



Implications
The increasing availability of feed-grade amino acid (AA) allows a reduction of CP content in animal diets. Besides the beneficial effects of dietary protein reduction on animal health and environment, the use of feed-grade AA provides a more balanced supply of AA relative to their requirements. This study shows that responses of the zootechnical performance of piglets towards dietary supplementation of free l-tryptophan (l-Trp) and l-valine (l-Val) are dependent of the dietary concentrations of these AA and that dietary amino excess for leucine (Leu), isoleucine (Ile) and histidine (His) reduces performance of piglets on low-protein diets. This demonstrates the importance of supplying adequate dietary levels of AA in line with the piglet’s requirements, avoiding dietary AA deficiency.
Introduction
The reduction of CP levels in diets is acknowledged to reduce nitrogen and ammonia emission from pig production, and therefore limits the environmental impact such as via less eutrophication to the aquatic environment and acidification of soils (Monteiro et al., Reference Monteiro, Bertol, de Oliveira, Dourmad, Coldebella and Kessler2017). Moreover, it is well known that low-CP diets have a positive impact on the health status of weaned piglets (Gu and Li, Reference Gu and Li2004; Nyachoti et al., Reference Nyachoti, Omogbenigun, Rademacher and Blank2006; Wellock et al., Reference Wellock, Fortomaris, Houdijk and Kyriazakis2006).
The increased availability of feed-grade AA allows a further reduction of the dietary CP content and an increase of the nitrogen efficiency. Therefore, it is crucial to have knowledge on indispensable AA requirements of piglets in order to obtain a similar growth performance as with diets containing a higher CP content. For instance, a meta-analysis estimated the requirement of Trp at 22% as a ratio to standardized ileal digestible (SID) lysine (Lys), to maximize growth performance of piglets (Simongiovanni et al., Reference Simongiovanni, Corrent, Le Floc’h and van Milgen2012). Another meta-analysis showed that a value of 69% SID Val to Lys ratio enables to obtain optimal performance of piglets (van Milgen et al., Reference van Milgen, Gloaguen, Le Floc’h, Brossard, Primot and Corrent2013). It was also noticed that piglets are sensitive to a deficiency in Val (Gloaguen et al., Reference Gloaguen, Le Floc’h, Corrent, Primot and Van Milgen2012) or an excess in Leu (Wessels et al., Reference Wessels, Kluge, Hirche, Bartelt, Corrent and Stangl2015), which leads to a reduction of feed intake (FI) and therefore a deterioration of growth performance. Moreover, other studies established the requirement of Leu (Gloaguen et al., Reference Gloaguen, Le Floc’h, Corrent, Primot and Van Milgen2012 and Reference Gloaguen, Le Floc’h and van Milgen2013; Wessels et al., Reference Wessels, Kluge, Hirche, Bartelt, Corrent and Stangl2015), Ile (Nørgaard et al., Reference Nørgaard, Shrestha, Krogh, Sloth, Blaadbjerg, Poulsen, Tybirk and Corrent2013; Soumeh et al., Reference Soumeh, van Milgen, Sloth, Corrent, Poulsen and Nørgaard2014) and His (Li et al., Reference Li, Zhang and Gong2002) at 100%, 53% and 32% on a SID Lys ratio, respectively. Thus, the requirements of these AA are established and scientifically reported. However, it is also known that particular metabolic interactions between individual AA can impact piglet performance. As an example, a deficiency in Trp results in a drop in FI and lowers growth performance, especially in high-protein diets and when concentrations of large neutral AA (LNAA; phenylalanine (Phe), Tyr, Val, Leu, Ile) are in excess (Henry et al., Reference Henry, Sève, Colléaux, Ganier, Saligaut and Jégo1992; Jansman et al., Reference Jansman, Van Diepen and Melchior2010). An excess of dietary Leu is also accompanied by decreased plasma levels of Ile and Val and an increase in branched-chain AA (BCAA) catabolism, leading to a decrease of piglet performance (Wiltafsky et al., Reference Wiltafsky, Pfaffl and Roth2010). Many studies have been performed to establish the AA requirements (Trp, Ile, Val, Leu) in piglets but only a few focussed on the impact of AA excess on piglet performance and their possible interactions. The present studies hypothesized that the dietary levels of Val and Trp affect the response of FI and growth performance (Experiment 1), and that the extent of dietary imbalance for Ile, His and Leu reduces performance of piglets in the post-weaning phase in dependence on the dietary level of Val (Experiment 2).
