


Introduction
Orycrocephalid trilobites play an important role in Cambrian chronostratigraphy because of their relatively wide geographic distributions. The first appearance datum (FAD) of Oryctocephalus indicus (Reed, Reference Reed1910) has been proposed as the primary tool for recognizing the conterminous base of Cambrian Series 3 and Stage 5 (Zhao et al., Reference Zhao, Yuan and McCollum2001b, Reference Zhao, Yuan, Peng, Guo, Zhu, Peng and Wang2004; Peng et al., Reference Peng, Babcock and Cooper2012), and the FAD of another orytocephalid, Ovatoryctocara granulata (Chernysheva, Reference Chernysheva1962), is used as another intercontinental tie point below the base of Stage 5, which has been proposed as an alternative level to define the conterminous base of Series 3 and Stage 5 (Fletcher, Reference Fletcher2003; Shabanov et al., Reference Shabanov, Korovnikov, Pereladov and Fefelov2008). The base of the Duyunian Stage, a regional stage used for South China, is marked by the FAD of the oryctocephalid Arthricocephalus chauveaui (Peng, Reference Peng2000). However, the concepts of Arthricocephalus and its type species A. chauveaui have long been unsettled, as discussed herein. Confusion in the concept of A. chauveaui was introduced by the fallacious designation of a lectotype (Lane et al., Reference Lane, Blaker and Zhang1988) that was not illustrated in the original publication (Bergeron, Reference Bergeron1899), and which is shown here to belong to another oryctocephalid species. Poor understanding of the generic concept of Arthricocephalus lead not only to suppression of the genus Oryctocarella as a junior synonym of Arthricocephalus (Suvorova, Reference Suvorova1964), but also to incorrect assignment of Oryctocarella specimens to Arthricocephalus (e.g., Qian, Reference Qian1961; Lu and Qian, Reference Lu and Qian1964; Qian and Lin in Zhou et al., Reference Zhou, Liu, Meng and Sun1977; Yin in Yin and Li, Reference Yin and Li1978; Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980; Zhang and Zhou, Reference Zhang and Zhou1985; Blaker and Peel, 1977; Yuan et al., Reference Yuan, Zhao, Li and Huang2002; McNamara et al., Reference McNamara, Yu and Zhou2003; Peng et al., Reference Peng, Zhao, Wu, Yuan and Tai2005a, Reference Peng, Zhao and Yang2006), and to erection of new genera having concepts identical with Arthricocephalus.
This article clarifies the generic concepts of Arthricocephalus and Oryctocarella, prompted by a restudy of the original type series of Arthricocephalus chauveaui, the type species of Arthricocephalus. In addition, topotypic material of Oryctocarella sibirica, the type species of Oryctocarella, has been collected and studied, leading to further elucidation of the concept of this monospecific genus.
Revised concept of Arthricocephalus
Bergeron (Reference Bergeron1899) erected Arthricocephalus, with A. chauveaui Bergeron, Reference Bergeron1899 as the type species by monotypy. Bergeron (Reference Bergeron1899, p. 514) stated that the genus and the type species were based on 10 sclerites preserved on the bedding surface of a small slab of limestone (Fig. 1). The slab is composed of lime mudstone, and measures 6.2 cm by 3.8 cm. It was collected from the Balang Formation, north of Tongren, eastern Guizhou, South China, and sent to France by M. Chauveau for study by Bergeron. The sclerites on the slab include five exoskeletons in varying degress of articulation, a small external mold of an incomplete meraspid cephalon, and an external mold of a holaspid pygidium (Figs. 1, 2.1–2.12). Bergeron (Reference Bergeron1899) evidently considered the specimens to be conspecific, and used all of them to establish the morphologic limits of A. chauveaui. The original publication included one illustrated specimen, a dorsal exoskeleton lacking the free cheeks (reproduced here as Fig. 2.1).

Figure 1 Slab with the type series of Arthricocephalus chauveaui Bergeron, Reference Bergeron1899, reassigned to three species, each in a separate genus; from the Balang Formation, north of Tongren, eastern Guizhou, China. Museum of the Geological Department, Claude Bernard University, Lyon, France, E.M. 90001. Sclerites (1–6, 8) were renumbered as E.M. 90001a–g by Lane et al. (Reference Lane, Blaker and Zhang1988). An external mold of incomplete cephalon (7) was not previously illustrated and now numbered E.M. 90001h. (1–2) Arthricocephalus chauveaui Bergeron, Reference Bergeron1899: (1) disarticulated holaspid exoskeleton; (2) external mold of holaspid pygidium. (3–7) Oryctocarella duyunensis (Qian, Reference Qian1961): (3, 4) two meraspid exoskeletons; (5) external mold of cranidium; (6) thoracopygon; (7) external mold of incomplete cephalon. (8) Duyunaspis duyunensis Zhang and Qian in Zhou et al., Reference Zhou, Liu, Meng and Sun1977, meraspid exoskeleton. See also caption of Figure 2 for detailed description of each specimen. Scale bar=10 mm.

Figure 2 Original illustration (1) and type series (2–12) of Arthricocephalus chauveaui Bergeron, Reference Bergeron1899, from the Balang Formation, north of Tongren, eastern Guizhou, China; all sclerites are on a single slab, all in dorsal view unless stated otherwise. White arrowhead indicates posterior margin of thorax; arrowhead with black outline indicates the posterior margin of partially released segment of transitory pygidium; black triangles indicate posteromedial notch on pygidial border; b=border, c=connective suture, d=doublure. (1–3, 10, 11) Arthricocephalus chauveaui Bergeron, Reference Bergeron1899: (1) line-drawing of exoskeleton, illustrated as fig. 9 in Bergeron (Reference Bergeron1899); (2) E.M. 90001a, type specimen, designated as lectotype, disarticulated holaspid exoskeleton with inverted cranidium, librigena and rostral-hypostomal plate; (3) reversal of image in 2 (lectotype), from negative film; (10) latex cast from external mold of rostral-hypostomal plate and cephalic doublure of specimen in 2 and 3 (lectotype), ventral view; (11) E.M. 90001g, latex cast of external mold of pygidium; dark curved region at margin is the mold of the pygidial doublure; part of the upturned border is preserved at left upper corner of pygidium. (4–9) Oryctocarella duyunensis (Qian, Reference Qian1961), specimens determined by Lane et al. (Reference Lane, Blaker and Zhang1988) as A. chauveaui: (4) E.M. 90001c, disarticulated thoracopygon, posterior margin of thorax uncertain (arrows indicate alternative possibilities); specimen may be meraspid degree 10 or early holaspid (11 segments); (5) E.M. 90001b, inverted cranidium, external mold in ventral view, illustrated and designated as the lectotype of Arthricocephalus chauveaui by Lane et al. (Reference Lane, Blaker and Zhang1988, pl.1, fig. 2); (6) latex cast from external mold of cranidium in 5; (7, 8) two meraspid exoskeletons lacking librigenae: (7) E.M. 90001d, meraspid degree 9; (8) E.M. 90001e, meraspid degree 8; (9) E.M. 90001h, latex cast of external mold of incomplete meraspid cephalon in Figure 1.7. (12) Duyunaspis duyunensis Zhang and Qian in Zhou et al., Reference Zhou, Liu, Meng and Sun1977: E.M. 90001f, meraspid exoskeleton, specimen identified as meraspid degree 5 or 6 of A. chauveaui by Lane et al. (Reference Lane, Blaker and Zhang1988). All scale bars represent 1.0 mm.
Bergeron’s (Reference Bergeron1899) original concept of A. chauveaui was generally accepted by subsequent authors including Lane et al. (Reference Lane, Blaker and Zhang1988), Blaker and Peel (Reference Blaker and Peel1997), and McNamara et al. (Reference McNamara, Yu and Zhou2003). Lane et al. (Reference Lane, Blaker and Zhang1988) regarded all of the sclerites present on Bergeron’s slab (Fig. 1) as syntypes and illustrated nearly all of them photographically. They selected one cranidium (Figs. 1.5, 2.5, 2.6) as the lectotype of A. chauveaui and regarded all other specimens except for a small cranidium (Figs. 1.7, 2.9) on the slab (Figs. 1.1–1.4, 1.6, 1.8, 2.4, 2.7, 2.8, 2.11, 2.12) as paralectotypes. The specimen selected by Lane et al. (Reference Lane, Blaker and Zhang1988) as the lectotype is not the specimen originally illustrated as a line drawing by Bergeron (Reference Bergeron1899, fig. 9; reproduced here as Fig. 2.1).
Restudy of the material on Bergeron’s slab shows that three species are present, each referable to a separate genus (Figs. 1, 2). The exoskeleton and only specimen illustrated by Bergeron (Reference Bergeron1899, fig. 9; Fig. 2.1) can be inferred to be the intended type specimen (a lectotype by ICZN convention). The cranidium selected by Lane et al. (Reference Lane, Blaker and Zhang1988) as the lectotype (Figs. 1.5, 2.5, 2.6), by contrast, belongs to Oryctocarella duyunensis. The designating of this specimen by Lane et al. (Reference Lane, Blaker and Zhang1988) created confusion about the generic concept because most authors seem to have relied on Bergeron’s line drawing for guidance on the species and generic concepts.
Photographs of Bergeron’s (Reference Bergeron1899) specimens published by Lane et al. (Reference Lane, Blaker and Zhang1988, pl. 1, figs.1–5) as A. chauveaui show great morphologic differences among the sclerites in the type series, suggesting different species are included. Bergeron’s slab was not re-examined in the preparation of the 1988 paper (P.D. Lane, personal communication, 2016). In 2002, SP examined the type material of Arthricocephalus chauveaui at the museum of the Geological Department, Claude Bernard University, Lyon, France, and rephotographed all exposed sclerites. The examination demonstrated that Bergeron’s line drawing of A. chauveaui (Fig. 2.1) is generally accurate, as concluded by Lane et al. (Reference Lane, Blaker and Zhang1988, p. 558). The examination also indicated that the line drawing was based principally on the largest exoskeleton on the slab. That specimen (Fig. 2.2, 2.3, 2.10) is easily identifiable among the sclerites on the slab. The specimen that evidently served as the basis for the illustration and much of the original description (Bergeron, Reference Bergeron1899) is an exuvium of the exoskeleton having the cranidium, librigenae, and hypostome disarticulated from the thoracopygon and overturned (Figs. 1.1, 2.2, 2.3, 2.10). Features showing that the exoskeleton is the basis for Bergeron’s illustration and description include: (1) outline of glabella, which is pestle-shaped, expanded forward anterior to the S3 furrow, and defined laterally by curved axial furrows; (2) nature of the posterior three glabella furrows (S1–S3), which are transversely continuous across the glabella and reach to the axial furrows; (3) shape of the posterior three glabellar lobes (L1–L3), which are narrow, subrectangular, about three times wider than long, and subequal in length; (4) length of the “anterior lobe” of the glabella (formed by fusion of L4 and the frontal lobe), which is twice as long as L3; (5) segmentation pattern of the thorax, with eight segments, the posteriormost of which is incompletely released; (6) semicircular outline of the pygidium (subequal to the cranidium in length); (7) segmentation of the pygidial axis (five rings and a terminal piece); (8) faint interpleural furrows of the pygidium; and (9) smooth, continuous arc forming the posterior margin of the pygidium.
Bergeron’s (Reference Bergeron1899) original description of A. chauveaui provides further evidence that the species’ description was principally based on the largest exoskeleton on the slab. Characters noted by Bergeron include: (1) expansion in width of the anterior glabellar lobe and expansion of its length to twice that of the other lobes, implying the glabella is expanded forward anterior to S3; (2) eight thoracic rings (segments); (3) six rings in the pygidial axis; (4) cranidial length of 2.5 mm; and (5) pygidial length of 2.5 mm. None of the other exoskeletons present on the slab has this combination of characters. The sum of information published by Bergeron (Reference Bergeron1899) indicates that the largest exoskeleton on the original limestone slab was the basis for most of the species description and for the illustration, and it is inferred that this exoskeleton was the intended type specimen.
Examination of Bergeron’s type exoskeleton reveals some misinterpretation of features in the original illustration (Bergeron, Reference Bergeron1899, fig. 9). The ridge-like structure along the pygidial border as illustrated is actually the ventral doublure (compare Fig. 2.1, 2.11). This misinterpretation may have been influenced in part by the appearance of the largest pygidium on the slab (Fig. 1.2), which is preserved as an external mold. The latex cast of the pygidial external mold reveals that the “ridge” is the ventral doublure (Fig. 2.11). This pygidium is the only sclerite on Bergeron’s (Reference Bergeron1899) slab that is conspecific with the largest exoskeleton. Fulcra and facets on the thoracic segments were not illustrated by Bergeron (Reference Bergeron1899), although they are present. The fulcra and facets are not well exposed on the type specimen, and are not presented on the other thoracic material on the slab except for the smallest exoskeleton (Fig. 2.12).
Apart from the specimen illustrated here in Figure 2.2 and 2.3, there are four additional exoskeletons on Bergeron’s slab (E.M. 90001c [thoracopygon associated with cranidium, 90001b], 90001d, 90001e, 90001f), but none of them matches the original illustration and description. A number of differences between these exoskeletons and the largest exoskeleton on the slab (E.M. 90001a) indicate that they are not the specimens on which Bergeron’s illustration was based, and that they are neither conspecific nor congeneric with A. chauveaui. Three exoskeletons (E.M. 90001b and 90001c, 90001d, 90001e; Figs. 1.3–1.6, 2.4–2.8) are reassigned herein to Oryctocarella duyunensis (Qian, Reference Qian1961). New, abundant and well-preserved material from the Balang Formation helps to clarify the differences between A. chauveaui and O. duyunensis (Fig. 3), and each of the distinguishing characteristics of O. duyunensis is present on the three sub-largest exoskeletons of Bergeron’s slab. These exoskeletons differ from A. chauveaui in: (1) having a narrow cylindrical glabella that is parallel-sided and defined laterally by straight axial furrows; (2) having pit-like glabellar furrows that are isolated from the axial furrows and two or three pairs of furrows (S1, S2, S3) connected transversely by furrows, but not extending abaxially to the axial furrows; (3) having straight rather than curved axial furrows; (4) having proportionally long L1–L3 and a short “anterior lobe” (L4 plus frontal lobe, nearly as long as L3); (5) having an upturned rather than ridge-like cranidial border; (6) having palpebral lobes in a relatively anterior position; (7) having an ocular ridge relatively closer to the anterior border furrow; (8) having a thorax with more than eight segments (up to 10 or 11; Fig. 2.4); (9) having a smaller pygidium that is shorter than the cephalon; (10) having a pygidial axis with three rather than five axial rings; (11) having well-defined interpleural furrows; (12) having a medial notch at the posterior margin of the pygidium; and (13) lacking a pygidial border rather than having an upturned border (Figs. 2.4, 2.7–2.8, 3.2). The thoraxes in each of these three exoskeletons are apparently non-fulcrate, differing in this character from the fulcrate thorax with geniculate segments of the largest exoskeleton. This difference was suggested by Whittington (Reference Whittington1995, p. 555) who found that the “weakly geniculated pleurae” described by Lane et al. (Reference Lane, Blaker and Zhang1988, p. 559) for A. chauveaui are not present on some specimens assigned to Arthricocephalus by Zhang et al. (Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980). Cephala on each of the three sub-largest exoskeletons appear to have proparian facial sutures, whereas on E.M. 90001a the sutures are gonatoparian.
