


Distillers dried grains with solubles (DDGS) is a by-product produced by dry-grind ethanol plants and has become a valuable low-cost feed ingredient for replacement of maize, soyabean meal and inorganic phosphate in diets of pigs. Sows are ideal candidates for diets containing DDGS because of its moderately high fibre content. DDGS contains approximately three times the concentrations of fat, protein, fibre and minerals compared with maize( Reference Spiehs, Whitney and Shurson 1 ), and many studies have indicated that supplementation of DDGS to sows during gestation and lactation had no negative effect on the sow and litter performance( Reference Song, Baidoo and Shurson 2 – Reference Wang, Su and Shi 4 ). However, maize DDGS contains approximately 10 % maize oil( Reference Song and Shurson 5 ), and the lipids in DDGS predominantly comprise PUFA, which are prone to preoxidation( Reference Weber and Kerr 6 ). During drying, DDGS are heated at relatively high temperatures, potentially leading to the formation of secondary lipid oxidation products( Reference Song and Shurson 5 ). Therefore, DDGS might be prone to significant lipid peroxidation. However, most studies have ignored the potential risk of oxidative stress upon the DDGS to the diet of sows and offspring.
l-Carnitine plays a key role in cell metabolism by regulating the mitochondrial transport of long-chain free fatty acids and the generation of ATP by β-oxidation( Reference Rebouche 7 ). Evidence indicated that dietary l-carnitine is beneficial to sows, by enhancing reproduction performance and milk production( Reference Ramanau, Kluge and Eder 8 – Reference Musser, Goodband and Tokach 10 ). Furthermore, other studies have demonstrated that l-carnitine supplementation may enhance gastrointestinal protective( Reference Dokmeci, Akpolat and Aydogdu 11 , Reference Akpolat, Gulle and Topcu-Tarladacalisir 12 ), antioxidant( Reference Kalaiselvi and Chinnakkannu 13 ), anti-inflammatory( Reference Ribas, Vargas and Wajner 14 ) and radioprotective( Reference Akpolat, Topcu-Tarladacalisir and Dokmeci 15 ) properties.
In our previous study, we have found that the DDGS diet revealed negative effects on weights of piglets at birth and weaning, but it did not influence the number of total piglets, piglets born alive and piglets at weaning, and sows treated with l-carnitine had higher birth weight and weaning weight regardless of sows’ dietary treatments (soyabean meal v. DDGS). Furthermore, l-carnitine supplementation of sows causes also an increase of l-carnitine concentrations in milk and the carnitine status of their offspring. In addition, there are no published data on the effects of maternal dietary treatments and l-carnitine in sows on intestinal function of their offspring. Therefore, the aim of the present study is to evaluate the effect of dietary treatment and l-carnitine in gestating and lactating sows on the intestinal barrier functions of their offspring and the possible interaction between dietary treatment and l-carnitine on the intestinal barrier functions including antioxidant capacity, concentrations of cytokines, intestinal microbiota and tight junction mRNA expression of their offspring.
Methods
The DDGS used for the experiment was provided by Jilin Alcohol Company Limited. The DDGS was analysed for crude protein (295·7 g/kg), crude fat (100·4 g/kg), crude fibre (71·3 g/kg) and P (6·1 g/kg). The soyabean meal was analysed for crude protein (430·4 g/kg), crude fat (19·2 g/kg), crude fibre (58·8 g/kg) and P (6·3 g/kg). l-Carnitine (Carniking™, 50 % purity) was obtained from Lonza Inc. The contents of crude protein, crude fat, crude fibre and P were assayed in our laboratory according to the procedures described by the AOAC (Association of Official Analytical Chemists)( 16 ).
Animals and experimental design
This study was performed in strict accordance with recommendations of the National Research Council Guide(62), and all of the animal experimental procedures were approved by the Ethical and Animal Welfare Committee of Heilongjiang Province, China (2008).
