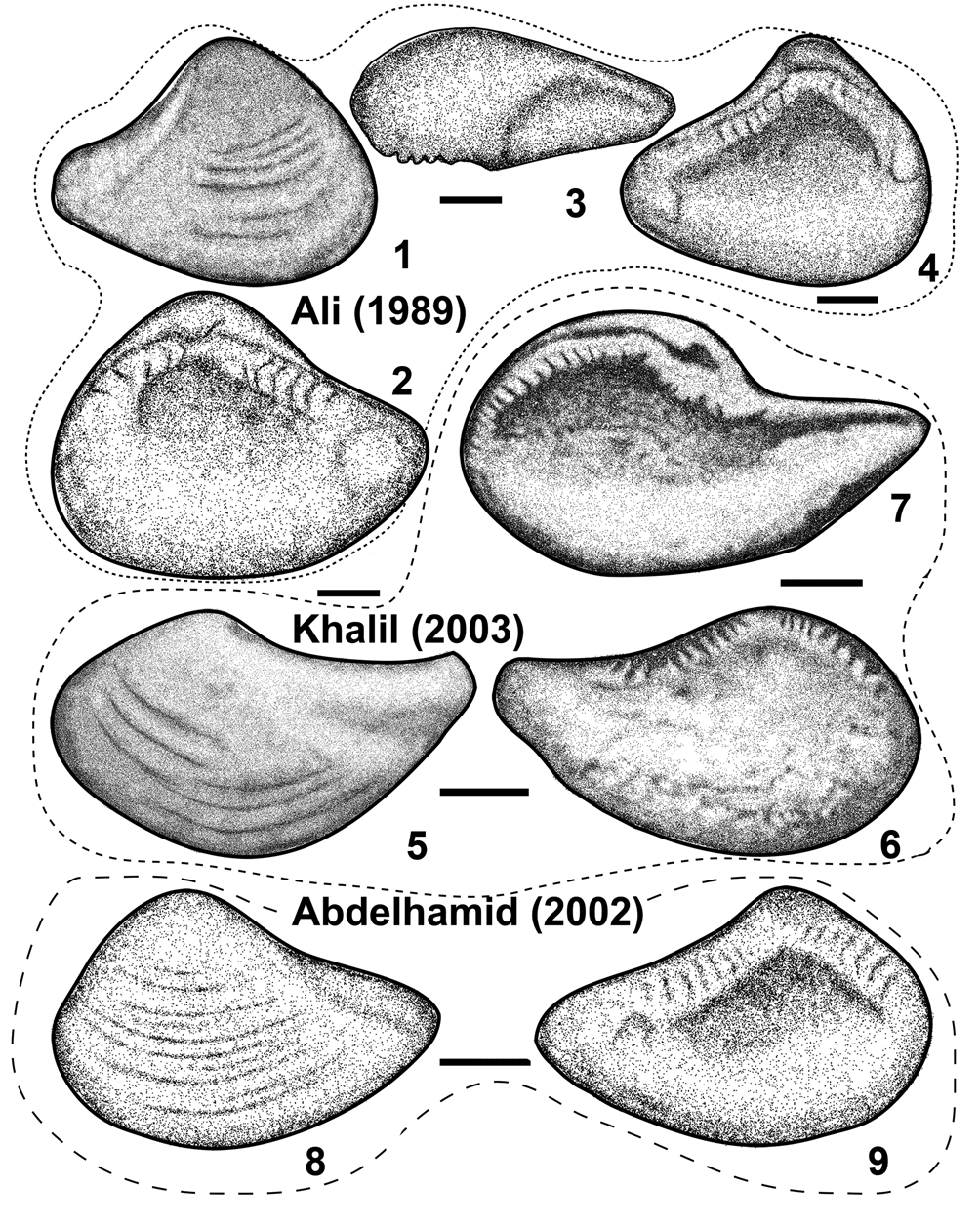
Figure 19. (1–4) Sketches of Nuculana rostrata Ali, Reference Ali1989, from the lower Bathonian Safa Formation, Gebel Maghara (homonym of the type species of Nuculana, viz. N. rostrata [Gmelin, 1791] = Dacryomya lacryma [J. de C. Sowerby, Reference Sowerby1824]). (5–7) Cuspidaria sp., Khalil, Reference Khalil2003, from upper Bathonian rocks, Gebel Maghara (= D. lacryma [J. de C. Sowerby, Reference Sowerby1824]). (8, 9) Nuculana rostrata Ali et al., Reference Ali, Azab and Hussain1997, of Abdelhamid (Reference Abdelhamid2002) from the upper Bathonian‒Callovian of Gebel Maghara (= Dacryomya lacryma [J. de C. Sowerby, Reference Sowerby1824]). Scale bars = 2 mm.
Nuculana (Dacryomya) cf. D. lacryma (Sowerby) of Choubert and Faure-Muret (Reference Choubert and Faure-Muret1967) from the Middle Jurassic of Morocco differs from the holotype of Sowerby in having strong commarginal ribs, a shorter rostrum, and much greater inflation. Dacryomya lacryma differs from D. diana (d'Orbigny, 1850) in having a longer rostrum, a deep escutcheon delimited by a sharp ridge, and a strongly convex anterodorsal margin. Dacryomya heberti (Martin, Reference Martin1860) of Hodges (Reference Hodges2000, p. 22, pl. 1, figs. 31–43, 46, 48–50, aff. fig. 30, text-figs. 16–23) from the Lower Jurassic (Pliensbachian) of England differs from the present species in being less elongated, lacking the umbonal posterior ridge, and in having a more posteriorly placed umbo. A strongly inflated valve and a short rostrum distinguish the Toarcian Nuculana (Dacryomya) thompsoni Cox (Reference Cox1965, p. 26, pl. 1, figs. 4a–c) from Kenya from the present species. The figured specimens of N. (D.) dodsoni Cox (Reference Cox1965, p. 27, pl. 1, figs. 2a–c) from Bathonian‒Callovian rocks of the same area are internal molds and differ from D. lacryma in being more rostrate.
Genus Ryderia Wilton, Reference Wilton1830
Type species
Leda renevieri Oppel, Reference Oppel1856, from the Lower Lias of Awre, Gloucester, southern England; subsequent designation by Cox (Reference Cox1936).