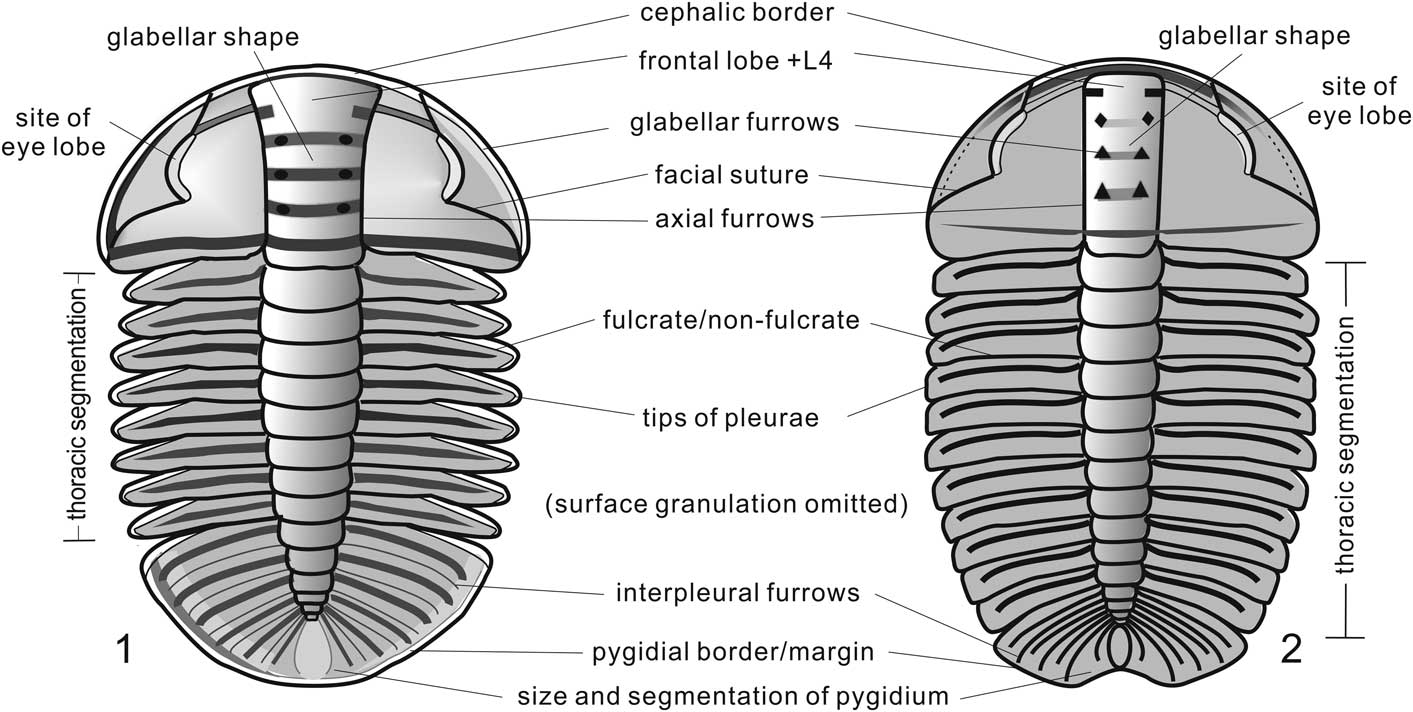
Figure 3 Reconstructions of Arthricocephalus chauveaui (1) based on NIGP 163354, and Oryctocarella duyunensis (2) based on NWU-DYXJT 1823, both from the Balang Formation of western Hunan, China. These species show differences in glabellar outline (forwardly expanding or pestle-shaped, 1, compared to cylindrical, 2); development of the glabellar furrows (transglabellar, 1, compared to pit-like, 2); course of the axial furrow (curved, 1, compared to straight, 2); course of the facial suture (gonatoparian, 1, compared to proparian, 2); shape of the anterior cranidial border (upturned posterolaterally, 1, compared to anteriorly, 2); position of the palpebral lobe (situated at the level of the glabellar mid-point, 1, compared to located anteriorly, 2); presence,1, or absence, 2, of fulcra; thoracic segmentation (eight segments, 1, rather than 11 segments, 2); tips of pleurae (pointed, 1, rather than slightly rounded, 2); size and segmentation of pygidium (isopygous, with five pleurae, 1, compared to micropygous, with three pleurae, 2); pygidial interpleural furrows (faint, 1, rather than well defined, 2); pygidial border (upturned, 1, rather than absent, 2); development of the posterior pygidial margin (equally curved, 1, rather than with a median notch, 2); and surface prosopon (smooth to finely granulose, 1, as compared to coarsely and densely granulose, 2; surface granulation on the reconstruction of each species has been omitted).
The smallest exoskeleton on Bergeron’s (Reference Bergeron1899) slab (E.M. 90001f, Figs. 1.8, 2.12) also is not conspecific with A. chauveaui although it has long been considered to be a meraspid of this species (e.g., Lane et al., Reference Lane, Blaker and Zhang1988; Blaker and Peel, Reference Blaker and Peel1997; McNamara et al., Reference McNamara, Yu and Zhou2003). The specimen also differs considerably from specimens on Bergeron’s (Reference Bergeron1899) slab reassigned to Oryctocarella duyunensis. The specimen is here reassigned to Duyunaspis duyunensis Zhang and Qian in Zhou et al., Reference Zhou, Liu, Meng and Sun1977 (p. 132, pl. 41, figs. 5, 6), the type species of Duyunaspis. The exoskeleton differs from both A. chauveaui and O. duyunensis in having a proportionally large glabella that is wider than the fixigena; shallow and weakly impressed glabellar furrows; a proportionally wide thoracic axis that is nearly as wide as the pleural area; and a relatively small pygidium with a broad axis. The specimen of D. duyunensis has a medial notch on the posterior margin of the pygidium, similar to O. duyunensis, but the pygidial axis in specimens of O. duyunensis is much narrower, being only about half as wide as the pleural area. The thorax of the smallest exoskeleton on Bergeron’s slab has fulcrate segments, similar to A. chauveaui, but the fulcra in the smallest specimen lie much closer to axial furrow than they do in the largest exoskeleton.
We conclude that Bergeron’s (Reference Bergeron1899) original illustration and description of A. chauveaui are based principally on the largest exoskeleton in the type series. According to Article 74.6.1 of the International Code of Zoological Nomenclature (ICZN, 1999), which states that “The inference that the specimen is a ‘holotype’ or ‘the type’ may be by reference to an illustration or description of specimen,” the largest exoskeleton on Bergeron’s slab is inferred as ‘type’ of A. chauveaui. In accordance with Article 74.6, “the assumption that the species-group taxon was based upon a single type specimen is deemed to have designated that specimen as the lectotype.” For these reasons, the largest exoskeleton on Bergeron’s (Reference Bergeron1899) limestone slab (E.M. 90001a) is designed the lectotype of Arthricocephalus chauveaui. Arthricocephalus is considered to be a monotypic genus.
An external mold of a large pygidium (E.M. 90001g; Figs. 1.2, 2.11) on Bergeron’s (Reference Bergeron1899) slab remains assigned to Arthricocephalus chauveaui; it is a paralectotype. Other sclerites on the slab, which are presumed to be part of Bergeron’s suite of examined specimens and all originally referred to A. chauveaui, are also considered to be paralectotypes of A. chauveaui. All of these specimens, however, are here reassigned as either Oryctocarella duyunensis (E.M. 90001b–90001e, 90001h) or Duyunaspis duyunensis (E.M. 90001f).
Incorrect designation of lectotype
Circumstances surrounding the designation of a lectotype of Arthricocephalus chauveaui by Lane et al. (Reference Lane, Blaker and Zhang1988), and subsequent treatment of Bergeron’s (Reference Bergeron1899) suite of material, has resulted in ambiguity of the concepts of both A. chauveaui Bergeron, Reference Bergeron1899 and Arthricocephalus Bergeron, Reference Bergeron1899. Photographs of some Bergeron’s sclerites were first published by Lane et al. (Reference Lane, Blaker and Zhang1988). In that same article, Lane et al. (Reference Lane, Blaker and Zhang1988, p. 558) stated that they accepted the specimen upon which Bergeron’s line drawing was based (Bergeron, Reference Bergeron1899, fig. 9) as the lectotype of the species because it was the only illustration Bergeron published of A. chauveaui. As originally published (Bergeron Reference Bergeron1899, fig. 9; reproduced herein as Fig. 2.1), the exoskeleton was separated into three parts—cranidium, thorax, and pygidium—but examination of the limestone slab makes it clear that the illustration is of an exoskeleton separated into a thoracopygon with an associated, inverted cranidium (Fig. 2.2, 2.3). Bergeron’s (Reference Bergeron1899) description also seems to apply to this specimen. Because the individual specimen on which most of Bergeron’s (Reference Bergeron1899) original published information was based can be identified from the original slab of examined material, there can be little doubt that it was the intended ‘type,’ and by implication should be the lectotype of the species.
Lane et al. (Reference Lane, Blaker and Zhang1988) seem to have followed this line of reasoning, but a mixup evidently occurred at publication stage, leading to designation of an inverted cranidium, E.M. 90001b (Fig. 2.5, 2.6) as the lectotype of Arthricocephalus chauveaui. That specimen is here reassigned to Oryctocarella duyunensis (Qian, Reference Qian1961). The article by Lane et al. (Reference Lane, Blaker and Zhang1988) was published in Chinese and English, and the Chinese portion of the paper was translated from a draft originally written in English. In the English description, Lane et al. (Reference Lane, Blaker and Zhang1988) clearly referred to the exoskeleton (E.M. 90001a) rather than the cranidium (E.M. 90001b) as the specimen selected to be the lectotype. As described (Lane et al., Reference Lane, Blaker and Zhang1988, p. 559), the cranidium has a “subrectangular glabella” (not a cylindrical glabella as present in E.M. 90001b) and “four pairs of deep glabellar pits, S1–S3 reaching to axial furrows and connected across glabella also connected across glabella by shallow transverse furrow” (not isolated from the axial furrows); the thorax has “pleurae with weak as geniculation” (i.e., segments are fulcrate rather than non-fulcrate, implying the presence of thoracic facets). As discussed above, the only exoskeleton in Bergeron’s (Reference Bergeron1899) suite of examined specimens that meets all these criteria is E.M. 90001a.
In the Chinese translation, the discussion related to the designation of the specimen in Bergeron’s (Reference Bergeron1899) figure 9 as the lectotype, and the description of the glabellar furrows meeting the axial furrows, were omitted. According to P.D. Lane (written communication, 2016), designation of a lectotype was somehow changed from that in the draft, without his knowledge, both in the description and plate explanation. As published, a cranidium, E.M. 90001b, was indicated as the lectotype (Lane et al., Reference Lane, Blaker and Zhang1988, p. 555, 558, 559, pl. 1, right in fig. 2; herein, Figs. 1.5, 2.5, 2.6). The Chinese part of the article was published prior to the English portion, and the altered designation of lectotype, which was at odds with the written morphological description published later in English, was included in it. Repetition of this designation at the time when the English portion of the article was published exacerbated the problem.
Because the cranidium (E.M. 90001b) was erroneously selected as the lectotype of Arthricocephalus chauveaui, but in fact belongs to Oryctocarella duyunensis, it has been the source of some misinterpretation of the concepts of both Arthricocephalus and A. chauveaui. Blaker and Peel (Reference Blaker and Peel1997, p. 110, fig. 62.1) and Geyer (Reference Geyer2005, fig. 3) interpreted the largest exoskeleton on Bergeron’s (Reference Bergeron1899) slab (E.M. 90001a) as the basis for Bergeron’s (Reference Bergeron1899) illustration, indicating it as the lectotype of the species, and discussed the concept of A. chauveaui in light of that view. M.R. Blaker was a co-author of the Lane et al. (Reference Lane, Blaker and Zhang1988) paper, and his 1997 monograph (Blaker and Peel, Reference Blaker and Peel1997) showed that he did not realize there had been a change in the designation of a lectotype for A. chauveaui.
Yuan et al. (Reference Yuan, Zhao, Li and Huang2002, p. 120) and McNamara et al. (Reference McNamara, Yu and Zhou2003, p. 107) accepted the cranidium (E.M. 90001b) with its associated thoracopygon (E.M. 90001c) to be the lectotype and discussed a different concept of Arthricocephalus chauveaui. The cranidium and its associated thoracopygon may originate from a single individual, but Lane et al. (Reference Lane, Blaker and Zhang1988, p. 555, 559) definitely specified the cranidium to be the lectotype. This thoracopygon differs considerably from the thorax and pygidium illustrated by Bergeron (Reference Bergeron1899, fig. 9;=E.M. 90001a). The differences from Bergeron’s illustrated specimen include a greater number of thoracic segments, non-fulcrate segments, and a pygidium that is shorter and less segmented. The species concept discussed by Yuan et al. (Reference Yuan, Zhao, Li and Huang2002, p. 120) and McNamara et al. (Reference McNamara, Yu and Zhou2003, p. 107) applies to Oryctocarella duyunensis.
We conclude that the designation of the crandium (E.M. 90001b) from Bergeron’s (Reference Bergeron1899) limestone slab by Lane et al. (Reference Lane, Blaker and Zhang1988) is invalid. As discussed above, Bergeron’s original illustration (1899, fig. 9) allows us to infer that E.M. 90001a was the intended ‘type,’ deemed a lectotype, of A. chauveaui despite the fact that Bergeron (Reference Bergeron1899) obviously regarded all specimens on the slab as belonging to the same species. According to Article 74.1.3 of the Zoological Code (ICZN, 1999), “The valid designation of a lectotype permanently deprives all other specimens that were formerly syntypes of that nominal taxon of the status of syntype.” Because the intended ‘type’ can be unequivocally identified from the syntypic series through comparison to the original illustration (Bergeron, Reference Bergeron1899, fig. 9), subsequent designation of any other specimen as a lectotype would be invalid and misleading. Furthermore, in accordance with Recommendation 74B of the Code (ICZN, 1999), “Other things being equal, an author who designates a lectotype should give preference to a syntype of which an illustration has been published.” Bergeron’s illustration (1899, fig. 9) was evidently based on a single specimen, E.M. 90001a, and this specimen therefore should have priority for designation as a lectotype over any non-illustrated sclerite in the original syntypic series.
Historical review of Arthricocephalus and its junior synonyms
Saito (Reference Saito1934, p. 232, pl. 25, figs. 26–29) questionably referred some specimens from northwestern Korea to Arthricocephalus. According to Rasetti (Reference Rasetti1959, p. O220) and Blaker and Peel (Reference Blaker and Peel1997, p. 109), these specimens do not belong to Arthricocephalus but to Cheiruroides Kobayashi, Reference Kobayashi1935. Rasetti (Reference Rasetti1959, p. O524) regarded the genus Arthricocephalus to be unrecognizable and therefore failed to include it in the family Oryctocephalidae.
Qian (Reference Qian1961) assigned some specimens from the Balang Formation at Palang, Duyun, southeastern Guizhou, China, to a new species of Arthricocephalus, A. duyunensis. Qian’s (Reference Qian1961) material is from a locality ~100 km southwest of Tongren, the type locality of A. chauveaui. As revised here, these specimens are all referable to Oryctocarella, showing that Qian’s (Reference Qian1961) paper influenced subsequent work on Arthricocephalus and created confusion about the concepts of both Oryctocarella and Arthricocephalus (see Qian and Lin in Zhou et al., Reference Zhou, Liu, Meng and Sun1977; Yin and Li, Reference Yin and Li1978; Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980; Liu, Reference Liu1982; Ju, Reference Ju1983; Zhang and Zhou, Reference Zhang and Zhou1985; Blaker, Reference Blaker1986; Blaker and Peel, Reference Blaker and Peel1997; Yuan et al., Reference Yuan, Zhao, Li and Huang2002, Reference Yuan, Zhao and Yang2006; McNamara et al., Reference McNamara, Yu and Zhou2003; Geyer, Reference Geyer2005; Peng et al., Reference Peng, Zhao, Wu, Yuan and Tai2005a, Reference Peng, Zhao, Wu, Yuan and Taib). Subsequent synonymization of Oryctocarella with Arthricocephalus (Suvorova, Reference Suvorova1964; Shergold, Reference Shergold1969; Blaker and Peel, Reference Blaker and Peel1997) apparently stems in part from this broad interpretation of Arthricocephalus.
Tomashpolskaya (in Khalfin, Reference Khalfin1960, p. 199, pl. 23, fig. 5) erected a new species, Oryctocara sibirica, and this species was later used (Tomashpolskaya and Karpinski, Reference Tomashpolskaya and Karpinski1961) as the type species of the new genus Oryctocarella. Suvorova (Reference Suvorova1964, p. 235) considered the genus to be a junior synonym of Arthricocephalus. This suppression proved to be incorrect and, as revised here, Oryctocarella is revived as an independent genus.