A total of 120 cross-bred (Landrace×Large White crossed with Duroc boar) multiparity gestation sows with an average parity of 4·2 (sem 0·2) and their offspring were used in the study. At service, sows (230 (sem 4·2) kg) were blocked by body weight (BW) and service date and randomly assigned to four groups of thirty each. The experiment was designed as 2×2 factorial with two dietary treatments (soyabean meal v. DDGS) and two l-carnitine levels (0 v. 100 mg/kg in gestating diets and 0 v. 200 mg/kg in lactating diets). The experimental treatments were as follows – from 1 to 90 d of gestation: (1) 160 g/kg soyabean meal; (2) 250 g/kg DDGS; (3) 160 g/kg soyabean meal+100 mg/kg l-carnitine; and (4) 250 g/kg DDGS+100 mg/kg l-carnitine; from 90 d of gestation to 21 d of lactation: (1) 278 g/kg soyabean meal; (2) 400 g/kg DDGS; (3) 278 g/kg soyabean meal+200 mg/kg l-carnitine; and (4) 400 g/kg DDGS+200 mg/kg l-carnitine. The composition and analysis of the dietary treatments (Tables 1 and 2) were compared using validated analytical methods (National Standards of the People’s Republic of China, GB/T 19540-2004).
Table 1 Composition of experiment diets for sows (as fed basis)

DDGS, maize distillers dried grains with soluble; CP, crude protein.
* Provided the following per kg of diet: 3·6 mg of vitamin A, 62·5 μg of vitamin D3, 65·8 mg of vitamin E, 3·6 mg of vitamin K, 1·7 mg of thiamin, 7·0 mg of riboflavin, 1·6 mg of pyridoxine, 0·3 mg of vitamin B12, 35 mg of niacin, 15 mg of pantothenic acid, 3·5 mg of folic acid, 0·4 mg of biotin, 0·33 mg of antioxidant, 120 mg of Fe as FeSO4·H2O, 21·25 mg of Cu as CuSO4·5H2O, 39·75 mg of Mn as MnSO4·H2O, 101·25 mg of Zn as ZnSO4·H2O, 0·3 mg of Se as Na2SeO3, 0·4 mg of I as KI.
† Analysed values.
‡ Calculated values according to the tables of feed composition and nutritive values in China (Xiong et al.(63)).
Table 2 Composition of diets for offspring (as fed basis)

CP, crude protein.
* Provided the following per kg of diet: 3·3 mg of vitamin A, 68·9 μg of vitamin D3, 66 mg of vitamin E, 4·4 mg of vitamin K, 1·1 mg of thiamin, 9·9 mg of riboflavin, 2·2 mg of pyridoxine, 0·1 mg of vitamin B12, 55 mg of niacin, 55 mg of pantothenic acid, 1·7 mg of folic acid, 0·4 mg of biotin, 0·33 mg of antioxidant, 105 mg of Fe as FeSO4·H2O, 6 mg of Cu as CuSO4·5H2O, 28·62 mg of Mn as MnSO4·H2O, 110·06 mg of Zn as ZnSO4·H2O, 0·3 mg of Se as Na2SeO3, 0·14 mg of I as KI.
† Provided the following per kg of diet: 2·6 mg of vitamin A, 41·3 μg of vitamin D3, 33 mg of vitamin E, 3·3 mg of vitamin K, 1·1 mg of thiamin, 5·5 mg of riboflavin, 2·0 mg of pyridoxine, 0·03 mg of vitamin B12, 33 mg of niacin, 22 mg of pantothenic acid, 1·0 mg of folic acid, 0·1 mg of biotin, 0·33 mg of antioxidant, 114 mg of Fe as FeSO4·H2O, 6 mg of Cu as CuSO4·5H2O, 14·31 mg of Mn as MnSO4·H2O, 70 mg of Zn as ZnSO4·H2O, 0·3 mg of Se as Na2SeO3, 0·14 mg of I as KI.
‡ Provided the following per kg of diet: 2·6 mg of vitamin A, 41·3 μg of vitamin D3, 33 mg of vitamin E, 3·3 mg of vitamin K, 1·1 mg of thiamin, 5·5 mg of riboflavin, 2·0 mg of pyridoxine, 0·03 mg of vitamin B12, 33 mg of niacin, 22 mg of pantothenic acid, 1·0 mg of folic acid, 0·1 mg of biotin, 0·33 mg of antioxidant, 90 mg of Fe as FeSO4·H2O, 5 mg of Cu as CuSO4·5H2O, 14·31 mg of Mn as MnSO4·H2O, 69 mg of Zn as ZnSO4·H2O, 0·3 mg of Se as Na2SeO3, 0·14 mg of I as KI.