Material and methods
The experimental protocols of both experiments were approved by the Animal Care and Use Committee of Wageningen University and Research, the Netherlands.
Animal, housing and management
A total of 320 castrated piglets (Great Yorkshire × Pietrain)×Dalland (Experiment 1) and 384 males piglets Topigs 20 and Topigs 40 (Experiment 2) were assigned to eight pens with eight replicates per treatment. Experiments 1 and 2 were carried out in two and three batches, respectively. Experiment 1 consisted in a pre-experimental period of 7 days and an experimental period of 28 days, starting at 34 days of age. Experiment 2 started at weaning (28 days of age) and lasted 34 days. For both experiments, piglets were distributed among pens to obtain the same mean BW per treatment per pen within a block of five (Experiment 1) or six pens (Experiment 2).
The average initial weight of piglets in Experiments 1 and 2 was 8.3 and 8.0 kg, respectively. During the experiments, piglets were housed in an artificially heated, ventilated and lighted pig unit with a specific-pathogen-free status. They were fed ad libitum and had free access to water via an automatic drinking service. In Experiment 1, all piglets were preventively treated against Streptococcus suis via the drinking water with the antibiotic Octacilline (Eurovet, Bladel, The Netherlands). This treatment was applied during days 1 to 5 of the experimental period for the first batch and during days 4 to 10 for the second batch.
Experimental diets
In Experiment 1, five experimental diets were formulated based on chemical analyses of the main ingredients for dry matter following the procedure from the International Organization for Standardization (www.iso.org; ISO 6496-1999) and for nitrogen according to the Dumas procedure (method V18-120, 1997 of the AFNOR group) using a Rapid N cube (Elementar France, Villeurbanne, France). Amino acid contents was determined using a JLC-500/V AminoTac Amino Acid Analyzer (Jeol, Croissy-sur-Seine, France) using the AFNOR method (standard NF EN ISO 13903). Methionine and cysteine were hydrolysed after oxidation with performic acid. Amino acids were separated by ion exchange chromatography and determined by photometric detection after derivatization by ninhydrin. Total Trp was analysed by HPLC after an alkaline hydrolysis with barium hydroxide using the AFNOR method (XP V18-114, 1998). Further data of the nutritional value of feed ingredients were taken from Centraal Veevoederbureau (CVB) (2007). Diets were based on wheat, barley, maize, soya bean meal, peas, whey powder and cane molasses (Table 1). During the pre-experimental period, all piglets received a high CP diet (diet 5), containing 19.5% CP and formulated to meet all AA