



Introduction
Nutrients in forest litterfall have been linked to patterns of nutrient use at regional and global scales (Vitousek Reference Vitousek1982, Reference Vitousek1984; McGroddy et al. Reference McGroddy, Daufresne and Hedin2004; Parsons et al. Reference Parsons, Congdon, Shoo, Valdez-Ramirez and Williams2014), and changes in foliar and litterfall nutrient concentrations and ratios have been related to climatic drivers such as air temperature and rainfall (Austin & Vitousek Reference Austin and Vitousek1998; Reich & Oleksyn Reference Reich and Oleksyn2004; Santiago et al. Reference Santiago, Schuur and Silvera2005; Wright et al. Reference Wright, Reich, Cornelissen, Falster, Groom, Hikosaka, Lee, Lusk, Niinemets, Oleksyn, Osada, Poorter, Villar, Warton and Westoby2005; Townsend et al. Reference Townsend, Cleveland, Asner and Bustamante2007; Yuan & Chen Reference Yuan and Chen2009; Sardans et al. Reference Sardans, Rivas-Ubach and Peñuelas2011; Luo et al. Reference Luo, Elser, Lü, Wang, Bai, Yan, Wang, Li, Zimmermann, Han, Xu, Li, Wu and Jiang2015; Yue et al. Reference Yue, Fornara, Yangi, Peng, Li, Wu and Peng2017). These studies have mostly used climatic gradients, several focusing on widely distributed plant species and considering mean annual precipitation as the climatic driver. A different approach, to avoid confounding factors, such as changes in plant species composition or soil conditions, may be useful to improve our understanding of the role of rainfall variability and its seasonal distribution on leaf or litterfall nutrients. This involves long-term climate and vegetation data from the same forest site, as well as a perspective that goes beyond mean annual rainfall metrics (Schwartz et al. Reference Schwartz, Lintner, Feng and Powers2020). These studies however are scant. Cusser et al. (Reference Cusser, Helms, Bahlal and Haddad2021) examined the contribution of long-term data in the understanding of ecosystem properties and processes. They concluded that data collected during at least 10 years were needed to properly evaluate processes over a range of climatic extremes or changes that are slow to develop. Furthermore, they hypothesised that biogeochemical processes may take even longer (> 20 years) to show agreement in the direction of significant trends. Several studies have acknowledged the relevance of long-term (> 10 years) ecosystem-scale studies for the understanding of plant stoichiometry (Sardans et al. Reference Sardans, Rivas-Ubach and Peñuelas2012) and the impact of annual rainfall variability on nutrient cycling (Wood et al. Reference Wood, Lawrence and Clark2005). The seasonal distribution of rainfall and its changes among years have also been considered key for the understanding and modelling of ecological processes and of the effects of climate change (Feng et al. Reference Feng, Porporato and Rodríguez-Iturbe2013; White & Hastings Reference White and Hastings2020).
The understanding of nutrient responses in forest leaves and litterfall to changes in precipitation may be especially relevant, given that dry-season precipitation rates have changed globally in recent decades (Murray-Tortarolo et al. Reference Murray-Tortarolo, Jaramillo, Maass, Friedlingstein and Sitch2017), that global change models predict an increasing frequency and magnitude of climatic extreme events as part of the intensification of the global hydrological cycle (Ciais et al. Reference Ciais, Sabine, Bala, Bopp, Brovkin, Canadell, Chhabra, DeFries, Galloway, Heimann, Jones, Le Quéré, Myneni, Piao, Thornton, Stocker, Qin, Plattner, Tignor, Allen, Boschung, Nauels, Xia, Bex and Midgley2013; Wu et al. Reference Wu, Christidis and Stott2013; Knapp et al. Reference Knapp, Hoover, Wilcox, Avolio, Koerner, La Pierre, Loik, Luo, Sala and Smith2015; Allen et al. Reference Allen, Dupuy, Gei, Hulshof, Medvigy, Pizano, Salgado-Negret, Smith, Trierweller, Van Bloem, Waring, Xu and Powers2017), and that decreasing rainfall has been projected in the tropical dry forest (TDF) distribution region (Esperón-Rodríguez et al. Reference Esperón-Rodríguez, Beaumont, Lenoir, Baumgartner, McGowan, Correa-Metrio and Camac2019; Jaramillo & Murray-Tortarolo Reference Jaramillo, Murray-Tortarolo, Busse, Giardina, Morris and Page-Dumroese2019; Xu et al. Reference Xu, McDowell, Fisher, Wei, Sevanto, Christoffersen, Weng and Middleton2019).
