


Introduction
Macaws are among the largest and most threatened parrots (BirdLife International, 2013) and populations are in decline as a result of illegal trapping and habitat loss (Snyder et al., Reference Snyder, McGowan, Gilardi and Grajal2000). Many of the 16 species of macaws require large tracts of relatively pristine habitat for their survival (Forshaw, Reference Forshaw1989). Macaws are adversely affected by anthropogenic activities such as establishment and expansion of roads, towns and cities, collection for the pet trade, hunting, and clearing, alteration and degradation of habitat (Gonzalez, Reference Gonzalez2003).
Three species of macaws coexist in the Beni savannahs of Bolivia: the blue-throated Ara glaucogularis, blue-and-yellow Ara ararauna and red-and-green Ara chloropterus macaws (Hanagarth & Beck, Reference Hanagarth and Beck1996; Yamashita & de Barros, Reference Yamashita and de Barros1997). All three have experienced population declines in the Beni savannahs since the 1960s as a result of local hunting for feathers, poaching for the pet trade, and habitat loss (Ingels et al., Reference Ingels, Parkes and Farrand1981; Boussekey et al., Reference Boussekey, Morvan and Saint-Pie1997; Hesse & Duffield, Reference Hesse and Duffield2000).
Rediscovered in the wild in 1992 (Jordan & Munn, Reference Jordan and Munn1993), the blue-throated macaw has undergone a significant population decline and is reported to be the rarest of the macaws in the wild (Hesse & Duffield, Reference Hesse and Duffield2000). The estimated population of this Critically Endangered Bolivian endemic is 115 individuals (BirdLife International, 2013). Since 1997 governmental and non-governmental organizations have been working to increase the wild population of the species (Duffield & Hesse, Reference Duffield and Hesse1997; Hesse & Duffield, Reference Hesse and Duffield2000; Herrera et al., Reference Herrera, Vargas, Sandoval, Perskin and Rendón2007; Berkunsky, Reference Berkunsky2008, Reference Berkunsky2010). Each organization has its own monitoring scheme that usually includes annual surveys of sites where the species’ presence was confirmed previously, and surveys of new areas in search of new populations.
The blue-and-yellow and red-and-green macaws are two of the most striking Neotropical parrots and are well known in the pet trade (Gonzalez, Reference Gonzalez2003; Brightsmith & Bravo, Reference Brightsmith and Bravo2006). Both are categorized as Least Concern on the IUCN Red List (BirdLife International, 2013) but local populations have been reduced in parts of their range, especially in areas near human population centres (Forshaw, Reference Forshaw1989; Juniper & Parr, Reference Juniper and Parr1998). The blue-and-yellow macaw may be the most resilient of the three species to human activity, whereas the red-and-green macaw appears to be the most sensitive (Karubian et al., Reference Karubian, Fabara, Yunes, Jorgenson, Romo and Smith2005; Franchin, Reference Franchin2009). Studies have examined multiple aspects of their biology, including activity patterns (Roth, Reference Roth1984), breeding biology (Brightsmith & Bravo, Reference Brightsmith and Bravo2006; Renton & Brightsmith, Reference Renton and Brightsmith2009) and habitat use (Karubian et al., Reference Karubian, Fabara, Yunes, Jorgenson, Romo and Smith2005; Brightsmith & Villalobos, Reference Brightsmith and Villalobos2011). The influence of habitat, season and detectability on abundance has been described for the blue-and-yellow and red-and-green macaws (Lee & Marsden, Reference Lee and Marsden2012). However, no information regarding occupancy and detectability has been recorded for the blue-throated macaw.
As a result of the difficulties in monitoring macaw species, estimates of abundance and occupancy are scarce. Most studies on macaw abundance disregard detectability and observations are usually based on encounter rates, which are dependent on sampling effort, rather than counts based on spatial sampling units (Karubian et al., Reference Karubian, Fabara, Yunes, Jorgenson, Romo and Smith2005).