requirements according to the ideal AA profile of Gloaguen et al. (Reference Gloaguen, Le Floc’h and van Milgen2013). From day 34 until the end of the experimental period, piglets were offered one of the five experimental pelleted diets. For the diets 1 to 4, basal ingredients were mixed as one batch and were formulated to be deficient in Val (SID Val : Lys=61%) and Trp (SID Trp : Lys=18%). Then, diet 1 was supplemented with 1.0 g/kg l-Val to obtain diet 2, with 0.5 g/kg l-Trp to obtain diet 3 and with 1.0 g/kg l-Val and 0.5 g/kg l-Trp to obtain diet 4. The dietary concentration of SID Lys was calculated to be slightly limiting at 1.03%. Diet 5 was formulated to be sufficient for all essential AA and was only supplemented with free l-Lys, dl-Met, l-Thr and l-Trp. The diets were analysed in duplicate by Ajinomoto Eurolysine S.A.S. Customer Laboratory (Amiens, France) for CP and total AA contents as further described above for the feed ingredients used. Based on these analyses, and information on the ileal AA digestibility of the feed ingredients and using the AA profile for basal endogenous AA losses per kilogram of dry matter intake according to the CVB (2007), dietary levels of SID AA were recalculated (Table 2). In diets 2 and 4, SID Val to SID Lys ratio (SID Val : Lys) was slightly above the assumed requirement (75.4% and 73.7% respectively), whereas SID Val : Lys was marginal relative to the assumed requirement in diets 1 and 3 (66.7%). For diets 3 and 4, SID Trp : Lys was at least at the requirement of piglets (22.8% and 23.1% SID Trp : Lys, respectively) and below the requirement for diets 1 and 2 (17.5% and 17.4% SID Trp : Lys, respectively). The level of net energy in the diets was kept constant among treatments (9.67 MJ/kg). Dietary CP content for diets 1, 2, 3, 4 and 5 was 17.7%, 17.8%, 17.6%, 17.8% and 19.5%, respectively.
Table 1 Ingredient and analysed nutrient composition of the respective experimental diets for the piglets for treatments 1 to 5 in Experiment 1 studying the interaction of dietary tryptophan and valine
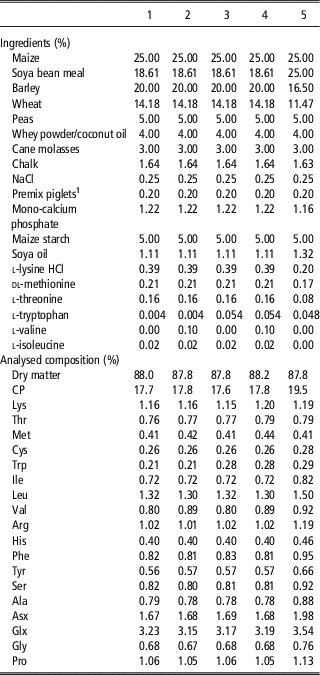
1 The vitamin–mineral premix supplied per kilogram feed: 10 000 IU vitamin A, 2000 IU vitamin D3, 20 IU vitamin E, 1.5 mg vitamin K, 1.0 mg vitamin B1, 4 mg vitamin B2, 15 mg d-pantothenic acid, 25 mg niacin, 25 µg biotin, 20 µg vitamin B12, 0.2 mg folic acid, 1.5 mg vitamin B6, 150 mg choline chloride, 150 mg Fe, 20 mg Cu, 65 mg Zn, 30 mg Mn, 0.15 mg Co, 0.5 mg I, 0.3 mg Se, 200 mg Fx aroma Uni (flavour).
Table 2 Calculated amino acid concentrations and profiles relative to lysine on a standardized ileal digestible (SID) basis in the experimental diets for piglets used in Experiment 1 studying the interaction of dietary tryptophan and valine