Qian and Lin (in Lu et al., Reference Lu, Zhang, Qian, Zhu, Lin, Zhou, Qian, Zhang and Wu1974) erected a monotypic subgenus Arthricocephalus (Arthricocephalites) with the new species A. (Arthricocephalites) xinzhaiheensis as its type species. Differential characteristics of the subgenus were not published until six years later. At that time Qian and Lin (in Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, p. 275–279) stated that A. (Arthricocephalites) differed from A. (Arthricocephalus) in having a longer palpebral lobe, a narrower fixigena, and a shorter (exsag.) posterior area of the fixigena. As now understood, their concept of A. (Arthricocephalus) is identical with that of Oryctocarella, whereas their concept of A. (Arthricocephalites) is identical with that of Arthricocephalus (as revised here).
The unfortunate designation of cranidium E.M. 90001b as the lectotype of Arthricocephalus chauveaui Bergeron, Reference Bergeron1899 by Lane et al. (Reference Lane, Blaker and Zhang1988) has led to misunderstanding about the concept of Arthricocephalus. The cranidium is here referred to Oryctocarella duyunensis (Qian, Reference Qian1961). Some generic names have been mistakenly interpreted as junior synonyms of Arthricocephalus (see Blaker and Peel, Reference Blaker and Peel1997; Yuan et al., Reference Yuan, Zhao, Li and Huang2002), and some species have been mistakenly interpreted as Arthricocephalus (see Blaker and Peel, Reference Blaker and Peel1997; Yuan et al., Reference Yuan, Zhao, Li and Huang2002; McNamara et al., Reference McNamara, Yu and Zhou2003; Yuan et al., Reference Yuan, Zhao and Yang2006; Geyer and Peel, Reference Geyer and Peel2011).
Blaker and Peel (Reference Blaker and Peel1997, p. 109) were unable to determine any consistent differences between A. (Arthricocephalus) and A. (Arthricocephalites), citing individual variability in the three features identified by Qian and Lin (in Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980) as differentiating the subgenera. For this reason, Blaker and Peel (Reference Blaker and Peel1997) suppressed A. (Arthricocephalites) as a junior synonym of Arthricocephalus. However, they included specimens now referable to Arthricocephalus (Blaker and Peel, Reference Blaker and Peel1997, fig. 62.1) and Oryctocarella (Blaker and Peel, Reference Blaker and Peel1997, fig. 62.2, 62.3) in A. chauveaui, leading to a broad concept of Arthricocephalus that embraces what we now consider to be Arthricocephalus and Oryctocarella.
In their 1997 paper, Blaker and Peel also erected a new genus, Haliplanktos, with Arthricocephalus (Arthricocephalites) jishouensis Zhou in Zhou et al., Reference Zhou, Liu, Meng and Sun1977 as its type species (Blaker and Peel, Reference Blaker and Peel1997, p. 112). Haliplanktos, as revised here, is regarded as a junior synonym of Arthricocephalus, and A. (A.) jishouensis is apparently based on a meraspid of A. chauveaui (Lei, Reference Lei2016, and discussion below).
Yuan et al. (Reference Yuan, Zhao, Li and Huang2002, p. 120–123) elevated A. (Arthricocephalites) and A. (Arthricocephalus) to generic rank and did not fully accept Blaker and Peel’s (Reference Blaker and Peel1997) synonymization of the taxa. They argued that Arthricocephalus differs from Arthricocephalites in having a proportionately wide cranidium with a broad fixigenae; narrow librigenae; a cylindrical glabella; short palpebral lobes; proparian facial sutures; a thorax having a narrow axis and eight to 11 segments; a small (micropygous [or heteropygous; see Kobayashi, Reference Kobayashi1942]) pygidium with four to five axial rings and well-defined interpleural furrows. In contrast, Arthricocephalites, in their view, is characterized by having a narrower crandium with narrow fixigenae; wide librigenae; a forwardly expanding glabella; moderately long palpebral lobes; a gonatoparian facial suture; a thorax with a relatively wide axis and five to eight segments; a large (isopygous) pygidium with six to eight axial rings and faint or obsolescent interpleural furrows. The concept of Arthricocephalus used by Yuan et al. (Reference Yuan, Zhao, Li and Huang2002) equates to that of Oryctocarella as revised here, whereas their concept of Arthricocephalites equates to that of Arthricocephalus as revised here.
McNamara et al. (Reference McNamara, Yu and Zhou2003) considered A. (Arthricocephalites) (=Arthricocephalites of Yuan et al., Reference Yuan, Zhao, Li and Huang2002) and Arthricocephalus (sensu Yuan et al., Reference Yuan, Zhao, Li and Huang2002) to be synonymous. Their concept of Arthricocephalus embraced taxa assigned here to both Arthricocephalus and Oryctocarella.
The concept of Arthricocephalus used by Geyer and Peel (Reference Geyer and Peel2011), being guided by the publication of Lane et al. (Reference Lane, Blaker and Zhang1988), equates to that of Oryctocarella as used here. Geyer and Peel (Reference Geyer and Peel2011) considered Haliplanktos to be a valid genus, whereas as revised here, it is considered to be a junior synonym of Arthricocephalus.
Large collections of oryctocephalid trilobites, including some topotypic material, have been amassed from the Balang Formation in eastern Guizhou and western Hunan onward from about 1961 (see Qian, Reference Qian1961; Zhou in Zhou et al., Reference Zhou, Liu, Meng and Sun1977; Yin and Li, Reference Yin and Li1978; Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980; Yuan et al., Reference Yuan, Zhao, Li and Huang2002, Reference Yuan, Zhao and Yang2006, Reference Yuan, Zhao, Peng, Zhu and Lin2009; McNamara et al., Reference McNamara, Yu and Zhou2003, Reference McNamara, Yu and Zhou2006; Peng et al., Reference Peng, Zhao and Yang2006; Ma et al., Reference Ma, Peng, Zhao, Da and Sun2011; Lei and Peng, Reference Lei and Peng2014; Lei, Reference Lei2016), and this material helps to clarify the morphology and characterize the various taxa now under discussion. Together with restudy of Bergeron’s (Reference Bergeron1899) slab from the Balang Formation of eastern Guizhou, the new material provides clarification about the concept of Arthricocephalus, and leads us to revive Oryctocarella as an independent, well-recognizable genus, and to regard Arthricocephalites and Haliplanktos as junior synonyms of Arthricocephalus.
Validity of Oryctocarella
Tomashpolskaya and Karpinski (Reference Tomashpolskaya and Karpinski1961) erected the genus Oryctocarella (with Oryctocara sibirica Tomashpolskaya in Khalfin, Reference Khalfin1960 as the type species) for an oryctocephalid trilobite based on material from the Kuznetsk Alatau of the Altay-Sayan Foldbelt, southwestern Siberia. At that time, at least 10 variously complete exoskeletons were known. Oryctocarella was characterized by having eight thoracic segments (similar to the number of segments in Arthricocephalus chauveaui). The Siberian material appears to be congeneric with material from eastern Guizhou that was incorrectly assigned to Arthricocephalus by Qian (Reference Qian1961), leading Suvorova (Reference Suvorova1964, p. 235) to suppress the monospecific Oryctocarella as a junior synonym of Arthricocephalus. Blaker and Peel (Reference Blaker and Peel1997, p. 109) likewise suppressed Oryctocarella, noting that the holotype exoskeleton of O. sibirica is “comparable in all morphological features to A. chauveaui.” Shergold (Reference Shergold1969, p. 40) noted that O. sibirica has segmentation identical with Arthricocephalus chauveaui, and that Suvorova (Reference Suvorova1964) had considered the two genera as possible synonyms. However, Peng et al. (Reference Peng, Zhu, Babcock and Korovnikov2015) considered that Oryctocarella should be revived as a valid genus because it is significantly distinguishable in exoskeletal characters from Arthricocephalus.
Unfortunately, the type material of Tomashpolskaya and Karpinski (Reference Tomashpolskaya and Karpinski1961) is lost (Korovnikov and Novozhilova, Reference Korovnikov and Novozhilova2017), and the original figures of O. sibirica are poor, making it hard to determine its diagnostic features, especially the exact number of thoracic segments. From the original illustration of the holotype (Tomashpolskaya and Karpinski, Reference Tomashpolskaya and Karpinski1961, pl. 1, fig. 2), it appears that O. sibirica has at least nine thoracic segments. Recently collected topotypic material of O. sibirica from its type locality in the Kuznetsk Alatau, Altay-Sayan Foldbelt (see Fig. 6) reveals that the species has nine or possibly 10 thoracic segments. In addition, topotypes show that O. sibirica shares a number of important characters with O. duyunensis, allowing clear distinction from A. chauveaui (see Fig. 3). Distinguishing characters include a cylindrical glabella, pit-like glabellar furrows that are isolated from the axial furrows, a non-fulcrate thorax, and a relatively short pygidium with clearly defined interpleural furrows. A pygidial border and border furrow are not defined. These characters clearly differentiate Oryctocarella from Arthricocephalus so that, following Peng et al. (Reference Peng, Zhu, Babcock and Korovnikov2015) and Korovnikov and Novozhilova (Reference Korovnikov and Novozhilova2017), the genus must be regarded as valid and discrete.
A large number of specimens from China previously assigned to Arthricocephalus are referable to Oryctocarella. They include specimens illustrated as Arthricocephalus duyunensis Qian, Reference Qian1961 , A. granulus Qian and Lin in Zhou et al., Reference Zhou, Liu, Meng and Sun1977 , A. jiangkouensis Yin in Yin and Li, Reference Yin and Li1978, A. tenuis Zhang and Zhou, Reference Zhang and Zhou1985, and A. cf. A. granulus sensu Zhang and Zhou, Reference Zhang and Zhou1985. Some of these described species are regarded here as synonymous with Oryctocarella duyunensis (Qian, Reference Qian1961).
Materials and methods
Figured specimens were collected from the Cambrian Balang Formation in various localities of South China and from the lower part of the traditional Middle Cambrian in a single locality of Siberia, Russia. The localities are indicated by these acronyms: BL=Bulin, Huayuan, northwestern Hunan, China; DZ=Nangao, Danzhai, eastern Guizhou, China; DM=Dolgii Mys (Long Cape) Mountain, Khakassia, Batenevsky Ridge, Kuznetsk Alatau, Altay-Sayan Foldbelt, Russia; GY=Palang (previously misspelled as Balang), Duyun, southeastern Guizhou, China; HN-GZ=Border area between Hunan and Guizhou provinces, China (exact collecting locality unknown); KH=Panxin and Feilongshan, Songtao, eastern Guizhou, China; LYX=Luoyixi, Guzhuang, northwestern Hunan, China; MZC=Mozichong, Huayuan, northwestern Hunan, China; TJBL=Balang, Taijiang, eastern Guizhou, China; TR=Tongren, eastern Guizhou, China; WW=Wawu, Tongren, Guizhou, China (village that has administrative jurisdiction over the type locality of A. chauveaui); XHZA=Paiwu (Zila A), Huayuan, northwestern Hunan, China; XHZB=Paiwu (Zila B), Huayuan, northwestern Hunan, China; XS=Xiaosai, Yuqing, eastern Guizhou, China.
Specimens in Bergeron’s (Reference Bergeron1899) type series of Arthricocephalus chauveaui were coated with magnesium oxide and then photographed with an Olympus OM4 film camera. Topotypes of Oryctocarella sibirica were photographed with a digital camera. All other specimens were coated with black ink and followed by magnesium oxide, then photographed under a Zeiss stereomicroscope (Model Axio Zoom V16) having a digital head (AxioCam MrM) and a circle light around the lens providing uniform lighting. Measurements were made from images of sclerites using the ruler tool in Adobe Photoshop CS6. Scale bars were either photographed along with the images of specimens by film photography or produced automatically by digital photography.
Repositories and institutional abbreviations
Illustrated and referred specimens are deposited in the following institutions: CSGM, Central Siberian Geological Museum, Novosibirsk, Russia; E.M., Geological Department, Claude Bernard University, Lyon, France; Gt, Guizhou Institute of Regional Geological Survey, Guiyang, Guizhou, China; NIGP, Nanjing Institute of Geology and Palaeontology, Nanjing, China; NWU, Geological Department, Northwest University, Xi’an, China.
Spellings of author names
English spellings of the names of some Chinese authors have changed through time. The Pinyin (Chinese Phonetic Alphabet), introduced in 1958, was accepted in 1982 as the International Standard for the Spelling of Chinese Characters (ISO7098), and it is Pinyin spellings that are used here. Names of Chinese authors affected by the change to Pinyin spellings include Guo (changed from Kuo), Li (from Lee), Qian (from Chien), Xiang (from Hsiang), Zhang (from Chang), and Zhu (from Chu).
Transliterated names of Russian authors follow commonly accepted spellings.
Systematic paleontology
Terminology mostly follows that of Whittington and Kelly (Reference Whittington and Kelly1997). In the following descriptions, “long” and “short” are used with reference to longitudinal (sagittal, sag., and exsagittal, exsag.) dimensions, and “broad” (or “wide”) and “narrow” are used with reference to transverse (tr.) dimensions (Whittington, Reference Whittington1997, p. O2).
Order Corynexochida Kobayashi, Reference Kobayashi1935
Family Oryctocephalidae Beecher, Reference Beecher1897
Subfamily Oryctocarinae Hupé, Reference Hupé1953
Genus Arthricocephalus Bergeron, Reference Bergeron1899
1899 Arthricocephalus Reference BergeronBergeron, p. 514.
1974 Arthricocephalus (Arthricocephalites) Qian and Lin in Reference Lu, Zhang, Qian, Zhu, Lin, Zhou, Qian, Zhang and WuLu et al., p. 94.
1988 Arthricocephalus Bergeron; Reference Lane, Blaker and ZhangLane et al. (part), p. 555.
1997 Haliplanktos Reference Blaker and PeelBlaker and Peel, p. 112.
2002 Arthricocephalites Qian and Lin in Reference Yuan, Zhao, Li and HuangYuan et al., p. 122.
Type species
Arthricocephalus chauveaui Bergeron, Reference Bergeron1899, p. 514, text-fig. 9, from the Balang Formation, near Tongren, eastern Guizhou, China; by monotypy.
Other species
Arthricocephalus (Arthricocephalites) xinzhaiheneis Qian and Lin in Lu et al., Reference Lu, Zhang, Qian, Zhu, Lin, Zhou, Qian, Zhang and Wu1974, p. 95, pl. 36, fig. 7, from the Balang Formation, at Panxin, Songtao, eastern Guizhou, China.
Diagnosis
Isopygous or subisopygous genus of Oryctocarinae with arched cranidial anterior margin; glabella parallel-sided along L1–L2 or L1–L3, expanded forward anterior to S2 or S3, defined laterally by outwardly curved axial furrow; S1–S3 pit-like or narrow furrows, connected by transglabellar furrows terminating at the pits or continuing laterally to the axial furrows; palpebral lobe moderately long, slightly oblique; palpebral area slightly narrower than glabella; facial suture gonatoparian; librigena narrow. Thorax fulcrate, with eight segments; axis moderately wide. Pygidium nearly as long as cranidium, axis with five rings plus a terminal piece; pleural furrows wide and clearly defined, interpleural furrow faint; pygidial border upturned and poorly defined by wide border furrow; posterior margin without medial notch.
Remarks
As summarized above, identification of E.M. 90001a as Bergeron’s (Reference Bergeron1899) intended ‘type’ (now lectotype) of Arthricocephalus chauveaui leads to revision of the generic concepts of Arthricocephalus and Oryctocarella. New synonymies are introduced, and Haliplanktos is considered to be a junior synonym of Arthricocephalus.
Arthricocephalus resembles Oryctocarella, which is revived here as a separate genus. Characters that distinguish Arthricocephalus from Oryctocarella include a forwardly expanding glabella, curved axial furrows, glabellar furrows reaching the axial furrows, a fulcrate thorax with eight segments, and a relatively large pygidium with poorly defined interpleural furrows and a well-defined border. Oryctocarella, in contrast, has a cylindrical glabella, straight axial furrows, pitted S1–S3 glabellar furrows that are isolated from axial furrows, a non-fulcrate thorax with nine or 10 segments, and a relatively short pygidium with well-defined interpleural furrows and a poorly defined pygidial border.