Site occupancy modelling has become increasingly useful to ecologists because species detection or non-detection is determined over multiple sampling occasions, which allows estimates of detection probabilities (MacKenzie et al., Reference MacKenzie, Nichols, Royle, Pollock and Bailey2005; Kroll et al., Reference Kroll, Duke, Runde, Arnett and Austin2007; Weller, Reference Weller2008). Site occupancy modelling also provides a flexible framework that facilitates modelling of occupancy as a function of covariate information (MacKenzie et al., Reference MacKenzie, Nichols, Royle, Pollock and Bailey2005; Olea & Mateo-Tomás, Reference Olea and Mateo-Tomás2011). The use of covariates, such as distance to villages, elevation and human presence, can provide information about the factors influencing habitat use by species (Zeller et al., Reference Zeller, Nijhawan, Salom-Pérez, Potosme and Hines2011). Understanding how these variables affect the probabilities of occupancy and detection of the species is critical for the design of efficient monitoring protocols.
Here, we use occupancy models that account explicitly for detectability (MacKenzie et al., Reference MacKenzie, Nichols, Royle, Pollock and Bailey2005), to determine detection probabilities and occupancy of blue-throated, blue-and-yellow and red-and-green macaws in the Beni savannahs. We also investigate the coexistence of the three species. Finally, we discuss how our results could be applied to the on-going monitoring programme for the blue-throated macaw and for monitoring programmes for other macaw species.
Study area
We conducted surveys in the Beni savannahs (Fig. 1), a 160,000 km2 region of seasonally inundated lowlands occupying the Beni–Mamoré–Iténez basin in south-west Amazonia, Bolivia (Mayle et al., Reference Mayle, Langstroth, Fisher and Meir2007). Mean annual precipitation is 1,300–2,000 mm across the region, with precipitation occurring mostly during September–May (Hanagarth & Beck, Reference Hanagarth and Beck1996). The area is inhabited at low densities, with cattle-ranching being the primary economic activity (Mayle et al., Reference Mayle, Langstroth, Fisher and Meir2007).

Fig. 1 Location of the 29 sampling hexagons surveyed in the Beni savannahs, in northern Bolivia.
The landscape of the Beni savannahs is dominated by flat, low-lying areas of open, treeless savannah (Mayle et al., Reference Mayle, Langstroth, Fisher and Meir2007). Forest patches are scarce and restricted to areas that are sufficiently high to avoid flooding. Most forest patches are eroded relics of natural levees or terraces of abandoned river channels, which constitute fragments of gallery forest (Hanagarth & Beck, Reference Hanagarth and Beck1996). The Beni savannahs are poorly represented in Bolivia's protected areas, none of which overlap with our study site. Protected lowlands with little savannah are found at the borders of the ecoregion at the Beni Biological Station, 150 km from the study area (Beck & Moraes, Reference Beck, Moraes, Davis and Heywood1997).
Methods
From a randomly selected starting point we generated a hexagonal sampling grid that encompassed a significant proportion of the Beni savannahs region. The area of the hexagon (232.7 km2) was based on the mean recorded home range for breeding pairs of blue-throated macaws (IB, pers. obs.).
We conducted surveys from 8 September to 9 November 2011, at the end of the dry season, to coincide with the peak of the reproductive season of large macaw species in the Beni region (IB, pers. obs.). During the breeding season, breeding pairs remain in the proximity of nesting areas, which facilitates counts.
We surveyed 29 hexagons, covering 6,750 km2 (Fig. 1). Access limitations and time constraints prevented us from conducting repeat surveys in seven of the hexagons. Landscape heterogeneity resulted in a variable number of forest patches in each hexagon. Forest patches were surveyed by four trained observers (IB, GD, JADL and FG) during 06.00–10.00 and 14.00–18.00.
In each hexagon we selected several variables that could be affecting levels of occupancy of macaws (Table 1). We expected a higher probability of occupancy in hexagons where food and nesting resources were present and threats absent. The presence of other avian species or groups of species has been used as an indicator of habitat condition (Roberge & Angelstam, Reference Roberge and Angelstam2006). The presence of cavity-nesting species is commonly linked to availability of nesting resources (Cornelius et al., Reference Cornelius, Cockle, Politi, Berkunsky, Sandoval-Vargas and Ojeda2008). We assumed that hexagons with more nesting resources would support a higher number of cavity-nesting species, and therefore we used the number of medium and large cavity-nesting species as a covariate variable.