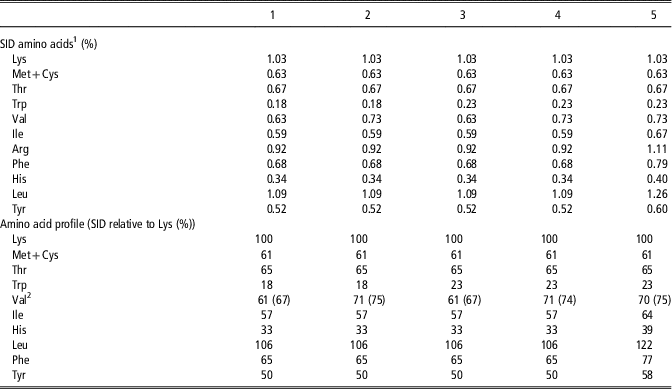
1 Calculated values based on data from CVB (2007) and analysed amino acid concentrations in the dietary ingredients.
2 Values in brackets are recalculated values based on analysed valine concentrations in the experimental diets.
In Experiment 2, six experimental diets were formulated after chemical analyses of the main ingredients on concentrations of dry matter, CP and AA by Ajinomoto Eurolysine S.A.S similar as described for samples of feed ingredients in Experiment 1 and their nutritional values according to CVB (2007). Diets were based on maize, wheat, barley and soya bean meal (Table 3). For the preparation of diets 1 to 6, the basal ingredients were mixed as a single batch of basal diet. Experimental diets were prepared by supplementing the basal diet with required amounts of free AA, in exchange of maize starch. All diets were pelleted with addition of steam. The calculated level of SID Lys was 1.13% in each of the experimental diets and was supposed slightly limiting for maximum performance of piglets (Boisen, Reference Boisen2003; Barea et al., Reference Barea, Brossard, Floc’h, Primot, Melchior and Van Milgen2009). The levels of other indispensable AA in all diets were adequate or in excess relative to the concentration of SID Lys according to the proposed ideal AA profile of Gloaguen et al. (Reference Gloaguen, Le Floc’h and van Milgen2013) (Table 4). Diets 1 to 3 were formulated to be limiting in Val (SID Val : Lys=60%) and diets 4 to 6 contained a higher level of Val (SID Val : Lys=70%). Compared to diets 1 and 4, diets 2, 3, 5 and 6 were additionally supplemented with free l-Ile (0.4 or 0.8 g/kg), l-His (0.24 or 0.48 g/kg) and l-Leu (1.5 or 3.0 g/kg), with increasing dose levels covering a range of excess that can be encountered in commercial diets. Dietary CP content in diets 1 to 6 varied between 17.5% and 18.1% and the net energy content was calculated to be 9.7 MJ/kg for each diet.
Table 3 Ingredient and nutrient composition of the respective experimental diets for the piglets for treatments 1 to 6 in Experiment 2 studying the effects of dietary amino acid imbalance
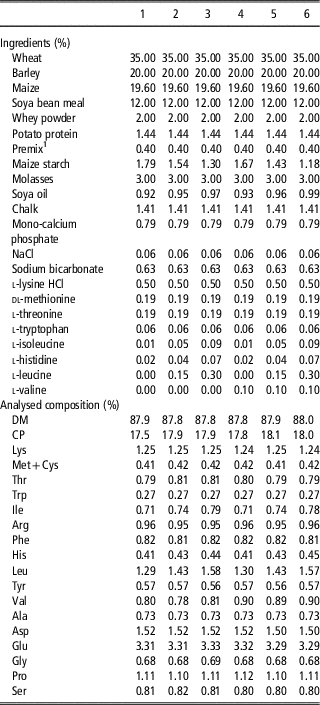
DM=dry matter.
1 The vitamin–mineral premix supplied per kilogram feed: 10 000 IU vitamin A, 2000 IU vitamin D3, 40 IU, vitamin E, 1.5 mg vitamin K3, 1.0 mg vitamin B1, 4 mg vitamin B2, 15 mg d-pantothenic acid, 30 mg niacin, 50 µg biotin, 20 µg vitamin B12, 0.3 mg folic acid, 1.5 mg vitamin B6, 150 mg choline chloride, 150 mg Fe, 160 mg Cu, 65 mg Zn, 30 mg Mn, 0.5 mg I and 0.3 mg Se.
