


Introduction
Carcass composition and growth traits are of great economic importance in commercial pig production. Hence, these traits receive considerable interest in pig breeding programmes. With the advent of molecular genetic tools, selection may be further improved based on mutations in candidate genes or quantitative trait loci (QTL), e.g. the IGF-2 mutation in pigs affecting the percentage lean meat (Van Laere et al., Reference Van Laere, Nguyen, Braunschweig, Nezer, Collette, Moreau, Archibald, Haley, Buys, Tally, Andersson, Georges and Andersson2003). However, improvement of performance traits may be associated with undesirable effects on meat quality, as has been clearly evidenced in the case of the RYR1 and the RN mutation (De Smet et al., Reference De Smet, Pauwels, Vervaeke, Demeyer, De Bie, Eeckhout and Casteels1995; Sellier, Reference Sellier1998; Hamilton et al., Reference Hamilton, Ellis, Miller, McKeith and Parret2000). Therefore, the search for new gene-based selection tools aiming at improving performance traits has to be accompanied by an evaluation of the correlated effects on pork quality.
In the pig, an association between a MC4R Asp298Asn missense mutation and fatness, increased daily gain and increased feed intake was first demonstrated by Kim et al. (Reference Kim, Larsen, Short, Plastow and Rothschild2000). This seems a promising tool to improve daily gain, especially in the Belgian pig industry where the slow-growing Piétrain breed is very frequently used as the sire breed. The MC4R receptor is expressed in the appetite-regulating region of the brain where it regulates feed intake and energy balance. The Asp298Asn mutation occurs within a highly conserved region where Asp298 is required for normal MC4R signalling. The Asn298 variant decreases MC4R signalling without differences in the ligand binding (Kim et al., Reference Kim, Reecy, Hsu, Anderson and Rothschild2004). The effect of the Asp298Asn substitution in MC4R is not reported to be uniform, but rather a blend of effects on average daily gain (ADG), feed intake, lean growth, carcass fat and carcass length (Kim et al., Reference Kim, Larsen, Short, Plastow and Rothschild2000; Park et al., Reference Park, Carlborg, Marklund and Andersson2002; Hernández-Sánchez et al., Reference Hernández-Sánchez, Visscher, Plastow and Haley2003; Stachowiak et al., Reference Stachowiak, Szydlowski, Obarzanek-Fojt and Switonski2005; Bruun et al., Reference Bruun, Jørgensen, Nielsen, Andersson and Fredholm2006). The aim of the current study was to evaluate the effect of the MC4R missense mutation on carcass and meat quality traits in a commercial four-way cross population. Moreover, independent of this mutation, the relationships between ADG and carcass and meat quality traits were evaluated in this population. It is well known that some meat quality traits may be negatively affected in very lean pigs, although the effects of intense selection for ADG and leanness on meat quality traits depend on the specific meat quality trait and are not unequivocal in literature (Sellier, Reference Sellier1998).
Material and methods
Animals and experimental set-up
Animals were the offspring from a commercial four-way cross. Hybrid sows originated from a Landrace × Large White intercross mainly selected for number of pigs born alive, number of pigs weaned and longevity. Sires originated from a line based on Large White and Piétrain, mainly selected for ADG and carcass lean meat content. The distribution among the five sires and gender is given in Table 1. The animals were all born on the same farm and were transferred after 2 months to the finishing farm. The pigs were fed a commercial diet ad libitum and were slaughtered at a target live weight of 110 kg. A total of 1155 animals were slaughtered in the abattoir of Ghent University on 25 different slaughter days (approximately 45 pigs per slaughter day). Pigs were bled following manual electrical stunning (240 V). Before slaughter, feed was withdrawn overnight and the pigs were allowed to rest after transportation for a minimum of 1 h. Based on the time of slaughter, the time of feed withdrawal and the arrival time of the pigs at the slaughterhouse, starvation time and lairage time were calculated in hours and decimal minutes. Starvation time ranged between 4.3 and 22.0 h (average 16.6) and the lairage time varied between 0.4 and 15.8 h (average 4.1).