Remarks
Douvillé (Reference Douvillé1916) erected Leda decorata from Middle Bathonian rocks of Gebel Maghara, Egypt. Later on, Abdelhady and Fürsich (Reference Abdelhady and Fürsich2014, fig. 6E, F) assigned Leda decorata to Ryderia. According to Damborenea and Pagani (Reference Damborenea and Pagani2019), the records of Ryderia from the Middle Jurassic of Egypt are doubtful and, therefore, they suggested that Douvillé's species probably belongs to the genus Nuculana. Based on the diagnostic features of Ryderia (a very long, narrow rostrum, a well-developed umbonal ridge, a well-developed corselet, an entire pallial line, and fine commarginal striae; Hodges, Reference Hodges2000, p. 40, 45), the present material (138 well-preserved specimens) can be placed in Ryderia without any doubt (Fig. 18.8–18.23). The genus Ryderia has been recorded from Rhaetian–Toarcian strata (Cox, Reference Cox1965; Liu, Reference Liu1995; Ivimey-Cook et al., Reference Ivimey-Cook, Hodges, Swift, Radley, Swift and Martill1999; Fürsich et al., Reference Fürsich, Berndt, Scheuer and Gahr2001; Yin and McRoberts, Reference Yin and McRoberts2006; Mander et al., Reference Mander, Twitchett and Benton2008; Ros et al., Reference Ros, Márquez-Aliaga and Damborenea2014). According to Hodges (Reference Hodges2000, p. 40), Ryderia extends from the Carboniferous to the Lower Jurassic (upper Pliensbachian) of Europe, South America, Mexico, Afghanistan, Japan, China, and New Zealand. Ros et al. (Reference Ros, Márquez-Aliaga and Damborenea2014, p. 19) did not agree with Hodges, because he did not provide references to confirm his statement. In addition, the oldest well-documented records of Ryderia are from Rhaetian beds (e.g., Ivimey-Cook et al., Reference Ivimey-Cook, Hodges, Swift, Radley, Swift and Martill1999) and it extends to the lower Toarcian (e.g., Fürsich et al., Reference Fürsich, Berndt, Scheuer and Gahr2001). Damborenea and Pagani (Reference Damborenea and Pagani2019, p. 934) pointed out that Middle Jurassic records of Ryderia are doubtful. The latter view is not accepted here, because Dietze et al. (Reference Dietze, Gräbenstein, Franz, Schweigert and Wetzel2021, p. 48, pl. 27, fig. 6) recorded Ryderia doris (d'Orbigny, 1849) from the early Middle Jurassic (Aalenian, ammonite Opalinum Zone) of southwestern Germany. In the present study, the occurrence of Ryderia is extended to the lower Upper Jurassic of Egypt (Fig. 20). It has been recorded from the Toarcian Shusha Formation, middle‒upper Bathonian Kehailia Formation, and from the Lower Kimmeridgian Masajid Formation of Gebel Maghara. Because there are no previous records of Ryderia from Africa (Ros et al., Reference Ros, Márquez-Aliaga and Damborenea2014, fig. 4; Fig. 20), this is also the first record of the genus from Africa.
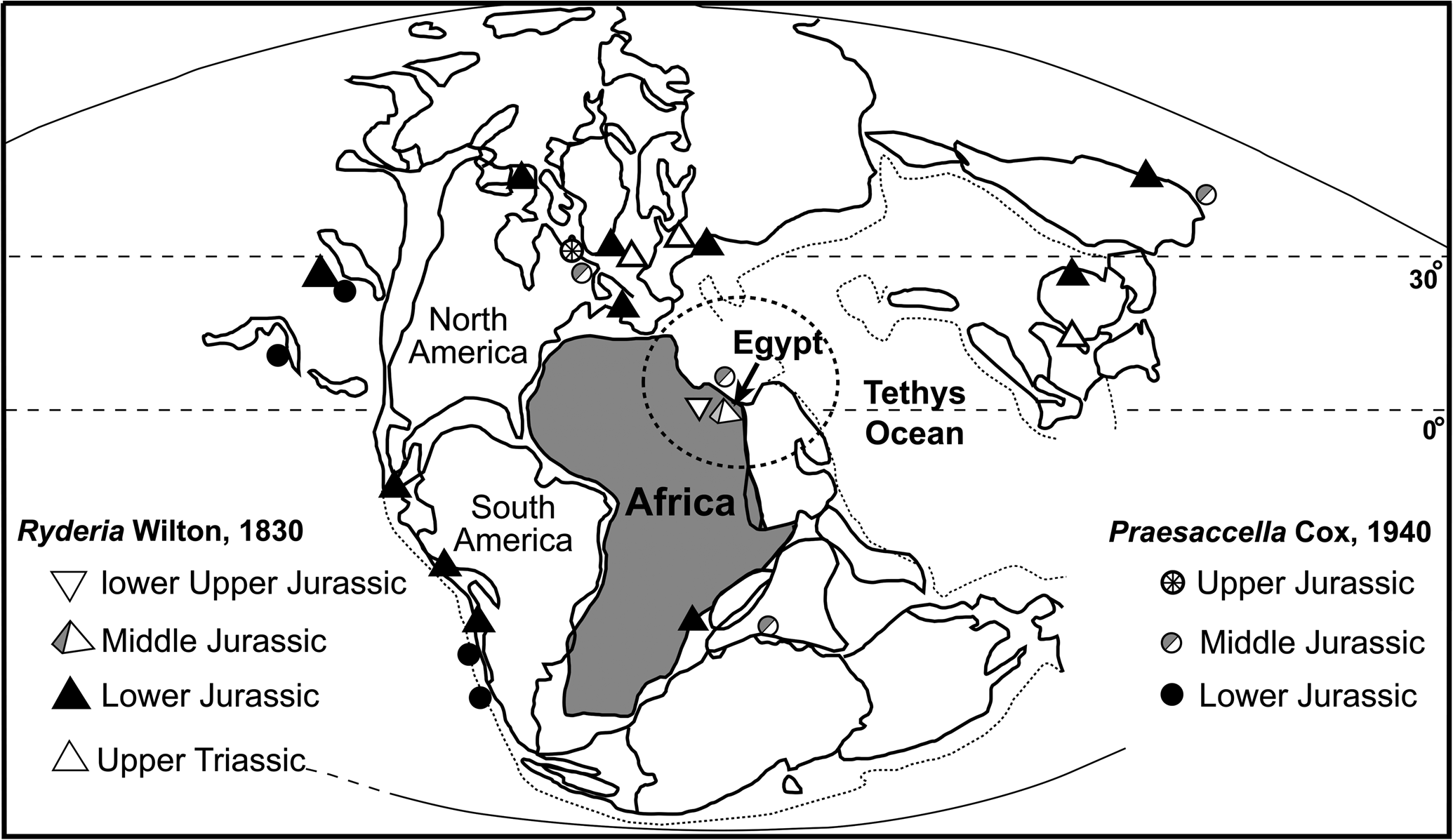
Figure 20. Paleogeographic distribution of Ryderia Wilton, Reference Wilton1830, and Praesaccella Cox, Reference Cox1940, during Upper Triassic and Jurassic time. See Figure 15 for references.