Table 1 Variables (with their type and description) surveyed in each sampling hexagon in the Beni savannahs, Bolivia (Fig. 1).

Three species of palms (Acrocomia aculeata, Attalea phalerata and Copernicia alba) are commonly used for food and nesting by most local parrot species (Yamashita & de Barros, Reference Yamashita and de Barros1997). Previous findings have suggested that the blue-throated macaw is associated with both A. phalerata and A. aculeata, which are sources of food and nesting sites for this species (Boussekey et al., Reference Boussekey, Morvan and Saint-Pie1997; Yamashita & de Barros, Reference Yamashita and de Barros1997). The presence of these common species of palm trees in the area was used as a covariate associated with resources.
Cattle ranching, which is the main human activity in the region, is an indirect threat to macaws. People regularly burn the savannah to produce grazing land, and fires destroy trees in forest patches. Hunting pressure also appears to be higher near settlements. Local people explore the surroundings of their villages in search of places to hunt. Presence of cattle and distance to villages (from the centre of the hexagon to the centre of the settlement or village) were therefore used as covariates associated with threats.
Trained observers walked 200-m transects in each forest patch. Transects were allocated following the direction of the major dimension of the patch. In most cases one transect was sufficient to cover the entire patch, and we surveyed a maximum of three transects in large patches. For each hexagon we recorded the detection or non-detection of each target bird and palm species, and direct and indirect evidence (faeces and tracks) of cattle. Visual and vocal detections were recorded for all medium and large cavity nesters of the following families: Cathartidae, Falconidae, Tytonidae, Strigidae, Anatidae, Psittacidae, Trogonidae, Ramphastidae and Picidae. We also recorded the number of blue-throated, blue-and-yellow and red-and-green macaws detected, and recorded the duration of each visit, to account for differences in sampling effort.
The abundance of macaw species should reflect a relationship between the availability of resources and the presence of threats. We expected that hexagons where blue-and-yellow and red-and-green macaws were abundant would have a higher probability of occupancy by blue-throated macaws. To model occupancy of the blue-throated macaw we used the sum of the maximum number of detected individuals of blue-and-yellow and red-and-green macaws as a covariate. We applied the same hypothesis for the other two macaw species.
For the blue-throated macaw we defined historical occupation as an additional binary variable for each hexagon. Historical occupation was equal to 1 in hexagons where blue-throated macaws were reported previously (Hesse & Duffield, Reference Hesse and Duffield2000; Berkunsky, Reference Berkunsky2008). A number of studies have monitored the presence of the species (Duffield & Hesse, Reference Duffield and Hesse1997; Hesse & Duffield, Reference Hesse and Duffield2000; Herrera et al., Reference Herrera, Vargas, Sandoval, Perskin and Rendón2007; Berkunsky, Reference Berkunsky2008, Reference Berkunsky2010), and our study area includes the whole known breeding range of the blue-throated macaw.
Occupancy can be defined simply as the proportion of patches in which a species is found (MacKenzie et al., Reference MacKenzie, Nichols, Royle, Pollock and Bailey2005). For each macaw species we estimated both naïve occupancy and detection (i.e. the proportion of visits where a species was detected). As we conducted three surveys in most hexagons (18 of 29) we built detection histories of three visits for each hexagon and each of the three macaw species. There were missing data in the detection histories of 11 hexagons.
We modelled occupancy of each macaw species, using a method that accounts explicitly for detectability, and we used the maximum likelihood approach to estimate occupancy values (MacKenzie et al., Reference MacKenzie, Nichols, Royle, Pollock and Bailey2005). The basis of the model is that there are two stochastic processes occurring that affect whether a species is detected at a site. A site may be either occupied or unoccupied by the species; if it is occupied then at each visit there is some probability of detecting the species.