§ Analysed values.
The sows were kept in single crates until day 30 of pregnancy. From day 30 to day 110 of pregnancy, the sows were kept in groups of six in pens that had fully slatted floors. On day 110 of pregnancy, the sows were moved to the farrowing accommodation where they were housed in single farrowing pens. The farrowing room temperature was maintained at approximately 18–20°C. Within 2 d after farrowing, all piglets were earmarked according to the ear marks of their mothers. Piglets removed from sows and piglets given to sows were selected on the basis of their BW. At day 3, piglets received an Fe injection. At day 7, male piglets were castrated. The piglets were weaned on day 21. Then, 400 piglets (100 piglets/treatment) with similar weight were moved to post-weaning rooms set at 27°C for the first 3 d and then gradually reduced to 22°C. Piglets were housed in pens with woven-wire flooring with ten piglets per pen. After 55 d, a total of 120 offspring with similar weight (regardless of sex) were moved to finishing barns with total concrete slat flooring. In all, thirty piglets were selected from each sow treatment. There were six pens per treatment and five pigs per pen. All offspring were fed the same diets (Table 2) and provided free access to water and the standard diet throughout the experiment. All of the experimental diets were formulated to meet or exceed the recommended requirements of the National Research Council(62). The basal concentration of l-carnitine was <5 mg/kg in all feed mixtures used.
Sample collection
Piglets were selected from each treatment as follows: twenty-four similar average-weight piglets (at birth mean 1·4 (sem 0·01) kg and weaning mean 5·6 (sem 0·06) kg) regardless of sex were selected (six piglets per treatment) and slaughtered by an intra-arterial injection of pentobarbital (200 mg/kg) after general anaesthesia. The twenty-four slaughtered newborn piglets were selected from twenty-four sows (six sows per treatment, one piglet per sow) without fostered piglets. The selected newborn piglets were removed from their mothers immediately after birth and without sucking colostrum to exclude the effects of nutrients in the colostrum on their intestinal barrier functions. The slaughtered weaning piglets were selected from their mothers (six sows per treatment, one piglet per sow), which corresponded to the mothers of slaughtered newborn piglets, respectively. In all, twenty-four growing–finishing pigs with a similar weight of 110 (sem 4·6) kg and regardless of sex from the four treatments (six pens per treatment, a pig per pen) were transported to the abattoir for slaughter. The twenty-four slaughtered growing–finishing pigs were born of sows (one piglet per sow), which corresponded to the mothers of slaughtered newborn and weaning piglets according to their ear marks. The abdomen was immediately opened to remove the entire small and large intestines. The small intestine and the caecum were carefully dissected from the mesentery, emptied and sampled. On the basis of previous reports on piglets and rats( Reference Cetinkaya, Bulbuloglu and Kantarceken 35 , Reference Strzalkowski, Godlewski and Hallay 36 ), we designated the middle part of the jejunum. One part of mucosal scrapings from the jejunum was prepared frozen in liquid N2 and stored at −80°C for further analyses of cytokines and tight junction protein mRNA expressions. The other part of the mucosal scrapings from the jejunum was sealed into pockets and preserved at −20°C until used for the evaluation of antioxidant status. The contents (approximately 3 g) of the caecum, which was divided into three equally long segments, were collected from the middle section and immediately frozen in liquid N2 and stored at −80°C until analysis for microbial DNA.
Analytical methods
Approximately 1 g of mucosal scrapings was homogenised after being suspended in 9 ml of PBS. After centrifugation at 3000 rpm for 15 min, the supernatant was taken for the measurement of the total superoxide dismutase (SOD), reduced glutathione (GSH), malondialdehyde (MDA) and cytokine concentrations. All antioxidant parameters were measured with the method of assay kits supplied by Nanjing Jiancheng Bioengineering Institute( Reference Mao, Lv and Yu 17 , Reference Che, Xuan and Hu 18 ). Mucosal levels of IL-1β, IL-6, IL-12 and TNF-α were measured using ELISA kits in accordance with the manufacturers’ instructions (Experimental Equipment Co. Ltd).