Table 1 Distribution of the pigs (n = 1155) according to the genotype for the MC4R genotype, gender and sire
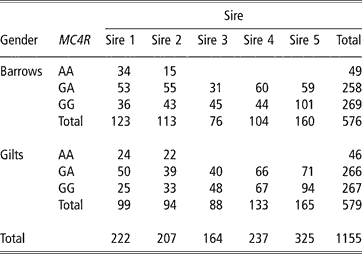
Abbreviation is: MC4R = melanocortin-4 receptor genotype.
Animal and carcass quality data
Live weight (kg) and warm carcass weight (kg) were recorded before stunning and at the end of the slaughter line, respectively. Dressing yield (%) was calculated based on cold carcass weight (98% of the warm carcass weight). ADG (g/day) was calculated as the live weight divided by the number of days from birth to slaughter. Average daily carcass lean meat gain (ADLMG, g/day) was calculated as the cold carcass weight multiplied by the percentage lean meat estimated with the CGM device (Capteur Gras/Maigre, Sydel, Lorient, France), divided by the number of days from birth to slaughter.
Instrumental carcass grading was performed using the CGM device (Sydel, France) and the SKGII apparatus (Eurocontroll Belgium, Vilvoorde, Belgium). With the CGM apparatus, fat and muscle thickness (mm) were measured at the level of 3rd to 4th last rib using light reflection and the carcass lean meat percentage (LMP) was estimated by using the regression formulae as approved for use in Belgian abattoirs. The SKGII device combines four physical measurements (fat thickness (mm), ham width (mm), loin width (mm) and ham angle (deg.)) to estimate the type index as a measure for conformation (Branscheid and Sack, Reference Branscheid and Sack1988). The type index has to be interpreted as lower values indicating more muscular conformation. In addition, fat depth (mm) was manually measured on the cutting edge at rib 1, rib 7 and lumbar vertebra 3. The number of ribs and vertebra and carcass length (cm), assessed as the distance from the posterior edge of the symphysis pubis to the cranial edge of the first rib, were recorded as well.
Meat quality measurements
The pH was measured 40 min and 24 h after slaughtering in the longissimus at the level of the 3rd–4th last rib (pH1L and pH2L) and in the ham (semimembranosus) (pH1H and pH2H) using a Knick Portamess 654 pH meter (Knick, Berlin, Germany) equipped with an Mettler Toledo LoT406-M6-DXK-S7/25 electrode (Urdorf, Switzerland). Temperature of the carcass was also measured at 40 min after slaughtering in the longissimus (Temp1L) (°C). Conductivity (μS) was measured at 24 h post mortem in the longissimus (PQM2L) and semimembranosus (PQM2 H) using a pork quality meter (PQM; Tecpro GmbH, Aichach, Germany). The average of the measurements of pH, PQM and temperature on both carcass sides for each animal was used for further calculations.
One day post mortem (24 h), a piece of the longissimus from the left carcass side at the level of the last thoracic vertebra and first two lumbar vertebra was removed (approximate thickness 7 cm), deboned and sliced. One slice was used for colour measurements using the six-point Japanese pork colour scale (FHK Co., Tokyo, Japan) (1 = PSE, 6 = DFD) and by determining the CIELAB colour co-ordinates (L*, a* and b*) in quadruplicate with a HunterLab MiniScan device after 30 min blooming time (D65 light source, 10° standard observer, 45°/0° geometry, 1-in light surface, white standard; Hunter, Reston, USA). Drip loss was assessed in duplicate as the uptake of fluid (mg) using the filter paper method of Kauffman et al. (Reference Kauffman, Eikelenboom, van der Wal, Merkus and Zaar1996). Thaw loss (%) was determined as the difference between the initial weight of a sample before frozen storage at −18°C in a vacuum bag and the weight after overnight thawing at 4°C, divided by the initial weight. Cooking loss (%) was determined as the difference of the sample weight before and after cooking the sample in a open plastic bag in water at 70°C for 40 min followed by cooling in cold running tap water for 15 min, divided by the initial weight. Tenderness was evaluated by shear force determination on the cooked samples. Shear force (N) and total work (area under the curve, N·mm) were measured on cylindrical cores (∅1.27 cm) taken parallel and sheared perpendicular to the longitudinal orientation of the muscle fibres, using a Lloyd TA 500 Texture Analyser (Lloyd Instruments, Fareham, Hampshire, UK) equipped with a triangular Warner–Bratzler shear device. A mean of eight measurements were used for further analysis. Total work was divided into myofribrillar work (N·mm) and collagen work (N·mm) as an estimate of the respective contributions of myofibrillar and collagen component to toughness. Myofibrillar work is the area under the first peak and the area difference between total work and myofibrillar work is designated as collagen work (Claeys et al., Reference Claeys, De Smet, Balcaen and Demeyer2000). Finally, intramuscular fat (IMF) content was measured on thawed longissimus homogenates with near infrared spectroscopy, calibrated with IMF values determined by the Soxhlet method on 184 samples (mean 1.42%, s.d. 0.62%, min. 0.37%, max. 4.43%). The calibration and measurements were performed as described by Meulemans et al. (2002) for technological and organoleptical properties of pork. The best predictive model was selected based on the lowest standard error of cross-validation (SECV) and the highest determination coefficient of cross-validation (R 2cv), which were 0.27 and 0.80, respectively. This model was then used to predict the ether content of all other samples.