In water-limited ecosystems such as TDF, the typical seasonal distribution of rainfall defines periods of resource abundance and scarcity. Since rainfall varies greatly, both seasonally and annually (Murphy & Lugo Reference Murphy and Lugo1986; Miles et al. Reference Miles, Newton, DeFries, Ravilious, May, Blyth, Kapos and Gordon2006; Maass & Burgos Reference Maass, Burgos, Dirzo, Young, Mooney and Ceballos2011; Maass et al. Reference Maass, Ahedo-Hernández, Araiza, Verduzco, Martínez-Yrízar, Jaramillo, Parker, Pascual, García-Méndez and Sarukhán2018), it allows exploring the effects of extreme variation in water availability on forest nutrients and stoichiometry. Our long-term ecosystem research project conducted over the last 40 years in a well-preserved old-growth TDF in Chamela, western Mexico, has shown high inter- and intra-annual rainfall variability, a well-defined onset of the rainy season, but an extremely variable dry-season length (Maass et al. Reference Maass, Ahedo-Hernández, Araiza, Verduzco, Martínez-Yrízar, Jaramillo, Parker, Pascual, García-Méndez and Sarukhán2018). These variations strongly influence the seasonality pattern of litter production (Martínez-Yrízar et al. Reference Martínez-Yrízar, Jaramillo, Maass, Búrquez, Parker, Álvarez-Yépiz, Araiza, Verduzco and Sarukhán2018) and organic matter decomposition (Anaya et al. Reference Anaya, Jaramillo, Martínez-Yrízar and García-Oliva2012). We have also documented how changes in seasonal and interannual water availability affect a variety of soil and plant biogeochemical processes, especially the response of soil and plant N and P (García-Méndez et al. Reference García-Méndez, Maass, Matson and Vitousek1991; Jaramillo & Sanford Reference Jaramillo, Sanford, Bullock, Mooney and Medina1995; Campo et al. Reference Campo, Jaramillo and Maass1998; Rentería et al. Reference Rentería, Jaramillo, Martínez-Yrízar and Pérez-Jiménez2005; Anaya et al. Reference Anaya, García-Oliva and Jaramillo2007, Reference Anaya, Jaramillo, Martínez-Yrízar and García-Oliva2012; Murray-Tortarolo Reference Murray-Tortarolo2011; Rentería & Jaramillo Reference Rentería, Jaramillo, Martínez-Yrízar and Pérez-Jiménez2011). Most of these studies, however, have analysed short-term (≤ 3 years) processes or responses.
Here, we examine litterfall N and P concentrations and their stoichiometry at the forest scale in response to the seasonal distribution of rainfall based on a 20-year data set from our long-term research. We performed this study in the context of a natural experiment (sensu Diamond Reference Diamond1983), which allowed examination of conditions that cannot otherwise be created through experimental manipulation. We tackled three questions: 1) Do rainy-season litterfall N and P concentrations and their stoichiometry change under different levels of rainy-season precipitation? 2) Do dry-season litterfall N and P concentrations and their stoichiometry change in response to variation in the preceding rainy-season precipitation? 3) How do litterfall N and P concentrations from the dry season and their stoichiometry respond to out-of-season precipitation (OSP)?
