


Mountains are high relief habitats that occur across all continents. Their impressive features define landscapes and human societies. These high elevation,Footnote 1 topographically complex habitats provide key ecosystem services (Körner & Ohsawa Reference Körner, Ohsawa, Hassan, Scholes and Ash2006), host high levels of diversity and endemism (Antonelli et al. Reference Antonelli, Kissling and Flantua2018), and are characterized by many specialized and charismatic species, in addition to many generalist species that are distributed across broad elevation gradients (Boyle & Martin Reference Boyle and Martin2015). Mountain regions are highly valued by people in terms of their natural beauty and wildlife, and they are common tourist destinations year-round. However, these regions are under threat from a range of factors (Alba et al. Reference Alba, Kasoar and Chamberlain2022), including climate change (e.g., Gottfried et al. Reference Gottfried, Pauli and Futschik2012; Freeman et al. Reference Freeman, Scholer, Ruiz-Gutierrez and Fitzpatrick2018), changes in livestock management (MacDonald et al. Reference MacDonald, Crabtree and Wiesinger2000; Laiolo et al. Reference Laiolo, Dondero, Ciliento and Rolando2004), increasing pressure from tourism and recreational activities (Rixen & Rolando Reference Rixen and Rolando2013), and exploitation of natural resources, including renewable energy (Svadlenak-Gomez et al. Reference Svadlenak-Gomez, Badura and Kraxner2013), all of which may have implications for mountain bird populations.
In this chapter, we first define our key terms of reference, including what we consider to be ‘mountains’ and ‘mountain birds’. We then summarize the importance of mountains to biodiversity in general and to birds in particular, focussing on key drivers of avian community assembly and variation along elevation gradients encompassing a wide range of habitats (i.e., from relatively low elevations to the highest mountain peaks). Subsequently, we provide an overview of the particular conditions faced by mountain birds at higher elevations, especially at and above the treeline. Finally, we identify some of the key anthropogenic pressures that have shaped high elevation habitats historically. In so doing, we set the scene for the diversity of topics covered in the following chapters.
1.1 Defining a Mountain
What is a mountain? This is a simple question for which there is no simple answer. Several researchers have attempted to define methods and delineate estimates of regional or global mountain areas, typically involving the key characteristics of elevation and steepness of terrain (e.g., Kapos et al. Reference Kapos, Rhind, Edwards, Price, Ravilious, Price and Butt2000; Körner et al. Reference Körner, Paulsen and Spehn2011; Karagulle et al. Reference Karagulle, Frye and Sayre2017; Körner et al. Reference Körner, Jetz and Paulsen2017; Sayre et al. Reference Sayre, Frye and Karagulle2018), although the importance put on specific characteristics varies (Körner et al. Reference Körner, Urbach and Paulsen2021). The definition of Kapos et al. (Reference Kapos, Rhind, Edwards, Price, Ravilious, Price and Butt2000) and Blyth et al. (Reference Blyth, Groombridge, Lysenko, Miles and Newton2002) developed for the United Nations Environmental Programme (UNEP), is based on defining different mountain classes, largely in relation to elevation, the minimum being 300 m to be included as part of a mountain system. This classification (which we term K1 following Sayre et al. Reference Sayre, Frye and Karagulle2018), results in 24.3 per cent of global terrestrial surface being classed as mountainous (Plate 1). This does, however, exclude areas that have many ecological characteristics of mountains. Körner et al. (Reference Körner, Paulsen and Spehn2011) developed a different classification (termed K2) for the Global Mountain Biodiversity Assessment, mostly based on terrain ruggedness, that resulted in the inclusion of a greater area at lower elevations (particularly coastal mountains), but an overall lower area of global mountain systems (12.3 per cent of global terrestrial surface) compared to Kapos et al. (Reference Kapos, Rhind, Edwards, Price, Ravilious, Price and Butt2000). This was due to the exclusion of high elevation plateaus, intermontane valleys and hilly forelands (Plate 2).

Plate 1 Classification of global mountain areas based on Kapos et al. (2000; K1), grouped into seven elevation strata (two strata are combined in the figure). The 7th class (isolated inner basins/plateau < 25 sq. km) was introduced in the 2002 revision of the original 2000 system and is excluded from the calculation of global land surface (g.l.s). Every grid cell above 2500 m was considered ‘mountain’ and all land below 300 m asl was considered as non-mountainous. At intermediate elevations, the classification was based on combinations of elevation, slope and local elevation range (a 7 km radius around each cell). According to this typology, 24.3% of global land surface (g.l.s.) outside Antarctica is classified as ‘mountain’. Note that, unlike K2 and K3 (Plates 2 & 3), K1 mapped relief both for Greenland and Antarctica. Map downloadable at: https://rmgsc.cr.usgs.gov/gme/ (USGS 2021). See Chapter 1 for citations.
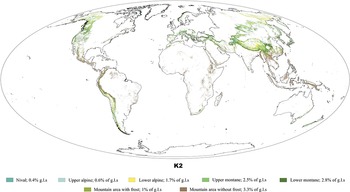
Plate 2 Classification of global mountain areas into seven climatic belts, based on Körner et al. (2011; K2). K2 was developed using ruggedness as the determining factor. According to this classification, 12.3% of the terrestrial surface (outside Antarctica) is mountainous, less than half of the mountain area reported in K1. Körner et al. (2017) attributed the larger area identified as mountainous in K1 to inclusion of high plateaus, intermontane valleys and hilly forelands. Greenland was retained in the mapping for its coastal mountain ranges. GIS raster layer downloadable at: https://ilias.unibe.ch/goto_ilias3_unibe_cat_1000514.html; release 2016 version 3. See Chapter 1 for citations.
Using a higher resolution (250 m versus 1,000 m), Karagulle et al. (Reference Karagulle, Frye and Sayre2017) based their classification (termed K3) for the US Geological Survey on gentle slopes (a virtual mean inclination), ruggedness and profile type (the amount of gently sloping land in upland areas), resulting in an estimate of 30.4 per cent mountain cover of global terrestrial surface (Plate 3). Testolin et al. (Reference Testolin, Attore and Jiménez-Alvaro2020) used an even higher resolution (30 m) to identify a global alpine zone (areas above the treeline) based on unforested areas and modelled estimates of the limits of regional treelines, using the classification of Körner et al. (Reference Körner, Paulsen and Spehn2011) as an initial template. Excluding Arctic and Antarctic mountains, this resulted in an estimated 2.6 per cent of the global terrestrial surface being covered by alpine zones which matches well the alpine areas defined in K2. Plate 4 shows the classification of Testolin et al. (Reference Testolin, Attore and Jiménez-Alvaro2020) superimposed on a composite map of the other three main classifications (K1–K3; Kapos et al. Reference Kapos, Rhind, Edwards, Price, Ravilious, Price and Butt2000; Körner et al. Reference Körner, Paulsen and Spehn2011; Karagulle et al. Reference Karagulle, Frye and Sayre2017) and thus gives an estimate of the maximum extent of mountainous area combining different ‘mountain’ definitions.

Plate 3 Classification of global mountain areas based on Karagulle et al. (2017; K3), grouped into four types based on elevation and isolation (high mountains; scattered high mountains; low mountains; scattered low mountains). The former two classes cover 11.3%, of the Earth’s land surface (excluding Antarctica), while the latter two cover 19.1%, for a total of 30.4% (Sayre et al. 2018). K3 was developed at a finer spatial resolution (250 m) than K1 and K2 and was based on global landforms characterized using three classification parameters: slope, ruggedness and profile. Data downloadable at https://rmgsc.cr.usgs.gov/gme/ (USGS 2021). See Chapter 1 for citations.