We evaluated the simplest model for each macaw species, in which both detection and occupancy probabilities were assumed to be constant. We also incorporated site covariates (Table 1) into the model through a logit link function. We evaluated all potential models without interactions and considered only models with 2–4 parameters (including the intercept and probability of detection). The number of parameters was limited by the number of detection histories in the data set (MacKenzie et al., Reference MacKenzie, Nichols, Royle, Pollock and Bailey2005). Two parameters were needed to estimate model intercepts (i.e. occupancy and detection); thus, a maximum of two parameters was available to model covariate variables. We used the corrected Akaike information criterion (AICc) to identify and rank the most parsimonious models. We assessed the fit of our models, using a goodness-of-fit test based on bootstrapping and Pearson's χ2 (MacKenzie & Bailey, Reference MacKenzie and Bailey2004).
For each sampling hexagon we obtained estimates of occupancy (
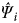 $\hat \Psi _i $
). Using those estimations we applied a bootstrap method to calculate an overall estimate of occupancy (Ψ), with a 95% confidence interval. We computed a conditional probability (Ψ
i
), taking detection history into account. For single-state models, Ψ
cond = 1 for hexagons with at least one detection because species presence was established unambiguously. Finally, we plotted the conditional probability of each hexagon for each species studied. We ran models in PRESENCE v. 3.1 (MacKenzie et al., Reference MacKenzie, Nichols, Royle, Pollock and Bailey2005).
$\hat \Psi _i $
). Using those estimations we applied a bootstrap method to calculate an overall estimate of occupancy (Ψ), with a 95% confidence interval. We computed a conditional probability (Ψ
i
), taking detection history into account. For single-state models, Ψ
cond = 1 for hexagons with at least one detection because species presence was established unambiguously. Finally, we plotted the conditional probability of each hexagon for each species studied. We ran models in PRESENCE v. 3.1 (MacKenzie et al., Reference MacKenzie, Nichols, Royle, Pollock and Bailey2005).
Results
We recorded macaws in 66% of the 29 hexagons surveyed. We counted 25 blue-throated macaws in seven hexagons, 156 blue-and-yellow macaws in 17 hexagons, and 33 red-and-green macaws in 10 hexagons. The naïve occupancy was 24% for blue-throated, 59% for blue-and-yellow, and 34% for red-and-green macaws. The naïve probabilities of detection were 0.73 ± SE 0.12 (n = 5) for blue-throated, 0.61 ± SE 0.08 (n = 13) for blue-and-yellow, and 0.47 ± SE 0.11 (n = 6) for red-and-green macaws. The number of individuals detected per visit per sampling hexagon was 2–9 blue-throated, 1–56 blue-and-yellow, and 1–15 red-and-green macaws. We found C. alba in 28 hexagons (97%), and A. phalerata and A. aculeata in 27 hexagons (93%). We present continuous covariate variables and their statistical descriptions in Table 2.
Table 2 Mean ± SE of continuous covariates used to model the occupancy of large macaws in surveyed hexagons in the Beni savannahs, Bolivia (Fig. 1).

The top models for the three macaw species include the maximum number of individuals and cavity-nester richness (Table 3). The positive values in estimated coefficients of β for the covariates in all models indicate a direct association between the covariates and macaw occupation. Distance-to-village is also included in the top models for the blue-and-yellow and red-and-green macaws (Table 3). Occupation by blue-throated macaws also showed an association with historical occupation.
Table 3 Top occupancy models and coefficients of covariates for blue-throated, blue-and-yellow, and red-and-green macaws in the Beni savannahs, Bolivia (Fig. 1).

1 Covariates: M, maximum number of individuals of other macaw species; D, distance to the nearest village; R, richness of cavity-nesting species; H, historical occupation by the blue-throated macaw
2 Differences in AICc value relative to the top model
3 AICc weight
4 Number of parameters in the model
5 Proportion of sampling units where the species was detected
Mean constant detection probabilities of top models were 0.86 ± SE 0.09 for blue-throated, 0.64 ± SE 0.08 for blue-and-yellow, and 0.48 ± SE 0.13 for red-and-green macaws. Detection probability was not associated with the duration of surveys for any of the species. In the models ranked above the model that assumes constant occupancy and detection probabilities across sites and surveys (i.e. Ψ(.)p(.)) the goodness-of-fit test was not significant (ĉ<2, Table 3), which suggests the model provides an adequate description of the data. In all top-ranked models ψ confidence intervals included the naïve estimation of occupancy.