New material from northwestern Hunan shows that the holotype of A. (Arthricocephalites) jishouensis, the type species of Haliplanktos, is a meraspid degree 6 of Arthricocephalus chauveaui (Lei, Reference Lei2016). New material also shows that A. (Arthricocephalites) xinzhaiheensis, the type species of A. (Arthricocephalites), shares general diagnostic characters with A. chauveaui, differing only in the nature of the S1–S3 glabellar furrows, which are pitted and connected with transverse furrows between pits. Both species are reassigned here to Arthricocephalus.
Representatives of Arthricocephalus are known from South China (Guizhou and Hunan provinces) and North Greenland, and range in position from Cambrian Stage 4 (Duyunian of the South China regional standard) to Stage 5 (Taijiangian of South China).
Arthricocephalus chauveaui Bergeron, Reference Bergeron1899
Figures 1.1, 1.2, 2.1–2.3, 2.10, 2.11, 3.1, 4

Figure 4 Arthricocephalus chauveaui Bergeron, Reference Bergeron1899 from the Balang Formation in northwestern Hunan (1–5, 7, 8, 10–12) and eastern Guizhou (6, 9), China. White arrowhead indicates posterior margin of thorax; arrowhead with black outline indicates posterior margin of partially released segment of transitory pygidium; all in dorsal view; b=border, d=doublure; f=facet: (1) NIGP 164838, exoskeleton, meraspis degree 4, LYX-1; (2) NIGP 164839, exoskeleton, meraspis degree 4, LYX-1; (3) NIGP 163346, exoskeleton, meraspis degree 5, XHZA407-3, original of Lei (2015, fig. 3.3); (4) NIGP 164952, distorted exoskeleton, meraspis degree 5, LYX-1; (5) NIGP 164840, exoskeleton, meraspis degree 6, LYX-1; (6) NIGP 38233, exoskeleton, meraspis degree 6, HN-GZ, holotype of Arthricocephalus (Arthricocephalites) pulchellus Zhang and Qian in Zhang et al. (Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 92, fig. 3); illustrated originally as Arthricocephalus chauveaui by Lu et al. (Reference Lu, Zhang, Zhu, Qian and Xiang1965, pl. 17, fig. 1), reillustrated as Arthricocephalus taijiangensis by Yuan et al. (Reference Yuan, Zhao and Li2001, pl. 2, fig. 5; 2006, fig. 2c) and as Arthricocephalus pulchellus by McNamara et al. (Reference McNamara, Yu and Zhou2003, pl. 2, fig. 12); (7) NIGP 164841, exoskeleton, meraspis degree 6, LYX-1; (8) enlargement of the right side of the thorax of NIGP 164841 in 7, showing pleural facets; (9) NIGP 164842, exoskeleton, meraspis degree 7, WWf2; (10) NIGP 163349, exoskeleton, holaspid, latex cast from external mold, XHZA329-13; (11), NIGP 163354, exoskeleton, holaspid, XHZA325-27; (12) NIGP 163357, latex cast of external mold of incomplete holaspid exoskeleton lacking librigenae and with displaced cranidium, XHZA416-9, original of Lei (Reference Lei2016, fig. 3.14). All scale bars represent 1.0 mm.
1899 Arthricocephalus chauveaui Reference BergeronBergeron, p. 514, fig. 9.
1957 Arthricocephalus chauveaui; Reference LuLu, p. 265, pl. 139, fig. 11.
1962 Arthricocephalus chauveaui; Reference Lu, Zhu and QianLu et al., p. 27, pl. 1, fig. 9, pl. 6, fig. 10.
1963 Arthricocephalus chauveaui; Reference Lu, Qian and ZhuLu et al., p. 64, pl. 7, figs. 7a–d.
1963 Arthricocephalus chauveaui; Reference Egorova, Xiang, Li, Nan and GuoEgorova et al., p. 20, pl. 2, fig. 9.
non 1964 Arthricocephalus chauveaui; Reference Lu and QianLu and Qian, p. 26, pl. 1, fig. 5.
1965 Arthricocephalus chauveaui; Reference Lu, Zhang, Zhu, Qian and XiangLu et al., p. 108, pl. 17, fig. 1.
1977 Arthricocephalus (Arthricocephalites) jishouensis Zhou in Reference Zhou, Liu, Meng and SunZhou et al., p. 131, pl. 42, figs. 4–6.
non 1977 Arthricocephalus chauveaui; Reference Zhou, Liu, Meng and SunZhou et al., p. 130, pl. 42, fig. 3.
1978 Arthricocephalus (Arthricocephalites) tongrenensis Yin in Reference Yin and LiYin and Li, p. 442, pl. 155, fig. 17.
1978 Arthricocephalus taijiangensis Yin in Reference Yin and LiYin and Li, p. 441, pl. 156, fig. 8.
non 1980 Arthricocephalus chauveaui; Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and YuanZhang et al., p. 275, pl. 92, figs. 1, 2.
1980 Arthricocephalus (Arthricocephalites) pulchellus Zhang and Qian in Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and YuanZhang et al., p. 276, pl. 92, fig. 3.
1982 Arthricocephalus chauveaui; Reference LiuLiu, p. 300, pl. 213, fig. 9.
1982 Arthricocephalus (Arthricocephalites) jishouensis; Reference LiuLiu, p. 300, pl. 213, figs. 7, 8, 11.
1988 Arthricocephalus chauveaui; Reference Lane, Blaker and ZhangLane et al. (part), p. 555, pl. 1, figs. 1, 5; non figs. 2–4, 6.
1997 Arthricocephalus chauveaui; Reference Blaker and PeelBlaker and Peel (part), p. 109, fig. 62.1; non figs. 62.2, 62.3, 63, 64.1–64.4.
1997 Haliplanktos jishouensis (Zhou); Reference Blaker and PeelBlaker and Peel, p. 112, fig. 64.5–64.7.
2001 Arthricocephalites taijiangensis; Reference Yuan, Zhao and LiYuan et al., pl. 2, fig. 5.
2002 Arthricocephalites taijiangensis; Reference Yuan, Zhao, Li and HuangYuan et al., p. 123, pl. 30, figs. 3–5, pl. 31, figs. 2–6.
non 2002 Arthricocephalus chauveaui; Reference Yuan, Zhao, Li and HuangYuan et al., p. 121, pl. 31, fig. 1.
non 2003 Arthricocephalus chauveaui; Reference McNamara, Yu and ZhouMcNamara et al., p. 106, pl. 1, figs. 1–15, pl. 2, figs. 5, 7, text-fig. 3A–K.
2003 Arthricocephalus pulchellus; Reference McNamara, Yu and ZhouMcNamara et al., p. 115, pl. 2, figs. 10–12, text-fig. 6A, 6B.
2005 Arthricocephalus chauveaui; Reference GeyerGeyer (part), p. 87, fig. 3.7; non fig. 3.1–3.4, 3.8, 3.9.
2005 Haliplanktos jishouensis; Reference GeyerGeyer, p. 88, fig. 3.5, 3.6.
non 2005a Arthricocephalus chauveaui; Reference Peng, Zhao, Wu, Yuan and TaiPeng et al., p. 1161, fig. 2e.
non 2005b Arthricocephalus chauveaui; Reference Peng, Zhao, Wu, Yuan and TaiPeng et al., p. 1056, fig. 2e.
2006 Arthricocephalus xinzhaiheensis Qian and Lin; Reference Yuan, Zhao and YangYuan et al. (part), p. 617, fig. 2b; non fig. 2a.
non 2006 Arthricocephalus chauveaui; Reference Yuan, Zhao and YangYuan et al., p. 615, fig. 1d, 1e.
2006 Arthricocephalites taijiangensis; Reference Yuan, Zhao and YangYuan et al., p. 617, fig. 2c–j.
non 2006 Arthricocephalus chauveaui; Reference Peng, Zhao and YangPeng et al., p. 241, pl. 1, fig. 11.
2009 Arthricocephalus chauveaui; Reference Yuan, Zhao, Peng, Zhu and LinYuan et al., p. 214, fig. 1c, 1cc’.
2011 Arthricocephalites taijiangensis; Reference Ma, Peng, Zhao, Da and SunMa et al., p. 746, fig. 3c.
2011 Haliplanktos jishouensis; Reference Geyer and PeelGeyer and Peel, p. 499, fig. 15A–F.
2013 Arthricocephalus xinzhaiheensis; Reference LeiLei, p. 72, figs. 24, 25.
2015 Arthricocephalus chauveaui; Reference Peng, Zhu, Babcock and KorovnikovPeng et al., p. 89, pl. 1, figs. 1–4.
2016 Arthricocephalus chauveaui; Reference LeiLei (part), p. 496, figs. 3.1–3.4, 3.6, ?3.8, 3.9–3.12, 3.14, ?3.15, 4.1–4.3, 4.5, 4.6, 4.8, ?4.9–4.16; non figs. 3.5, 3.7, 3.13, 4.2, 4.7.
2016 Arthricocephalites xinzhaiheensis; Reference Shen, Peng, Wen and LiuShen et al. (part), p. 16, fig. 1.8–1.11; non fig. 1.1–1.7.
2016 Arthricocephalites intermedius Zhou; Reference Shen, Peng, Wen and LiuShen et al., p. 15, fig. 2.1–2.3, 2.6–2.13, ?2.4, ?2.5.
Lectotype
Disarticulated exoskeleton (E.M. 90001a; Figs. 1.1, 2.2, 2.3, 2.10); also illustrated by Lane et al. (Reference Lane, Blaker and Zhang1988, pl. 1, fig. 1), Blaker and Peel (Reference Blaker and Peel1997, fig. 62.1), and Geyer (Reference Geyer2005, fig. 3.7), from the Balang Formation, north of Tongren, eastern Guizhou, China; original of Bergeron (Reference Bergeron1899, fig. 9).
Diagnosis
Arthricocephalus with glabellar furrows continuous transversely across glabella, reaching to axial furrow; glabella expanded forward of S2 or S3; preocular area nearly half as long (exsag.) as frontal lobe (sag.); palpebral lobe slightly oblique to almost exsagittal in holaspid stage with posterior end at a level corresponding to S1; posterior branch of facial suture strongly divergent, curving posterolaterally to meet cephalic margin at genal corner or faintly anterior to it. Dorsal surface smooth or covered with fine granules.
Occurrence
Arthricocephalus chauveaui has been reported from the Jiangnan Slope Belt of eastern Guizhou and western Hunan, South China, and (as Haliplanktos jishouensis) from North Greenland. In Guizhou and Hunan, it occurs in dark-gray, calcareous mudstone and shale (weathering yellow-green) in the middle part of the Balang Formation (Cambrian Stage 4 or Duyunian according to South China regional stratigraphy). In Greenland, it occurs in the Olenellus Zone of the Henson Gletscher Formation (Cambrian Stage 4).
Description
Exoskeleton ovate, length ~1.5 times width. Cephalon semicircular; glabella forwardly expanding or pestle-shaped, expanded anterior to S2 or S3, with transverse occipital furrow. Four pairs of glabellar furrows present; S1–S3 transverse across glabella, extending to axial furrows and deepened laterally into paired pits near axial furrows, S4 narrow incision, reaching to axial furrow abaxially. Occipital ring wider and slightly shorter than L1, S0 and S1–S4 subequally spaced, frontal lobe slightly longer than L4. Palpebral lobe medium-sized, of ~0.33–0.36 (N=6) of cephalic length in adult specimens, gently curved, moderately to gently oblique and directed moderately forward, with posterior tip in adult specimens opposite S1; palpebral area about three-quarters width of glabella at L2. Anterior branch of facial suture short and straight, diagonally convergent forward; posterior branch strongly divergent for about two-thirds its length, then curving smoothly to meet cephalic margin at genal corner or faintly anterior to it. Librigena narrow, lateral border narrow and upturned, with well-defined border furrow. Anterior cranidial border ridge-like and narrow, upturned, with well-defined border furrow; posterior border slightly shorter than occipital ring, becoming much longer abaxially, with well-developed and moderately long (exsag.) posterior furrow.
Rostral-hypostomal plate (Fig. 2.10) shield-shaped; middle body of hypostoma large, elliptical, gently convex, divided by a faint medial furrow, posterior portion short crescentic; anterior wing triangular, about half as long as hypostome, lateral and posterior borders narrow (tr., sag.), with deep border furrows. Rostral plate narrow, ridge-like, separated from ridge-like cephalic doublure by short connective suture.
Thorax fulcrate, with eight segments. Axis gently narrowing posteriorly, slightly expanded medially, pleural region ~1.5 as broad as axis, fulcrum present about halfway across width (tr.) of pleura; anterior and posterior bands of pleura subequally long (exsag.), separated by deep and long pleural furrow; anterior margin of pleura curves backward to a sharply pointed or acute posterolateral tip.
Pygidium semicircular, as long as cephalon or slightly shorter, length/width ratio ~0.48–0.53 (N=8). Axis consists of five rings and a tiny terminal piece, tapering rapidly rearward, of about two-thirds of pygidial length. Pleural region moderately convex, with five or six pleurae consisting of subequal anterior and posterior bands; pleural furrow nearly straight, well impressed and broad (exsag.); interpleural furrows less well developed, sometimes faint. Lateral and posterior borders upturned; border furrows broad, shallow, of unequal depth due to faintly elevated parts in continuation of the pleural bands. Pygidial doublure narrow, belt-like.
Dorsal surface smooth or covered with fine but densely spaced granules.
Materials
Eight exoskeletons including five in collection LYX-1 (NIGP 164838–164841, 164952), one in collection WWf2 (NIGP 164842), and two in collection XHZA329-13 and XHZA325-27, respectively (NIGP 163349, 163354). Reillustrated specimens include the holotype exoskeleton of Arthricocephalus (Arthricocephalites) pulchellus Zhang and Qian in Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980 (NIGP 38233) and two exoskeletons assigned by Lei Reference Lei(2016) to Arthricocephalus chauveaui Bergeron (NIGP 163346, 163357).
Remarks
A new diagnosis and description of Arthricocephalus chauveaui are presented here because the concept of the species and the genus are significantly revised. Most species that were previously referred to A. (Arthricocephalites), Arthricocephalites, and Haliplanktos are now synonymized with A. chauveaui either because they closely resemble the lectotype exoskeleton of A. chauveaui (E.M. 90001a), or because they closely resemble meraspides of the species.
Material from the Balang Formation of northwestern Hunan and eastern Guizhou shows that the holaspid exoskeleton of Arthricocephalus chauveaui has eight thoracic segments. This suggests that some species referred to Arthricocephalus, A. (Arthricocephalites), and Haliplanktos and the assigned species with fewer than eight thoracic segments are all based on meraspides. Those species are: Arthricocephalus taijiangensis, A. pulchellus, A. (Arthricocephalites) jishouensis (also published as Haliplanktos jishouensis), and A. (Arthricocephalites) tongrenensis. Arthricocephalus taijiangensis (Yin and Li, Reference Yin and Li1978, pl. 156, fig. 8) is based on an individual having five thoracic segments. Arthricocephalus (Arthricocephalites) tongrenensis (Yin and Li, Reference Yin and Li1978, pl. 157, fig. 17) is based on an individual having seven thoracic segments. Holotypes of A. (Arthricocephalites) jishouensis (Zhou et al., Reference Zhou, Liu, Meng and Sun1977, pl. 42, fig. 4) and A. (Arthricocephalites) pulchellus (Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 92, fig. 3) are individuals having six thoracic segments.
Blaker and Peel (Reference Blaker and Peel1997, p. 109) referred the type material of Ovatoryctocara granulata (Chernysheva Reference Chernysheva1962, pl. 5, figs. 6–8) to Arthricocephalus chauveaui. As discussed above, their concept of A. chauveaui embraced both A. chauveaui and Oryctocarella duyunensis. Based on a rather large number of specimens now known from Russia, Greenland, and Newfoundland (e.g., Fletcher, Reference Fletcher2003; Korovnikov and Shabanov, Reference Korovnikov and Shabanov2008; Naimark et al., Reference Naimark, Shabanov and Korovnikov2011; Geyer and Peel, Reference Geyer and Peel2011), it is evident that Ovatoryctocara granulata is distinct from both A. chauveaui and O. duyunensis. It differs in having a prominent preocular field that is relatively long (as compared to the short preocular field in A. chauveaui and O. duyunensis), in having a glabella that is commonly defined by sinuous axial furrows, a pair of longitudinal furrows connecting S1–S3, pit-like glabellar furrows exsagittally, and a palpebral lobe that is moderately close to the glabella. The thorax of O. granulata has only four segments in the holaspid stage, and the pygidium is proportionally longer than that of either A. chauveaui or O. duyunensis; it has more axial rings and lacks a border.