Plate 4 Classification of global alpine areas (green) based on Testolin et al. (2020), overlaid onto a map representing all other mountain areas (brown) that combines the other three classifications (K1-3; Plates 1–3). Testolin et al. (2020) characterized alpine biomes according to climate, NDVI and satellite images (Google Earth Engine) at a 30 m spatial resolution to exclude forested areas (all pixels with forest cover > 0% per cent were removed). This approach matches well the alpine areas defined in K2 (Plate 2). See Chapter 1 for citations.
It should be noted that only K1 includes all of Greenland or Antarctica. These areas were excluded from K2 (except for coastal mountains of Greenland) and K3 because their overall aims were not to identify ruggedness per se (a purely topographic view), but to apply the classifications to fields such as forestry (Kapos et al. Reference Kapos, Rhind, Edwards, Price, Ravilious, Price and Butt2000), biodiversity and climatic life zones on earth (Körner et al. Reference Körner, Paulsen and Spehn2011, Reference Körner, Jetz and Paulsen2017), and human populations living in or near mountains (Körner et al. Reference Körner, Urbach and Paulsen2021). We argue that Greenland and Antarctica should be included in future mountain mapping exercises as they hold relevant mountain features (high elevation sites at high latitudes), they host mountain birds (e.g., golden eagle Aquila chysaetos and rock ptarmigan Lagopus muta in Greenland, snow petrel Pagodroma nivea in Antarctica), and many ice-covered sites currently without birds are subject to fast ice-melting processes and are likely to become suitable in the near future.
Which of these methods is preferred depends on the objectives of a given study (Sayre et al. Reference Sayre, Frye and Karagulle2018), but there are situations where clear and objective definitions of mountain areas are needed (Körner et al. Reference Körner, Jetz and Paulsen2017). In this book, we focus on the ecology of the bird species that use these zones for at least a part of their life cycle. Our goals are most in line with the definition of Körner et al. (Reference Körner, Paulsen and Spehn2011), that is, the K2 classification in Plate 2, in that we are primarily concerned with mountain biodiversity quantity and condition, species–habitat relationships and species–climate relationships. However, we do not formally adopt a strict and static definition of a ‘mountain’ which could risk the exclusion of important examples from low mountains (e.g., coastal, or where boreal mountains grade into arctic tundra) or from high elevation plateaus where species are still subject to many of the same constraints (in particular climatic) as mountain birds in steeper terrain. For example, the K2 classification does not include the whole Tibetan Plateau as it does not meet the requirements for terrain ruggedness, but ecologically we would consider this area as mountainous.
Our philosophy mirrors that of Nagy & Grabherr (Reference Nagy and Grabherr2009) in that we are mainly concerned with areas that can be considered part of mountain systems from an ecological, rather than a topographic, point of view. In other words, mountain systems should have significant influences on the ecology of habitats and species due to factors associated with a combination of elevation and topography with respect to the surrounding landscape. For much of this book, we maintain a focus (albeit not exclusively) on areas above the natural elevational limit of continuous forest, where the treeline ecotone forms the lower limit of our main area of interest. Thus, the Testolin et al. (Reference Testolin, Attore and Jiménez-Alvaro2020) classification probably matches that focus most closely. However, it does not include treeline ecotone areas, and in particular those that have been formed at elevations lower than the climatic limit of the treeline, which are also of interest (Chapter 4). It also underestimates the area of alpine zones that have less rock and bare ground, particularly in the tropics (Chapter 3).
1.2 Mountain Biodiversity
Mountainous areas tend to have disproportionately high biodiversity, covering around a quarter of the world’s terrestrial surface (Kapos et al. Reference Kapos, Rhind, Edwards, Price, Ravilious, Price and Butt2000), supporting an estimated one-third of terrestrial biodiversity (Körner Reference Körner2004), and harbouring almost 50 per cent of terrestrial biodiversity hot-spots globally (Myers et al. Reference Myers, Mittermeier, Mittermeier, da Fonseca and Kent2000). Mountain specialists (i.e., those dependent on and restricted to high elevation habitats for key parts of their annual cycle) often show very narrow geographic (and vertical) distributions. The range of individual species may sometimes be restricted to a single mountain or valley (Antonelli et al. Reference Antonelli, Kissling and Flantua2018), or more typically a narrow elevational range, hence mountains are important centres of endemism (Körner et al. Reference Körner, Jetz and Paulsen2017) and speciation (Fjeldså et al. Reference Fjeldså, Bowie and Rahbek2012; Rahbek et al. Reference Rahbek, Borregaard and Antonelli2019). Mountains thus often harbour a greater proportion of threatened species than other habitats (Franzén & Molander Reference Franzén and Molander2012). Biodiversity is also increased by the upshifting of generalist species (those normally occurring over a wide range of elevations) that have lost their low elevation habitat due to anthropogenic impacts, such as farmland birds in France (Archaux Reference Archaux2007).
What drives the high biodiversity in mountains? From an evolutionary perspective, geological heterogeneity and its interaction with historical long-term fluctuations in climate has led to enhanced speciation rates and hence high diversity in mountainous regions (Rahbek et al. Reference Rahbek, Borregaard and Antonelli2019). At a fairly large scale (1° latitude), tetrapod species richness is closely and positively correlated with temperature, precipitation and topographic relief (Antonelli et al. Reference Antonelli, Kissling and Flantua2018), showing the importance of the complexity of mountain environments (evolutionary processes are considered further in Chapter 9). At finer scales, high biodiversity arises over relatively small spatial scales (e.g., one or a few kilometres) as a consequence of the steep terrain and subsequent zonation along elevation gradients (Section 1.2.1).
Species diversity, in particular species richness, varies strongly with elevation. There are competing hypotheses to explain such patterns, and typically these are linked closely to hypotheses explaining trends in relation to latitude. Moist, tropical regions have a more stable year-round climate which, over evolutionary time, may result in greater divergence and niche packing with fine-scale specialization. More fluctuating, higher latitude environments facilitate generalists with broad niches. Analogously, the more fluctuating climatic conditions at higher elevations may contribute to broader niches (Mermillon et al. Reference Mermillon, Jähnig and Sander2022) and decreasing species richness along elevation gradients. However, the latitude gradient shows a fairly constant decrease in species richness towards the poles, whereas there is much more evidence of an intermediate peak in terms of elevation patterns, suggesting that latitudinal and elevational trends are driven, at least in part, by different factors (Rahbek Reference Rahbek1995).
Temperature is in general the most important factor driving biodiversity trends along elevation gradients (Peters et al. Reference Peters, Hemp and Appelhans2016; Laiolo et al. Reference Laiolo, Pato and Obeso2018). Ambient temperature varies with elevation, or more strictly air pressure, in a fairly predictable way termed the adiabatic lapse rate. Typically, there is an approximately 0.6°C decrease for every 100 m increase in elevation, with local variation caused by humidity, wind exposure, cloud cover and other factors (e.g., Dillon et al. Reference Dillon, Frazier and Dudley2006; Colwell et al. Reference Colwell, Brehm, Cardelús, Gilman and Longino2008). Since temperature may constrain the number of organisms that a given area can support, the decrease in temperature at higher elevations may limit the richness of a given community and affect its community structure (White et al. Reference White, Dey, Mohan, Stephens and Price2019). Water availability (precipitation, soil water retention and evaporation) is an additional critical climatic factor (McCain Reference McCain2009; Antonelli et al. Reference Antonelli, Kissling and Flantua2018), influencing, for example, tree formation at high elevation. Primary productivity, which decreases with temperature (and hence elevation) and is also affected by precipitation, is integrated with these two abiotic drivers. High elevations have lower productivity, hence there is insufficient energy to support species rich communities (Newton Reference Newton2020; Schumm et al. Reference Schumm, White, Supriya and Price2020). Indeed, there is evidence that bird species richness is closely correlated with measures of productivity (e.g., Acharya et al. Reference Acharya, Sanders, Vijayan and Chettri2011; Abebe et al. Reference Abebe, Cai and Wale2019). However, these relationships show considerable geographic variation – stability, in situ speciation and accumulation of species over a long time are considered to be more important drivers of species richness within regions with high landscape complexity (Rahbek et al. Reference Rahbek, Borregaard and Antonelli2019).