Statistics
The effect of the MC4R mutation was analysed with a univariate GLM with MC4R genotype (MC4R), gender (G) and sire (S) as fixed factors and slaughter day as random factor for the meat quality traits. The effect of starvation and lairage time were evaluated in preliminary analyses, but these factors did not appear to yield additional information above slaughter day, and therefore were not included in the model. No effects of the two-way interactions were found in the preliminary analyses, and therefore these terms were also excluded from the final model. When the MC4R mutation had a significant effect, means were compared with the Tukey’s comparison of mean test. The Pearson’s correlation coefficients between the residuals of all univariate models were used to examine the relationships between the carcass and meat quality traits after correction for the fixed effects. All data were analysed using SPSS 12.0 for windows (Statistical Packages for the Social Sciences, 2003).
Results
Growth and carcass parameters
Pigs were slaughtered at an average (±s.d.) live weight of 109 ± 8 kg. The allelic frequencies of the MC4R mutation were 0.69 for the 298Asp variant or G-allele and 0.31 for the 298Asn variant or A-allele. Live weight was not different between the MC4R genotypes (Table 2). However, the days to reach this slaughter weight and thus also ADG differed significantly according to the MC4R mutation. Based on the least-square means, animals with the AA genotype grew approximately 25 g/day faster than their GG genotype counter mates. Animals heterozygous for the MC4R mutation were intermediate and different from the homozygous genotypes (P < 0.05).
Table 2 Least square means ± s.e. for growth and carcass quality traits by the MC4R genotype

Abbreviations are: s.e. = standard error, MC4R = melanocortin-4 receptor genotype, GLM = general linear model, G = gender and S = sire effect, ADG = average daily gain, ADLMG = average daily carcass lean meat growth.
a,b,cMeans for the effect of line lacking a common superscript letter differ (P < 0.05).
†ADG was calculated as the live weight divided by the number of days from birth to slaughter.
‡ADLMG was calculated as the carcass weight multiplied by the percentage lean meat estimated with the CGM (Capteur Gras/Maigre) device, divided by the number of days from birth to slaughter.
§Fat and meat thickness were recorded at the level of the 3rd to 4th last rib using light reflection and % lean meat was estimated by using the official regression formulae.
∥Loin width, ham width, fat thickness and ham angle are SKGII measures that are used in a regression formula for the estimation of the type index.
***P < 0.001, **P < 0.01, *P < 0.05 and empty cells = P > 0.1.
The increased ADG of the MC4R mutation was accompanied by a lower LMP and a higher fat thickness, measured with the CGM device as well as measured manually (P < 0.05). The CGM muscle thickness was not influenced. The traits related to conformation (ham width, ham angle, type index and carcass length) were not affected by the MC4R mutation. The number of ribs and vertebra were also not different. The fat thickness and LMP values of the heterozygous animals were intermediate and were different from the values of the GG animals (P < 0.05). As a resultant from the opposite effects of the MC4R mutation on ADG and LMP, differences between MC4R genotypes for ADLMG were rather small (P < 0.1), but AA animals still displayed higher values than GG animals with the heterozygous animals being intermediate (P < 0.05).