In their synonymy of A. chauveaui, Blaker and Peel (Reference Blaker and Peel1997, p. 110) referred some additional Russian specimens to A. chauveaui. They include a pygidium assigned to Neopagetina nomokonovi by Semashko (Reference Semashko1969, pl. 1, fig.13) and a cranidium and a pygidium referred to Oryctocara (Ovatoryctocara) sp. by Chernysheva (Reference Chernysheva1971, pl. 13, figs. 8, 9). However, the illustrations of these specimens show that they do not belong to either Arthricocephalus, as revised here, or to Oryctocarella. They are more likely specimens of the genus Ovatoryctocara.
As revised, some whole or partial specimens that were previously assigned to A. chauveaui are now referred to Oryctocarella duyunensis Qian, Reference Qian1961 (e.g., Qian, Reference Qian1961; Lu and Qian, Reference Lu and Qian1964; Zhou et al., Reference Zhou, Liu, Meng and Sun1977; Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980; Lane et al., Reference Lane, Blaker and Zhang1988; Blaker and Peel, Reference Blaker and Peel1997; Yuan et al., Reference Yuan, Zhao, Li and Huang2002, Reference Yuan, Zhao and Yang2006; McNamara et al., Reference McNamara, Yu and Zhou2003; Geyer, Reference Geyer2005; Peng et al., Reference Peng, Zhao, Wu, Yuan and Tai2005a, Reference Peng, Zhao, Wu, Yuan and Tai2005b; Reference Peng, Zhao and Yang2006).
Arthricocephalus xinzhaiheensis Qian and Lin in Lu et al., Reference Lu, Zhang, Qian, Zhu, Lin, Zhou, Qian, Zhang and Wu1974

Figure 5 Arthricocephalus xinzhaiheensis Qian and Lin in Lu et al., Reference Lu, Zhang, Qian, Zhu, Lin, Zhou, Qian, Zhang and Wu1974 from the Balang Formation in eastern (1, 2, 6–8) and southeastern (3) Guizhou and northeastern Hunan (4, 5, 9–13), China; all in dorsal view. White arrowhead indicates the posterior margin of thorax; arrowhead with black outline indicates the posterior margin of partially released segment of transitory pygidium; b=border, d=doublure, f=facet: (1) holotype, NIGP 38234, exoskeleton, KH071, original of Lu et al. (Reference Lu, Zhang, Qian, Zhu, Lin, Zhou, Qian, Zhang and Wu1974, pl. 36, fig. 7); (2) NIGP 38235, exoskeleton, KH070, original of Arthricocephalus (Arthricocephalites) xinzhaiheensis Qian and Lin (Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 92, fig. 6); (3) NIGP 21481, exoskeleton, DZ-53-upper, original of Arthricocephalus (Arthricocephalites) intermedius Zhou (Lu et al., Reference Lu, Zhang, Qian, Zhu, Lin, Zhou, Qian, Zhang and Wu1974, pl. 36, fig. 8); (4) NIGP 163350, exoskeleton, XHZA 416-14, original of Lei (Reference Lei2016, fig. 3.7); (5) NIGP 164843, exoskeleton, meraspis degree 6, LYX-1; (6) NIGP 135415, exoskeleton, meraspis degree 5, GY(GPN2), original of Arthricocephalus balangensis (McNamara et al., Reference McNamara, Yu and Zhou2003, pl. 2, fig. 6); (7) NIGP 135416, exoskeleton, meraspis degree 6, GY(GPN2), original of Arthricocephalus balangensis (McNamara et al., Reference McNamara, Yu and Zhou2003, pl. 2, fig. 8); (8) NIGP 135417, exoskeleton, meraspis degree 7, GY(GPN2), original of Arthricocephalus balangensis (McNamara et al., Reference McNamara, Yu and Zhou2003, pl. 2, fig. 9); (9) NIGP 163356, incomplete exoskeleton, meraspis degree 7, XHZA 351-15, original of Lei (Reference Lei2016, fig. 4.13); (10–11) NIGP 163348, holaspid exoskeleton (10), and latex cast from its external mold (11), XHZA404-13, original of Lei (Reference Lei2016, fig. 3.5); (12) NIGP 163365, cephalon with displaced right librigena and lacking left librigena, XHZA416-5, original of Lei (Reference Lei2016, fig. 4.7); (13) NIGP 163362, cranidium, XHZA404-20, original of Lei (Reference Lei2016, fig. 4.4). All scale bars represent 1.0 mm.
1974 Arthricocephalus (Arthricocephalites) xinzhaiheensis Qian and Lin in Reference Lu, Zhang, Qian, Zhu, Lin, Zhou, Qian, Zhang and WuLu et al., p. 95, pl. 36, fig. 7.
1974 Arthricocephalus (Arthricocephalites) intermedius Zhou in Reference Lu, Zhang, Qian, Zhu, Lin, Zhou, Qian, Zhang and WuLu et al., p. 95, pl. 36, fig. 8.
1978 Arthricocephalus (Arthricocephalites) intermedius; Reference Yin and LiYin and Li, p. 441, pl. 156, fig. 10.
1978 Arthricocephalus (Arthricocephalites) xinzhaiheensis; Reference Yin and LiYin and Li, p. 441, pl. 157, fig. 1.
1980 Arthricocephalus (Arthricocephalites) xinzhaiheensis; Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and YuanZhang et al. (part), p. 276, pl. 92, figs. 5, 6; non pl. 93, fig. 1.
1980 Arthricocephalus (Arthricocephalites) intermedius; Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and YuanZhang et al., p. 278, pl. 92, fig. 4.
?1980 Arthricocephalites (Arthricocephalites) sp. 2, Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and YuanZhang et al., p. 279, pl. 93, fig. 8.
2002 Arthricocephalites xinzhaiheensis; Reference Yuan, Zhao, Li and HuangYuan et al., p. 123, pl. 30, figs. 6, 7.
non 2003 Arthricocephalus xinzhaiheensis; Reference McNamara, Yu and ZhouMcNamara et al., p. 111, pl. 2, figs. 1–4, text-fig. 4A–D.
2006 Arthricocephalites xinzhaiheensis; Reference Yuan, Zhao and YangYuan et al. (part), p. 617, fig 2a; non fig. 2b.
2016 Arthricocephalites xinzhaiheensis; Reference Shen, Peng, Wen and LiuShen et al., (part), fig. 1.1–1.7; non fig. 1.8–1.11.
2016 Arthricocephalus chauveaui Bergeron; Reference LeiLei (part), p. 496, figs. 3.5, 3.7, 3.13, 4.2, 4.7; non figs. 3.1–3.4, 3.6, ?3.8, 3.9–3.12, 3.14, ?3.15, 4.1–4.3, 4.5, 4.6, 4.8, ?4.9–4.16.
Holotype
Exoskeleton (NIGP 38234, Fig. 5.1) from the Balang Formation, Xinzhaihe, Songtao, northeastern Guizhou, China; by monotypy. The holotype was figured by Lu et al. (Reference Lu, Zhang, Qian, Zhu, Lin, Zhou, Qian, Zhang and Wu1974, pl. 36, fig. 7), and refigured by Yin and Li (Reference Yin and Li1978, pl. 157, fig. 1), Zhang et al. (Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 92, fig. 5), and Yuan et al. (Reference Yuan, Zhao, Li and Huang2002, pl. 30, fig. 6). It is not the exoskeleton illustrated by Yuan et al. (Reference Yuan, Zhao and Yang2006, fig. 2a) as the holotype of the species.
Diagnosis
Arthricocephalus with glabellar furrows S1–S3 pit-like, connected by weak transverse furrows, separated from axial furrow; palpebral lobe variably oblique in meraspis and holaspis with posterior end at a level corresponding to the posterior half or mid-length of L2; dorsal surface smooth or covered by densely spaced granules.
Occurrence
Arthricocephalus xinzhaihenesis co-occurs with A. chauveaui in the Balang Formation of the Jiangnan Slope Belt, eastern Guizhou and western Hunan, South China. It is present in dark-gray, calcareous mudstone and shale (weathering yellow-green) in the middle part of the Balang Formation (Cambrian Stage 4 or Duyunian according to South China regional stratigraphy).
Materials
One exoskeleton (NIGP 164843) in collections LYX-1. Reillustrated specimens include the holotype exoskeleton of Arthricocephalus (Arthricocephalites) xinzhaiheensis Qian and Lin in Lu et al., Reference Lu, Zhang, Qian, Zhu, Lin, Zhou, Qian, Zhang and Wu1974 (NIGP 38234), the holotype exoskeleton of A. (Arthricocephalites) intermedius Zhou in Lu et al., Reference Lu, Zhang, Qian, Zhu, Lin, Zhou, Qian, Zhang and Wu1974 (NIGP 21481), an exoskeleton assigned to A. (Arthricocephalites) xinzhaiheensis by Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980 (NIGP 38235); three exoskeletons assigned to Arthricocephalus balangensis Lu and Qian in Yin and Li, Reference Yin and Li1978 (not Qian and Lin, as documented in Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980) by McNamara et al., Reference McNamara, Yu and Zhou2003 (NIGP 135415–135417); and four specimens assigned to Arthricocephalus chauveaui Bergeron by Lei (Reference Lei2016), including two exoskeletons (NIGP 163348, 163356), an incomplete cephalon (NIGP 163365), and an incomplete cranidium (NIGP 163362).
Remarks
Lei (Reference Lei2016) suppressed Arthricocephalus xinzhaiheensis as a junior synonym of A. chauveaui. In most respects, such as shape of the glabella, morphology of the glabellar furrows, course of the facial suture, presence of thoracic fulcra, shape and subdivision of the pleurae, segmentation of the thorax and pygidium, faint interpleural furrows on the pygidium, and the upturned cephalic and pygidial borders, A. xinzhaiheensis closely resembles A. chauveaui. However, the species is distinct from A. chauveaui in having S1–S3 glabellar furrows that do not extend abaxially to the axial furrow, although they are joined by weak transverse furrows medially on the glabella. The palpebral lobe of A. xinzhaiheensis seems to be more oblique in adults, and the anterior tip of the palpebral lobe appears to be closer to the glabella than it is in A. chauveaui and the posterior tip places further back. In the holotype of A. xinzhaiheensis the axis of the thorax (Fig. 5.1) is proportionately broader than in A. chauveaui, but another specimen (Fig. 5.5) suggests variation in this character. In addition, the specimen in Figure 5.5 is indistiguishable from A. chauveaui in the proportion of the thoracic axis. In A. xinzhaiheensis the forward expansion of the glabella is variable.
Specimens assigned to Arthricocephalus balangensis by McNamara et al. (Reference McNamara, Yu and Zhou2003, pl. 2, figs. 6, 8, 9, text-fig. 5; Fig. 5.6, 5.7) differ significantly from the holotype of A. balangensis and are here transferred to A. xinzhaiheensis based on the morphology of the glabella and glabellar furrows, and on the presence of fulcra in the thorax. The holotype of A. balangensis (Yin and Li, Reference Yin and Li1978, pl. 157, fig. 10; refigured by Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 93, fig. 2; Fig. 7.1, 7.2) is similar to A. xinzhaiheensis in having pit-like S1–S3 joined by transverse furrows across the middle of the glabella. However, the glabella of A. balangensis is subrectangular or subcylindrical in shape, and defined by nearly straight, parallel axial furrows. Importantly, the thorax of A. balangensis lacks fulcra. The thorax normally has 11 segments, but in some examples one more segment may be developed. These characters support a reassignment of A. balangensis to Oryctocarella (and recombination as Oryctocarella balangensis, see below).
Specimens assigned to Arthricocephalus xinzhaihenesis by McNamara et al. (Reference McNamara, Yu and Zhou2003, pl. 2, figs. 1–4, text-fig. 4A–D) are reassigned here as Oryctocarella balangensis (see below), and specimens assigned to A. xinzhaihenesis by Yuan et al., Reference Yuan, Zhao and Yang2006 (fig. 2b) are reassigned here as Arthricocephalus chauveaui. The specimens illustrated by McNamara et al. (Reference McNamara, Yu and Zhou2003), from Palang, Duyun, southeastern Guizhou, are all meraspid exoskeletons; the largest one has only seven thoracic segments (McNamara et al., Reference McNamara, Yu and Zhou2003, pl. 2, fig. 4; Fig. 5.8). In one exoskeleton assigned to A. xinzhaihenesis by Yuan et al. (Reference Yuan, Zhao and Yang2006), the glabellar furrows S1–S3 are transglabellar and reach to the axial furrows. This character is regarded as indicative of A. chauveaui.
Shen et al. (Reference Shen, Peng, Wen and Liu2016) referred numerous specimens to the genus Arthricocephalites, either as Arthricocephalites xinzhaihenesis or as A. intermedius. Most of these specimens, in fact, represent Arthricocephalus chauveaui because their glabellar furrows S1 through S3 are transglabellar.
Arthricocephalites (Arthricocephalites) sp. 2 (Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 93, fig. 8) is questionably assigned to Arthricocephalus. It has a glabella with pit-like glabellar furrows and a fulcrate thorax with facets. However, the interpleural furrows in the pygidium are clearly incised, making it difficult to confidently assign the specimen to a certain species.
As discussed above, S1 through S3 are isolated from the axial furrows in Arthricocephalus xinzhaihenesis, unlike the transglabellar development of S1 through S3 in Arthricocephalus chauveaui. In oryctocephalids, the nature of S1 through S3 appears to be a critical specific character. In addition, the palpebral lobes and ocular ridges of A. xinzhaihenesis are more obliquely directed to the length of axis than those of A. chauveaui, suggesting that A. xinzhaihenesis is a species separate from A. chauveaui. However, the differences between both species in other features seem minimal, and both species share almost the same stratigraphic occurrences as well. For these reasons, the possibility that they represent sexual dimorphs of a single species can not be ruled out.
Genus Oryctocarella Tomashpolskaya and Karpinski, Reference Tomashpolskaya and Karpinski1961
1961 Oryctocarella Reference Tomashpolskaya and KarpinskiTomashpolskaya and Karpinski, p. 156.
1988 Arthricocephalus Bergeron; Reference Lane, Blaker and ZhangLane et al., p. 555 (part).
Type species
Oryctocara sibirica Tomashpolskaya in Khalfin, Reference Khalfin1960 (p. 199, pl. 23, fig. 5), from the lower part of the traditional middle Cambrian, Batenevsky Ridge, Kuznetsk Alatau, Altay-Sayan Foldbelt, Russia; by original designation.
Other species
Arthricocephalus duyunensis Qian, Reference Qian1961 (p. 97, pl. 1, fig. 19, pl. 2, figs. 5, 7–10; non pl. 1, fig. 20, pl. 2, fig. 6 [=Arthricocephalus chauveaui Bergeron, Reference Bergeron1899]), from the Balang Formation, Palang, Duyun, eastern Guizhou, China; Arthricocephalus (Arthricocephalites) balangensis Qian and Lin in Yin and Li, Reference Yin and Li1978 (p. 442, pl. 157, fig. 10), from the Balang Formation, Palang, Duyun, eastern Guizhou, China.
Diagnosis
Micropygous genus of Oryctocarinae with gently curved or transverse anterior margin, surface sculpture granulated or smooth. Glabella subrectangular to cylindrical, defined laterally by nearly straight axial furrows; S1–S4 pit-like or narrow incision, with S1–S3 distant from axial furrow, connected by weak or faint transverse furrows. Palpebral lobe notably oblique, located near or anterior to glabellar mid-length, palpebral area subequal in width with glabella. Facial suture proparian. Thorax non-fulcrate, consisting of 9–11, or rarely 12, segments; axis about half as wide as pleural region. Pygidium much shorter than cranidium, axis about half of pygidial length, with three rings and a tiny terminal piece; interpleural furrows well defined; borders absent, margin may have a posteromedial notch.