A range of other hypotheses have been proposed to explain variations in species richness with elevation. Rapoport’s rule states that the latitudinal range size of animals and plants is greater at higher latitudes (Stevens Reference Stevens1989). This has been extended to range sizes in relation to elevation, that is, species of higher elevations show a greater elevational range as they are adapted to a wider range of conditions (Stevens Reference Stevens1992). This results in greater species richness at lower elevations as higher elevation species are more likely to ‘spill down’ to lower elevations (Acharya et al. Reference Acharya, Sanders, Vijayan and Chettri2011). There are also hypotheses that are more related to spatial effects, rather than biological effects per se. For example, some have argued that lower species richness at higher elevations in mountains is due to the species-area relationship and the fact that a ‘typical’ conical-shaped mountain has a greater area at the base than close to the summit (Šekercioğlu et al. Reference Šekercioğlu, Primack and Wormworth2012). An alternative hypothesis is the Mid-Domain Effect (Colwell & Lees Reference Colwell and Lees2000), which proposes that the ranges of species are randomly distributed within a given area, thus more ranges will overlap near the middle of the area than at the edges, resulting in a mid-elevation species richness peak. There has been only limited support for Rapoport’s rule (Gaston et al. Reference Gaston, Blackburn and Spicer1998; Achayra et al. 2011), the species-area relationship (Elsen & Tingley Reference Elsen and Tingley2015) and the Mid-Domain Effect (McCain Reference McCain2009; Reynolds et al. Reference Reynolds, Swanson and Rooney2021) for explaining patterns in species richness along elevation gradients. Environmental drivers (e.g., productivity and climate, in particular water and temperature) are thus likely to be more important (McCain Reference McCain2009), although a range of complex factors interact to produce location-specific patterns (Reynolds et al. Reference Reynolds, Swanson and Rooney2022).
Whilst much research on biodiversity trends along the elevation gradient has focussed on species richness, other studies have instead considered variations in functional diversity, that is, the role of organisms in communities and ecosystems (Petchey & Gaston Reference Petchey and Gaston2006), usually expressed through the analysis of species traits (e.g., diet type, clutch size, foraging niche, migratory strategy). Trends in functional diversity along elevation gradients vary according to latitude. In the tropics, bird communities show a disproportionately high functional diversity in relation to their species richness (i.e., functional overdispersion) in stable lowland habitats, but the opposite pattern (functional clustering) in higher elevation habitats (Jarzyna et al. Reference Jarzyna, Quintero and Jetz2021). However, increasing functional overdispersion is shown in temperate and boreal bird communities at higher elevations (above c. 2,000 m, Martin et al. Reference Martin, Altamirano and de Zwaan2021). Temperate mountains are therefore functionally rich and distinctive ecosystems, despite their overall low species richness. These findings further suggest that higher latitude mountains are disproportionately susceptible to the loss of critical ecological functions because they harbour species assemblages with high functional distinctiveness and low species richness (Jarzyna et al. Reference Jarzyna, Quintero and Jetz2021).
1.2.1 Zonation Along the Elevation Gradient
Mountains are defined by their greater elevation with respect to the surrounding landscape, thus a key characteristic, in particular in relation to biodiversity, is the rapid change in environmental conditions along the elevation gradient – and obviously the steeper the gradient, the more rapidly conditions will change over a given spatial scale. The decrease in temperature with elevation is one of the key environmental factors that affects variation in biotic communities along elevation gradients (see earlier in section 1.2). Additionally, wind speed, air pressure, partial pressure of oxygen and UV radiation vary more-or-less predictably with elevation (Nagy & Grabherr Reference Nagy and Grabherr2009; Chapter 2).
The changing conditions over small spatial scales result in fairly distinct vegetation zones along the elevation gradient that are normally bounded by the upper limit of particular growth forms dictated by the environmental conditions. In a natural state (i.e., with little or minimal human influence), these correspond to the bioclimatic zones listed in Table 1.1. There are two features separating different zones that are of particular relevance to the scope of this book. First, the timberline, which is the upper limit of closed forest. Much of this book is concerned with the area above the timberline (i.e., it forms the lower limit of the bioclimatic zones considered). Second, the treeline, the approximate line that links the highest groups of mature trees, which is often limited by temperature (Körner & Paulsen Reference Körner and Paulsen2004). The treeline typically represents an area of marked change in bird communities (e.g., Altamirano et al. Reference Altamirano, de Zwaan, Ibarra, Wilson and Martin2020; Martin et al. Reference Martin, Altamirano and de Zwaan2021). Given the inconsistencies in the use of these terms to describe vegetation zones and boundaries around the treeline, we discuss them in more detail in Chapter 4.
Table 1.1 Habitat zonation and key divisions between zones along the elevation gradient (based largely on Nagy & Grabherr Reference Nagy and Grabherr2009), as used in this book.
Zone | Description |
|---|---|
| Lowland | Areas not classified as mountain. |
| Montane forest | Closed canopy forest, mature trees – note that transitions may occur between different types of forest within this zone (e.g., subtropical and temperate broad-leaved forest; Acharya et al. Reference Acharya, Sanders, Vijayan and Chettri2011). |
| Timberline | The line where the closed forest ends, marking the transition between montane forest and treeline ecotone. |
| Treeline ecotone | The zone between the timberline and the tree species line. Also sometimes termed the upper subalpine, this is typically characterized by a mosaic of trees, shrubs and meadows. |
| Treeline | The approximate line that links the highest growing groups of mature trees. |
| Tree species line | The maximum possible elevation of tree growth (including seedlings and saplings). |
| Alpine | The treeless area above the tree species line that is dominated by dwarf-shrub communities (sometimes termed lower alpine) and grassland, steppe-like and meadow communities (sometimes termed upper alpine). |
| Snowline | The elevation at which there is permanent snow cover (often considered equal to the upper limit of the alpine zone; Körner Reference Körner2012). |
| Nival | Patchy vegetation, often cushion or rosette plants, within a largely unvegetated landscape (some authors separate nival and subnival zones according to the snowline). |
| Aeolian | Beyond the elevation limit at which vascular plants grow. Wind is important in providing nutrient input and maintaining food chains. |
The zones set out in Table 1.1 are, of course, generalizations – there are many situations where some of them are absent, often due to human activity (see Section 1.5), but also due to ecological or climatic conditions (e.g., the extent of treeline habitat for temperate mountains is often very limited; Nagy & Grabherr Reference Nagy and Grabherr2009). There are also regional or local climatic constraints that may influence zonation such as aspect. In dry climates, the forest may be largely absent (e.g., some central Asian ranges, Potapov Reference Potapov2004; the dry central Andes, Chapter 9). Furthermore, the limit of the alpine zone is influenced by slope exposure. For mountain ranges that are generally orientated from east to west (e.g., Himalayas, European Alps, Pyrenees), the alpine zone is typically lower on northern facing slopes in the northern hemisphere and on southern facing slopes in the southern hemisphere (Nagy & Grabherr Reference Nagy and Grabherr2009). There are oceanic influences on the treeline as well, mediated by precipitation patterns that influence the elevation of the different zones in major mountain chains that are orientated from north to south (e.g., the Andes, Chapter 9) and also mountains on islands. Zonation may also vary according to the geographic position of a particular location within a mountain range, whereby central areas have warmer temperatures and thus higher elevations for any given zone relative to external slopes (the mass elevation effect; Körner Reference Körner2012). Some examples of elevation gradients in mountains from different geographic regions are shown in Figure 1.1. Despite these variations, the definitions in Table 1.1 serve as a useful reference for the typical zonation found along elevation gradients in many mountains.