Our first question addresses the integrated process by which nutrient uptake during leaf production and cycling in litterfall, due to senescence or storm and herbivore damage (Bullock & Solís–Magallanes Reference Bullock and Solís–Magallanes1990), respond to changes in water availability during the rainy season (June–October), which concentrates on average 89% of the long-term annual precipitation (Maass et al. Reference Maass, Ahedo-Hernández, Araiza, Verduzco, Martínez-Yrízar, Jaramillo, Parker, Pascual, García-Méndez and Sarukhán2018). Our second question explores the degree to which dry-season litterfall nutrient concentrations, indicative of nutrient resorption and conservation, vary as a result of changes in the rainy-season precipitation amount in years without OSP inputs. Finally, our third question addresses the extent to which unusual rainfall events during months normally considered part of the dry season (November–May; Maass et al. Reference Maass, Ahedo-Hernández, Araiza, Verduzco, Martínez-Yrízar, Jaramillo, Parker, Pascual, García-Méndez and Sarukhán2018) affect litterfall nutrients. Litterfall pattern in the Chamela TDF is markedly seasonal, with a maximum during the dry season. However, in years with unusual rains (i.e., years with OSP) the amount of dry-season litterfall at the end of the phenological year (March–June) represents a larger fraction of annual production than in years without OSP and similar annual rainfall (Supplementary Figure 1). Yet, the extent to which OSP may also affect nutrient cycling in litterfall is unknown.
Materials and methods
Study area and field design
The study was conducted at the long-term ecological research site of the Chamela-Cuixmala Biosphere Reserve (hereafter, Chamela), on the Pacific coast of Mexico (19°29’N; 105°03’W). The landscape is dominated by low hills (< 300 m elevation) with steep slopes (21° to 41°) (Cotler et al. Reference Cotler, Durán, Siebe, Noguera, Vega Rivera, García-Aldrete and Quesada-Avendaño2002). Average annual temperature is 25.6°C ± 3.4°C (SD; 1980–2015). Monthly mean minimum and maximum temperatures are 16.4°C (March) and 32.6°C (August), respectively. Mean annual rainfall (January–December) is 800 ± 266 SD mm (1983–2018), highly seasonal, with 89% falling between June and October, and September (mean of 212 mm) as the wettest month on average (all data from the Chamela, IBUNAM, and the “Watershed Project”, UNAM, meteorological stations; see also Maass et al. Reference Maass, Ahedo-Hernández, Araiza, Verduzco, Martínez-Yrízar, Jaramillo, Parker, Pascual, García-Méndez and Sarukhán2018). The onset of rains in June triggers the growing season in the Chamela TDF. In contrast to the small variation in mean annual temperature, annual rainfall is highly variable from year to year (Supplementary Figure 2). Only 7% of rainfall events are > 50 mm, but these events, associated with hurricane activity, deliver 42% of the total precipitation (Maass & Burgos Reference Maass, Burgos, Dirzo, Young, Mooney and Ceballos2011). Soils are young, shallow (0.5–1 m depth), predominantly sandy loams, and classified as Typic Ustorthents (USDA system). Soil total N varies between 0.14 and 0.18% and soil PO4- between 9.1 and 18.2 ppm (Mehlich II extractant) in the long-term monitoring forest plots (Solís-Villalpando Reference Solís-Villalpando1993). Soil water content is strongly correlated with monthly rainfall (Galicia et al. Reference Galicia, López-Blanco, Zarco-Arista, Filip and García-Oliva1999). Total N mean concentration in bulk deposition in a year with average rainfall is 1.40 mg L-1 and mean nitrate is 0.08 mg L-1 (Jaramillo unpubl. data). Significant sources of reactive atmospheric N from industries or major agricultural areas are far from our research site.
The dominant vegetation is a highly diverse old-growth TDF (1,149 species of vascular plants), with trees 4–15 m tall and a well-developed understory (Lott & Atkinson Reference Lott, Atkinson, Noguera, Vega Rivera, García-Aldrete and Quesada-Avendaño2002). Most species are deciduous and remain leafless during the dry season each year (Bullock & Solís–Magallanes Reference Bullock and Solís–Magallanes1990; Martínez-Yrízar et al. Reference Martínez-Yrízar, Jaramillo, Maass, Búrquez, Parker, Álvarez-Yépiz, Araiza, Verduzco and Sarukhán2018). Annual litter production varies between 332 ± 27 and 654 ± 18 g m-2 (mean and 1SE; Supplementary Figure 2). Monthly litterfall ranges from a minimum of 24.9 ± 1.7 (August) to a maximum of 52.0 ± 3.7 g m-2 (December; mean ± 1SE; period 1987–2014). However, the month of maximum litterfall in the dry season varies in response to changes in the seasonal distribution of rainfall among years (Martínez-Yrízar unpubl. data).