Table 4 Calculated amino acid concentrations and profiles relative to lysine on a standardized ileal digestible (SIDW) basis in the experimental diets for the piglets of Experiment 2 studying the effects of dietary amino acid imbalance
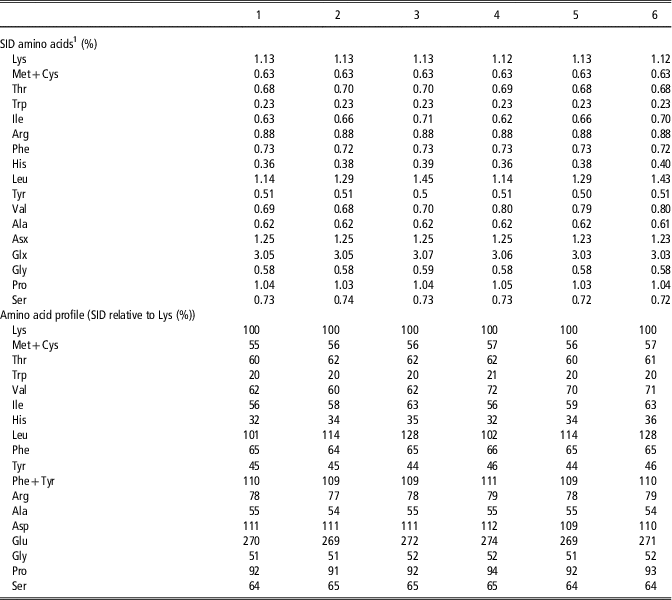
1 Calculated values based on data from CVB (2007) and analysed concentrations of total amino acids in experimental diets.
Animal performance
In both experiments, individual BW was measured at the start and at the end of the experimental period. In Experiment 1, piglets were weighed on days 0, 14 and 28 to determine the BW gain (BWG) for weeks 1 to 2, weeks 3 to 4 and for the overall period. In Experiment 2, piglets were weighed on days 0, 6, 20 and 34 post-weaning to determine the BWG for the 1st week, weeks 2 to 3, weeks 4 to 5 and for the overall period. Feed consumption was recorded per pen at each time point of animal weighing. From data on FI and BW, feed conversion ratio (FCR) was calculated. General health status of piglets was examined and mortality was reported.
Statistical analyses
The pen was the experimental unit in the both studies. Results (mean values per pen) were analysed statistically accordingly. For both experiments, effects of experimental factors on FI, BWG and FCR were analysed by ANOVA models (Genstat 5, release 18.1; Payne et al., Reference Payne, Lane, Ainsley, Bicknell, Digby, Harding, Leech, Simpson, Todd, Verrier and White1993). Data were analysed using block (initial live weight; four blocks per round (Experiment 1) and two or three blocks per round (Experiment 2)). A block is a set of pens in which each of the experimental treatment groups was equally represented. Experiment 1 was analysed as a 2×2 factorial design with the dietary level of Val and Trp and their interaction as experimental factors (treatments 1 to 4). In addition, data of all treatments were analysed using an ANOVA model with experimental treatment (treatment 1 to 5) as experimental factor. If the overall treatment effects were significant, differences between treatments means were compared further using the Duncan’s multiple range test. In Experiment 2, the results were also analysed using block, dietary Val level, dietary AA imbalance and their interaction as experimental factors as a 2×3 factorial design.
Effects and differences were considered significantly different at P<0.05 and were indicated as trends when P<0.10.
Results
Nutrient composition of diets
Analysed nutrient composition of the diets for Experiments 1 and 2 are given in Tables 1 and 3, respectively. In Experiment 1, chemical composition of the diets was in agreement with the calculated composition, with the exception of a higher analysed Val concentration in the diets than calculated. In Experiment 2, analysed contents of CP and AA in the experimental diets were higher than calculated values. Analysed content of CP in experimental diets were between 1.8% and 2.1% higher than calculated. For all AA, analysed values were higher by on average 13% on a relative basis, meaning that concentrations for all AA expressed in ratio to Lys were still in line with expected values.
Experiment 1
Results on piglet performance over the first and second 2 weeks periods and over the entire experimental period are presented in Table 5.