A. Suntar-Khayata Range, Eastern Siberia, showing gentle elevation gradients resulting in a wide treeline ecotone.

B. Peruvian Andes, with patches of Polylepis woodland.

C. Gradient from montane forest to the alpine zone in the Italian Alps, where grazing has a major impact on vegetation structure and in particular on the elevation of the treeline ecotone.

D. A high elevation lake in the Tantalus Range, British Columbia, Canada, within a diffuse treeline ecotone transitioning into alpine shrubs and a rocky nival zone towards the peak.
Figure 1.1 Examples of the elevation zones that are the main focus of this book.
At very high latitudes, Arctic mountains do not have a treeline as they are beyond the latitudinal limit of tree growth. Indeed, latitude is the main determinant of the elevation of these various zones (Table 1.1); the treeline in tropical mountains can occur at very high elevations (Nagy & Grabherr Reference Nagy and Grabherr2009), whereas in sub-arctic areas at high latitudes, the treeline is at sea-level. Furthermore, this classification does not apply in many areas due to human influence (see Section 1.5).
1.3 Mountain Birds
1.3.1 What Is a Mountain Bird?
Defining a mountain is difficult, so it follows that defining a mountain bird is equally challenging. Objective definitions of mountain birds have been developed based on definitions of mountain areas as outlined above and their overlap with the range maps of the geographical distribution of species. In this way, mountain birds are identified as those with a large proportion of their range in mountain areas (e.g., Scridel et al. Reference Scridel, Brambilla and Martin2018; Lehikoinen et al. Reference Lehikoinen, Brotons and Calladine2019; Alba et al. Reference Alba, Kasoar and Chamberlain2022). However, such range maps are usually restricted to breeding season distributions and thus do not represent the use of mountains by birds throughout the year. The number of species that use mountains may be particularly high. One field study of temperate mountains in the Americas during the breeding season detected 44 to 63 per cent of the regional species pool in western Canada and southern Chile, respectively (Martin et al. Reference Martin, Altamirano and de Zwaan2021). At a continental level and including migrants, Boyle & Martin (Reference Boyle and Martin2015) found that c. 35 per cent of the birds that breed in North America use mountains at some point in their annual life cycle.
In this book, we are interested in how mountain habitats are used by birds. We define a mountain bird in this book as a bird species where at least some populations somewhere in their distribution spend at least one critical stage of their life cycle at or above the elevational limit of continuous forest (i.e., above the timberline). In doing so, we recognize that our knowledge of avian use of mountains is incomplete from a seasonal point of view (as research is biased towards breeding seasons) and from a geographic point of view (as many of the world’s mountain ranges are under-researched – see Section 1.3.2).
1.3.2 Extent of Knowledge of Birds using Alpine Habitats Compared to Other Systems
Given the particular logistical challenges to mountain research, it has been suggested that knowledge of mountain birds is relatively poor compared to other major habitat types (European Environment Agency 2010; Chamberlain et al. Reference Chamberlain, Arlettaz and Caprio2012; Scridel et al. Reference Scridel, Brambilla and Martin2018). For example, nearly one quarter of all alpine breeding species have no nest records or have less than five nests described, in addition to deep data deficiency for most other basic life-history traits (Chapters 2 and 3). A systematic search of published articles in the Web of Science online database between 2011 and 2021 was undertaken to determine the level of relatively recent research on birds in mountains compared to other major habitat types. Of the ten different habitats considered, the number of publications on birds restricted to mountain/alpine habitat (n = 403) was comparable to the total from grasslands and lake/river, and was higher than Arctic/Antarctic and desert habitats (Figure 1.2). Birds associated with forests had the most publications, followed by farmland and urban habitats.

Figure 1.2 The number of research articles on Web of Science (articles referenced in the Science Citation Index, all languages) between 2011 and 2021 according to different search terms based on habitats. The general topic search (TS) term was ‘TS=((bird* OR avian*) AND (HAB1* OR HAB2*) AND (ecology OR conservation))’, where HAB1 and HAB2 represent the search terms on the x-axis (with the exception of desert for which there was only a single habitat term in the search). Only a maximum of two habitat-based terms were used in order to try to produce a more comparable search. A study was only included if the research therein was restricted to a given habitat (e.g., a landscape-level study including both forest and farmland would not have been included).
At first sight, the contention that mountain birds are under-studied compared to birds in other habitats does not seem to be supported. However, considering the number of publications in mountain/alpine habitat according to elevation zone (Table 1.1), it is clear that much research has been carried out exclusively in montane forests (50% of 403 studies), and on elevation gradients across zones (20%), but comparatively little has been conducted specifically in the alpine zone (13%) and even less in the treeline ecotone (8%; Figure 1.3). Hence, our knowledge of the ecology of mountain birds at high elevation does indeed seem to be lower than those in most other major habitat types based on research carried out in the last ten years. Only desert habitats (n = 83; Figure 1.2) had fewer publications than those specifically undertaken either at the treeline or in the alpine zone (n = 84).

Figure 1.3 The number of research articles on mountain birds grouped according to elevation zone (Table 1.1). ‘Open general’ refers to largely treeless habitats that are usually anthropogenic in nature and that occur below the climatic treeline in a given location. Gradient studies encompass more than one elevation zone. N = 403.
A further examination of the ‘mountain’ and ‘alpine’ references was carried out in order to assess geographical biases in research. Most studies had been conducted in Asia and Europe, with somewhat fewer in North and South America (Figure 1.4). At the national level, there were more studies in China (n = 45) than any other country. There were very few studies (<10%) in Africa, Oceania (including Australia) or studies that were carried out in multiple continents. Similarly, a larger proportion of studies was conducted in temperate climatic zones (45%) compared to tropical (35%) or subtropical (13%) zones, and very few were undertaken in boreal or arctic mountains (<3%). These percentages were compared to the percentage of global mountain area in each continent based on the classification of Körner et al. (Reference Körner, Jetz and Paulsen2017). There is a clear bias towards mountain bird research in Europe, which has a far higher percentage of papers relative to the percentage of global mountain area (Figure 1.4), whereas research in Asian mountains is under-represented in proportion to their global mountain coverage. Research in other continents was more-or-less representative of the contribution to global mountain coverage. We should, however, acknowledge that our search terms were in English, hence the search was biased towards publications written in English or with abstracts and titles in English, which may have underestimated representation in some regions.

Figure 1.4 The percentage of research articles on mountain birds grouped according to the continent in which the research took place (dark grey bars) and the percent of global mountain area in each continent (light grey bars) as defined by Körner et al. (Reference Körner, Jetz and Paulsen2017), where Australia has been combined with Oceania. ‘Multiple’ indicates a study that took place on more than one continent. N = 403.
1.3.3 Bird Communities Along the Elevation Gradient
The most common pattern of variation in bird species richness with elevation is that of a mid-elevation peak (Rahbek Reference Rahbek1995; but see Quintero & Jetz Reference Quintero and Jetz2018). However, there is much variation, and the precise pattern may be influenced by a range of factors. Humidity has a marked influence on species richness at large scales. In particular, a low-elevation plateau (i.e., species richness is constant at lower elevations and then decreases after a particular threshold) or decrease in richness with respect to elevation is shown in humid mountains, whereas a mid-elevation peak (i.e., the highest species richness at an intermediate elevation) is more likely in dry mountains (McCain Reference McCain2009). At larger scales, species richness is also correlated with topographic relief, heterogeneity in soils and geological substrate properties (Antonelli et al. Reference Antonelli, Kissling and Flantua2018; Rahbek et al. Reference Rahbek, Borregaard and Antonelli2019), which are in turn related to evolutionary processes (Chapter 9). The influence of methods should also be considered, as estimates of elevational species richness are often affected by variation in sampling effort and sample size (Nogués-Bravo et al. Reference Nogués-Bravo, Araújo, Romdal and Rahbek2008; Quintero & Jetz Reference Quintero and Jetz2018). Raw species richness may give patterns that differ from those where adjustments are made to control for area considered or the geographic range of species. For example, the common intermediate peak (Rahbek Reference Rahbek1995) is driven largely by wide ranging species. When controlling for species range size, the global elevational pattern of species richness is weakly and linearly decreasing (Quintero & Jetz Reference Quintero and Jetz2018).