Five small contiguous watersheds (12–28 ha each) have been gauged for long-term ecological research since 1981 in Chamela (Sarukhán & Maass Reference Sarukhán, Maass and Leff1990; Martínez-Yrízar et al. Reference Martínez-Yrízar, Jaramillo, Maass, Búrquez, Parker, Álvarez-Yépiz, Araiza, Verduzco and Sarukhán2018). One 2,400−m2 permanent plot (80 × 30 m) was established at the middle position in each watershed, with its long axis perpendicular to the stream channel, covering both slopes (generally North- and South-facing slopes). Thus, each slope includes a 1,200 m2 (40 × 30 m) sub-plot, divided into 10 × 10 m quadrats.
Since 1982, litterfall has been collected monthly using 50−cm diameter, 1−m tall conical fibreglass mesh traps (1.4 mm mesh size). Twenty-four litter traps were set up per plot, 12 traps (one per quadrat) on each slope. For this study, we used litterfall samples collected between July 1987 and June 2014 from which a 20-year complete nutrient (N and P) data set was available covering four different decades (1987–1989; 1996–2010; 2013 and 2014; Supplementary Table 1). Due to the occurrence of a category 2 hurricane (Jova) in 2011 which caused significant disturbance to the forest (Álvarez-Yépiz et al. Reference Álvarez-Yépiz, Martínez-Yrízar and Fredericksen2018), this and the following year (2012) were excluded from the analyses. However, two additional post-Jova years (2013 and 2014) were included since canopy cover had fully recovered by 2013 (Parker et al. Reference Parker, Martínez-Yrízar, Álvarez-Yépiz, Maass and Araiza2018). The annual litterfall data correspond to “phenological years”, each starting in July (month of complete leaf production) to June (end of complete leaf-fall) of the following year (Martínez-Yrízar et al. Reference Martínez-Yrízar, Jaramillo, Maass, Búrquez, Parker, Álvarez-Yépiz, Araiza, Verduzco and Sarukhán2018).
Rainfall data and sample chemical analyses
The annual rainfall data correspond to “hydrological years”, each starting in June (onset of the rainy season) and finishing in May of the following year (end of the dry season; Maass et al. Reference Maass, Ahedo-Hernández, Araiza, Verduzco, Martínez-Yrízar, Jaramillo, Parker, Pascual, García-Méndez and Sarukhán2018). The period considered for this study included the wettest (1,506 mm in 2014) and the driest (334 mm in 2005; Supplementary Table 1) years recorded in the Chamela region in our 40-year record (Maass et al. Reference Maass, Ahedo-Hernández, Araiza, Verduzco, Martínez-Yrízar, Jaramillo, Parker, Pascual, García-Méndez and Sarukhán2018; Maass & Ahedo unpubl. data). To address each of our research questions, we used three different data subsets (Supplementary Table 1). For question 1, we used rainfall and litterfall data only from the rainy season of each year (n = 20 years). To answer question 2, we considered only those years in which > 98% of rainfall occurred during the rainy season (n = 9 years) so that the changes in nutrient concentrations in dry-season litterfall (November–June) could be attributed only to the growing-season rainfall. Finally, to address question 3, a subset of 12 years was used fulfilling the following criteria: First, together they represented a precipitation range from wet to dry. Second, they could be grouped according to three levels of total annual rainfall: wet, average, and dry years, defined as follows. “Wet years” had annual precipitation >30% on average (range 950–1131 mm, n = 4 years) than the long-term mean of 795 mm. “Average years” had annual precipitation < 10% above on average (range 808–888 mm, n = 4 years) the long-term mean, and “dry years” had annual precipitation >34% below on average (range 492–561 mm; n = 4 years) the long-term mean. Third, within each year type (i.e., wet, average, and dry), two years registered > 98% of the annual rainfall in the rainy season (June–October) and two registered significant rainfall (9.3–37.4% of the total) in the dry season (November–May). Thus, we attempted to control the total amount of rainfall within each year type to test for the effect of OSP on litterfall nutrients.