Table 5 Effects of dietary treatments on feed intake (FI, kg/day), BW gain (BWG, g/day) and feed conversion ratio (FCR) of piglets in Experiment 1
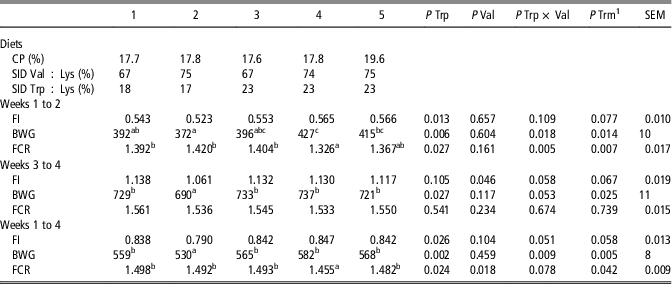
SID=standardized ileal digestible.
1 P-value for the overall treatment (diet) effect over the five experimental treatments. If the treatment effect was significant, treatment means were compared using Duncan’s multiple range test.
abcValues for the experimental treatments within a row with different superscripts differ significantly at P<0.05.
Considering treatments 1 to 4, during the first 2 weeks, FI was lower for piglets on the low-Trp diets (P<0.05), while for BWG and FCR there was a significant Trp × Val interaction (P<0.05 and P<0.01, respectively). Increasing the dietary level of Val decreased BWG and increased FCR when supplementing low-Trp diets with additional Val, whereas it improved BWG and decreased FCR in Trp sufficient diets. During weeks 3 and 4, FI decreased when increasing dietary Val level (P<0.05), whereas BWG enhanced when the dietary Trp level was increased (P<0.05). Feed conversion ratio was not affected by dietary level of Trp and Val. Over the entire experimental period, FI was affected by dietary Trp level (P<0.05) and FCR by both dietary Trp and Val level (both P<0.05), where increasing Trp level improved values while increasing dietary Val level reduced FI and increased FCR. For BWG, there was an interaction between dietary Trp and Val level (P<0.05), similar as described for BWG over weeks 1 to 2 of the experimental period. Piglets fed the low-CP diet with adequate levels of Val and Trp (treatment 4) had the same BWG but showed an improved FCR (P<0.05) compared to piglets fed the high-CP reference diet (treatment 5) over the entire experimental period.
Experiment 2
Results on piglet performance over the different periods of the experiment are presented in Table 6. During the first 6 days of the experimental period, increasing dietary Val level improved FI, BWG and FCR (P<0.01, P<0.001 and P<0.01, respectively). Increasing dietary AA excess for Ile, His and Leu decreased BWG and increased FCR. No interaction was observed between both experimental factors over this period. Over weeks 2 to 3, 4 to 5 and the entire experimental period, dietary level of Val increased FI and BWG (P<0.001) and increasing dietary AA excess reduced FI (P<0.05 for weeks 2 to 3 and 4 to 5 and P<0.01 for weeks 0 to 5). In addition, BWG was reduced by dietary AA excess over the entire period (P<0.05). Only, for FCR, an interaction was found between dietary Val level and AA excess over weeks 4 to 5 (P<0.05), FCR values being highest the medium level of excess of Ile/His/Leu when using the low-Val diets, and for the lowest level of excess in the Val sufficient diets, respectively.
Table 6 Effects of dietary treatments on feed intake (FI, kg/day), BW gain (BWG, g/day), feed conversion ratio (FCR) and BW (kg) of piglets in Experiment 2
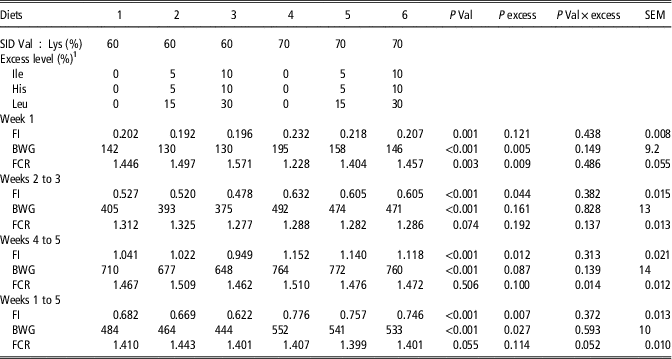
1 Excess level is expressed as the percentage of dietary excess of Ile, His and Leu relative to their requirement on a standardized ileal digestible (SID) basis (Gloaguen et al., Reference Gloaguen, Le Floc’h and van Milgen2013).