Total DNA extraction from the middle section of the caecum samples was extracted using the TIANamp Stool DNA Kit (Tiangen Biotech, Ltd) according to the instructions of the manufacturer. The quantity and quality of DNA was assessed using a NanoPhotometer-P330 spectrophotometer (Implen GmbH). Primers for total eubacteria, Lactobacillus spp., Enterobacteriaceae, bifidobacteria spp. and Clostridium cluster XIVa were designed according to previously published data( Reference Lee, Zo and Kim 19 – Reference Matsuki, Watanabe and Fujimoto 24 ). The primers (Table 3) were used for the estimation of select bacterial groups by quantitative real-time PCR using the ABI 7500 Real-Time PCR system (Applied Biosystems Limited). For bacterial groups, real-time PCR used the SYBR® Premix Ex TaqTM II system (TaKaRa® Bio Catalog). The PCR was run in a volume of 20 μl, with 10 μl of SYBR® Premix Ex TaqTM, 0·8 μl of each primer (0·4 μm), 0·4 μl of ROX Reference Dye II (TaKaRa), 6·0 μl of double-distilled water and 2 μl of DNA template. The thermal cycling conditions involved an initial denaturation step at 95°C for 5 min, followed by forty cycles of 95°C for 15 s and 72°C for 30 s. Dissociation analyses of the PCR products were carried out to confirm the specificity of the resulting PCR products. Quantification was performed in duplicate, and the mean values were calculated. The results were reported as log10 16S ribosomal DNA gene copies per gram fresh matter.
Table 3 Primers used for real-time PCR to detect bacterial numbers

Total RNA was extracted from approximately 100 mg of frozen jejunal tissues using the reagent box of the Total RNA Kit (E.Z.N.A.®, Omega Bio-tek, Inc.), according to the manufacturer’s instructions. The concentration of RNA was measured by using a spectrophotometer, and the purity was ascertained by the A260:A280 ratio. The total RNA from each sample was RT complementary DNA (cDNA) using the Prime Script® RT reagent Kit (TaKaRa® Bio Catalog), according to the manufacturer’s instructions, and the cDNA was used for RT-PCR. The SYBR Green I RT-PCR Kit (TaKaRa® Bio Catalog) was used to measure the mRNA expression of tight junctions (claudin 1, zonula occludens-1 (ZO-1) and occludin) relative to the expression of the β-actin endogenous control. Specific primers were designed using the Primer Express® software (PE Applied Biosystems) and were synthesised by Sangon Biological Engineering Co. Ltd. Primer sequence data are presented in Table 4. For analyses on an ABI PRISM 7500 SDS thermal cycler (Applied Biosystems), the reactions were performed with 2·0 μl of first-strand cDNA and 0·4 μm of sense and anti-sense primers in a final volume of 20 μl. The samples were centrifuged briefly and run on the PCR machine using the default fast programme (one cycle at 95°C for 30 s, forty cycles of 95°C for 5 s and 60°C for 34 s). The relative gene expression levels were determined using the
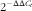 $${\rm 2}^{{{\minus}\Delta \Delta C_{t} }} $$
method(
Reference Livak and Schmittgen
25
).
$${\rm 2}^{{{\minus}\Delta \Delta C_{t} }} $$
method(
Reference Livak and Schmittgen
25
).
Table 4 Sequence of primers for tight junction genes

ZO-1, zonula occludens-1.
Statistical analysis
Bacterial gene copies were transformed (log10) before statistical analysis. Data were analysed by ANOVA using the general linear model procedure of SPSS 17.0 (IBM-SPSS Inc.). The results were presented by mean values with their standard errors. The statistical model included the main effects of maternal diet, l-carnitine and piglet age, the interactions of maternal diet×l-carnitine, maternal diet×piglet age, l-carnitine×piglet age and maternal diet×l-carnitine×piglet age. Differences were considered significant at P<0·05.