Ryderia decorata (Douvillé, Reference Douvillé1916)
Figures 18.8–18.26, 21.1–21.3
- *Reference Douvillé1916
Leda Decorata Douvillé, p. 61, pl. 5, figs. 56–62.
- .Reference Hirsch1980
Nuculana decorata; Hirsch, p. 130, pl. 1, fig. 9.
- non Reference Parnes1981
Ryderia sp. cf. R. decorata; Parnes, p. 27, pl. 3, figs. 27–28.
- .Reference Abdelhamid2002
Nuculana decorata; Abdelhamid, p. 337, pl. 5, figs. 5, 6.
- v.Reference Abdelhady2014
Ryderia decorata; Abdelhady, p. 72, fig. 5.4E, F.
- v.Reference Abdelhady and Fürsich2014
Ryderia decorata; Abdelhady and Fürsich, p. 181, fig. 6E, F.

Figure 21. (1–3) Ryderia decorata (Douvillé, Reference Douvillé1916) from the Jurassic (Toarcian–lower Kimmeridgian) rocks of Gebel Maghara; (1) BSPG 2014V 414/1, Masajid Formation, western Bir Maghara, dorsal view showing a well-developed corselet (Ct), (2, 3) BSPG 2014V 243/4, Kehailia Formation, Gebel Homayir; dorsal view and close-up, respectively, showing a well-ornamented escutcheon (Esc), located within corselet (Ct), and separated from main valve by corselet ridge. (4–8) Rollieria? sp. cf. Rollieria aequilateralis (Roemer, Reference Roemer1836) from the Middle–Upper Jurassic of Gebel Maghara. (4, 5) BSPG 2014V 324/1, Bir Maghara Formation, Gebel Arousiah, right lateral and left lateral views, respectively; (6–8) BSPG 2014V 351/1, Safa Formation, Bir Maghara, right valve exterior, dorsal view, and close-up and sketch, respectively, showing the convexodont teeth, a small resilifer (R), and rectangular anterior and posterior muscle scars (Aa, Pa). (9–15) Rollieria? sp. indet. from the Middle Jurassic of Gebel Maghara. (9–11) BSPG 2014V 352/1, Bir Maghara Formation, Gebel Arousiah, incomplete left lateral view, right valve, and dorsal view, respectively, (12–15) BSPG 2014V 353/1, Kehailia Formation, Gebel Homayir, right lateral view, left lateral view, dorsal view, and close-up showing the poorly preserved taxodont hinge (arrowed), respectively. (16–18) Nuculana (Nuculana) sp. indet., BSPG 2014V 322/1, middle‒upper Bathonian Kehailia Formation, Gebel Mowerib, left lateral view, dorsal view, and left valve interior, respectively. (19–31) Praesaccella juriana Cox, Reference Cox1940, from the Middle Jurassic of Gebel Maghara. (19, 20) BSPG 2014V 355/1, Bir Maghara Formation, Gebel Arousiah, left valve and right valve, respectively, (21–30) Kehailia Formation, Gebel Mowerib; (21, 22) BSPG 2014V 218/1, right valve and dorsal views, respectively, (23) BSPG 2014V 218/2, exterior view of right valve with gastropod and bivalve encrustations, (24, 25) BSPG 2014V 218/3, left valve and right valve, respectively, (26, 27) BSPG 2014V 218/4, right valve and dorsal views, respectively, (28–30) BSPG 2014V 218/5, left valve, right valve, and dorsal view, respectively; (31) BSPG 2014V 354/1, Kehailia Formation, Gebel Homayir, left lateral view showing the taxodont hinge. (32) Costinuculana magharensis Ayoub-Hannaa, Fürsich, and Abdelhamid, 2017, BSPG 2014V 356/1, Kehailia Formation, Gebel Homayir, left lateral view. Scale bars = 3 mm.
Holotype
Leda decorata Douvillé, Reference Douvillé1916, pl. 5, figs. 56–62, from the Middle Jurassic (Bathonian) of Gebel Maghara, Sinai Peninsula, Egypt.
Occurrence
Toarcian–lower Kimmeridgian of Gebel Maghara (Douvillé, Reference Douvillé1916; present study).