Discussion
Age effect
In both experiments, differences in animal performance were observed with regard to the effects of the dietary treatments between the first 2 weeks of the experimental period compared to considering the second part and the entire experimental period. In Experiment 1, the BWG of piglets fed the adequate Val and Trp, low-CP diet (treatment 4) was significantly higher than of piglets fed the Val and Trp deficient diet (treatment 1) only during weeks 0 to 2 of the experiment. In addition, FCR was significantly lower for piglets fed the adequate Val and Trp, low-CP diet (treatment 4) than for piglets fed the Val and Trp deficient diet only during weeks 0 to 2 (treatment 1), but not in weeks 3 to 4 of the experimental period. This suggests that the impact of a relative deficiency in Val and Trp in the diet was higher in the 1st weeks post-weaning. The former might be related to a gradual drop in AA requirements in time after weaning, which might be different for individual AA in both absolute sense and relative to Lys. Alternatively, it could suggest that piglets in the immediate post weaning phase show a higher sensitivity towards AA deficiencies compared to older piglets. In Experiment 2, increasing AA imbalance for Ile, Leu and His decreased growth performance of piglets, in particular during the 1st week when they were fed a Val adequate diet. During the remainder of the experiment, increasing AA imbalance for Ile, Leu and His numerically reduced FI and BWG in particular in piglets fed deficient Val diets. Again, the former might be related to relative changes in the requirement of individual AA in the post-weaning phase or piglets could be more sensitive to the AA imbalance during the 1st weeks post-weaning when Val was supplied adequately and in the period thereafter when dietary Val supply was deficient.
Imbalance for branched-chain amino acids
The study of Gloaguen et al. (Reference Gloaguen, Le Floc’h, Corrent, Primot and Van Milgen2012) showed that piglets rapidly detect Val deficiency or BCAA imbalance in the diet, leading to a reduction of FI and resulting in a reduction of growth performance. Results in Experiment 2 are in accordance with this study as increasing AA imbalance for Ile, Leu and His decreased both FI and BWG by on average 6% at the highest level of excess for Ile, His and Leu. In addition, the study showed a clear response of the piglets towards an increase dietary Val level from 60% to 70% relative to SID Lys and indicates that a value of 60% for Val relative to Lys is below requirement for maximum growth performance. A value of 70% for the requirement of Val has been suggested in more recent studies (Barea et al., Reference Barea, Brossard, Floc’h, Primot, Melchior and Van Milgen2009; Gloaguen et al., Reference Gloaguen, Le Floc’h, Corrent, Primot and Van Milgen2012; Soumeh et al., Reference Soumeh, van Milgen, Sloth, Corrent, Poulsen and Nørgaard2015), which is higher than the value of 63% recommended by National Research Council (2012).
Physiologically, the effects of AA imbalance for Ile, Leu and His as evaluated in the present study, can be related to different interactions between AA at systemic level in piglets. It has been shown that Leu can stimulate its own degradation via the branched-chain keto acid dehydrogenase (BCKDH), an enzyme complex, that catalyses the irreversible degradation of all BCAA, including Val and Ile (Wiltafsky et al., Reference Wiltafsky, Pfaffl and Roth2010). Wessels et al. (Reference Wessels, Kluge, Hirche, Kiowski, Schutkowski, Corrent, Bartelt, König and Stangl2016a) showed that excessive dietary supply of Leu increased the activity of BCKDH in the brain, liver and cardiac muscle in post-weaning piglets and reduced the concentrations of free Val and Ile in most tissues. Therefore, imbalances in the dietary supply of BCAA might induce metabolic losses of specific indispensable AA, resulting in AA deficiency and preventing maximum body protein deposition and growth performance.