We estimated conditional occupancy probabilities using the top-ranked model for each species (Fig. 2). Twenty of 29 hexagons surveyed showed a probability of occupancy by macaws of > 0.2. We observed a greater spatial continuity in the distribution of blue-and-yellow and red-and-green macaws compared to the conditional occupancy probabilities of blue-throated macaws.

Fig. 2 Maps of conditional occupancy probability for the blue-throated Ara glaucogularis, blue-and-yellow Ara ararauna and red-and-green macaws Ara chloropterus in the Beni savannahs, Bolivia (Fig. 1). The conditional occupancy probabilities were estimated by the best-ranked model for each species.
The blue-throated macaw was always associated with another species. In hexagons occupied by blue-throated macaws we used the probabilities of detection to estimate the number of macaws. We found a relationship of 5.4 : 1 between numbers of blue-and-yellow and blue-throated macaws detected and a relationship of 1.8 : 1 between numbers of red-and-green and blue-throated macaws detected.
Discussion
Macaws occupied two thirds of the surveyed hexagons and the blue-and-yellow macaw was the most common of the large macaws. The blue-throated macaw occupies small isolated areas and its distribution has not changed since 2000 (Hesse & Duffield, Reference Hesse and Duffield2000). Furthermore, the number of blue-throated macaws recorded (29 individuals) was in the range reported by the census conducted during 1993–1998 in the same area (Hesse & Duffield, Reference Hesse and Duffield2000). In all hexagons occupied by blue-throated macaws there was a high probability of occupancy of both blue-and-yellow and red-and-green macaws. Our results agree with previous records of joint presence of blue-throated and blue-and-yellow macaws (Jordan & Munn, Reference Jordan and Munn1993; Yamashita & de Barros, Reference Yamashita and de Barros1997; Herrera et al., Reference Herrera, Vargas, Sandoval, Perskin and Rendón2007).
Considering the whole area the blue-and-yellow macaw was the most abundant species, outnumbering the red-and-green macaw by 3.5 : 1 and the blue-throated macaw by 8.4 : 1. Blue-and-yellow macaws outnumbered blue-throated macaws at a lower ratio than the 20 : 1 reported by Jordan & Munn (Reference Jordan and Munn1993).
This is the first time that probabilities of detection have been modelled for macaws. We found similar naïve and modelled probabilities of detection for each species. Detection probability was not associated with sampling effort (i.e. with the duration of the surveys). Differences in detection probabilities between species may be associated with flock size and density. Blue-and-yellow macaws, which usually move in larger flocks, showed a higher probability of detection than red-and-green macaws, which are usually found in pairs. The high detection probability of the blue-throated macaw may be a consequence of its tendency to occur together with other macaw species, in areas with higher macaw density. These and other variables should be investigated to determine how they affect detection probability.
Palm tree species and cattle were distributed widely throughout our sampling grid, whereas macaws occupied smaller fractions of the surveyed area. At the landscape scale we were unable to explore whether palm species are reliable indicators of macaw presence. Studies of habitat use at smaller scales may be needed to examine suggested associations between palms and macaws (e.g. A. phalerata and the blue-throated macaw; Hesse & Duffield, Reference Hesse and Duffield2000).
The positive effect of cavity-nester species richness and abundance of other large macaw species on the occupancy of the three species studied here suggests a positive conservation status of occupied sites and a high availability of resources for macaws. This positive relationship is congruent with the general assumption that macaws require relatively pristine habitats (Forshaw, Reference Forshaw1989) and suggests that macaws may be good biological indicators of forest conservation status.
The effect of distance-to-village on macaw occupancy has been used as a valid indirect measure of anthropogenic effects on wild animal populations in several studies (Arroyo-Rodríguez et al., Reference Arroyo-Rodríguez, Mandujano and Benítez-Malvido2008; Margalida et al., Reference Margalida, Donázar, Bustamante, Hernández and Romero-Pujante2008). We found a positive association between distance-to-village and occupancy for the blue-and-yellow and red-and-green macaws; thus, these species could be experiencing higher pressure from hunters. Alternatively, this may be associated with low availability of resources near populated areas. In the case of the blue-throated macaw the absence of a clear effect of distance-to-village could be related to the low number of occupied sites, which may decrease statistical power for detection of a significant effect.