For plants, temperature and water availability typically determine the position of key vegetation zones (Körner & Paulsen Reference Körner and Paulsen2004) and these vegetation communities are then a key determinant of animal communities. Direct effects of temperature may also limit species distributions in some animals, especially invertebrates (Hodkinson Reference Hodkinson2005), as may the other factors that vary systematically along the elevation gradient (see Section 1.2.1). There is little evidence that these factors directly affect the distribution of many birds (but see Chapter 2). Instead, it seems highly likely that they will impact birds indirectly through effects on plants (Nagy & Grabherr Reference Nagy and Grabherr2009) and invertebrates (Hodkinson Reference Hodkinson2005), influencing both nesting habitat and food supply.
There is evidence that variations in avian diversity along elevation gradients are influenced by vegetation structure (Chapters 3 and 4). For example, Acharya et al. (Reference Acharya, Sanders, Vijayan and Chettri2011), working on an exceptionally long gradient (300–4,700 m) in the Eastern Himalayas, found a peak in species richness at around 2,000 m, close to the transition between subtropical and temperate broad-leaved forest. The richness patterns were best explained by vegetation characteristics (plant species richness, tree basal area, shrub density) and evapotranspiration. Ceresa et al. (Reference Ceresa, Kranebitter, Monrós, Rizzolli and Brambilla2021) found a significant indirect effect of local temperature via vegetation on bird abundance for all 15 species considered along elevation gradients in the Italian Alps, but only generalist forest species showed evidence of direct effects of temperature. Similarly, Ferger et al. (Reference Ferger, Schleuning, Hemp, Howell and Böhning-Gaese2014) found evidence of both direct and indirect effects of climate on bird diversity along a gradient (870–4,550 m) on Mount Kilimanjaro extending to the alpine zone, where both vegetation structure and food resources were important indirect drivers. These studies support an indirect effect of climate (i.e., temperature and moisture) on bird diversity along the elevation gradient mediated through vegetation responses and hence food resources. Indeed, there is evidence from the Himalayas that species richness patterns are associated with declines in arthropod diversity and resource abundance along the elevation gradient (Schumm et al. Reference Schumm, White, Supriya and Price2020).
Interspecific competition is acknowledged as a potentially important driver of animal communities, and it likely plays a role in patterns of functional diversity along elevation gradients (Jarzyna et al. Reference Jarzyna, Quintero and Jetz2021). Although this topic has received relatively little attention, largely due to the difficulties in quantifying interactions among species (McCain & Grytnes Reference McCain and Grytnes2010), Freeman (Reference Freeman2020) indicates high elevation birds may be just as competitive as low elevation birds. Supriya et al. (Reference Supriya, Price and Moreau2020) present intriguing evidence that the mid-elevation peak in bird diversity in the Himalayas is caused by competition with Asian weaver ants Oecophylla smaragdina. Songbirds and ants have a high dietary overlap, and experimental removal of ants increased arthropod prey. At mid-elevations that were beyond the elevation range of the ants, arthropod abundance was positively correlated with bird abundance. This study, carried out in montane forest, shows the potential for competition driving elevational diversity patterns.
1.4 Environmental Challenges for Mountain Birds
Mountains present various environmental challenges to birds. For breeding or year-round resident birds, these are associated with conditions that may be extreme, and are often highly variable and unpredictable. Mountain bird species, or mountain populations (i.e., compared to other populations of the same species at lower elevations), therefore often have specific adaptations to enable them to survive under the particular abiotic and biotic conditions that mountains present. This can include physiological and life-history adaptations as well as some unique behavioural strategies in high elevation specialists (Chapter 2). For example, the glacier finch Idiopsar speculifera nests in glaciers at elevations of up to 5,300 m in the Peruvian Andes (Hardy & Hardy Reference Hardy and Hardy2008; Box 1.1), one of only very few species known to nest on ice (others include emperor penguin Aptenodytes forsteri and white-fronted ground tyrant Muscisaxicola albifrons; Hardy et al. Reference Hardy, Hardy and Castañeda Gil2018). Furthermore, mountains can influence bird migration, both as a physical barrier and as a corridor (Aschwanden et al. Reference Aschwanden, Schmidt and Wichmann2020), and also as a seasonal habitat that enables migratory stopovers for birds migrating both north and south (Boyle & Martin Reference Boyle and Martin2015; de Zwaan et al. Reference de Zwaan, Wilson, Gow and Martin2019).
Box 1.1 Glacier Bird of the High Andes
Few birds are as well adapted to the highest alpine environments as the glacier finch Idiopsar speculifer, a distinctive species living in the High Andes of Peru, Bolivia, and northern Chile. Formerly known as white-winged diuca finch Diuca speculifera, the ecology of this species is strongly associated with glaciers (Figure B1.1). Glacier finches are among the highest nesting passerines in the Western Hemisphere, routinely constructing nests directly on glacier ice at elevations up to 5,300 m (Hardy & Hardy Reference Hardy and Hardy2008). They also roost within glaciers (Hardy et al. Reference Hardy, Hardy and Castañeda Gil2018), and feed on insects on glacier surfaces (Hardy et al. Reference Hardy, Dufour and Oosterbroek2020).

Figure B1.1 A recently fledged glacier finch Idiopsar speculifera (left) being fed by an adult near the nest at Quelccaya Ice Cap, Peru.
The glacier finch is relatively large, slate gray overall with a white throat, a white arc under red eyes, and has dark grey primaries with partially white bases that appear as a white wing patch on perched birds (Fjeldså & Krabbe Reference Fjeldså and Krabbe1990). Its primary foraging habitat is puna grassland, often in wetlands or bogs known as bofedales (Gibbons et al. Reference Gibbons, Jia and Villalba2016), where pairs or small groups move slowly over open ground looking for food. Their distinctive plumage and behaviour render the species easily identifiable, suggesting that its geographic range should be relatively accurately established. However, the 4,000 to 5,500 m elevations where glacier finches are typically found requires acclimatization by humans, hindering detailed studies of their distribution and ecology.
Each day towards dusk the birds move upslope, gathering along glacier margins in preparation for roosting within crevasses and cavities. Typically, pairs arrive at the margin and perch near or on the ice, only entering the actual roost sites when it is dark. This behaviour seems to happen year around, including during the breeding season, and was first described in 1953 (Niethammer Reference Niethammer1953). Roosts are thought to offer protection from low night-time temperatures, snowfall and predation (Hardy et al. Reference Hardy, Hardy and Castañeda Gil2018).
Glacier finches may nest exclusively on glaciers. All of the c. 100 nests observed have been built on glaciers in the Cordillera Vilcanota of southern Peru (Hardy & Hardy 2008; Castañeda Reference Castañeda2015; Hardy et al. Reference Hardy, Hardy and Castañeda Gil2018). Active nests have been found during April in cracks on near-vertical ice faces or deep within crevasses, with clutch sizes of two or three eggs. Nests are a massive (c. 160 g dry mass) collection of grasses and twigs. During one period of extended nest observation, feeding of chicks was evenly shared between the two parents, with both adults arriving at the nest simultaneously and regurgitating food to the chicks before departing with faecal sacs. At this nest, feeding bouts averaged just over one hour (Hardy et al. Reference Hardy, Hardy and Castañeda Gil2018), as most foraging flights were likely greater than 500 m. Does the species nest exclusively on glaciers? Extensive additional research is required; however, their range distribution closely coincides with that of glaciers (e.g., Dussaillant et al. Reference Dussaillant, Berthier and Brun2019).