Remarks
Oryctocarella is here regarded as a valid genus. Differences between the type species of Oryctocarella and Arthricocephalus are discussed at length above, and differences between A. chauveaui and O. duyunensis are illustrated in Figure 3 . Oryctocarella is distinguished from Arthricocephalus in having a pygidium that is much shorter than the cephalon; a cylindrical glabella; pit-like glabellar furrows, most of which do not reach to the axial furrows; a larger number of thoracic segments; and well-defined interpleural furrows in the pygidium. In addition, the thoracic segments lack fulcra, and a pygidial border is not clearly developed in Oryctocarella.
Oryctocarella sibirica (Tomashpolskaya in Khalfin, Reference Khalfin1960)

Figure 6 Oryctocarella sibirica (Tomashpolskaya in Khalfin, Reference Khalfin1960), topotypes from the lower part of the Karasuk Formation, Region of Dolgii Mys range (DM), Kuznetsk Alatau, Altay-Sayan Foldbelt, Russia; all in dorsal view. White arrowhead indicates the posterior margin of thorax; arrowhead with black outline indicates the posterior margin of partially released segment of transitory pygidium; ltr=last thoracic axial ring; 9=ninth thoracic segment: (1) neotype CSGM2075/2, proposed here, incomplete cranidium, original specimen of Korovnikov and Novozhilova (Reference Korovnikov and Novozhilova2017, pl. 5, fig. 2); (2) CSGM2075/17, incomplete cranidium; (3) CSGM2075/16, latex cast from external mold of incomplete cranidium; (4) CSGM 2075/5, incomplete exoskeleton, meraspis degree 8, original specimen of Korovnikov and Novozhilova (Reference Korovnikov and Novozhilova2017, pl. 5, fig. 5); (5) CSGM2075/6, thoracopygon, holaspid?, with nine thoracic segments, original specimen of Korovnikov and Novozhilova (Reference Korovnikov and Novozhilova2017, pl. 5, fig. 6); (6) enlarged posterior part of CSGM2075/6 in 5, showing the broken last thoracic axial ring (ltr) on the ninth segment. All scale bars represent 1.0 mm.
1960 Oryctocara sibirica Tomashpolskaya in Reference KhalfinKhalfin, p. 199, pl. Cm-23, fig. 5.
1961 Oryctocarella sibirica; Reference Tomashpolskaya and KarpinskiTomashpolskaya and Karpinski, p. 156, pl. 1, figs. 1–6.
2015 Oryctocarella sibirica; Reference Peng, Zhu, Babcock and KorovnikovPeng et al., p. 89, pl. 1, fig. 5.
2017 Oryctocarella sibirica; Reference Korovnikov and NovozhilovaKorovnikov and Novozhilova, p. 267, pl. 5, figs. 1–7; text-fig. 3.a, 3.b.
Neotype
The original type material, including the holotype (an exoskeleton) and more than 10 additional variously complete exoskeletions, of which five were figured by Tomashpolskaya and Karpinski (Reference Tomashpolskaya and Karpinski1961, pl. 1, figs. 1, 3–6), is apparently lost. An incomplete cranidium, CSGM 2075/2 (Fig. 6.1) from a recent collection of topotypes, is designated as the neotype (see discussion below).
Diagnosis
Oryctocarella with subrectangular or cylindrical glabella; preglabellar field absent; palpebral ridge faint, close to anterior border furrow; palpebral lobe located anterior to glabellar mid-length, with posterior tip opposite mid-length of L2 (~34% of cephalic length); palpebral area narrower than glabella; posterior section of facial suture nearly transverse; cranidial posterior border widens abaxially. Thorax with nine segments; anterior pleural band equal to or slightly shorter than posterior band. Pygidial margin may have slight inward curvature medially, but lacks medial notch. Dorsal surface densely granulose.
Occurrence
Known only from outcrop 863, Region of Dolgii Mys Mountain, Batenevsky Ridge, Kuznetsk Alatau, Altay-Sayan Foldbelt, southwestern Siberia, Russia; basal Amgan Stage (traditional middle Cambrian) of the Siberian regional stratigraphy, corresponding to the Cambrian Stage 4.
Materials
Two unfigured cranidia (CSGM2075/16, CSGM 2075/17) and three specimens illustrated by Korovnikov and Novozhilova (Reference Korovnikov and Novozhilova2017), which include an incomplete cranidium (CSGM2075/2), an incomplete exoskeleton (CSGM2075/5), and a thoracopygon (CSGM2075/6); all in a single collection from the basal Amgan Stage of Siberian regional stratigraphy (provisional Cambrian Stage 4), Region of Dolgii Mys (Long Cape) Mountain, Batenevsky Ridge, Kuznetsk Alatau, Altay-Sayan Foldbelt, Russia, the type locality of the species.
Remarks
The type species of Oryctocarella, Oryctocara sibirica, was originally based on a single exoskeleton, the holotype (Tomashpolskaya in Khalfin, Reference Khalfin1960, p. 199, pl. 23, fig. 5, No. 863(1)58; refigured in Tomashpolskaya and Karpinski, Reference Tomashpolskaya and Karpinski1961, pl. 1, fig. 2) from the traditional lower middle Cambrian, Region of Dolgii Mys Mountain, Batenevsky Ridge, Kuznetsk Alatau, southwestern Siberia, Russia; by monotypy. When Tomashpolskaya and Karpinski (Reference Tomashpolskaya and Karpinski1961) erected the genus Oryctocarella, with Oryctocara sibirica as its type species, additional specimens from the same collection as the holotype were illustrated (Tomashpolskaya and Karpinski, Reference Tomashpolskaya and Karpinski1961, pl. 1, figs. 1, 3–6). At least 10 variously complete exoskeletons were in the collection with the holotype (see Tomashpolskaya and Karpinski, Reference Tomashpolskaya and Karpinski1961). The original material, which was housed at the Geological Museum of the Tomsk Polytechnic Institute (TPI, now Tomsk Polytechnic University), now appears to be lost. A search on our request by I. Korovnikov, Novosibirsk through the collections at Tomsk Polytechnic University, failed to recover any material of O. sibirica from Tomashpolskaya’s original collection. Because of the absence of the original material, a neotype (Fig. 6.1) is selected from a collection of topotypes collected by I. Korovnikov from Tomashpolskaya’s original locality in the Dolgii Mys Mountain region. This new material (Korovnikov and Novozhilova, Reference Korovnikov and Novozhilova2017, pl. 5, figs. 1–7; Fig. 6), although not well preserved, is sufficient to demonstrate most morphological characters of O. sibirica, and that Oryctocarella should be regarded as a valid genus.
Oryctocarella sibirica was originally reported to have eight thoracic segments. One of the topotypes (Fig. 6.5, 6.6) shows that the thorax contains at least nine segments. In this specimen the axial ring of the ninth segment is present, but broken.
Differences between the type species of Oryctocarella, O. sibirica, and other species now referred to the genus, Oryctocarella balangensis (Qian and Lin in Yin and Li, Reference Yin and Li1978) and O. duyunensis (Qian, Reference Qian1961), are discussed below.
Oryctocarella balangensis (Lu and Qian in Yin and Li, Reference Yin and Li1978)

Figure 7 Oryctocarella balangensis (Lu and Qian in Yin and Li, Reference Yin and Li1978) from the Balang Formation in southeastern (1–3, 5) and eastern (4, 6–13) Guizhou, South China; all in dorsal view. White arrowhead indicates the posterior margin of thorax; arrowhead with black outline indicates the posterior margin of partially released segment of transitory pygidium; b=border, d=doublure: (1) holotype NIGP 38240, incomplete exoskeleton, original of Lu and Qian in Yin and Li (Reference Yin and Li1978, pl. 157, fig.10), GY210; (2) enlargement of part of cephalon in 1; (3) NIGP 38241, cephalon previously illustrated by Zhang et al. (Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 93, fig. 3), GY222; (4) NIGP 164947, incomplete cranidium on same slab as specimen in 8, XS-Xiao152; (5) NIGP 11485, cranidium previously illustrated by Qian (Reference Qian1961, pl. 1, fig. 20) as Arthricocephalus chauveaui, GY206; (6) NIGP 11492, cranidium previously illustrated by Qian (Reference Qian1961, pl. 2, fig. 6) as Arthricocephalus chauveaui, GY204; (7) NIGP 38239, cranidium previously illustrated by Zhang et al. (Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 93, fig. 1) as Arthricocephalus (Arthricocephalites) xinzhaiheensis, GY214; (8) NIGP 38244, cranidium previously illustrated by Zhang et al. (Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 93, fig. 7) as one of two ‘holotypes’ of Arthricocephalus (Arthricocephalites) xiaosaiensis Qian, retrodeformation with inferred strain ellipse, XS-Xiao152; (9) NIGP 38248, cranidium previously illustrated by Qian in Zhang et al. (Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 94, fig. 3) as one of two ‘holotypes’ of Arthricocephalus (Arthricocephalites) xiaosaiensis Qian, XS-Xiao152; (10) NIGP 38251, pygidium with six thoracic segments, previously illustrated by Zhang et al. (Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 94, fig. 6) as Arthricocephalus (Arthricocephalites) xiaosaiensis, XS-Xiao152; (11, 13) NIGP 164948, external mold of incomplete exoskeleton, two views, reversed images, XS-Xiao152, in association with the pygidium in 10; (12) NIGP 164949, reversed image of external mold of thoracopygon, XS-Xiao152, on same slab as the NIGP 38251 pygidium in 10 and thoracopygon in 11. All scale bars represent 1.0 mm.
1961 Arthricocephalus duyunensis Reference Zhao, Yu, Yuan, Yang and GuoQian (part), p. 97, pl. 1, fig. 20, pl. 2, fig. 6; non pl. 1, fig. 19, pl. 2, figs. 5, 7, 8, 10, ?fig. 9.
1978 Arthricocephalus (Arthricocephalites) balangensis Lu and Qian in Reference Yin and LiYin and Li, p. 442, pl. 157, fig. 10.
1980 Arthricocephalus (Arthricocephalites) xinzhaiheensis Qian; Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and YuanZhang et al., p. 276, pl. 93, fig. 1; non pl. 92, figs. 5, 6.
1980 Arthricocephalus (Arthricocephalites) balangensis; Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and YuanZhang et al., p. 277, pl. 93, figs. 2, 3.
1980 Arthricocephalus (Arthricocephalites) xiaosaiensis Qian in Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and YuanZhang et al. (part), p. 278, pl. 93, fig. 7, pl. 94, figs. 3, 4, 6; non fig. 5 (?=Palaeolenus fengyanensis Zhu, Reference Zhu1962).
?2003 Arthricocephalus xinzhaiheensis; Reference McNamara, Yu and ZhouMcNamara et al., p. 111, pl. 2, figs. 1–4, text-fig. 4A–D.
non 2003 Arthricocephalus balangensis; Reference McNamara, Yu and ZhouMcNamara et al., p. 114, pl. 2, figs. 6, 8, 9, text-fig. 5.
Holotype
Incomplete exoskeleton (Yin and Li, Reference Yin and Li1978, pl. 157, fig. 10, NIGP 38240; reillustrated by Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 93, figs. 2; Fig. 7.1, 7.2) from the Balang Formation, Palang, Duyun, southeastern Guizhou, China; by monotypy.
Diagnosis
Oryctocarella with glabella subrectangular or cylindrical in outline; axial furrows parallel-sided or gently constricted medially; S0 and S1–S3 furrows pit-like, connected by faint transverse furrows; palpebral lobe located at about midlength of glabella, ~0.29–0.31 (N=6) of cephalic length, posterior end opposite posterior one-third of L2; palpebral ridge close to anterior cranidial border furrow; posterior branch of facial suture cutting lateral cephalic border opposite S1 or midlength of L1. Thorax non-fulcrate, with 11 or possibly 12 segments; axis three-quarters width of pleural region. Pygidial axis composed of three rings and a tiny terminal piece; posterior margin without a medial notch. Dorsal surface smooth or covered with fine granules.
Occurrence
From yellow-green calcareous shale in the middle part of the Balang Formation, eastern Guizhou, South China (Jiangnan Slope Belt); Cambrian Stage 4, Duyunian Stage of the regional stratigraphic scheme.
Materials
Three associated sclerites (unfigured), including an incomplete exoskeleton, a thoracopygon, and an incomplete cranidium (NIGP 164947–164949), from the Balang Formation near Xiaosai, Yuqing, eastern Guizhou, South China; refigured sclerites including the incomplete holotype exoskeleton of Arthricocephalus (Arthricocephalites) balangensis (Lu and Qian in Yin and Li, Reference Yin and Li1978) (NIGP 38240); a cephalon illustrated as Arthricocephalus (Arthricocephalites) balangensis by Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980 (NIGP 38241); two cranidia assigned as Arthricocephalus chauveaui by Qian, Reference Qian1961 (NIGP 11485, NIGP 11492); three incomplete cranidia, one of them assigned to Arthricocephalus (Arthricocephalites) xinzhaiheensis by Qian in Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980 (NIGP 38239), and two “holotypes” designated for the new species Arthricocephalus (Arthricocephalites) xiaosaiensis Qian in Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980 (NIGP 38244, NIGP 38248); one pygidium assigned as Arthricocephalus (Arthricocephalites) xiaosaiensis Qian in Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980 (NIGP 38251); and an unfigured incomplete cranidium in association with NIGP 38244.
Remarks
This species, originally introduced as Arthricocephalus (Arthricocephalites) balangensis, is now transferred to Oryctocarella. It was referred to Arthricocephalus (Arthricocephalites) principally because its glabella was thought to be expanded forward anterior to S4 (Yin and Li, Reference Yin and Li1978, p. 442; Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, p. 277). Our new observations indicate that this species does not belong to A. (Arthricocephalites) (a junior synonym of Arthricocephalus), but to Oryctocarella. Examination of the holotype (Fig. 7.1, 7.2) shows that the glabella is subrectangular in outline, defined by nearly straight axial furrows, and is not expanded anteriorly. A cephalon illustrated by Zhang et al. (Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 93, fig. 3; Fig. 7.3) indicates an expansion of the anterior part of the glabella, but this is clearly an artefact of compaction. The axial furrows on this specimen are not symmetrical to the axis, providing evidence of differential distortion.
Other characters further support assignment of this species to Oryctocarella. The thorax of this species was originally described as having at least 10 segments. However, new material shows that the thorax consists of eleven, or possibly twelve segments (Fig. 7.11, 7.12). The thoracic segments lack clear fulcra. This species is better characterized by pit-like glabellar furrows that are separated from the axial furrows, but connected by faint to obsolescent transverse furrows. The occipital furrow is weak and deepens into pits in a short distance before reaching to the axial furrow. The facial suture is proparian.
This species shows variability in surface sculpture. Most specimens do not show a prosopon. One cephalon (Fig. 7.3) assigned to the species by Zhang et al. (Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 93, fig. 3), and two cranidia (Fig. 7.5, 7.6), previously assigned to A. chauveaui by Qian (Reference Qian1961, pl. 1, fig. 20, pl. 2, fig. 6), bear fine, densely spaced granules.
Exoskeletons of meraspides referred to Arthricocephalus xinzhaiheensis by McNamara et al. (Reference McNamara, Yu and Zhou2003, pl. 2, figs. 1–4) are questionably assigned to Oryctocarella balangensis herein. The largest of these specimens (McNamara et al., Reference McNamara, Yu and Zhou2003, pl. 2, fig. 4) is of a meraspid degree 8. It has a cylindrical glabella with pit-like S1–S4, a proparian facial suture (with the right hand librigena somewhat displaced), and a relatively broad thoracic axis. All these features are consistent with assignment to O. balangensis. However, the interpleural furrows in the pygidium are poorly defined, and this character is more reminiscent of Arthricocephalus.
Two cranidia (Fig. 7.5, 7.6), previously assigned to Arthricocephalus chauveaui by Qian (Reference Qian1961, pl. 1, fig. 20, pl. 2, fig. 6), are here transfered to O. balangensis. They were collected from the same section as the holotype of A. balangensis at Palang, Duyun, and are morphologically indistinguishable from the cranidium of the holotype. McNamara et al. (Reference McNamara, Yu and Zhou2003, p. 114) correctly referred these cranidia to Arthricocephalus balangensis (now O. balangensis), although those authors used a broader concept for A. balangensis, which included some specimens (McNamara et al., Reference McNamara, Yu and Zhou2003, pl. 2, figs. 6, 8, 9) here reassigned to Arthricocephalus xinzhaiheensis.