Results
There were no significant interaction effects between dietary treatment and l-carnitine in gestating and lactating sows on the antioxidant capacity, concentrations of cytokines and the tight junction protein expression of jejunal mucosa in their offspring.
Antioxidant capacity
Table 5 shows that dietary supplementation with l-carnitine in gestating and lactating sows increased the SOD activity of the jejunal mucosa in newborn piglets and weaning piglets on day 21 (P<0·05). Dietary supplementation with l-carnitine in gestating and lactating sows decreased the concentration of MDA of the jejunal mucosa in newborn and weaning piglets on day 21 (P<0·01). There were no differences in the antioxidant capacity of jejunal mucosa in growing–finishing pigs. Age significantly influenced the antioxidant capacity of jejunal muscosa in their offspring (P<0·001), but there were no interaction effects between age and treatments (P>0·05) (Table 5).
Table 5 Effects of l-carnitine and/or maize distillers dried grains with solubles diets (DDGS) in diets of gestating and lactating sows on antioxidant capacity in the jejunal mucosa of their offspring

SBM, soyabean meal diets; D, dietary treatments; L, l-carnitine; A, age; D×A, dietary treatments×age interaction; L×A, l-carnitine×age interaction; D×L×A, dietary treatments×l-carnitine×age interaction; MDA, malondialdehyde; SOD, total superoxide dismutase; GSH, reduced glutathione.
Concentrations of cytokines
Table 7 shows that dietary supplementation with l-carnitine in gestating and lactating sows decreased the concentrations of IL-1β, IL-12 and TNF-α in the jejunal mucosa of newborn piglets (P<0·05) and decreased the concentrations of IL-6 (P<0·05) and TNF-α (P<0·01) in the jejunal mucosa of weaning piglets on day 21. There were no differences in the concentrations of cytokine in the jejunal mucosa in growing–finishing pigs. Age significantly influenced the concentrations of cytokines in their offspring (P<0·001), and there were significant interaction effects between l-carnitine and age in the concentration of IL-6, IL-12 and TNF-α (P<0·05). However, there were no interactions between dietary treatments and age, and also there were no interaction effects among the dietary treatments, l-carnitine and age (P>0·05) (Table 6).
Table 6 Effects of l-carnitine and/or maize distillers dried grains with solubles diets (DDGS) in diets of gestating and lactating sows on the concentrations of cytokines in the jejunal mucosa of their offspring

SBM, soyabean meal diets; D, dietary treatments; L, l-carnitine; A, age; D×A, dietary treatments×age interaction; L×A, l-carnitine×age interaction; D×L×A, dietary treatments×l-carnitine×age interaction; MDA, malondialdehyde; SOD, total superoxide dismutase.
Bacterial numbers
There was an interaction between dietary treatment and l-carnitine on the number of total eubacteria in the digesta of the caecum in weaning piglets on day 21 (P=0·046). The number of total eubacteria in weaning piglets was significantly increased by l-carnitine only in the soyabean meal diet (P<0·05), but there was no significant effect of l-carnitine in the DDGS diet (P>0·05). Table 8 shows that dietary supplementation with l-carnitine increased the numbers of Lactobacillus spp. and bifidobacteria spp. in the digesta of the caecum in weaning piglets on day 21 (P<0·05). There were no differences in bacterial numbers in the digesta of the caecum in growing–finishing pigs. Age significantly influenced the number of Lactobacillus spp., Enterobacteriaceae and bifidobacteria spp. in their offspring (P<0·01), but there were no effects in the number of total eubacteria and Clostridium cluster XIVa (P>0·05). There was no interaction between treatments and age (P>0·05) (Table 7).
Table 7 Effects of l-carnitine and/or maize distillers dried grains with solubles diets (DDGS) in diets of gestating and lactating sows on bacterial numbers in the caecal digesta of their offspringFootnote *

SBM, soyabean meal diets; D, dietary treatments; L, l-carnitine; A, age; D×A, dietary treatments×age interaction; L×A, l-carnitine×age interaction; D×L×A, dietary treatments×l-carnitine×age interaction.