Glacier finch ecology appears heavily reliant upon the glacierized environment at high elevations of the Andes, for food resources and protection against both predation and the harsh climate. In the strongly seasonal precipitation regime typical of the central Andes, glacier runoff during the dry season sustains bofedales and their associated biodiversity. Even the ice morphology of glaciers appears to be important for roosting and nesting sites; extensive nest site observations at Quelccaya Ice Cap (e.g., Hardy et al. Reference Hardy, Hardy and Castañeda Gil2018) have revealed a strong preference for the protection of crevasses and very steep, fissured margins. Currently, however, Andean glaciers are melting and retreating upslope at an alarming rate (Rabatel et al. Reference Rabatel, Francou and Soruco2013; Dussaillant et al. Reference Dussaillant, Berthier and Brun2019), and the disappearance of the Chacaltaya Glacier in Western Bolivia demonstrates the consequences for this species. Chacaltaya is where glacier finch behaviour on a glacier was first observed (Neithammer Reference Niethammer1953), yet the glacier had disappeared by 2010 (Rabatel et al. Reference Rabatel, Francou and Soruco2013). There are no recent eBird reports of glacier finches from the immediate area. In light of the species’ close association with glaciers, and rapid Andean environmental changes underway, the IUCN Red List Category of ‘Least Concern’ conservation status (BirdLife International 2021) appears inadequate. The future of the glacier finch will likely depend upon the ability of the species to adapt to a very different environment, one with a diminished number of glaciers and less meltwater runoff during the dry season to nourish bofedales.
1.4.1 Weather
Mountain birds, especially at higher latitudes, are typically subject to greater degrees of fluctuations in weather conditions, both at broad (seasonal) and fine (daily) temporal scales (Martin & Wiebe Reference Martin and Wiebe2004). The frequency of extreme weather events increases with elevation (Martin et al. Reference Martin, Wilson and MacDonald2017) and can pose a significant challenge to mountain birds, in particular in terms of its impacts on nesting success. Severe storm events, often involving late season snowfalls, can cause abandonment (e.g., horned lark Eremophila alpestris, Martin et al. Reference Martin, Wilson and MacDonald2017; red-faced warbler Cardellina rubrifrons, Decker & Conway Reference Decker and Conway2009) and direct nestling mortality (e.g., buff-bellied pipit Anthus rubescens, Hendricks & Norment Reference Hendricks and Norment1991; Lapland longspur Calcarius lapponicus, Jehl & Hussel Reference Jehl and Hussell1966). Similarly, prolonged rainfall can be a major cause of nest failure (e.g., in Savannah sparrow Passerculus sandwichensis, Martin et al. Reference Martin, Wilson and MacDonald2017). For high elevation specialists, hot dry weather can also have negative impacts – for example, annual survival in female white-winged snowfinch Montifringilla nivalis is negatively correlated with warm, dry summers (Strinella et al. Reference Strinella, Scridel, Brambilla, Schano and Korner-Nievergelt2020).
Unpredictable weather conditions at high elevation mean that the timing of breeding for many mountain species varies substantially from year to year, for example, up to one month’s difference among years in the clutch initiation dates of ptarmigan Lagopus spp. in North America (Martin & Wiebe Reference Martin and Wiebe2004). Timing of snowmelt in particular will dictate onset of nesting for alpine zone species (e.g., Martin Reference Martin, Johnson and O’Neil2001; de Zwaan et al. Reference de Zwaan, Wilson, Gow and Martin2019; Sander et al. Reference Sander, Chamberlain and Mermillon2021, Reference Sander, Jähnig and Lisovski2023), and probably also for other shrub and tree nesting species. Some migrant species, including long-distance, short-distance and altitudinal migrants, wait at lower elevations for conditions to change (e.g., ring ouzel Turdus torquatus, Barras et al. Reference Barras, Liechiti and Arlettaz2021). In the horned lark, birds that used staging areas relatively close to their breeding grounds for periods of 30 to 60 days on their spring migration had greater productivity, a strategy that may enable individuals to monitor environmental conditions and optimize their arrival date to their breeding site (de Zwaan et al. Reference de Zwaan, Wilson, Gow and Martin2019). Early breeding can be risky due the possibility of nest failure caused by inclement weather. In late-onset years, reproductive success can be severely limited in comparison with birds in lower elevation habitats as the breeding window is short and there may not be the possibility to attempt re-nesting after early nest failures (Martin & Wiebe Reference Martin and Wiebe2004). As a consequence, annual reproductive success may be highly variable. Bollmann & Reyer (Reference Bollmann and Reyer2001) found that predation on water pipit Anthus spinoletta nests did not vary significantly from year to year. However, failure rates due to heavy rainfall and snow varied between 1 and 20 per cent annually. Such effects may have selected for slower life-histories in high elevation birds, both within and across species (physiological and behavioural adaptations to coping with conditions at high elevation are considered in detail in Chapter 2).
Clearly weather, that is, relatively short- or medium-term fluctuations in atmospheric conditions, can have major consequences for mountain bird reproductive success, survival and movement (dispersal or migration). It follows that longer term trends (over many years) in atmospheric conditions, (i.e., climate change), will also have concomitant longer term consequences for bird populations. Indeed, mountains are expected to be affected disproportionately by climate change in many regions (but predictions vary – see Pepin et al. Reference Pepin, Arnone and Gobiet2022), and there is some evidence that this is having impacts on mountain birds (Freeman et al. Reference Freeman, Scholer, Ruiz-Gutierrez and Fitzpatrick2018; Scridel et al. Reference Scridel, Bogliani and Pedrini2017). These impacts are likely to increase for alpine zone species in particular in the future (Chamberlain et al. Reference Chamberlain, Negro, Caprio and Rolando2013). This topic is addressed fully in Chapter 6.
1.4.2 Food Resources
For high latitude mountains, the shorter growing season is likely to be an important constraint that will limit the temporal availability of food and hence affect breeding phenology, migratory behaviour (see Section 1.4.3) and ultimately may influence life-history strategies (e.g., Bears et al. Reference Bears, Martin and White2009; Sandercock et al. Reference Sandercock, Martin and Hannon2005). The melting snow itself may provide an important resource for a range of species (Antor Reference Antor1995; Resano-Mayor et al. Reference Resano-Mayor, Korner-Nievergelt and Vignali2019; Chapter 3). The timing of snowmelt may influence the period for which food is available. For rock ptarmigan Lagopus muta, seasonal variation in the quality of food is important – early snowmelt leads to earlier breeding and higher breeding success. This may be influenced by the quality of plant matter in their diet, whereby earlier snowmelt leads to a longer availability of plants with high digestible protein content (García-González et al. Reference García-González, Aldezabal, Laskurain, Margalida and Novoa2016). For invertebrate feeders, there are numerous examples of lowland passerine species that time their breeding to coincide with peak seasonal abundance of their invertebrate prey (e.g., Both et al. Reference Both, Bouwhuis, Lessells and Visser2006, Reference Both, van Asch, Bijlsma, van den Burg and Visser2009). In a mountain environment, there is little evidence of such clear seasonal peaks. Instead, food availability seems to be dictated by interactions between snowmelt and/or soil moisture (Barras et al. Reference Barras, Marti and Ettlin2020) and vegetation development, and thus can vary substantially from year to year. For ground foragers of the alpine zone such as white-winged snowfinch and water pipit, it seems more likely that birds time their breeding to coincide with the peak in food availability, which may be influenced by vegetation structure (and hence access to prey), rather than prey abundance per se (Brodmann et al. Reference Brodmann, Reyer and Baer1997; Resano-Mayor et al. Reference Resano-Mayor, Korner-Nievergelt and Vignali2019; Chapter 3). In general, however, trophic links between vegetation, invertebrates and birds are poorly studied in mountains.