The number of litterfall samples for N and P determinations, a total of 24 per plot per month, was reduced to six compound samples per plot by pooling material of four adjacent litter traps. In turn, the monthly litterfall samples were pooled by 4-month periods in each phenological year: the rainfall period (July–October), when the canopy is fully green, the transition period (November–February), when most of the leaves drop, and the dry period (March–June) when the forest becomes leafless. Thus, N and P concentrations were determined in 18 composite litterfall samples per plot per year (6 samples per plot and period × 3 periods), in each of the five watersheds. Litterfall samples, which included leaves, twigs, flowers, and fruits, were ground in a mill to pass a 40-mesh screen. Total N and P concentrations (mg g-1) were determined colorimetrically with an autoanalyser after acid digestion by a semi-Kjeldahl method (Bran-Luebbe Auto Analyser III, Norderstedt, Germany; method No. 696-82W; Technicon Industrial Systems 1977). Total P was determined by the molybdate method after ascorbic acid reduction (Murphy & Riley Reference Murphy and Riley1962). Litterfall N:P ratios were calculated on a mass basis.
Statistical analyses
Litterfall nutrient concentrations were averaged for each period (rainy, transition, and dry) using the five plots as replicates for statistical analyses. To address questions 2 and 3, nutrient concentrations were averaged over the transition and dry periods, hereafter referred to as “dry-season” concentrations. We employed two statistical approaches to analyse the data. To evaluate the impact of rainy-season precipitation on nutrient concentrations and ratios in forest litterfall from the rainy and dry seasons (questions 1 and 2), we used non-parametric Kendall correlation tests on the rainfall and nutrient variables. This was to account for the lack of temporal independence of the samples. To assess the effect of OSP (OSP) on dry-season nutrients and ratios (question 3), we used a bootstrap analysis (i.e., random sampling with replacement method) in which the randomly sampled values of a given season with OSP (e.g., litterfall N, P, or N:P ratio) were subtracted from the values of a season without OSP and divided by the mean value of the season without OSP. We then calculated the mean of the difference, which is the effect size. The procedure was applied to the three year-types independently (dry, average, and wet) and was replicated 1,000 times for each variable (i.e., N and P concentrations and N:P ratios); the sampling size was 20 (2 years with OSP + 2 years without OSP × 5 replicate plots) for each year type (wet, average, and dry). The logic behind our approach was that if nutrient concentrations or ratios were not different in years with and without OSP when randomly subtracted from each other, the resulting distribution should not be different from zero in more than 5% of the cases (Ho). In contrast, a detectable effect of OSP should lead to positive or negative differences and differ from zero in more than 95% of the cases (Ha). The results are presented as boxplots (Supplementary Figure 3). If the whiskers do not touch the zero line, the effect is statistically significant (p < 0.05) for that variable. All statistical tests were performed with R (R Core Team, 2016). Data are shown in the original scale of measurement.
Results
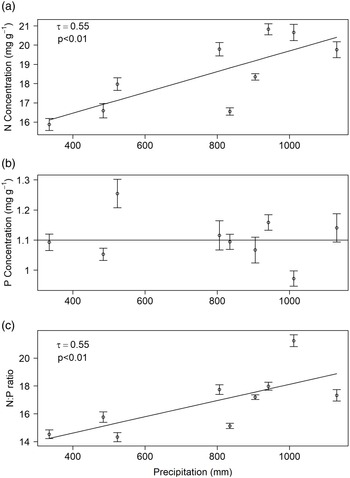
The relationship between both rainy-season rainfall and litterfall N concentrations was positive and statistically significant (p < 0.01; Figure 1a). In contrast, neither litterfall P concentrations nor N:P ratios from the period July–October changed predictably in response to total rainy-season rainfall (Figure 1b, c). Dry-season litterfall N concentrations and N:P ratios both increased with the previous rainy-season rainfall (p < 0.01 and p < 0.01, respectively; Figure 2a, c). In contrast, dry-season P concentrations in litterfall did not change in response to rainy-season rainfall (Figure 2b).