Gilts grew slower than barrows (P < 0.001). Gender also influenced LMP, fat thickness as well as muscle thickness (P < 0.001), with the gilts having a leaner carcass compared with the barrows. Moreover, carcass length was increased in gilts (P < 0.001) and the SKGII measures were also different for barrows and gilts resulting in a lower and thus better type index for gilts (P < 0.001). ADLMG was slightly higher for gilts than for barrows (P < 0.001) (data not shown).
There was also an effect of sire on ADG and ADLMG (P < 0.001), and on LMP and carcass conformation traits (P < 0.05), except for muscle thickness and ham angle. When only the two sires having progeny with the three genotypes were considered, the differences in ADG and ADLMG were also significant (P < 0.001, data not shown).
Meat quality traits
The least-square means for meat quality traits by MC4R mutation are shown in Table 3. No differences were found for the MC4R genotypes for all meat quality traits except for shear force parameters and IMF content. The pH and PQM at 40 min and 24 h post mortem were similar for the three genotypes, except a small difference (P < 0.1) for pH1L and PQM2L. The meat from animals with the GG genotype had higher shear force values compared with meat from animals with genotypes AA and GA (P < 0.1). The higher shear force values were due to a higher myofibrillar work component (P < 0.05), whereas the collagen work component was not affected. The shear force values for AA and GA animals were similar. The GG animals had a lower IMF content compared with AA animals, with the heterozygous animals being intermediate (P < 0.05).
Table 3 Least-square means±s.e. for meat quality traits by the MC4R genotype

Abbreviations are: s.e. = standard error, MC4R = melanocortin-4 receptor genotype, GLM = general linear model, G = gender, and S = sire effect, IMF = intramuscular fat.
a,b,cMeans for the effect of line lacking a common superscript letter differ (P < 0.05).
†IMF content measured with near-infrared spectroscopy on 55 AA, 332 GA and 358 GG animals.
***P < 0.001, **P < 0.01, *P < 0.05 and empty cells = P > 0.1.
Slaughter day, included in the model as a random factor, was significant (P < 0.001) for all meat quality traits (data not shown). Also, sire had a significant effect on all meat quality traits, except for the shear force parameters. Gender had a significant influence on several meat quality traits, although the differences were small. The meat from gilts was less pale and more red, had lower thawing losses, higher shear force values compared with barrows and contained less IMF (data not shown).
Relationship between lean meat gain parameters and meat quality traits
The residual correlation coefficients between lean meat gain parameters and meat quality traits are shown in Table 4. As expected, the relationships between ADG on one hand and LMP and type index on the other hand were negative, and those between ADG, muscle thickness and ADLMG were positive.
Table 4 Residual correlation coefficients between lean meat gain parameters and meat quality traitsFootnote †

Abbreviations are: ADG = average daily gain, LMP = lean meat percentage measured by CGM device, ADLMG = average daily carcass lean meat gain, CGM, CGM = Capteur Gras/Maigre device, IMF = intramuscular fat content measured with near infrared spectroscopy.
† Significant correlations are shown in bold text (P < 0.05).
No relationships were apparent between ADG and most meat quality traits, except a negative relationship with the shear force traits and cooking losses and a positive relationship with drip loss and IMF. The pH1H was negatively related to LMP and PQM2L was positively related with LMP. Shear force traits, cooking losses and L* values showed a positive relationship with LMP and IMF was negatively related with LMP. Fat thickness was negatively related to the shear force traits and cooking losses, and a positive relationship between fat thickness and IMF, pH2 in loin and ham was apparent. In contrast, muscle thickness was not related to shear force parameters and IMF, but positive residual correlation coefficients were found with Temp1L, pH2 ham, PQM2 in loin and ham, L* and b* values and drip and negative coefficients with pH1 in loin and ham and cooking losses. The type index, as a measure for carcass conformation, showed a negative relationship with Temp1L, PQM2 in loin and ham, L* value and drip and a positive relationship with pH1 in loin and ham, a* value and IMF.