In tropical mountains, there are relatively more frugivores and nectivores in the bird community. Periods of bad weather that reduce their food supplies or their ability to forage successfully can pose a significant risk as these food sources are low in protein and fat and hence must be consumed frequently and in quantity (Boyle Reference Boyle2018), a contributory factor to altitudinal migration behaviour (see Section 1.4.3). However, tropical conditions may mean a relatively constant food supply is available year-round, hence some species have fewer constraints on timing of breeding. For example, the tropical high-elevation wetlands of the Andes generally do not freeze because of the amount of solar heating during the day (and the thermal capacity of water). This means that some waterbirds can breed all year round (Chapter 9).
1.4.3 Migration
In temperate and boreal mountains, marked seasonality is a major influence on the bird community. Most species that breed above the treeline do not remain all year round. In many temperate and boreal mountains, most species (>75 per cent) are migrants avoiding the harsh winter conditions and thus the winter communities above the forest are made up of very few species. It should be noted that aside from studies in British Columbia (Wilson & Martin Reference Wilson and Martin2005; Boyle & Martin Reference Boyle and Martin2015), there have been few published studies on temperate or boreal mountain bird communities outside the breeding season. However, there are a few studies on the winter ecology of particular mountain species (especially grouse, e.g., Artlettaz et al. Reference Artlettaz, Patthey, Baltic and Leu2007; Bocca et al. Reference Bocca, Caprio, Chamberlain and Rolando2014). There is better understanding of migrants passing through mountains, and in particular passerines, thanks to long-term ringing stations in mountain passes, such as the Col de Bretolet in the Swiss Alps which has operated a ringing scheme continuously since 1958 (Hohl Reference Hohl2019). Data from such sites can be useful in terms of understanding species-specific seasonal migration patterns through mountains in terms of phenology and abundance (Jenni & Kéry Reference Jenni and Kéry2003).
Migrants can be broadly defined into long-distance or short-distance migrants (although this is a simplification as there can be diverse strategies within the same species or indeed the same population; Boyle Reference Boyle2018). Long-distance migrants are generally classified as those species for which the whole population makes seasonal long-distance latitudinal movements, often of 1,000 km or more, to different climatic zones. Short-distance migrants generally migrate away from the breeding area, but they remain in the wider geographic region. Many short-distance migrants in mountains include species that are altitudinal migrants, that is, they make ‘predictable, seasonal movements up and between breeding and wintering ranges within the same geographic region’ (Boyle & Martin Reference Boyle and Martin2015). This is a common strategy in mountain birds (e.g., 30 per cent of North American birds and 65 per cent of Himalayan birds exhibit altitudinal migration; Boyle Reference Boyle2018). This behaviour is facilitated by the marked changes in conditions over small spatial scales, thus the benefits that migrants flying longer distances may accrue by changing latitudes can be achieved with relatively little cost by changing elevation (Boyle Reference Boyle2018). The typical altitudinal migrant is usually perceived as a species that disperses to lower elevations outside the breeding season in response to harsh weather conditions. A good example is the wallcreeper Trichodroma muraria which can often be found at low elevation on older buildings of European towns and cities in the winter (Box 1.2).
Box 1.2 Wallcreeper – An Iconic Mountain Bird
The wallcreeper Tichodroma muraria (Figure B1.2) is probably the most sought-after species by birdwatchers in European Mountains. In part, this is because it is undoubtedly a striking species, but it also has a reputation for being very difficult to find due to its preferred habitat of inaccessible rock faces. In some ways, this species is a typical mountain bird, although it is difficult to classify into any of the zones along the elevation gradient (Table 1.1), as it can breed across a range of elevations, exceptionally even to fairly low levels (e.g., it has been recorded down to 350 m in Switzerland and 500 m in Slovakia; Cramp & Perrins Reference Cramp and Perrins1993; Saniga Reference Saniga2004). However, it is primarily a bird of high mountains and its restricted geographic distribution closely follows the highest Eurasian mountain ranges (Cramp & Perrins Reference Cramp and Perrins1993). Its most important requirement is the existence of sheer rock faces – it can inhabit gorges within mountain forests, although it is more widespread, but never common, on huge cliffs on mountainsides in the alpine zone at higher elevations, for example, 2,000–2,600 m in Switzerland (Keller et al. Reference Keller, Herrando and Voříšek2020), 2,450–3,000 m in Turkey (Cramp & Perrins Reference Cramp and Perrins1993) and above the treeline from 1,700–2,900 m in Catalonia (Aymerich et al. Reference Aymerich, Capdevila, Canut, Roig and Santandreu2012). Aside from rock faces, where it nests and obtains its prey (mostly spiders and small insects), its main requirement seems to be the proximity to water, a typical nest site being a cleft in a rock face above a torrent (Cramp & Perrins Reference Cramp and Perrins1993). It also favours rock faces that have some vegetation rather than being completely bare, and seems to be more common on limestone cliffs (Saniga Reference Saniga2004; Aymerich et al. Reference Aymerich, Capdevila, Canut, Roig and Santandreu2012).

Figure B1.2 Wallcreeper Tichodroma muraria.
The species exhibits altitudinal migration, usually moving to lower elevation valleys, plains and coastal cliffs in winter. It can even be found in towns and cities, in particular on old buildings such as ramparts, bridges and churches. The species is also capable of longer movements of a few hundred kilometres, probably following river valleys. In some records of longer movements, the species has turned up in places far from potential alpine breeding habitat, such as southern England (Dymond et al. Reference Dymond, Fraser and Gantlett1989). It is not known what proportion of the population migrates, either altitudinally or over longer distances, and whether this tendency is more closely associated with certain individuals (e.g., according to age or sex) in the population (Cramp & Perrins Reference Cramp and Perrins1993).
In common with many mountain birds, relatively little is known about this species. The general lack of knowledge of the ecology of mountain bird species or populations compared to their lowland counterparts is largely due to the logistic difficulties of working in mountain environments (Chamberlain et al. Reference Chamberlain, Arlettaz and Caprio2012). Even compared to other mountain species, the wallcreeper presents an extreme challenge due to its habitat. Despite this, there have been some studies on nesting in this species, in particular on nest site characteristics (Saniga Reference Saniga2004) and the nesting period (Cramp & Perrins Reference Cramp and Perrins1993). However, we know very little about the factors that drive reproductive success. Indeed, Alba et al. (Reference Alba, Kasoar and Chamberlain2022) could find no published study that assessed potential impacts of environmental change on this species. For such an iconic species, this is a gap in research that surely needs to be filled.
Whilst tropical mountains are not subject to the same level of seasonal variation in climatic conditions, migration is nonetheless a common strategy, and climate is still a main driver of migratory behaviour, in particular rainfall. Boyle (Reference Boyle2011) found that counts of migrants in lowland forests were positively correlated with montane rainfall, suggesting movements were associated with weather conditions at high elevation. In this case, heavy rain can limit foraging opportunities and hence energy intake, leading to greater propensity for altitudinal migration in wetter years (Boyle Reference Boyle2011). However, in much of the tropical Andes, heavy rainfall occurs commonly at lower elevations. In the alpine zone, it can rain for long periods, but it is usually not very heavy. In many areas, most small birds actually move up high to breed in the rainy season and down-slope migration happens in the dry (winter) season. The situation is similar in African mountains where most birds stay the whole year in the upper montane forest, but some insectivores disappear during the peak of the dry season (J. Fjeldså, pers. comm.).