Figure 1. Rainy-season rainfall (June–October) and growing-season (July–October) litterfall N concentrations (a), P concentrations (b), and N:P ratios (c) in the tropical dry forest at Chamela, Jalisco, Mexico. Each value represents the mean of five permanent plots and one standard error. Significant p-value for the regression model is shown; n = 20 years.

Figure 2. Rainy-season rainfall (June–October) and dry-season (November–June) litterfall N concentrations (a), P concentrations (b), and N:P ratios (c) in the tropical dry forest at Chamela, Jalisco, Mexico. Each value represents the mean of five permanent plots and one standard error; significant p-values for the regression models are shown. These models included data from years in which over 98% of annual rainfall occurred only during the rainy season; thus, n = 9 years and differs from Figure 1.
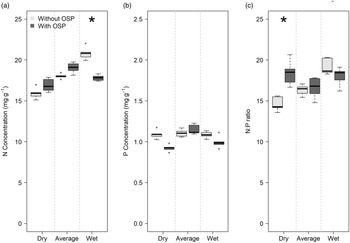
The OSP effect on nutrient concentrations and ratios in dry-season litterfall varied as a function of total annual rainfall and was significant in only two cases. In wet years, OSP significantly reduced N concentrations (p < 0.05) (Figure 3a). In dry years, OSP significantly increased litterfall N:P ratios (p < 0.05; Figure 3c). In average rainfall years, OSP had no significant effect (p > 0.05) on litterfall nutrient concentrations or N:P ratios. P concentrations in litterfall were not affected by OSP, irrespective of total annual rainfall (Figure 3b).

Figure 3. Litterfall N (a) and P (b) concentrations and N:P ratios (c) in dry (492–561 mm), average (808–888 mm), and wet (950-1131 mm) years, with and without out-of-season precipitation (OSP) in the tropical dry forest at Chamela, Jalisco, Mexico; n = 10 permanent plots in each case. Asterisks indicate a significant (p < 0.05) difference in nutrient concentrations or N:P ratios between years with and without OSP after bootstrap analyses. OSP = rainfall between November and May. See Supplementary Table 1 for the specific years, with their corresponding rainfall values, included in this figure.
Discussion
Our study assessed, over a total of 20 years during a 40-year period, the response of nutrient concentrations (N and P) in forest litterfall to changes in the amount and seasonal distribution of rainfall, an indirect measure of soil water availability in the Chamela TDF (Galicia et al. Reference Galicia, López-Blanco, Zarco-Arista, Filip and García-Oliva1999). We partitioned the forest response considering three different conditions, each related to one of our main research questions: a) the rainy growing season, b) the dry season in years with no OSP, and c) the dry season in years with OSP. These are sequentially discussed below.
Response of rainy-season litterfall nutrients to rainy-season rainfall variation
Our results indicated that litterfall N concentrations, but not litterfall P nor the N:P ratio, responded positively to increasing levels of water availability (i.e., rainfall amount). This pattern suggests that as plant N uptake proceeded during the growing season due to leaf growth and expansion, more N was circulated via litterfall as water availability increased with higher rainfall. Several N transformation processes may be involved in such a response. Anaya et al. (Reference Anaya, García-Oliva and Jaramillo2007) found that after the extended dry season, inorganic N and water-soluble organic N accumulate in litter and soil in the Chamela TDF. Also, when dry soil is wetted at the onset of the rainy season short-lived, small N losses (N2O and NO) and rapid microbial mineralisation occur in both standing litter and soil (García-Méndez et al. Reference García-Méndez, Maass, Matson and Vitousek1991; Davidson et al. Reference Davidson, Matson, Vitousek, Riley, Dunkin, García-Méndez and Maass1993; González-Ruiz Reference González-Ruiz1997). Additionally, during the rainy season, net N mineralisation, nitrate concentrations, and net nitrification increase in litter and soil, while soil N immobilisation decreases, and soluble N is leached progressively to the soil (Anaya et al. Reference Anaya, García-Oliva and Jaramillo2007; Montaño et al. Reference Montaño, García-Oliva and Jaramillo2007). Furthermore, our long-term data have shown a significant positive correlation between rainy-season rainfall, specifically rainfall events ≥10 mm, and organic matter decomposition rate in the Chamela TDF (Anaya et al. Reference Anaya, Jaramillo, Martínez-Yrízar and García-Oliva2012). Another important N source during the rainy season is likely related to symbiotic N fixation from the high number of N-fixing legume species abundant in this forest (Durán et al. Reference Durán, Balvanera, Lott, Segura, Pérez-Jiménez, Islas, Franco, Noguera, Vega Rivera, García-Alderete and Quesada-Avendaño2002; Lott & Atkinson Reference Lott, Atkinson, Noguera, Vega Rivera, García-Aldrete and Quesada-Avendaño2002). Nodule abundance and activity are positively linked to soil water content during the rainy season (González-Ruiz et al. Reference González-Ruiz, Jaramillo, Peña-Cabriales and Flores2008). Thus, the evidence suggests an available soil N pool for plant uptake and cycling with increased rainfall during the growing season and supports the finding that species apparently share coherent strategies to cope with changes in the abundance of N forms in their environment in response to changes in rainfall (Houlton et al. Reference Houlton, Sigman, Schuur and Hedin2007).