In contrast with ADG and LMP, ADLMG was not related to the shear force parameters and IMF but a positive relationship with PQM2 in loin and ham, L* value and drip was apparent. All these correlation coefficients were rather low and at best reached values above |0.1|. In summary, shear force traits, IMF and cooking losses appeared to be more influenced by differences in ADG and fat thickness, whereas PSE-related traits (pH1, PQM, drip, colour L* value) were more affected by muscularity traits (muscle thickness, type index).
Discussion
Effect of the MC4R mutation on growth, carcass and meat quality traits
The MC4R gene has been implicated in the regulation of feeding behaviour and body weight in humans and mice (Cone, Reference Cone1999). Moreover, the MC4R gene is frequently studied as a candidate gene for the control of economically important growth and performance traits in the pig. The wild-type allele (Asp298) is often associated with less backfat thickness, slower growth rate and lower feed intake, and the mutated allele (Asn298) has been associated with fatter, higher feed-consuming and faster-growing animals (Kim et al., Reference Kim, Larsen, Short, Plastow and Rothschild2000; Houston et al., Reference Houston, Cameron and Rance2004). Our study confirms these earlier reports. Although feed intake was not assessed in our study, similar findings with respect to growth rate and fat thickness suggest that these effects are mediated by changes in feed intake. However, it has also been hypothesised that the MC4R mutation could change the allocation of nutrients from protein deposition towards a higher fat deposition (Houston et al., Reference Houston, Cameron and Rance2004). This hypothesis is supported by our results. The MC4R mutation increased backfat thickness and IMF content and the higher LMP in the wild-type animals was clearly due to a lower backfat thickness, because muscle thickness was not affected. However, the effect of the MC4R gene seems to be line or breed specific. This is shown in the overview of literature data on MC4R given in Table 5. Inconsistent or even opposing effects of the MC4R mutation on ADG and backfat thickness have been reported between or within some studies depending on the breed or line (Kim et al., Reference Kim, Larsen, Short, Plastow and Rothschild2000, in a breed based on Meishan; Park et al., Reference Park, Carlborg, Marklund and Andersson2002, in a Wild Boar × Large White intercross; Chen et al., Reference Chen, Xiong, Zuo, Zheng, Li, Lei, Li, Deng and Jiang2005, in several breeds; Stachowiak et al., Reference Stachowiak, Szydlowski, Obarzanek-Fojt and Switonski2005, in Polish genotypes; see also Table 5). However, the majority of the data points to an increasing effect of the MC4R mutation on ADG, fat deposition and feed intake. This strengthens the hypothesis of Kim et al. (Reference Kim, Larsen, Short, Plastow and Rothschild2000) that the observed mutation is a causal mutation. The physiological explanation of the causal effect would be a disturbed appetite control due to an alteration in the binding efficiency of a ligand to MC4R (Kim et al., Reference Kim, Reecy, Hsu, Anderson and Rothschild2004). Increased feed intake is then leading to an increased ADG and fat deposition. Since growth and fat deposition are complex polygenic traits, breed- or line-specific allelic interactions that are responsible for variation in these polygenic traits might then explain the conflicting results in literature (Kim et al., Reference Kim, Larsen, Short, Plastow and Rothschild2000). Moreover, it is not excluded that the different or no effects in the given studies are due to the reduced number of animals that could limit the ability to detect small-sized effects.
Table 5 Overview of literature data concerning the effect of MC4R on carcass and meat quality traitsFootnote †

† Abbreviations are: MC4R = melanocortin-4 receptor genotype, LR = Landrace, LW = Large White, D = Duroc, P = Piétrain, M = Meishan, WB = Wild boar, QTL = quantitative trait loci, LMP = lean meat percentage, IMF = intramuscular fat, H = Hampshire, LE = Leicoma, T = Taihu.
a Difference is significant in the study (P < 0.1).
‡ Maximum number of animals in the experimental set-up of the study.
§ Growth can be expressed in the different reports as ADG or test daily gain or growth expressed as days to reach a specific weight.