Description
Shell small, elongated-ovate, distinctly longer than high (H/L: ?0.41–0.49; Table 11), very inequilateral, equivalved, moderately inflated. Posterior end tapering, forming a narrow and long rostrum (complete rostra not preserved in all specimens). Anterior margin strongly convex, meeting ventral margin in rounded curve. Ventral margin convex, becoming straight to slightly concave posteriorly. Posterodorsal margin straight to slightly concave, anterodorsal margin slightly convex. Beak hardly prominent, small, opisthogyrate, located anteriorly. Escutcheon (Esc) wide, elongated-cordate, rounded at umbonal end, tapering towards posterior, located within corselet (Ct), ornamented with fine ribs, occasionally producing a chevron pattern (Fig. 21.1–21.3). Corselet elongated-cordate in outline, slightly concave, separated from main valve by rounded posterior ridge, extending from umbo to a position approximately one-third along posterodorsal margin. Posterior umbonal sulcus (ventral of posterior ridge) narrow, deep, gradually widening towards ventral margin (Fig. 18.14). Lunule distinct, shallow, lanceolate, sharply pointed at extremities with two rows of teeth-like granules (external projections of anterior teeth; Fig. 22), arranged parallel to anterodorsal margins of both valves. Posterior adductor muscle scar sub-ovate, more elongated than anterior one, located close to posterodorsal margin. Anterior adductor muscle scar sub-rounded, located at anterior end. Visceral muscle scar (VMS) preserved on few internal molds as grooves extending from umbo towards ventral margin, fading approximately at the mid-height of shell (Fig. 18.24–18.26). Umbonal pedal muscle scars (UPS) well preserved, located dorso-ventrally below umbo (Fig. 18.25). Hinge taxodont, slightly gradidentate. Anterior part of hinge slightly convex dorsally with 17 convexodont teeth and sockets, gradually decreasing in size towards umbo (Fig. 18.18–18.20). Posterior part of hinge dorsally concave with ~10 chevron-like teeth and sockets, pointing and decreasing in size towards umbo. Chondrophore small, triangular, shallow, located below the smallest five anterior teeth below umbo (Fig. 18.20). Ornament consisting of numerous, regular, fine commarginal ribs (Fig. 18.17, 18.23).

Figure 22. Relationships between the two rows of tubercles or granules on the lunule and development of anterior teeth in Ryderia decorata (Douvillé, Reference Douvillé1916) from the Jurassic of Gebel Maghara.
Table 11. Measurements (in mm) of Ryderia decorata (Douvillé, Reference Douvillé1916).

Materials
One hundred and thirty-seven specimens, mostly articulated and in shell preservation, from the Toarcian–lower Kimmeridgian strata of the Maghara area: three specimens from the marl beds of the Toarcian Shusha Formation (BSPG 2014V 418/1–3); 63 specimens from the middle–upper Bathonian Kehailia Formation, Gebel Homayir (BSPG 2014V 242/1–3; 243/1–31; 341/1–6; 342/1–14; 347/1–4; 348/1–2; 350/1–3); 11 specimens from the same formation, Gebel Mowerib (BSPG 2014V 343/1; 346/1; 349/1–9); seven specimens from the Kehailia Formation, Gebel Arousiah (BSPG 2014V 320/1–4; 321/1–2; 344/1); two specimens from the Kehailia Formation, Gebel Engabashi (BSPG 2014V 3/1–2); 37 specimens from the basal part (marl and bioclastic marl beds) of the lower Kimmeridgian Masajid Formation, western Bir Maghara (BSPG 2014V 345/1–19; 415/1–5; 416/1–13); and 14 specimens from the upper part of the same formation, western Bir Maghara (BSPG 2014V 414/1–4; 423/1–10).
Measurements
See Table 11.
Remarks on measurements
In agreement with Hodges (Reference Hodges2000, p. 42), measuring the length of the species is often difficult, because the rostrum, being so long and thin, is commonly broken and often appears shorter than it actually was. Therefore, the H/L (shell outline) and Iav/L (shell inflation) ratios are not accurate. In the present study, the lengths of nearly complete specimens have been measured, but marked with a question mark.
Remarks
Douvillé (Reference Douvillé1916) erected Leda decorata from the middle Bathonian rocks of Gebel Maghara on the basis of a well-ornamented deep escutcheon, numerous fine commarginal ribs, and a large and moderately inflated valve (L >21.0 mm; Iav/L: ?0.31–0.36; Table 11). In addition, the lunule of R. decorata is narrow, shallow, and lanceolate with two rows of tubercles or granules arranged along the anterodorsal margins of both valves. According to Cox (Reference Cox1940, p. 31), the “row of pearls of lunule” described by Douvillé (Reference Douvillé1916) is due to some abnormal development of hinge-teeth, the positions of which are indicated by a series of projections on the exterior of the shell. This view is followed here, because the number of granules (external projections of teeth) are nearly the same as that of the anterior teeth. In addition, the external projections have the same arrangement as the anterior teeth in the two valves (tooth in left valve located opposite to socket in the right one), and occasionally have the same orientation as the anterior teeth (Fig. 22). Cox (Reference Cox1940) suggested that Leda decorata Douvillé is probably a junior synonym of Nuculana (Dacryomya) lacryma (J. Sowerby) from the Lower Callovian of India (Cox, Reference Cox1940, p. 29, pl. 2, figs. 2–5). Later, Jaitly et al. (Reference Jaitly, Fürsich and Heinze1995, p. 161) accepted Cox's view, which is not followed here. Some similarities exist between the two species, but also some differences. R. decorata has a long and narrow rostrum (rapidly narrowing posterior rostrum in D. lacryma), numerous fine commarginal ribs, well-ornamented escutcheon (smooth in D. lacryma), a well-developed umbonal posterior sulcus, extending from umbo to posteroventral corner, a shallow lanceolate lunule, a well-developed corselet (absent in D. lacryma), and is larger (Lmax> 21.0 mm as opposed to 13.5 mm in Cox, Reference Cox1940), less inflated, and more elongated (H/L: ?0.41–0.49 as opposed to 0.60; Cox, Reference Cox1940, p. 30). The most closely related species is R. doris (d'Orbigny, 1849) figured and described, for example, by Hodges (Reference Hodges2000, p. 40, pl. 3, figs. 1–4, 6–7, text-figs. 40–45) from the Lower Sinemurian of southeastern England and by Karapunar et al. (Reference Karapunar, Werner, Fürsich and Nützel2020, p. 15, pl. 2, figs. 9–19, pl. 3, figs. 1–8, pl. 10, fig. 12) from the Pliensbachian of southern Germany. Ryderia doris differs from R. decorata in having a very long, narrow rostrum (H/L = 0.26–0.37; Hodges, Reference Hodges2000, as opposed to 0.41–0.49), a shallow, smooth escutcheon (strongly ornamented in R. decorata), a well-developed rostral groove, and in being less inflated (Iav/L = 0.13–0.22; Karapunar et al., Reference Karapunar, Werner, Fürsich and Nützel2020, as opposed to ?0.30–0.36). In addition, R. decorata is stratigraphically younger (Toarcian–lower Kimmeridgian) than R. doris. Ryderia sp. cf. R. decorata (Douvillé, Reference Douvillé1916) of Parnes (Reference Parnes1981) from the Bajocian of Negev, southern Israel, has a strong posterior ridge extending from the umbo to the ventral end of the rostrum, a smooth shell surface, compressed valves particularly along the posterior area, and a narrower and much more elongated valve than Ryderia decorata.