The lack of a positive response in rainy-season litterfall P concentrations to greater water availability suggests that plant P uptake occurs mostly as a pulse after the onset of rains, when organic and inorganic soil P, accumulated during the dry season, are released (Singh et al. Reference Singh, Raghubanshi, Singh and Srivastava1989; Lodge et al. Reference Lodge, McDowell and McSwiney1994). In the Chamela TDF, soluble and microbial P showed high concentrations in litter and soil at the end of the dry season, and their experimental wetting with the equivalent of a significant rainfall event (e.g., 30 mm), released P from both pools representing up to 83% of the annual aboveground P return from vegetation to the soil (Campo et al. Reference Campo, Jaramillo and Maass1998). In contrast, soluble P pools in rainy-season litter and soil are quite low and rainfall apparently promotes P immobilisation (Campo et al. Reference Campo, Jaramillo and Maass1998). Moreover, other studies in this TDF have shown that labile soil P (inorganic and organic) does not change predictably during the growing season (Montaño Reference Montaño2008) and that repeated experimental soil wetting and drying do not significantly change soil inorganic P concentrations (González-Ruiz Reference González-Ruiz1997). Altogether, these results may explain why litterfall P concentrations did not respond to variation in rainy-season rainfall over the 20-year study period.
Response of dry-season litterfall nutrients to rainy-season rainfall variation
Nutrient concentrations in dry-season litterfall during years with no OSP convey how nutrients are recycled or conserved in response to rainy-season rainfall. Similar to the N concentrations in the growing season litterfall, dry-season litterfall N increased with higher rainy-season rainfall, likely due to reduced N resorption in response to greater N availability (see above). In contrast, the lack of a response in litterfall P concentrations to higher rainy-season rainfall suggests P is conserved despite substantial interannual variation in water availability. Also, the changing N:P ratios suggest stoichiometric flexibility driven by N cycling, with increasing ratios from dry to wetter years. These results contrast with those derived from inter-site comparisons documenting decreased N:P ratios with higher precipitation (Yuan & Chen Reference Yuan and Chen2015). The changes in litter N:P ratios should represent a critical factor in determining the relative importance of different microbial decomposers, as well as the patterns of surface litter decomposition and associated nutrient dynamics (Wood et al. Reference Wood, Lawrence and Clark2005; Güsewell & Gessner Reference Güsewell and Gessner2009) during the dry season.
Response of dry-season litterfall nutrients to OSP
Our analysis revealed that OSP affected N concentrations in litterfall under two conditions. The higher N:P ratios in dry years with OSP were consistent with the increasing N:P ratios in dry-season litterfall in response to rainy-season rainfall, likely due to increased N uptake with greater water availability. In contrast, the lower N concentrations with OSP during wet years suggest that increasing N mineralisation rates and thus litterfall N concentrations from dry to wet years reached a limit, after which higher rainfall with OSP resulted in N losses through leaching from litter. We propose this may represent a threshold to litterfall N concentrations in the Chamela TDF in years without natural disturbances (i.e., hurricanes). The general absence of OSP effects on dry-season litterfall nutrients also suggests that the second leaf flushing of many species and the forest productivity increase in response to OSP, and the species investment in reproduction while leafless (Bullock & Solís–Magallanes Reference Bullock and Solís–Magallanes1990; Martínez-Yrízar unpubl. data) depend on plant nutrients acquired during the growing season and conserved in the dry season.