¶ Although the same but more extensive population is used in this report, the numbers in Table 1 (allelic frequencies) and 3 (means) of this report give opposite results compared with the study from Kim et al. (Reference Kim, Larsen, Short, Plastow and Rothschild2000). Since the values are almost similar, we assume that the definition of the genotypes in this study is wrong. We therefore used the same conclusions as in Kim et al. (Reference Kim, Larsen, Short, Plastow and Rothschild2000) (e.g. A = allele 1 and G = allele 2 in Tables 1 and 3 of this report).
Based on the allelic frequencies, the MC4R mutation has apparently been subject to negative selection in the population on the current study, because the more favourable genotype for ADG had the lowest allelic frequency. Probably, selection has been more oriented to carcass leanness in this population. Similar allelic frequencies for MC4R have been reported in several other studies; however, also, largely different allelic frequencies were found (Table 5). Moreover, Houston et al. (Reference Houston, Cameron and Rance2004) showed that divergent selection for lean growth or feed conversion ratio altered the allelic frequencies after only seven generations of selection.
To the best of our knowledge, the current study is the first one that looked simultaneously at the effect of the MC4R mutation on carcass traits and a complete set of meat quality traits in a large number of animals. The report of Óvilo et al. (Reference Óvilo, Fernández, Rodríguez, Nieto and Silió2006) showed no effect on growth in a Spanish heavy breed, although a small effect on colour (L* and a*) was found but other meat quality traits, protein and moisture content, IMF, and pH were scarcely affected by MC4R. In the study of Otto et al. (Reference Otto, Roehe, Looft, Thoelking, Knap, Rothschild, Plastow and Kalm2007), the favourable homozygous AA genotype was associated with darker meat and with less drip loss, but the effect of the MC4R mutation on carcass parameters was not discussed in this study. Overall, the effects of the MC4R mutation on meat quality traits seem to be of minor importance. The post mortem pH and PQM values reflecting the course of glycolysis were not influenced by the MC4R genotype and consequently, no effects on drip loss or colour were to be expected. As was also reported by Otto et al. (Reference Otto, Roehe, Looft, Thoelking, Knap, Rothschild, Plastow and Kalm2007), there was a tendency for reduced drip losses in the AA genotype. Only meat tenderness seemed to be influenced by MC4R. The slightly faster growing and fatter AA genotype resulted in more tender meat, due to a lower force needed to shear the myofibrillar component. The collagen component of the shear force was not affected, suggesting that collagen content or solubility were not influenced. The relationship between meat tenderness and growth is dependent on the growth mechanism in the muscle (Koohmaraie et al., Reference Koohmaraie, Kent, Shackelford, Veiseth and Wheeler2002) and can be explained by muscle fibre size or the role of protein degradation during growth. Muscle fibre size or proteolytic enzymes were not assessed in the present study, hence their relative contribution to tenderness cannot be estimated. In addition, it is well known that the IMF content also can contribute to improved tenderness (Blanchard et al., Reference Blanchard, Willis, Warkup and Ellis2000; Lebret et al., Reference Lebret, Juin, Noblet and Bonneau2001), and it is possible that the slightly higher IMF content in the AA genotype is the solely mechanism for the slight decrease in shear force values in the present study.