Another very closely related species is N. rostrialis (Lamarck, Reference Lamarck1819), figured and described by some authors, including Goldfuss (1837, p. 155, pl. 125, figs. 8a–c) and Klöcker (Reference Klöcker1966, p. 220, fig. 1) from the Lower Jurassic of Germany, Bronn (Reference Bronn1836, p. 371, pl. 20, figs. 6a, b) from the Lower Jurassic of France, and Etter (Reference Etter1990, pl. 3, fig. 7) from the Lower Jurassic of Switzerland. Although the latter two species have the same shell outline, size and ornamentation, N. rostrialis differs in having a narrow, smooth escutcheon, a strong posterior ridge extending from the umbo to the ventral end of the rostrum, a smooth and sharply demarcated lunule, sharply pointed, slightly opisthogyrate beaks, and a slightly concave posterodorsal margin. Nuculana sp. figured by Hirsch (Reference Hirsch1980, p. 130, pl. 1, fig. 10) from the middle Callovian of Makhtesh Hatira, Israel, has a wider rostrum, faint radial ribs close to the ventral margin, and is larger (L = ?46.0 mm). The poorly preserved Nuculana (Ryderia) kenyana Cox, Reference Cox1965 (p. 27, pl. 1, figs. 6a–c) from the upper Toarcian of Kenya differs from the present species in having compressed valves, an angulated anterior margin, and lacks a lunule. With respect to general outline, Nuculana numismalis (Tate, Reference Tate1870) figured by Chen (Reference Chen1987, p. 40, pl. 2, fig. 1) from the Lower Jurassic of China is similar to the present species, but has a slightly concave posterodorsal margin, thick commarginal ribs, a less-convex ventral margin, and is much smaller than the present species. Ryderia texturata (Terquem and Piette, Reference Terquem and Piette1865) (p. 89, pl. 11, figs. 5, 6) from the Toarcian of France and Ryderia tehuelchana Damborenea and Pagani, Reference Damborenea and Pagani2019 (p. 934, figs. 5.1–5.5, 5.18, 7.4) from the Lower Jurassic of Chubut (Argentina) display a different ornamentation, particularly on the rostrum.
Family Sareptidae Stoliczka, Reference Stoliczka1870
Subfamily Yoldiinae Dall, Reference Dall1908
Genus Rollieria Cossmann, Reference Cossmann1920
Type species
Nucula palmae J. de C. Sowerby, Reference Sowerby1824, “Mountain Limestone” (Carboniferous) of Derbyshire, England; subsequent designation by Rollier, Reference Rollier1923.
Remarks
Rollieria was regarded as a subgenus of Nuculana by some workers (e.g., Cox et al., Reference Cox, Newell, Branson, Casey, Chavan, Coogan, Dechaseaux, Moore and Teichert1969, p. N237; Liu, Reference Liu1995). Hodges (Reference Hodges2000, p. 35) placed Rollieria as a separate genus within the subfamily Yoldiinae (family Yoldiidae) on the basis of a sub-central, slightly prosogyrate umbo, a shallow pallial sinus, an elongated valve, and the lack of an escutcheon and lunule. Later, Carter et al. (Reference Carter, Altaba, Anderson, Araujo, Biakov, Bogan and Campbell2011) assigned the subfamily Yoldiinae Dall, Reference Dall1908, to the family Sareptidae Stoliczka, 1870. The type species of J. Sowerby was collected from Carboniferous rocks of England. Hodges (Reference Hodges2000, p. 35–36) doubted the presence of Rollieria at that time because this is the only occurrence of the genus in strata older than Jurassic. He pointed out that, apart from its questionable occurrence in the Carboniferous, Rollieria has a stratigraphic range from the Lower Jurassic–Lower Cretaceous of Europe and India. In the present study, Rollieria is recorded from Middle–Upper Jurassic strata of Gebel Maghara for the first time.
Rollieria? sp. cf. Rollieria aequilateralis (Roemer, Reference Roemer1836)
Figure 21.4–21.8
- cf. Reference Roemer1836
Nucula aequilateralis Roemer, p. 101, pl. 6, fig. 13a–c.
Occurrence
Middle Jurassic of Germany and probably from the Middle–Upper Jurassic of Gebel Maghara (first record).
Description
Shell small, elongated-ovate, longer than high (H/L: ?0.60–0.62; Table 12) slightly equilateral, equivalved, and moderately inflated. Posterodorsal margin slightly convex to straight. Anterodorsal margin straight, meeting anterior margin at nearly a right angle. Anterior margin blunt, meeting ventral margin in rounded curve. Posterior margin broken off. No lunule and escutcheon (Fig. 21.7). Umbo low, moderately inflated, located slightly anterior of mid-length of valve. Beaks hardly prominent, prosogyrate. Anterior and posterior muscle scars partly preserved, subquadrate in outline, located close to anterior and posterior margin, respectively. Hinge taxodont with unequal teeth and sockets. Anterior and posterior hinge teeth meeting in an obtuse angle (125°), separated by small triangular resilifer, directed slightly posteriorly (Fig. 21.8). Posterior part of hinge longer than anterior one with ~17 convexodont teeth, decreasing in size towards beak. Anterior convexodont teeth larger and fewer (~7) than posterior ones. Shell surface with fine growth lines.
Table 12. Measurements (in mm) of Rollieria? sp. cf. Rollieria aequilateralis (Roemer, 1836).