In this context, we suggest that the small variation in dry-season litterfall P concentrations, mostly between 1.00 and 1.15 mg/g, irrespective of rainfall amount and seasonality (i.e., years with and without OSP), is indicative of forest P conservation. This is consistent with results from nutrient input–output budgets in the same studied watersheds of the Chamela TDF showing a positive balance for dissolved inorganic P and insignificant particulate P losses in a 6-year study (Campo et al. Reference Campo, Maass, Jaramillo, Martínez-Yrízar and Sarukhán2001), thus denoting P retention and biological control of P cycling in the forest. Also, estimates of nutrient residence times at other TDF sites in Chamela (Jaramillo & Sanford Reference Jaramillo, Sanford, Bullock, Mooney and Medina1995) or for several years at the long-term watershed plots (Jaramillo et al. Reference Jaramillo, Martínez-Yrízar, Maass, Nava-Mendoza, Castañeda- Gómez, Ahedo-Hernández, Araiza and Verduzco2018) confirmed the prevalence of P uptake, and thus plant regulation over immobilisation in the Chamela TDF. Plant regulation and conservation are especially relevant for forest P cycling when abundant runoff and high P losses occur associated with extreme storm events such as hurricanes (Jaramillo et al. Reference Jaramillo, Martínez-Yrízar, Maass, Nava-Mendoza, Castañeda- Gómez, Ahedo-Hernández, Araiza and Verduzco2018). Moreover, this suggests that maintenance of forest structure and composition are key for sustainable forest management and conservation.
Our results, from a long-term study at the same forest site, free from potentially confounding factors such as differences in species composition or soils when using climatic gradients, are not consistent with those from other studies. For example, when changes in litterfall nutrients in response to rainfall in tropical forests have been studied along climatic gradients, leaf litter N generally increased, but litter P generally decreased in sites with higher precipitation (Santiago et al. Reference Santiago, Schuur and Silvera2005). In other cases, short-term results have shown that current rainfall increased leaf litter P concentrations but had no effect on N concentrations (Wood et al. Reference Wood, Lawrence and Clark2005).
In conclusion, our study showed litterfall N variation, but P conservation, along steep changes in TDF water availability. Our approach, explicitly incorporating rainfall seasonality in the analysis, allowed establishing that OSP is not a major driver of dry-season nutrient concentrations in forest litterfall, stressing the relevance of nutrient resorption and re-mobilisation as biogeochemical controls of forest functioning in the Chamela TDF. Finally, our study also suggests that N more than P cycling would change in response to either an increased frequency of drought events (Xu et al. Reference Xu, McDowell, Fisher, Wei, Sevanto, Christoffersen, Weng and Middleton2019) or to potential shifts in annual precipitation and/or seasonal droughts in the dry forest of the Chamela-Cuixmala Biosphere Reserve (Esperón-Rodríguez et al. Reference Esperón-Rodríguez, Beaumont, Lenoir, Baumgartner, McGowan, Correa-Metrio and Camac2019) under scenarios of climate change.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0266467423000159
Data and material availability statement (data transparency)
The datasets analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgements
We thank Abel Verduzco and Filiberto Novoa for field-work assistance, Enriquena Bustamante for technical help, and Franciso Mora for statistical advice. We are grateful to the Estación de Biología Chamela, UNAM, for logistic support for more than four decades.
Financial support
Funds for our long-term project were provided by different grants through the years: Consejo Nacional de Ciencia y Tecnología (CONACYT) G27674-N, 83441, 174094; Universidad Nacional Autónoma de México (UNAM) IN206092, IN20811, and most recently by CONACYT 179045 and UNAM IN209117 grants.
Competing interest
The authors declare none.



 Open access
Open access