A cranidium from the Balang Formation at Palang, which was assigned to Arthricocephalus (Arthricocephalites) xinzhaiheensis by Zhang et al. (Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 94, fig. 1; Fig. 7.7) apparently suffered transverse compression and is also included in O. balangensis herein.
Arthricocephalus (Arthricocephalites) xiaosaiensis Qian in Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980 (p. 278, pl. 93, fig. 7, pl. 94, figs. 3–6; Fig. 7.8–7.10), from the Xiaosai section, Yuqing, eastern Guizhou, is a junior synonym of O. balangensis because the morphology of its doubly designated ‘holotype’ cranidia falls within the morphological plasticity of the cranidia of O. balangensis. The non-fulcrate thorax and micropygous pygidium in the incomplete thoracopygon that is associated with the ‘holotypes’ (Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 94, fig. 6; Fig. 7.10) also show that the species belongs to Oryctocarella. Additional figured specimens of the species (Fig. 7.11–7.13) from the same collection show a non-fulcrate thorax and micropygous pygidium as well.
One thoracopygon from the Xiaosai section, eastern Guizhou (Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 94, fig. 5) is not conspecific with other specimens assigned to A. (Arthricocephalites) xiaosaiensis (=O. balangensis) because of the presence of fulcra in the thoracic segments. This specimen is tentatively reassigned to Palaeolenus fengyanensis Zhu, Reference Zhu1962, a species that co-occurs with O. balangensis in collection XS-Xiao152 (Zhang et al., Reference Zhang, Yuan, Zhou, Qian and Wang1979, p. 92).
Oryctocarella balangensis resembles O. sibirica in the shape of the glabella, the morphology of the glabellar furrows, and the location of the palpebral lobe. It differs from the type species of the genus in having less well defined transverse furrows in the medial area of the glabella, and in having a smooth or finely granulated dorsal surface. These characters also differentiate O. balangensis from O. duyunensis, which bears a strong and dense granulation. Among these three species, only O. duyunensis is known to have a distinct posteromedial notch on the pygidial margin. Other, more subtle characters that seem to differentiate O. balangensis from O. sibirica are a more clearly defined palpebral ridge and a less divergent posterior branch of the facial suture in the former species.
Oryctocarella duyunensis (Qian, Reference Qian1961)
Figures 1.1–1.7, 2.4–2.9, 3.2, 8, 9

Figure 8 Oryctocarella duyunensis (Qian, Reference Qian1961) from the Balang Formation in northwestern Hunan (1–15) and eastern Guizhou (16–18), China; all in dorsal view. White arrowhead indicates the posterior margin of thorax; arrowhead with black outline indicates the posterior margin of partially released segment of transitory pygidium; black triangle indicates medial notch at pygidial margin. (1) NIGP 164844, exoskeleton, meraspis degree 2, MZC-1; (2) NIGP 164845, exoskeleton, meraspis degree 4, MZC-1; (3) NIGP 164846, exoskeleton, meraspis degree 8, MZC-1; (4) NIGP 164847, exoskeleton, meraspis degree 9, MZC-1; (5) NIGP 164848, exoskeleton, meraspis degree 9, MZC-1; (6) NIGP 164849, exoskeleton, meraspis degree 9, MZC-1; (7) NIGP 164850, exoskeleton, meraspis degree 9, MZC-1; (8) NIGP 164851, exoskeleton, holaspid, thorax with possibly twelve segments, MZC-1; (9) NIGP 164852, cranidium with one thoracic segment, meraspis, MZC-1; (10) NIGP 164853, cranidium with two thoracic segments, meraspis, MZC-1; (11) NIGP 164854, cranidium with one thoracic segment, meraspis, MZC-1; (12) NIGP 164855, transitory pygidium with the anterior segment almost completely released, MZC-1; (13) NIGP 164856, transitory pygidium with the anterior segment partly released, MZC-1; (14) NIGP 164857, pygidium, holaspid, MZC-1; (15) NIGP 164858, exoskeleton, meraspis degree 9, MZC-1; (16) NIGP 38246, exoskeleton, meraspis degree 10, holotype of Arthricocephalus granulus Qian and Lin in Zhou et al., Reference Zhou, Liu, Meng and Sun1977 (pl. 42, fig. 2), retrodeformation with inferred strain ellipse, KH020; (17) holotype NIGP 11494, exoskeleton, meraspis degree 9, originally assigned as Arthricocephalus duyunensis Qian, Reference Qian1961 (pl. 2, fig. 8), GY207; (18) NIGP 38236, exoskeleton, meraspis degree 9, holotype of Arthricocephalus horridus Qian and Lin in Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980 (pl. 92, fig. 7), GY203. All scale bars represent 1.0 mm.

Figure 9 Oryctocarella duyunensis (Qian, Reference Qian1961) from the Balang Formation at Bulin and Mozichong, Huayuan, northwestern Hunan, China; all in dorsal views unless stated otherwise. White arrowhead indicates the posterior margin of thorax; arrowhead with black outline indicates the posterior margin of partially released segment of transitory pygidium; black triangle indicates posteromedial notch; d=doublure: (1) NIGP 164859, exoskeleton, meraspis degree 6, BL-1; (2) NIGP 164860, exoskeleton, meraspis degree 7, BL-1; (3) NIGP 164861, exoskeleton, meraspis degree 7, BL-1; (4) NIGP 164862, exoskeleton, meraspis degree 7, BL-1; (5) NIGP 164863, exoskeleton, meraspis degree 9, MZC-2; (6) NIGP 164864, exoskeleton, meraspis degree 9, BL-1; (7) NIGP 164865, exoskeleton with detached cranidium, meraspis degree 9, BL-1; (8) retrodeformation of specimen in 9 with inferred strain ellipse; (9), NIGP 164866, exoskeleton, meraspis degree 10, BL-1; (10) retrodeformation of specimen in 11 with inferred strain ellipse; (11) NIGP 164867, exoskeleton, meraspis degree 9, BL-1; (12), retrodeformation of specimen in 13 with inferred strain ellipse; (13) NIGP 164868, exoskeleton, meraspis degree 9, BL-1; (14) retrodeformation of specimen in 15 with inferred strain ellipse; (15) NWU-DYXJT 1823, exoskeleton, holaspid, MZC-1; (16) NIGP 164869, metaprotaspis with distinctly divided axis, distinct eye ridge and dot-like palpebral lobe, BL-1; (17, 18) NIGP 164870, external mold of exoskeleton, ventral view, meraspis degree 10 (17), and enlargement of hypostome (18), BL-1. All scale bars represent 1.0 mm.
1961 Arthricocephalus duyunensis Reference QianQian (part), p. 97, pl. 1, fig. 19, pl. 2, figs. 5, 7, 8, 10, ?fig. 9; non pl. 1, fig. 20, pl. 2, fig. 6.
1964 Arthricocephalus chauveaui Bergeron, Reference Bergeron1899; Lu and Qian, p. 26, pl. 1, fig. 5.
1965 Arthricocephalus duyunensis; Reference Lu, Zhang, Zhu, Qian and XiangLu et al., p. 108, pl. 17, figs. 2, 3, 5, non 4.
1974 Arthricocephalus duyunensis; Reference Lu, Zhang, Qian, Zhu, Lin, Zhou, Qian, Zhang and WuLu et al., p. 95, pl. 36, fig. 9.
1977 Arthricocephalus granulus Qian and Lin in Reference Zhou, Liu, Meng and SunZhou et al., p. 130, pl. 42, figs. 1, 2.
1977 Arthricocephalus chauveaui; Reference Zhou, Liu, Meng and SunZhou et al., p. 130, pl. 42, fig. 3.
1978 Arthricocephalus duyunensis; Reference Yin and LiYin and Li, p. 440, pl. 155, fig. 10.
1978 Arthricocephalus jiangkouensis Yin in Reference Yin and LiYin and Li, p. 440, pl. 155, figs. 15, 16.
1978 Arthricocephalus (Arthricocephalites) granulus Qian and Lin; Reference Yin and LiYin and Li, p. 441, pl. 157, fig. 2.
1980 Arthricocephalus chauveaui; Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and YuanZhang et al., p. 275, pl. 92, figs. 1, 2.
1980 Arthricocephalus horridus Qian and Lin in Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and YuanZhang et al., p. 275, pl. 92, figs. 7–9.
1980 Arthricocephalus (Arthricocephalites) granulus; Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and YuanZhang et al., p. 277, pl. 90, fig. 8, pl. 93, fig. 4, pl. 94, figs. 1, 2.
1982 Arthricocephalus granulus; Reference LiuLiu, p. 300, pl. 214, figs. 1, 9.
?1983 Arthricocephalus (Arthricocephalites) granulus; Reference JuJu, p. 632, pl. 2, figs. 12, 13.
1983 Arthricocephalus (Arthricocephalites) sp., Reference JuJu, pl. 2, figs. 14, 15.
?1985 Arthricocephalus tenuis Reference Zhang and ZhouZhang and Zhou, p. 263, pl. 2, figs. 10–12.
1985 Arthricocephalus duyunensis; Reference Zhang and ZhouZhang and Zhou, p. 263, pl. 2, figs. 8, 9.
1985 Arthricocephalus sp., Reference Zhang and ZhouZhang and Zhou, p. 263, pl. 2, fig. 14.
1988 Arthricocephalus chauveaui; Reference Lane, Blaker and ZhangLane et al. (part), p. 555, pl. 1, figs. 2, 3, 6; non figs. 1, 4, 5.
1997 Arthricocephalus chauveaui; Reference Blaker and PeelBlaker and Peel (part), p. 112, figs. 62.2, 62.3, 63, 64.1–64.4; non fig. 62.1.
2001a Arthricocephalus granulus; Reference Zhao, Yu, Yuan, Yang and GuoZhao et al., p. 180, pl. 1, figs. 4–6.
2002 Arthricocephalus chauveaui; Reference Yuan, Zhao, Li and HuangYuan et al., p. 121, pl. 31, fig. 1.
2003 Arthricocephalus chauveaui; Reference McNamara, Yu and ZhouMcNamara et al., p. 106, pl. 1, figs. 1–15, pl. 2, figs. 5, 7, text-fig. 3A–K.
2005 Arthricocephalus chauveaui; Geyer, Reference Geyer2005 (part), p. 87, fig. 3.1–3.4, 3.8, 3.9; non fig. 3.7.
2005a Arthricocephalus chauveaui; Reference Peng, Zhao, Wu, Yuan and TaiPeng et al., p. 1161, fig. 2e.
2005b Arthricocephalus chauveaui; Reference Peng, Zhao, Wu, Yuan and TaiPeng et al., p. 1056, fig. 2e.
2006 Arthricocephalus chauveaui; Reference Yuan, Zhao and YangYuan et al., p. 615, fig. 1d, 1e.
2006 Arthricocephalus chauveaui; Reference Peng, Zhao and YangPeng et al., p. 241, pl. 1, fig. 11.
2006 Arthricocephalus granulus; Reference Yuan, Zhao and YangYuan et al., p. 616, fig. 1b, 1c.
2006 Arthricocephalus jiangkouensis; Reference Yuan, Zhao and YangYuan et al., fig. 1a.
2011 Arthricocephalus cf. chauveaui; Reference Geyer and PeelGeyer and Peel, p. 498, fig. 14A–C, D?, E, F.
2015 Oryctocarella duyunensis; Reference Peng, Zhu, Babcock and KorovnikovPeng et al., p. 89, pl. 1, figs. 6–9.
Holotype
Incomplete exoskeleton (Qian, Reference Qian1961, pl. 2, fig. 8; Fig. 8.17, NIGP 11484) from the Balang Formation, Palang, Duyun, eastern Guizhou, China; by original designation.
Diagnosis
Oryctocarella with subcylindrical glabella; axial furrows with subparallel sides or gently diverging forward; palpebral ridge well defined, preocular field short or absent; palpebral lobe in anterior position, with posterior tip in adult specimens opposite midlength or anterior part of L2; palpebral area subequal in width to glabella. Thorax with 11 or possibly 12 segments; axis about half as broad as pleural region; anterior pleural band shorter than posterior band. Pygidial axis composed of three rings and a terminal piece; posterior margin with a medial notch. Surface covered with densely spaced granules.
Occurrence
From dark-gray calcareous mudstone and shale (weathering yellow-green) in the lower and middle parts of the Balang Formation, eastern Guizhou, and western Hunan, South China (Jiangnan Slope Belt); Cambrian Stage 4, Duyunian Stage of the regional stratigraphic scheme. It also occurs in the Olenellus Zone-equivalent of the Henson Gletscher Formation, North Greenland (Cambrian Stage 4).
Description
Exoskeleton elliptic in outline, about twice as long as wide. Cephalon subsemicircular in outline, with rounded genal corners; glabella subcylindrical, about four times as long as broad, widest part generally across L4; axial furrow more or less straight, parallel-sided or faintly divergent forward to near glabellar front; occipital furrow weak, deepened at lateral ends as incisions. Four pairs of glabellar furrows present; S1–S3 pit-like, each pair connected by a weak or faint transverse furrow that stops within pit, without extending outward to reach axial furrows; S4 narrow incision reaching to axial furrow abaxially; occipital ring wider, but much shorter than L1; L1–L3 subequal in length; L4 shorter than L3, and frontal lobe shorter than L4; L4 plus frontal lobe slightly longer than L3. Palpebral lobe medium-sized, ~0.26–0.31 (N=11) of cephalic length in adult specimens, gently curved, about 40º oblique to sagittal line, lying anterior to glabellar mid-length, posterior tip opposite S2; palpebral area about as wide as glabella or slightly narrower. Anterior branch of facial suture short, with straight posterior portion, moderately convergent and curved in anterior portion; posterior branch diverging outward and somewhat rearward, meeting lateral cephalic margin opposite L1. Librigena narrow, with wire-like, poorly defined lateral border; somewhat upturned posteriorly and progressively elevated anteriorly to merge with upturned anterior border of cranidium. Anterior cranidial border furrow well defined; posterior cephalic border transverse, broadening abaxially; posterior border furrow well defined.
Hypostome (Fig. 9.17, 9.18) subovate with inflated middle body and broad lateral and posterior borders, posterior margin gently curved, anterior margin (?hypostomal suture) horizontal.
Thorax non-fulcrate, with 11 or possibly 12 segments. Axis about half as broad as pleural region, gently narrowing rearward, slightly expanded medially, widest at segments 3–6; pleura with obtusely rounded tip, unequally divided into a shorter (exsag.) anterior band and a longer (exsag.) posterior band by well-incised pleural furrow, which runs slightly oblique to the longitudinal direction of the pleura and shows a weak rearward swing near abaxial termination.
Pygidium in holaspis (Fig. 8.14) subelliptical, transverse, about twice as broad as long. Axis small, containing a prominent articulating half-ring, three rings and a terminal piece, tapering rapidly rearward, occupying about half of pygidial length; ring furrows weakly to moderately defined. Pleural region gently convex, with 3–4 pleurae; pleural furrows and interpleural furrows well defined, gently curved; interpleural furrows slightly less well developed than pleural furrows. Posterior margin with a medial notch.
Dorsal surface covered with densely spaced granules.
Materials
Hundreds of sclerites from three sections (Mozichong, Bulin and Paiwu), Huayuan, northwestern Hunan, of which 22 exoskeletons (NIGP 164844–164851; NIGP 164858–164870; NWU-DYXJT 1823), three cranidia (NIGP 164852–164854) and three pygidia (NIGP164855–164857) are illustrated; reillustrated specimens include the holotype exoskeleton of the species (NIGP 11494) and an exoskeleton designated as holotype of Arthricocephalus horridus Qian and Lin in Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980 (NIGP 38236).