Materials
Four specimens, three composite molds and two in shell preservation, from the Middle–Upper Jurassic of Gebel Maghara, a single specimen from the upper Bajocian Bir Maghara Formation, Gebel Arousiah (BSPG 2014V 324/1), a single left valve from the upper third part of the lower Bathonian Safa Formation, Bir Maghara (BSPG 2014V 351/1), and two specimens from the lower marl beds of the early Kimmeridgian Masajid Formation, western Bir Maghara (BSPG 2014V 412/1–2).
Measurements
See Table 12.
Remarks
Based on the diagnostic features of Rollieria mentioned by Hodges (Reference Hodges2000, p. 35) the present specimens might well be Rollieria. Due to the missing posterior area (incomplete shell outline), however, the generic assignment is somewhat doubtful. Therefore, the material is assigned to the genus with question mark. Nucula aequilateralis Roemer (1836, p. 101, pl. 6, figs. 13a–c) from the Upper Jurassic of Germany, which is comparable in shape and size, is elongated subtriangular, has a narrow, blunt anterior margin, a smooth shell surface, and lacks escutcheon and lunule. The incomplete preservation makes it difficult to place the present specimens in this species with certainty.
Rollieria? sp. indet.
Figure 21.9–21.15
Description
Shell small (L >9.0 mm; H = 6.7 mm), incomplete, outline apparently subovate, equivalved, inequilateral, and moderately inflated (Iav/H = 0.52). Posterior margin convex, meeting ventral margin in rounded curve. Anterior margin broken off. Ventral margin strongly rounded. Beaks small, slightly prosogyrate, located approximately anterior of mid-length. No lunule or escutcheon (Fig. 21.11, 21.14). Hinge taxodont with numerous teeth and sockets (Fig. 21.15). Shell surface with irregular commarginal growth lines.
Materials
Four incomplete composite molds from the Middle Jurassic of Gebel Maghara; a single specimen from the Bajocian Bir Maghara Formation, Gebel Arousiah (BSPG 2014V 352/1) and three specimens from the middle–upper Bathonian Kehailia Formation, Gebel Homayir (BSPG 2014V 353/1–3).
Remarks
None of the present specimens is complete. This incomplete preservation makes it difficult to place them in any of the known species. However, they are somewhat similar to Rollieria bronni (Andler, Reference Andler1858) of Hodges (Reference Hodges2000, pl. 2, figs. 31–35, text-figs. 35–38) from the Lower Jurassic of England in having rounded margins and a smooth surface, but the latter is more elongated. Recently, Karapunar et al. (Reference Karapunar, Werner, Fürsich and Nützel2020) regarded Rollieria bronni as a synonym of their new species R. goldfussi (Karapunar et al., Reference Karapunar, Werner, Fürsich and Nützel2020, p. 16, pl. 13, figs. 9–12, pl. 4, figs. 1, 2, text-figs. 10–12), which has been recorded from the Pliensbachian of southern Germany. Another closely similar species is R. aequilatera (Koch and Dunker, Reference Koch and Dunker1837) of Cox (Reference Cox1965, p. 29, pl. 1, fig. 5a–c) from the Toarcian of Kenya, but the latter has strongly rounded margins and compressed valves.
Family Nuculanidae Adams and Adams, Reference Adams and Adams1858
Genus Nuculana Link, Reference Link1807
Subgenus Nuculana Link, Reference Link1807
Type species
Arca rostrata Chemnitz, Reference Chemnitz1774 (= Arca pernula Müller, Reference Müller1779) by original designation (Link, 1807, p. 155).
Nuculana (Nuculana) sp. indet.
Figure 21.16–21.18
Description
Shell small (L = ?17.5 mm), longer than high, inequilateral, weakly inflated, with a long posterior rostrum. Posterodorsal margin slightly concave. Anterodorsal margin slightly convex. Ventral margin broken off. Beak less prominent, opisthogyrate, and located anteriorly. Escutcheon less distinct, shallow, and ornamented (Fig. 21.17). Lunule narrow, shallow, lanceolate. Hinge gradidentate with convexodont teeth. Anterior part of hinge dorsally convex, posterior hinge dorsally concave, both separated by triangular, narrow, deep resilifer with slightly concave and smooth area located anterior of resilifer (Fig. 21.18). Anterior teeth strong, convexodont, sharply pointed, and decreasing in size towards umbo. Ornamentation consisting of faint commarginal striae (Fig. 21.16).
Materials
A single incomplete specimen from the middle‒upper Bathonian Kehailia Formation, Gebel Mowerib, Gebel Maghara (BSPG 2014V 322/1).
Remarks
Structure of the teeth and general outline allow assignment of the present specimen to the genus Nuculana. It can be distinguished from other Jurassic Nuculana species by having a shallow and ornamented escutcheon, a very narrow lanceolate lunule, and a resilifer bordered anteriorly by a slightly concave, smooth area (Fig. 21.18). Because only an incomplete left valve is available, identification at the species level is not possible. The valve resembles Leda alpina d'Orbigny figured by Cottreau (Reference Cottreau1925, p. 143, pl. 17, fig. 3) from the Callovian of France in general outline, but the latter differs in being smaller and in having more-inflated umbones.
Genus Praesaccella Cox, Reference Cox1940
Type species
Nuculana (Praesaccella) juriana Cox, Reference Cox1940, from the Callovian of Kachchh, India, by original designation.
Remarks
Praesaccella was erected as a subgenus of Nuculana by Cox (Reference Cox1940, p. 32) based on an entire pallial line and relatively few hinge teeth. Recently, Damborenea and Pagani (Reference Damborenea and Pagani2019, p. 933) regarded Praesaccella as a separate genus when describing P. ovum (J. de C. Sowerby, Reference Sowerby1824) from the Lower Jurassic of Argentina. Most Jurassic Praesaccella species, such as Nuculana (Praesaccella) camelorum Cox, Reference Cox1965 (p. 28, pl. 2, fig. l0a, b) from the Toarcian or Bajocian of East Africa, N. (P.) juriana Cox, Reference Cox1940 (p. 33, pl. 2, figs. 6–9), and N. (P.) calloviensis (Kanjilal and Singh, Reference Kanjilal and Singh1973) (p. 469, pl. 1, figs. a–f) from Callovian–Oxfordian rocks of western India have an acutely pointed posterior end, which is almost at mid-height or slightly above the mid-height of the valve. The latter feature, therefore, should be included as diagnostic of the genus.