Relationship between lean meat gain parameters and meat quality traits
In many literature studies, a negative relationship is apparent between carcass conformational traits and meat quality (reviewed by Sellier, Reference Sellier1998). Keeping meat quality at an acceptable level during selection for leanness has been a problem in the pig breeding industry. For instance, numerous studies have reported the positive effect of the halothane genotype on carcass lean content and conformation; however, this positive effect was often accompanied by a negative effect on meat quality traits (De Smet et al., Reference De Smet, Pauwels, Vervaeke, Demeyer, De Bie, Eeckhout and Casteels1995 and Reference De Smet, Pauwels, De Bie, Demeyer, Callewier and Eeckhout1996; Leach et al., Reference Leach, Ellis, Sutton, McKeith and Wilson1996). Also in other studies aiming at increasing lean tissue growth by selection or by feeding strategies, a decrease in meat quality was noticed (Karlsson et al., Reference Karlsson, Enfält, Essén-Gustavsson, Lundström, Rydhmer and Stern1993; Oksbjerg et al., Reference Oksbjerg, Petersen, Sørensen, Henckel, Vestergaard, Ertbjerg, Moller, Bejerholm and Stoier2000). On the other hand, the IGF-2 mutation shows that selection for increased LMP does not have to have unfavourable effects on meat quality (Van den Maagdenberg et al., 2005 and unpublished results). Divergent selection for lean growth rate or efficiency of feed conversion can result in similar responses in carcass composition, but can have different effects on meat quality traits (Cameron et al., Reference Cameron, Nute, Brown, Enser and Wood1999). In contrast to ours, the increased ADG was not accompanied by changes in LMP in the studies of Karlsson et al. (Reference Karlsson, Enfält, Essén-Gustavsson, Lundström, Rydhmer and Stern1993) and Oksbjerg et al. (Reference Oksbjerg, Petersen, Sørensen, Henckel, Vestergaard, Ertbjerg, Moller, Bejerholm and Stoier2000). In our study, higher ADG was associated with lower LMP, mainly due to a higher fat thickness. No relationship between ADG and PSE-related meat quality traits (colour, drip loss) was present in the studied population. However, a numerically negative and economically interesting relationship with shear force traits and cooking losses was found for ADG and fat thickness. As a consequence, the LMP showed an inverse relationship with shear force traits and cooking losses. A small effect of LMP on the L* value was also apparent. The correlation coefficients between shear force and cooking losses on the one hand with LMP and fat thickness on the other hand are in the opposite direction compared with the genetic correlations mentioned in Sellier (Reference Sellier1998). This probably can be explained by the influence of the IMF content on tenderness (Blanchard et al., Reference Blanchard, Willis, Warkup and Ellis2000; Lebret et al., Reference Lebret, Juin, Noblet and Bonneau2001), because the IMF content is negatively related with the mechanical measurements for tenderness in this study (data not shown). Moreover, it has been shown before that increasing the rate of lean growth is associated with higher shear force values (Cameron et al., Reference Cameron, Nute, Brown, Enser and Wood1999). As mentioned before, the relationship between meat tenderness and muscle growth is primarily dependent on the mechanism responsible for differences in growth, i.e. hyperplasia v. hypertrophy and protein synthesis v. protein degradation (Koohmaraie et al., Reference Koohmaraie, Kent, Shackelford, Veiseth and Wheeler2002). The significant residual correlation coefficients between LMP and fat thickness on the one hand and shear force on the other hand dropped from, respectively, 0.091 and −0.124 (P < 0.05) to 0.049 and −0.062 (P > 0.05) when a correction was made for the IMF content in this study. A comparable decrease for total and myofibrillar work was apparent. However, although correction for the IMF content decreased the residual correlation coefficients between ADG and shear force traits from −0.135 to −0.102, the relationship between these two parameters remained significant (P < 0.05). This shows that variation in IMF content contributed to variation in tenderness, but is not the only factor responsible for lower tenderness with increased ADG.
The responses of variation in ADG, LMP and ADLMG on PSE-related meat quality traits were small in the current study. However, the type index, as a measure of conformation, and muscle thickness were related to some extent to the early post mortem traits, e.g. pH1, PQM2, and Temp1, and colour and water-holding capacity. As was also argued by De Smet et al. (Reference De Smet, Van de Voorde and Demeyer1997), selection for improved conformation, i.e. round shaped carcasses, is more detrimental for meat quality than leanness per se.
Conclusion
In conclusion, the MC4R mutation has a positive influence on ADG, however, at the expense of increased fat thickness. No effects of the MC4R mutation on meat quality traits are to be expected, except for a slightly higher shear force in wild-type animals. Residual correlation coefficients revealed that meat quality should be controlled in selection programmes that aim at increasing the efficiency of lean meat production. Carcass traits but not ADG seem to be related to meat quality.
Acknowledgements
This research is financially supported by the Institute for the Promotion of Innovation through Science and Technology, Brussels. The authors thank all the technicians of the lab and the slaughterhouse for their excellent assistance. Family Deprez and co-workers are gratefully appreciated for producing the piglets, and Rattlerow-Seghers and Gentec are thanked for the financial and technical support.