Remarks
Oryctocarella duyunensis was originally referred to Arthricocephalus (Qian, Reference Qian1961). Lane et al. (Reference Lane, Blaker and Zhang1988) regarded the species as a junior synonym of A. chauveaui, and selected a ‘lectotype’ for A. chauveaui (suppressed here) that in fact belongs to O. duyunensis. For three decades, the concepts of Arthricocephalus and A. chauveaui were thus based on specimens that are here referred to O. duyunensis. As discussed above and summarized in Figure 3, this species differs significantly from A. chauveaui. This species is similar to Oryctocarella sibirica (Tomashpolskaya in Khalfin, Reference Khalfin1960) (Fig. 6), the type species of Oryctocarella. In O. duyunensis, the palpebral lobe is located somewhat more anteriorly, and its posterior tip is opposite S2, whereas in O. sibirica the posterior tip of the palpebral lobe is opposite L2, and as a result, it appears to have more strongly diverging posterior branches of the facial suture. The neotype of O. sibirica (Fig. 6.1) and some other cranidia (Fig. 6.2, 6.3) differ from O. duyunensis in its glabellar shape (being subrectangular rather than cylindrical) and proportion (glabellar length ranging up to three times its width). There is considerable variability in these characters among the topotype material (Korovnikov and Novozhilova, Reference Korovnikov and Novozhilova2017, pl. 5, figs. 1, 3; Fig. 6.2, 6.3), but O. duyunensis differs further in having 11 or 12 thoracic segments in the holaspid stage and bearing a distinct posteromedial notch in the pygidial margin, whereas O. sibirica has only nine or possibly 10 thoracic segments, and evidently lacks a distinct posteromedial notch in the pygidial margin.
Genus Duyunaspis Zhang and Qian in Zhou et al., Reference Zhou, Liu, Meng and Sun1977
1977 Duyunaspis Zhang and Qian in Reference Zhou, Liu, Meng and SunZhou et al., p. 131.
1988 Arthricocephalus Bergeron; Reference Lane, Blaker and ZhangLane et al. (part), p. 555.
Type species
Duyunaspis duyunensis Zhang and Qian in Zhou et al., Reference Zhou, Liu, Meng and Sun1977, p. 132, pl. 41, figs. 5, 6, from the Balang Formation, Palang, Duyun, southeastern Guizhou; by original designation.
Other species
Duyunaspis paiwuensis Lei and Peng, Reference Lei and Peng2014 (p. 357, figs. 4, 5a), from the Balang Formation, Paiwu, Huayuan, northwestern Hunan.
Diagnosis
Micropygous genus of the Oryctocarinae with large, broadly subrectangular glabella; glabellar furrows present or absent, connected or separated from axial furrow; palpebral lobe long, occupying nearly half of glabellar length; palpebral field narrow, less than half of glabellar width. Thorax fulcrate, with nine segments; thoracic axis about as wide as pleural region; fulcrum located about one-third distance from axial furrow. Pygidium broad and short, axis with three rings plus a terminal piece; interpleural furrows poorly defined; lateral and posterior border poorly defined; posteromedial notch on margin commonly present. Dorsal surface smooth or covered with fine granules.
Remarks
McNamara et al. (Reference McNamara, Yu and Zhou2006) and Lei and Peng (Reference Lei and Peng2014) discussed Duyunaspis in detail. Two species assigned to this genus are considered properly recognizable: Duyunaspis duyunensis and D. paiwuensis Lei and Peng, Reference Lei and Peng2014 (p. 357, fig. 4). Lei and Peng (Reference Lei and Peng2014) provided an expanded concept of the genus, and that concept is followed here. Five species described from the Balang Formation in either northwestern Hunan or eastern Guizhou, South China, have been assigned to Duyunaspis, but, upon review by Lei and Peng (Reference Lei and Peng2014, p. 355), were determined to fall within the range of morphologic variation of D. duyunensis, and were therefore regarded as junior synonyms. We concur with that conclusion. The species are: (1) Duyunaspis guzhangensis Zhou (in Zhou et al., Reference Zhou, Liu, Meng and Sun1977, p. 132, pl. 41, fig. 7), based on a single exoskeleton collected from Guanba, Guzhang, northwestern Hunan; (2) Duyunaspis songtaoensis Qian and Lin (in Zhou et al., Reference Zhou, Liu, Meng and Sun1977, p. 132, pl. 41, fig. 8), based on a single exoskeleton from Xiunao, Songtao, eastern Guizhou; (3) Duyunaspis briaris Qian and Lin (in Yin and Li, Reference Yin and Li1978, p. 445, pl. 156, fig. 4), based on a single cranidium from Xiunao, Songtao, eastern Guizhou; (4), Duyunaspis obesis Qian and Lin (in Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, p. 274, pl. 90, fig. 6), which is an objective junior synonym of D. briaris because both species are based on the same holotype cranidium; and (5) Duyunaspis laevigatus Qian and Lin (in Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, p. 274, p. 91, figs. 7–9), known from three exoskeletons collected from Xiunao, Songtao, eastern Guizhou.
As discussed above, the smallest exoskeleton in Bergeron’s type series of Arthricocephalus chauveaui (Figs. 1.8, 2.12) is conspecific with D. duyunensis.
Duyunaspis was diagnosed by McNamara et al. (Reference McNamara, Yu and Zhou2006, p. 7) as having seven thoracic segments and five pygidial rings. New material illustrated both here and by Lei and Peng (Reference Lei and Peng2014), from the Balang Formation at Mozichong and Paiwu, Huayuan, northwestern Hunan, shows that holaspides have nine thoracic segments and three rings in the pygidial axis, indicating that all exoskeletons illustrated by McNamara et al. (Reference McNamara, Yu and Zhou2006) represent meraspid degree 7 individuals.
Duyunaspis is easily differentiated from both Arthricocephalus and Oryctocarella by having a proportionately broader glabella, a narrower fixigena, less well defined glabellar furrows, a longer palpebral lobe, and a broader thoracic axis. Each of the three genera can be distinguished by the number of thoracic segments in holaspides. Duyunaspis has nine thoracic segments, Arthricocephalus has eight thoracic segments and Oryctocarella has 11 or 12 thoracic segments. Both Duyunaspis and Arthricocephalus have fulcrate thoraxes, but the fulcra in Duyunaspis lie closer to the axial furrow than in Arthricocephalus. Oryctocarella differs from both Duyunaspis and Arthricocephalus in lacking fulcra. Both Duyunaspis and Oryctocarella are micropygous, whereas Arthricocephalus is isopygous. Species of Duyunaspis and Oryctocarella commonly show a posteromedial notch on the pygidial margin, but this characteristic is unknown in Arthricocephalus.
Duyunaspis duyunensis Zhang and Qian in Zhou et al., Reference Zhou, Liu, Meng and Sun1977

Figure 10 Duyunaspis duyunensis Zhang and Qian in Zhou et al., Reference Zhou, Liu, Meng and Sun1977 from the Balang Formation at Mozichong (1–10) and Paiwu (11–14), Huayuan, northwestern Hunan, China; all in dorsal view unless stated otherwise. White arrowhead indicates the posterior margin of thorax; arrowhead outlined in black indicates the posterior margin of partially released segment of transitory pygidium; black triangle indicates posteromedial notch in pygidial border; b=border, d=doublure; f=facet; 9=ninth thoracic segment: (1) NIGP 164871, latex cast from external mold of exoskeleton, meraspis degree 6, MZC-2; (2) NIGP 164872, exoskeleton, meraspis degree 5, MZC-2; (3) NIGP 164873, latex cast from external mold of exoskeleton, meraspis degree 7, MZC-1; (4) NIGP 164874, latex cast from external mold of exoskeleton, meraspis degree 8, MZC-1; (5) NIGP 164875, exoskeleton, meraspis degree 7, retrodeformation with inferred strain ellipse, MZC-1; (6) NIGP 164876, exoskeleton, meraspis degree 8, retrodeformation with inferred strain ellipse, MZC-2; (7) NIGP 164877, exoskeleton, meraspis degree 8, MZC-1; (8) NIGP 164878, exoskeleton, meraspis degree 8, retrodeformation with inferred strain ellipse, MZC-2; (9) NIGP 164879, exoskeleton, meraspis degree 8, MZC-1; (10) NIGP 164880, exoskeleton, meraspis degree 8, MZC-2; (11) NIGP 159524, exoskeleton, holaspid, retrodeformation with inferred strain ellipse, XHZA314-12; (12) NIGP 159525, incomplete exoskeleton lacking cranidium, holaspis, XHZB117; (13) enlargement of hypostome of specimen in 12, latex cast, ventral view; (14) enlargement of right side of thorax of specimen in 12, showing fulcra, facets and doublure at the margin of the thoracic segments. All scale bars represent 1.0 mm.
1977 Duyunaspis duyunensis Zhang and Qian in Reference Zhou, Liu, Meng and SunZhou et al., p. 132, pl. 41, figs. 5, 6.
1977 Duyunaspis guzhangensis Zhou in Reference Zhou, Liu, Meng and SunZhou et al., p. 132, pl. 41, fig. 7.
1977 Duyunaspis songtaoensis Qian and Lin in Reference Zhou, Liu, Meng and SunZhou et al., p. 132, pl. 41, fig. 8.
1978 Duyunaspis laevigatus Qian and Lin in Reference Yin and LiYin and Li, p. 444, pl. 156, fig. 3.
1978 Duyunaspis songtaoensis; Reference Yin and LiYin and Li, p. 444, pl. 156, fig. 5.
1978 Duyunaspis duyunensis; Reference Yin and LiYin and Li, p. 444, pl. 157, fig. 11.
1978 Duyunaspis braris Qian and Lin in Reference Yin and LiYin and Li, p. 445, pl. 156, fig. 4.
1980 Duyunaspis duyunensis; Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and YuanZhang et al., p. 273, pl. 91, figs. 5, 6.
1980 Duyunaspis obesis Qian and Lin in Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and YuanZhang et al., p. 274, pl. 90, fig. 6.
1980 Duyunaspis songtaoensis; Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and YuanZhang et al., p. 274, pl. 90, fig. 7; pl. 91, fig. 2.
1980 Duyunaspis laevigatus; Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and YuanZhang et al., p. 274, pl. 91, figs. 7–9.
1982 Duyunaspis songtaoensis; Liu, Reference Liu1982, p. 299, pl. 212, fig. 18.
1982 Duyunaspis guzhangensis; Reference LiuLiu, p. 299, pl. 212, fig. 14.
1982 Duyunaspis duyunensis; Reference LiuLiu, p. 299, pl. 213, fig. 10.
1988 Arthricocephalus chauveaui; Reference Lane, Blaker and ZhangLane et al. (part), p. 555, pl. 1, fig. 4; non pl. 1, figs. 1–3, 5, 6.
2001a Duyunaspis sp., Reference Zhao, Yu, Yuan, Yang and GuoZhao et al., p. 180, pl. 1, fig. 3.
2001 Duyunaspis duyunensis; Reference Yuan, Zhao and LiYuan et al., p. 224, pl. 1, fig. 6.
2006 Duyunaspis duyunensis; Reference McNamara, Yu and ZhouMcNamara et al., p. 7, pl. 2, figs. 5–18, text-fig. 5.
2014 Duyunaspis duyunensis; Lei and Peng, Reference Lei and Peng2014, p. 354, figs. 3, 5b.
2015 Duyunaspis duyunensis; Reference Peng, Zhu, Babcock and KorovnikovPeng et al., p. 89, pl. 1, fig. 10.
Holotype
Incomplete exoskeleton (Zhou et al., Reference Zhou, Liu, Meng and Sun1977, p. 132, pl. 41, fig. 5), NIGP 38228, from the Balang Formation, Palang, Duyun, southeastern Guizhou, China; by original designation.
Diagnosis
As for the genus.
Occurrence
From dark-gray calcareous mudstone and shale (weathering yellow-green) in the middle part of the Balang Formation, eastern and southeastern Guizhou, western and northwestern Hunan, South China (Jiangnan Slope Belt); Cambrian Stage 4, Duyunian Stage of the regional stratigraphic scheme.
Description
The description in McNamara et al. (Reference McNamara, Yu and Zhou2006, p. 11–12), which applies to specimens from the protaspid period through meraspid degree 7, is slightly emended here. Apart from additional thoracic segments, the specimens of meraspis degree 8 (with eight thoracic segments) and holaspides (with nine thoracic segments) both resemble the meraspid degree 7 exoskeleton.
Hypostome shield-shape; middle body inflated, divided into a long anterior lobe, and a crescentic posterior lobe by a faint medial furrow; macula deeply incised, located near mid-length of the middle body; lateral and posterior border narrow, strongly upturned; anterior wing triangular, with subequal length and width, anterior margin (hypostomal suture?) gently arched forward.
Pygidial border gently upturned, defined by a shallow and broad border furrow. Posterior margin with distinct, broad notch.
Materials
10 exoskeletons (NIGP 164871–164880).
Remarks
Based on a large collection of 766 exoskeletons from the Balang Formation at Paiwu, Huayuan, northwestern Hunan, Lei and Peng (Reference Lei and Peng2014, p. 355–356) synonymized five earlier described species of Duyunaspis with D. duyunensis. The characters previously used to distinguish the species were regarded by Lei and Peng (Reference Lei and Peng2014) as the result of either intraspecific variation or taphonomic alteration. The synonymized species are Duyunaspis guzhangensis Zhou (in Zhou et al., Reference Zhou, Liu, Meng and Sun1977, p. 132, pl. 41, fig. 7); Duyunaspis songtaoensis Qian and Lin (in Zhou et al., Reference Zhou, Liu, Meng and Sun1977, p. 132, pl. 41, fig. 8); Duyunaspis briaris Qian and Lin (in Yin and Li, Reference Yin and Li1978, p. 445, pl. 156, fig. 4); Duyunaspis obesis Qian and Lin (in Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, p. 274, pl. 90, fig. 6); and Duyunaspis laevigatus Qian and Lin (in Zhang et al., Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980, p. 274, pl. 91, figs. 7–9). Duyunaspis briaris and D. obesis are objective synonyms because both are based on the same holotype cranidium. The types of all five synonymized species are preserved in greenish-yellow shale or mudstone. Some variation among them is probably the result of intraspecific variation (Lei and Peng, Reference Lei and Peng2014, p. 359, table 1). However, the fossils are mostly flattened and distorted, leading to variation in proportions of the exoskeleton and variation in the expression of axial and glabellar furrows. The glabellar furrows have been either enhanced or nearly obliterated through taphonomic distortion. Glabellar furrow S1, which consists of isolated pits separated from the axial furrows in well-preserved cranidia, may attain a transglabellar extent after taphonomic compaction.
The thorax of Duyunaspis duyunensis has been reported to consist of either eight segments (Zhang and Qian in Zhou et al., Reference Zhou, Liu, Meng and Sun1977, p. 182) or seven segments (McNamara et al., Reference McNamara, Yu and Zhou2006, p. 10) in the holaspid period. Lei and Peng (Reference Lei and Peng2014, p. 355), however, showed that the holaspid exoskeleton has as many as nine thoracic segments (Lei and Peng, Reference Lei and Peng2014, fig. 3.10–3.16; Fig. 10.11, 10.12).
Duyunaspis paiwuensis (Lei and Peng, Reference Lei and Peng2014, fig. 4), from the lower part of the Balang Formation at Paiwu, northwestern Hunan, resembles D. duyunensis in the overall exoskeleton morphology. Duyunaspis paiwuensis differs from the latter species mainly in having a proportionally broader and shorter cephalon, a narrower and longer glabella, a narrower thoracic axis (narrower than the pleural region), and a fulcrum that is located further abaxially from the axial furrow.
Acknowledgements
We thank J. Vannier (Lyon) for providing access to facilities during SP’s visit, and Bertrand Lefebvre (Lyon) for preparing the latex cast of the Bergeron’s slab. We thank P.D. Lane (Keele) for recalling information about the preparation of his co-authored 1988 paper; Y. Li (Nanjing) for offering collection LYX-1; I.V. Korovnikov (Novosibirsk) for providing images of topotypic specimens of Oryctocarella sibirica; S. Clausen (Lille) for improving the English translation of Bergeron’s (Reference Bergeron1899) paper; and T.V. Pegel (Novosibirsk) for access to Russian literature. We are also greatly indebted to J. Jago (Adlaide) and G. Geyer (Würzburg) for their helpful comments and suggestions for improving the manuscript, and to B. Hunda (Cincinnati), and N. Hughes (Riverside) for their accurate editorial work. Funding was provided by the National Natural Science Foundation of China to SP (41330101, 41521061, 41290260) and to XZ (41672002), by the Ministry of Science and Technology of China (2015FY31010), and by Regional Geological Survey of Guizhou Province (12120114068201); a grant was provided by the Subsurface Energy Research Center, The Ohio State University to LEB.