Praesaccella has a wide geographic distribution; it has been recorded from the Lower Jurassic of Chile (Aberhan, Reference Aberhan1994, p. 10), Argentina (Damborenea, Reference Damborenea1987; Damborenea and Pagani, Reference Damborenea and Pagani2019), Middle Jurassic of western India (Cox, Reference Cox1940; Kanjilal and Singh, Reference Kanjilal and Singh1973; Jaitly et al., Reference Jaitly, Fürsich and Heinze1995), Spain (Delvene, Reference Delvene2001), Japan (Hayami, Reference Hayami1961), and Toarcian or Bajocian of East Africa (Cox, Reference Cox1965). The species is recorded from Middle Jurassic rocks (upper Bajocian–Bathonian) of Egypt for the first time (Fig. 20).
Praesaccella juriana Cox, Reference Cox1940
Figure 21.19–21.31
- *Reference Cox1940
Nuculana (Praesaccella) juriana Cox, p. 33, pl. 2, figs. 6–9.
- .Reference Jaitly, Fürsich and Heinze1995
Nuculana (Praesaccella) juriana; Jaitly et al., p. 159, pl. 2, figs. 9–12, text-fig. 12.
Holotype
Nuculana (Praesaccella) juriana Cox, Reference Cox1940, pl. 2, fig. 7, from the Callovian of Kachchh, western India.
Occurrence
Callovian–Oxfordian of India and Bajocian to late Bathonian of Gebel Maghara (present study, first record).
Description
Shell small, subtriangular, longer than high (H/L: 0.65–0.70; Table 13), inequilateral, equivalved, moderately inflated, and posteriorly elongated into short rostrum. Anterior margin strongly convex meeting ventral margin in rounded angle. Posterior end of rostrum narrowly rounded, occasionally angulated, and located slightly above mid-height of valve (Fig. 21.19, 21.23). Ventral margin regularly rounded. Posterodorsal margin slightly concave, gradually sloping posteriorly. Umbones triangular, depressed, and located anterior of mid-length. Beak sharply pointed, orthogyrate to slightly prosogyrate. Escutcheon lanceolate, narrow, shallow, and delimited by a moderately well-developed ridge extending from umbo to end of rostrum (Fig. 21.22, 21.27). No lunule. Anterior part of hinge dorsally convex, posterior one slightly concave and carries numerous teeth and sockets (Fig. 21.14). Ornamentation consisting of numerous very fine commarginal striae except for posterodorsal and umbonal areas (Fig. 21.19, 21.25).
Table 13. Measurements (in mm) of Praesaccella juriana Cox, Reference Cox1940.

Materials
Seventeen articulated specimens with shell preservation, from the Middle Jurassic of Gebel Maghara: a single specimen from the upper Bajocian Bir Maghara Formation, Gebel Arousiah (BSPG 2014V 355/1), 14 specimens from the middle–upper Bathonian Kehailia Formation, Gebel Mowerib (BSPG 2014V 218/1–14), and two specimens from the Kehailia Formation, Gebel Homayir (BSPG 2014V 354/1–2).
Measurements
See Table 13.
Remarks
With respect to general outline, size, and ornamentation, the present specimens fit very well with the specimens of Nuculana (Praesaccella) juriana described and figured by Cox (Reference Cox1940) from the Middle–Upper Jurassic of India. The present species differs from the very similar Leda argoviensis Moesch, Reference Moesch1867 (p. 302, pl. 4, fig. 12a–c) from the Oxfordian of the Swiss Jura Mountains by having a longer rostrum, a depressed umbo (triangular in L. argoviensis), and a broadly convex anterior margin. The strongly convex posterior margin of L. dewalquei Terquem and Piette, Reference Terquem and Piette1865, figured by Stefanini (Reference Stefanini1939, p. 141, pl. 16, figs. 3–6) from the Lower Jurassic of Somalia distinguishes it from the present species. Praesaccella ovum (J. de C. Sowerby, Reference Sowerby1824) of Damborenea and Pagani (Reference Damborenea and Pagani2019, p. 9, figs 6.1–6.6, 7.5) from the Lower Jurassic of Argentina has a rounded posterior end, a strongly opisthogyrate umbo, regular commarginal lines, and its shell is less elongated than the present species. Nuculana (P.) camelorum Cox, Reference Cox1965 (p. 28, pl. 2, fig. 10a, b) from Toarcian or Bajocian strata of Kenya is more rostrate, more inflated, and has a well-developed umbonal posterior carina.
Subfamily Veteranellinae Chen, Liu, and Lan, Reference Chen, Liu and Lan1983
Genus Costinuculana Ayoub-Hannaa, Abdelhady, and Fürsich, Reference Ayoub-Hannaa, Abdelhady and Fürsich2017
Type species
Costinuculana magharensis Ayoub-Hannaa, Abdelhady, and Fürsich, Reference Ayoub-Hannaa, Abdelhady and Fürsich2017, from the middle Bathonian of Gebel Maghara, North Sinai, Egypt, by monotypy.
Diagnosis
See Ayoub-Hannaa et al. (Reference Ayoub-Hannaa, Abdelhady and Fürsich2017, p. 435–437).
Remarks
The genus Costinuculana and the species C. magharensis have been described, figured, and discussed in detail by Ayoub-Hannaa et al. (Reference Ayoub-Hannaa, Abdelhady and Fürsich2017).
Costinuculana magharensis Ayoub-Hannaa, Abdelhady, and Fürsich, Reference Ayoub-Hannaa, Abdelhady and Fürsich2017
Figure 21.32
- v.Reference Abdelhady2014
Nuculoidea n. sp. Abdelhady, p. 72, fig. 5.4A, B.
- v.Reference Abdelhady and Fürsich2014
Nuculoidea n. sp. Abdelhady and Fürsich, p. 181, fig. 6a, b.
- *Reference Ayoub-Hannaa, Abdelhady and Fürsich2017
Costinuculana magharensis Ayoub-Hannaa, Abdelhady, and Fürsich, p. 437, figs. 4.1–4.9, 5.1–5.21, 6, 7.2.