* The results were reported as log10 16S ribosomal DNA gene copies/g fresh matter.
Gene expression of tight junction proteins
The results of the expression of tight junction proteins showed that dietary supplementation with l-carnitine in gestating and lactating sows increased the expression of ZO-1 mRNA in the jejunal mucosa of newborn piglets (P<0·01), and increased the expression of ZO-1 and occludin mRNA in the jejunal mucosa of weaning piglets on day 21 (P<0·05). There were no differences in the expression of tight junction proteins of the jejunal mucosa in growing–finishing pigs. Age influenced the expression of ZO-1 mRNA (P<0·05) in their offspring, but there were no effects in the expression of claudin 1 and occludin mRNA (P>0·05). In addition, there was no interaction between treatments and age (P>0·05) (Table 8).
Table 8 Effects of l-carnitine and/or maize distillers dried grains with solubles diets (DDGS) in diets of gestating and lactating sows on tight junction mRNA expression in the jejunal mucosa of their offspring

SBM, soyabean meal diets; D, dietary treatments; L, l-carnitine; A, age; D×A, dietary treatments×age interaction; L×A, l-carnitine×age interaction; D×L×A, dietary treatments×l-carnitine×age interaction; ZO-1, zonula occludens-1.
Discussion
The use of l-carnitine in the nutrition of pigs is increasingly discussed. There are several studies that found beneficial effects of l-carnitine supplementation on sows( Reference Musser, Goodband and Tokach 10 , Reference Eder, Ramanau and Kluge 26 – Reference Ramanau, Kluge and Spilke 28 ). A few studies observed beneficial effects of l-carnitine supplementation on the growth performance or body composition of growing pigs( Reference James, Tokach and Goodband 29 – Reference Birkenfeld, Ramanau and Kluge 31 ). However, the present study is the first one that investigates the effect of dietary treatment and l-carnitine in gestating and lactating sows on intestinal barrier functions of their offspring.
As is known, there is a strong correlation among oxidative stress, inflammation, mucosal integrity and intestinal permeability. The activity of SOD is known to serve protective function for the elimination of reactive free radicals, and thus it represents an important antioxidant defence in nearly all cells exposed to oxygen( Reference Cheung, Zheng and Li 32 ). GSH forms an important part of the non-enzymatic antioxidants. Similar to other sulfhydryl-containing products, GSH also has regulatory and protective roles in the body. It establishes the defence line of the body against tissue injury because of chemicals through its ROS scavenger, cell viability and membrane-stabilizing effects( Reference Szabo 33 ). MDA is an end product of free-radical chain reaction and lipid peroxidation( Reference Liu, Han and Huang 34 ), so it is frequently used in the measurement of lipid peroxide levels, and it provides a good correlation with the degree of lipid peroxidation( Reference Cetinkaya, Bulbuloglu and Kantarceken 35 ). In the present study, the results show that the activity of SOD was significantly increased in the newborn piglets and weaning piglets of sows supplemented with l-carnitine. The variational trend of MDA concentrations was opposite to that of SOD activity in piglets of gestating and lactating sows supplemented with l-carnitine. It has been clearly demonstrated that l-carnitine not only has an important role in the metabolism of lipids but is also a potent antioxidant( Reference Kalaiselvi and Chinnakkannu 13 ). The findings may indicate that the abundance of antioxidants supplemented with mother’s milk might reduce oxygen free radicals in piglets( Reference Strzalkowski, Godlewski and Hallay 36 ). The results of the present study are consistent with previous conclusions that l-carnitine administration reduced the concentration of MDA, prevented the depletion of GSH levels and increased SOD activity( Reference Dokmeci, Akpolat and Aydogdu 11 , Reference Akpolat, Gulle and Topcu-Tarladacalisir 12 , Reference Cetinkaya, Bulbuloglu and Kantarceken 35 ).