A further feature of mountains is the marked difference between day and night time temperatures. They may commonly experience frosts during the night and severe summer heat during the day (Nagy & Grabherr Reference Nagy and Grabherr2009). This presents a particular challenge for smaller species with relatively unpredictable food resources, especially in the tropics. Both hummingbirds and sunbirds are known to go into nocturnal torpor, substantially decreasing their metabolic rate as a strategy to cope with cold conditions at night (e.g., Carpenter Reference Carpenter1974; Downs & Brown Reference Downs and Brown2002). Alternatively, altitudinal migration may be undertaken on a diurnal rather than seasonal basis, for example, slender-billed starlings Onychognathus tenuirostris fly from the forests to the alpine zone in east African mountains to feed, but return to the forest at dusk (Young & Evans Reference Young and Evans1993). Whilst normally a forest nester, this species also shows an interesting flexibility in habitat selection, apparently nesting in cliffs in the alpine zone in years of mass flowering of Lobelia deckenii keniensis that produces copious nectar and seeds and hence attracts abundant insects.
1.5 Anthropogenic Impacts
Human land use has been a driver of vegetation dynamics and species distributions in mountains for millennia. Humans have been clearing mountain forests since at least the Neolithic period, initially for fuel collection and burning to maintain more open areas to facilitate hunting (Kessler & Herzog Reference Kessler and Herzog1998). There is evidence that the practice of transhumance, the seasonal movement of livestock and people from the valleys to mountain pastures in the summer, has existed for c. 7,000 years in the European Alps, when Neolithic herdsmen probably burned the forests at the timberline to expand pastures for grazing (Schwörer et al. Reference Schwörer, Colombaroli, Kaltenreider, Rey and Tinner2014). This practice has caused treelines to be much lower than their natural temperature-limited elevation in much of the European Alps, and as a consequence, there is often little or no treeline ecotone. Clearing for urbanization, deforestation and agriculture in the valleys have also impacted lower elevation mountain forests with often only a thin belt of mature forest remaining at mid elevations (Chapter 4). This is taken to further extremes in other mountain ranges where upper montane deforestation is almost complete (e.g., Cantabrian Mountains in Spain).
In both the Andes and the Himalayas, creating land for agriculture was also likely a motivation for clearing high elevation forests (Miehe et al. Reference Miehe, Miehe and Schültz2009; Valencia et al. Reference Valencia, Bush, Coe, Orren and Gosling2018) which has had long-term impacts on the mountain landscape. In the arid mountains of southern Asia, the open dwarf shrubland that makes up the treeline ecotone is likely to be due to forest clearance and grazing, creating conditions from which the forest could not recover after the initial deforestation (Miehe et al. Reference Miehe, Miehe and Schültz2009). In the Andes, while Polylepis woodland is to some extent naturally fragmented, this fire- and disturbance-intolerant genus was much more widespread before the arrival of humans whose activity led to hyper-fragmentation, creating the landscape that is evident today. It is estimated that up to 97 per cent of the original Polylepis woodland has been lost in some areas of the Bolivian Andes due to human activities (Gareca et al. Reference Gareca, Hermy, Fjeldså and Honnay2010).
Changes to the mountain habitat by humans can have positive effects on biodiversity, in particular by creating more diverse habitats, thus generating potential new ecological niche space (Araneda et al. Reference Araneda, Sielfeld, Bonacic and Ibarra2018). For example, creation of open habitats in mountains has provided a refuge for farmland birds whose populations are declining in lowlands due to agricultural intensification (Archaux Reference Archaux2007). This may be the case when there are traditional and sustainable agricultural or pastoral systems in place, as in the European Alps where seasonal grazing creates ecotones that generally support more species than forest or alpine zone habitats (Laiolo et al. Reference Laiolo, Dondero, Ciliento and Rolando2004). In the dry Andes, traditional Indigenous agriculture, including a diverse range of crops fed by irrigation systems, is associated with greater vegetative heterogeneity as well as high avian diversity and density (Araneda et al. Reference Araneda, Sielfeld, Bonacic and Ibarra2018). Similarly, traditional irrigated Bedouin gardens in the South Sinai mountains have a higher bird species richness and density than the surrounding landscape, mainly due to a greater occurrence of migrant species rather than residents (Norfolk et al. Reference Norfolk, Power, Eichhorn and Gilbert2015).
The extent of human impact on the world’s mountains has varied over time. In the European Alps, the maximum rate of forest conversion to open landscapes occurred at the end of the Middle Ages (Gobet et al. Reference Gobet, Tinner, Hochuli, van Leeuwen and Ammann2003). Indeed, the current forest cover is probably the highest it has been for a number of centuries (Bebi et al. Reference Bebi, Siedl and Motta2017). However, that is contrasted with other more recent changes that may have negative implications for biodiversity. The practice of transhumance is becoming economically unviable in many areas and as a consequence of reduced or absent grazing pressure, former ecotone habitats are being encroached by forest (Gerig-Fasel et al. Reference Gehrig-Fasel, Guisan and Zimmermann2007), with subsequent negative effects on the bird diversity of open habitats (Laiolo et al. Reference Laiolo, Dondero, Ciliento and Rolando2004). Conversely, there are other areas where management is becoming more intensive, particularly in terms of management of grasslands for hay cutting, where applications of artificial fertilizers are increasing (Andrey et al. Reference Andrey, Humbert, Pernollet and Arlettaz2014; Assandri et al. Reference Assandri, Bogliani, Pedrini and Brambilla2019). Thus, in the Alps at least, management systems are becoming polarized into either more intensive management, or lack of any management, with likely negative impacts on birds and on biodiversity in general.
Agricultural activities have been the most significant historical anthropogenic factors that have influenced high mountain habitats and their associated bird communities (e.g., Gobet et al. Reference Gobet, Tinner, Hochuli, van Leeuwen and Ammann2003; Miehe et al. Reference Miehe, Miehe and Schültz2009; Schwörer et al. Reference Schwörer, Colombaroli, Kaltenreider, Rey and Tinner2014). Hunting has also undoubtedly been an ever-present factor that continues to influence some mountain bird populations (Chapter 7). However, the last century has brought a number of new pressures caused by human activity. The leisure industry, and in particular skiing, developed in the latter decades of the nineteenth century and has become the main source of income for many temperate mountain communities (Elsasser & Messerli Reference Elsasser and Messerli2001). This has potential consequences for biodiversity in terms of habitat loss and degradation, and direct disturbance. Power generation is also an increasing pressure on mountain environments. The potential for hydropower has been recognized for several decades, and the associated changes to the fluvial environment can have impacts on biodiversity (e.g., Wu et al. Reference Wu, Chen and Xu2019). Wind energy is a more recent development and indeed a more recent potential threat. The risk of collision with wind turbines has been well researched in lowland (especially coastal) habitats, and it seems likely that a similar threat will be posed to raptors in mountain regions (Chapter 7). Finally, evidence is building that climate change poses multiple challenges to mountain birds (Chapter 6). In total, direct effects from multiple stressors and, in some cases in interaction with other drivers, are projected to be a serious threat to high elevation specialist birds in the future (Chapter 8).
In order to protect potentially vulnerable populations of mountain birds, the ecological research community needs to develop strategies to safeguard the future of these species from potential threats through conservation and management actions. This should be underpinned by sound scientific research. Whether the existing knowledge base is sufficient for this relatively under-studied subject area (Section 1.3.2) is a question that underpins the objectives of this book. We aim to identify the key conservation issues and the highest priority conservation actions through our review of the ecology and conservation of birds at high elevations (Chapters 2–5 and 9), including the threats they face now and in the future (Chapters 6–8). Based on the evidence presented in this book, we develop a clear road map to guide research on the ecology and conservation of mountain birds over the next decades (Chapter 10).