Holotype
Articulated specimen BSPG 2014V 1 (Ayoub-Hannaa et al., Reference Ayoub-Hannaa, Abdelhady and Fürsich2017, fig. 5.13–5.17) from the middle Bathonian of Gebel Maghara, Egypt.
Occurrence
Middle Bathonian Kehailia Formation, Gebel Maghara (Ayoub-Hannaa et al., Reference Ayoub-Hannaa, Abdelhady and Fürsich2017; present study).
Description
Shells small, elongated-ovate in outline, strongly inequilateral, posteriorly elongated with ribbed rostrum. Anterodorsal margin slightly convex, meeting anterior margin in strongly rounded curve. Posterodorsal margin long, slightly concave. Anterior margin narrow and strongly convex. Posterior margin broken off. Ventral margin broad, irregular, slightly convex towards anterior margin, slightly concave posteriorly. Umbones broad, strongly convex, and located anteriorly. Beak sharply pointed and strongly incurved posteriorly. Escutcheon wide, moderately deep, and well ornamented with fine riblets. Surface with well-developed sub-vertical ribs, asymmetrical in cross-section, covering an area ~45% of total valve length from posterior end, and separated by deep, narrow interspaces. Area between these radial ribs and anterior margin covered by reticulate ornamentation (Fig. 21.32).
Materials
A single specimen from the Middle Jurassic (Bathonian) of Gebel Maghara, Kehailia Formation, Gebel Homayir (BSPG 2014V 356/1).
Measurements
See Table 14.
Table 14. Measurements (in mm) of Costinuculana magharensis Ayoub-Hannaa, Abdelhady, and Fürsich, Reference Ayoub-Hannaa, Abdelhady and Fürsich2017.

Remarks
According to Ayoub-Hannaa et al. (Reference Ayoub-Hannaa, Abdelhady and Fürsich2017), the taxonomic characters of Costinuculana (type species: Costinuculana magharensis) are not present in the majority of Jurassic nuculanid genera that have been identified from different areas (e.g., Pandey and Singh, Reference Pandey and Singh1981; Fürsich, Reference Fürsich1982; Aberhan, Reference Aberhan1994, Reference Aberhan1998; Jaitly et al., Reference Jaitly, Fürsich and Heinze1995; Sha et al., Reference Sha, Fürsich, Smith and Wang1998). Costinuculana can be distinguished from other Jurassic nuculanids by having strong, rounded, and oblique radial ribs on the rostrum (occasionally bifurcated ventrally), a wide and deep escutcheon, which is delimited by blunt ridges and ornamented by fine riblets, and a narrow, lanceolate, sharply demarcated lunule. In the present material, the area between the oblique radial ribs and anterior margin is covered by reticulate ornamentation (Fig. 21.32). For more details concerning diagnosis, description, and comparison with closely related taxa, see Ayoub-Hannaa et al. (Reference Ayoub-Hannaa, Abdelhady and Fürsich2017).
According to Damborenea and Pagani (Reference Damborenea and Pagani2019), Costinuculana is closely related to Ryderia. Actually, Ryderia has a very narrow and much more elongated rostrum, a narrow and smooth escutcheon (deep, ornamented, and delimited by blunt ridges in Costinuculana, see Ayoub-Hannaa et al., Reference Ayoub-Hannaa, Abdelhady and Fürsich2017, figs. 4.1, 4.5, 4.7, 5.17), a less-distinct lunule (sharply demarcated in Costinuculana), and is less inflated (moderately to strongly inflated in the present material). In addition, the strength, number, and development of ribs on the rostrum of Costinuculana are quite different from those of species of Ryderia.
Conclusions
Based on detailed analyses of several thousand specimens and on multivariate analyses of morphological features, the Jurassic protobranch bivalves from Gebel Maghara (northern Sinai, Egypt) were systematically described, and the genus and species boundaries were clearly defined. In addition, the paleobiogeographic patterns of some Jurassic protobranch genera were mapped (Figs. 15, 20). Due to the excellent preservation of many specimens, morphological features could be described in great detail. Among the sixteen taxa, three species were new—Nuculoma douvillei n. sp., N. sinaiensis n. sp., and Palaeoneilo aegyptiaca n. sp. In addition, five species—Palaeonucula cuneiformis (J. de C. Sowerby), P. muensteri (Goldfuss), Dacryomya diana (d'Orbigny), D. lacryma (J. de. C. Sowerby), and Praesaccella juriana Cox—had not been recorded from the Jurassic rocks of Egypt before. In the past, Middle and Upper Jurassic records of the genus Palaeoneilo have not been accepted due to insufficient knowledge of morphological details. Based on well-preserved internal and external characters, the occurrence of Palaeoneilo in Middle Jurassic (Bajocian‒Bathonian) strata of Gebel Maghara can be confirmed. Similarly, Bathonian‒Kimmeridgian records of Dacryomya and Ryderia from the Gebel Maghara area considerably extend the ranges of these genera. The wide geographic distribution of the identified taxa suggests that they were not influenced to a great extent by latitudinal climate gradients.
Acknowledgments
We thank B. Leipner-Mata (Erlangen) for preparing the polished slabs and thin-sections. A.A. Abdelhady gratefully acknowledges the German Academic Exchange Service (DAAD) and the Egyptian Ministry of Higher Education and Scientific Research “GERLS” Programme for their financial support. We thank the reviewers, M. Aberhan and S. Damborenea, for their careful and constructive criticism of the manuscript.
Author contributions
The material for this study was collected by AAA and FTF and identified by WA-H, AAA, and FTF. The manuscript was largely written by WA-H. FTF extensively discussed taxonomic problems with WA-H and improved the manuscript. All authors read and commented on the final version of the manuscript.
Declaration of competing interests
The authors declare none.
Appendix A
Dimensions of Nuculoma douvillei n. sp. See Figure 3 for explanation of measurements and ratios; measurements in mm.

Appendix B
Dimensions of Nuculoma wynnei Cox, Reference Cox1940, and Nuculoma sinaiensis n. sp. See Figure 3 for explanation of measurements and ratios; measurements in mm.






































 Open access
Open access