Based on our modelling blue-throated macaw occupancy was best predicted by the maximum number of other macaws detected, whereas blue-and-yellow and red-and-green macaw occupancy were best predicted by distance-to-village and cavity-nester richness. For red-and-green and blue-and-yellow macaws the Akaike differences between the first three models were not significant (ΔAICc < 2).
Apparently the range of the blue-throated macaw is not expanding, as suggested by the significant association between its occupancy and its historical occupation. We only detected blue-throated macaws in seven of the 11 hexagons occupied historically. Surveys conducted during 1992–1998 did not report a probability of detection for the blue-throated macaw (Jordan & Munn, Reference Jordan and Munn1993; Yamashita & de Barros, Reference Yamashita and de Barros1997; Hesse & Duffield, Reference Hesse and Duffield2000) and the number of individuals we detected (29) is similar to the numbers reported by those surveys. If we assume a similar probability of detection for all known surveys, there has not been a significant change in population size. Contrary to predictions based on population viability analysis (Strem & Bouzat, Reference Strem and Bouzat2012) the blue-throated macaw population may not be declining, which is a positive result for conservation, but abundance may not be increasing.
Conditional occupancy probability maps allowed us to identify key areas of importance for the blue-throated macaw and the other two macaw species. An efficient and informative monitoring scheme is needed to design and evaluate conservation initiatives aimed at securing the future of these species, and occupancy methods provide an efficient monitoring tool. The high detectability of macaw species allowed us to estimate occupancy from a low number of visits per sampling site. We consider that the shape and size of our sampling unit (232.7 km2) are appropriate for modelling occupancy of macaws at the landscape level and we recommend this sampling unit for future monitoring initiatives. Three visits to each unit was a sufficient sampling effort. The use of covariates, such as distance to nearest village and richness of cavity-nesting species, allowed us to predict the distribution range of macaw species. We believe that an efficient monitoring protocol could be achieved at a relatively low cost over a large number of sites in the Beni savannahs. Management institutions in developing countries where the monitoring of threatened parrot populations requires innovative approaches could implement similar monitoring protocols, which require a low level of training and expertise, and PRESENCE, required for the analyses, is freely available.
Acknowledgements
We thank P. Alvarez, F. Ávila, J. Casto Perez, C. Cuaino, V. Cuaino, C. Durán, J. Durán, R. Fernandez, L. González, H. Hoyos, R. Ibañez, J.C. Lisboa, A. Nacif, J. Nacif, I. Natusch, H. Parada, J. Rivero, A. Roca, E. Román, F. Román, J. Román, T. Salas, W. Villavicencio, P. Zabala, and R. Zabala for allowing us to work on their properties, and D. Añes, N. Franco, L. Gonzalez, M. Herrera and A.E. Rafael for their kind help and suggestions during fieldwork. This work was supported by The World Parrot Trust, Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET PIP 112-20110-00358), ANPCyT (PICT 2011-0173), Universidad Nacional de La Plata (11/n572, 1/n702) and Universidad Nacional del Centro de la Provincia de Buenos Aires, Rufford Small Grants Foundation, IdeaWild, Birder's Exchange, and CREOI.
Biographical sketches
Igor Berkunsky's primary research interests are the conservation biology of relict and endemic organisms. Rosana Cepeda, Claudia Marinelli and Verónica Simoy are a statistics and mathematics team with many years of experience working in modelling and multivariate analysis. Gonzalo Daniele and Facundo Gandoy are experienced bird surveyors with an interest in conservation. Federico Kacoliris is interested in conservation biology, protected areas and herpetology. José Díaz Luque studies parrots and macaws in Bolivian lowlands, and leads parrot conservation projects in the Neotropics. Rosana Aramburú’s research is focused on Neotropical species of parrots. James Gilardi is interested in the ecology and conservation of parrots.