The gastrointestinal tract functions as a major immunological organ, as it must maintain tolerance to commensal and dietary antigens while remaining responsive to pathogenic stimuli. The intestinal immunity is closely related to other intestinal functions. Previous studies have suggested that a decrease in the inflammatory cytokine gene expression may promote a more stable ecosystem that may prevent the proliferation of some particular bacterial groups( Reference Walsh, Sweeney and O’shea 37 ). The over-production of pro-inflammatory cytokines has an adverse effect on intestinal mucosal integrity( Reference Liu, Huang and Hou 38 ). Recent studies have indicated that most pro-inflammatory cytokines, such as TNF-α, interferon-γ, IL-1β and IL-6, induce a pathologic opening of the intestinal tight junction barrier and increase intestinal epithelial permeability( Reference Al-Sadi, Boivin and Ma 39 ). In the present study, dietary supplementation with l-carnitine in gestating and lactating sows decreased mucosal IL-1β, IL-12 and TNF-α concentrations in newborn piglets and decreased mucosal IL-6 and TNF-α concentrations in weaning piglets. l-Carnitine has been shown to affect humoral and cellular immune responses in mice( Reference Athanassakis, Mouratidou and Sakka 40 ). It suppresses TNF-α and IL-12 release from human monocytes in a glucocorticoid-like manner( Reference Alesci, De Martino and Mirani 41 ). l-Carnitine has also been shown to significantly decrease TNF-α and DNA damage( Reference Abdelrazik, Sharma and Mahfouz 42 ). Liu et al.( Reference Liu, Wu and Zhang 43 ) showed that the oral administration of l-carnitine reduced the serum concentrations of TNF-α or IL-6. The findings in the present study might be because an increase of the l-carnitine concentration in the fetus and milk( Reference Ramanau, Kluge and Eder 8 , Reference Lahjouji, Elimrani and Lafond 44 ) as a result of maternal supplementation with l-carnitine decreased mucosal cytokine concentrations in newborn and weaning piglets.
From simple to complex and being gradually stable with the increase of age, balanced intestinal flora is important for growth and health of piglets( Reference Mathew, Upchurch and Chattin 45 ). The most important beneficial effect of the indigenous microflora is creating a barrier to make it more difficult for exogenous pathogenic bacteria to colonise the gastrointestinal tract and cause disease, a phenomenon known as colonisation resistance( Reference Brassart and Schiffrin 46 ). The intestinal microbiota also has essential functions in host metabolism and in directing immune system development( Reference Brown, Decoffe and Molcan 47 ). The pre-treatment of animals with bifidobacteria reduced pro-inflammatory cytokine release, the concentrations of endotoxin, intestinal epithelial cell apoptosis and disruption of tight junctions, resulting in recovered microbiota and mucosal integrity( Reference Wang, Zhang and Zuo 48 ). In vitro, the probiotic Lactobacillus has also been shown to up-regulate occludin and cingulin gene mRNA levels of Caco-2 cells, suggesting that bacteria could affect the intestinal barrier integrity by regulating the gene expression of the tight junction proteins( Reference Anderson, Cookson and McNabb 49 ). Indigenous Clostridium-dependent induction of Tregs may be required for maintaining immune homoeostasis in mice and humans( Reference Atarashi, Tanoue and Shima 50 ). The present study was aimed at detecting whether the l-carnitine in DDGS-based diet of gestating and lactating sows can affect the form of the intestinal flora in their offspring. As the results show, dietary supplementation with l-carnitine increased the number of Lactobacillus spp. and bifidobacteria spp. in the digesta of the caecum in weaning piglets. As we know, diet has a direct influence on the intestinal microbes of both composition and vitality of this community( Reference Bauer, Williams and Smidt 51 ). Interestingly, it has been found that breastfed European children have a microbiota depleted of Bacteroidetes and enriched in Enterobacteriaceae compared with rural breastfed African children, which the authors attributed to difference in maternal diet( Reference De Filippo, Cavalieri and Paola 52 ). It has been suggested that maternal diet can influence the intestinal microbiota of the offspring. Furthermore, sows supplemented with l-carnitine have more milk production and piglets born to sows supplemented with l-carnitine are able to suckle for longer, which enables them to obtain more milk than piglets born to control sows( Reference Birkenfeld, Doberenz and Kluge 53 ). Previous studies showed that sufficient feed intake during weaning encourages the proliferation of beneficial bacteria, thereby contributing to improved gut health( Reference Rist, Eklund and Bauer 54 ). Interestingly, the number of total eubacteria in weaning piglets was significantly increased by l-carnitine only in soyabean meal diet, but there was no significant effect of l-carnitine in the DDGS-based diet. Further research needs to be carried out to explore the effects of l-carnitine in the DDGS-based diet on intestinal microbiota.