Interaction between tryptophan and large neutral amino acids
Supplementing only free Trp to a Val and Trp deficient diet did not influence significantly growth performance of piglets. These results confirm the concept of the ideal AA profile and show that growth performance cannot be improved by Trp supplementation when another AA (Val) is limiting before Trp. A recent meta-analysis established a requirement at 22% SID Trp : Lys to maximize FI in piglets and therefore their growth performance (Simongiovanni et al., Reference Simongiovanni, Corrent, Le Floc’h and van Milgen2012). It has been shown that Trp interacts with the other LNAA (Leu, Ile, Val, Phe, Tyr) at the level of the brain and share common transporters across the blood brain barrier (Henry et al., Reference Henry, Sève, Colléaux, Ganier, Saligaut and Jégo1992; Fernstrom, Reference Fernstrom2013). Excess of LNAA in Trp deficient diets reduces transport and availability of Trp at brain level. Trp is a precursor of serotonin in the brain, which plays a role in appetite regulation (Sève, Reference Sève1999). Moreover, in weaned piglets Trp enhances the plasma level of ghrelin (Zhang et al., Reference Zhang, Yin, Li, Zhou and Li2007), a gastrointestinal hormone which increases growth hormone secretion and appetite (Salfen et al., Reference Salfen, Carroll, Keisler and Strauch2004). Although the diets in Experiment 1 were less deficient for Trp compared to the former studies, resulting in a dietary Trp : LNAA ratio from 4.9 to 6.5 in the present study compared to values in the range between 2.9 and 4.7 in the study of Sève (Reference Sève1999), an increase of Val concentration in the diet when Trp is deficient may have lowered Trp concentration in the brain resulting in a lower FI and concomitant BWG of piglets in Experiment 1. In Experiment 2, increasing dietary AA imbalance may also have reduced Trp concentration at brain level leading to a lower FI and growth performance of piglets. This was also demonstrated by Wessels et al. (Reference Wessels, Kluge, Hirche, Kiowski, Bartelt, Corrent and Stangl2016b) who showed that plasma concentration of Leu was negatively correlated with the concentration of Trp in the hypothalamus and that hypothalamic Trp was positively correlated with hypothalamic serotonin. They also showed that Trp and serotonin concentrations in the brain were decreased, with an increase of the level of Leu in the diet in excess of requirement (Wessels et al., Reference Wessels, Kluge, Hirche, Kiowski, Schutkowski, Corrent, Bartelt, König and Stangl2016a). These results demonstrate the value of the ideal protein concept for piglets, aiming to supply essential AA via the diet in agreement with the requirements of these nutrients, preventing their excess or deficiency and optimizing growth performance.
Conclusion
The present work confirms the interaction between dietary supply of Val and Trp on the zootechnical performance of post-weaning piglets. Increasing the dietary Val concentration from SID Val : Lys from 67% to 75% in low-protein diets increases BWG and tends to increase FI in piglets when Trp is supplied according to the requirement of piglets. Performance is reduced when increasing Val in Trp-deficient diets.
In addition, beside minimum dietary concentrations of indispensable AA, imbalances in dietary AA, in particular for Ile, Leu and His, should be considered in formulating low-CP diets for piglets. Imbalances for Ile, Leu and His, as observed in practical diets, reduced growth performance of piglets. Therefore, it is essential to provide all indispensable AA according to requirement values, preventing both deficiency and excess, to achieve maximum growth performance in piglets.
Acknowledgement
None.
Declaration of interest
The authors declare that they have no conflict of interest.
Ethics statement
The present studies were not considered animal trials for which formal authorization was required in the Netherlands.
Software and data repository resources
None of the data were deposited in an official repository.







 Open access
Open access