Tight junction proteins (claudin 1, ZO-1 and occludin) have a crucial role in intestinal barrier integrity and permeability by sealing the paracellular space between epithelial cells, thus preventing the paracellular diffusion of intestinal bacteria and other antigens across the epithelium( Reference Ulluwishewa, Anderson and McNabb 55 ). Wang et al.( Reference Wang, Zhang and Zuo 48 ) showed that the pre-treatment of animals with bifidobacteria reduced the disruption of tight junctions. Zhong et al. ( Reference Zhong, Cai and Cai 56 ) showed that the beneficial effect on the expression of occludin mRNA has been considered to be related to the greater number of lactobacilli and bifidobacteria, in agreement with the results in the present study. Pro-inflammatory cytokines have been shown to down-regulate gene expression of tight junction proteins( Reference Al-Sadi, Boivin and Ma 39 , Reference Chen, Mao and He 57 ) through the dysregulation of tight junction proteins, resulting in the increase in intestinal permeability( Reference Berin, Yang and Ciok 58 ). This is part of the reason why higher expression of ZO-1 and occludin mRNA were found in piglets born to sows supplemented with l-carnitine in the present study. Furthermore, the high concentration of l-carnitine in the offspring of sows supplemented with l-carnitine may exert an antioxidant property, decreasing lipid peroxidation and reactive oxygen species formation, possibly resulting in the increased expression of ZO-1 and occludin mRNA. The present study indicates that supplementation with l-carnitine in sows could enhance intestinal barrier integrity by up-regulating the tight junction protein mRNA of newborn and weaning piglets.
DDGS contains approximately three times the amount of most nutrients, such as protein, oil, fibre and minerals, compared with maize( Reference Spiehs, Whitney and Shurson 1 ). Therefore, DDGS is viewed as a potential feed resource to replace a portion of energy (maize) and protein (soyabean meal) ingredients( Reference Wang, Wang and Shi 59 ). In the past, DDGS had not been used extensively in non-ruminant animal diets because of high fibre content. However, recently, many researchers thought it would be more suitable for sows because of its high content of protein and fat and moderately high fibre content( Reference Li, Baidoo and Li 60 ). However, there was no study to evaluate the effect of maternal DDGS on intestinal barrier functions of offspring. The present study did not find effects of the DDGS diets of gestating and lactating sows on the intestinal barrier functions of their offspring.
The results of the present study indicate that dietary supplementation with l-carnitine in gestating and lactating sows had positive effects on intestinal barrier functions of newborn and weaning piglets. Of note, the positive effects of maternal l-carnitine supplementation did not last to the period of growing–finishing pigs. Within the growing process of offspring, these differences of maternal effects might be subsequently mitigated by the postnatal environment( Reference Kunz and King 61 ).
Acknowledgements
The authors wish to acknowledgement the assistance of Yinguo Li, Chunliang Zhao and Quansheng Liu.
This work was supported by the National Basic Research Program (grant number 2012CB124703), the China Agriculture Research System (grant number CARS-36), the National Science and Technology Support Program (grant number 2013BAD10B03) and the Program for Universities in Heilongjiang province (grant number 2012TD003, 11551z003). The funders had no role in the design, analysis or writing of this manuscript.
All authors participated in the development of the study concept and design; B. W., S. N. and A. S. were responsible for the execution of the study; B. W., S. N. and Z. Q. were involved in the animal experiments, analysis and data collection; Q. M. and Z. C. were responsible for the statistical analysis; B. W. and S. N. wrote the draft of the manuscript; Q. M., A. S. and Z. C. reviewed and revised the manuscript.
None of the authors has any conflicts of interest to declare.








