


Introduction
During the latest Cretaceous (late Maastrichtian), the shallow, subtropical sea that covered the present-day Maastricht area (southern Limburg) was inhabited by a range of vertebrates. The top of the food chain was occupied by mosasauroids, the largest marine reptiles of this time interval, with overall lengths in excess of 16 m (Lingham-Soliar, Reference Lingham-Soliar1995; Gallagher et al., Reference Gallagher, Jagt, Mulder and Schulp2004; Street & Caldwell, Reference Street and Caldwell2016). Other associated reptiles included plesiosaurs (Mulder et al., Reference Mulder, Bardet, Godefroit and Jagt2000), crocodiles (Mulder et al., Reference Mulder, Jagt, Kuypers, Peeters and Rompen1998, Reference Mulder, Jagt, Stroucken, Sullivan and Lucas2016) and turtles (Mulder, Reference Mulder2003; Janssen et al., Reference Janssen, Van Baal, Schulp, Jagt, Jagt-Yazykova and Schins2011). In addition, a fairly wide array of small to medium-sized cartilaginous fish such as sharks, rays, skates and sawfish (Elasmobranchii) and chimaeras (Holocephali) was represented (Herman, Reference Herman1977; Duffin & Reynders, Reference Duffin and Reynders1995), as well as various species of ray-finned fish (Actinopterygii) (Friedman, Reference Friedman, Jagt, Donovan and Jagt-Yazykova2012), in particular in the upper Gulpen Formation and the entire Maastricht and Kunrade formations.
Although teleosts are not rare in the type Maastrichtian (Fig. 1A), their record is confined mostly to isolated teeth, skull bones (occasionally associated, yet displaced), vertebrae and scales. Especially in the lower Maastricht Formation, the ichnofossil (burrow) Lepidenteron lewesiensis (Mantell, Reference Mantell1822) is quite common (see Jurkowska & Uchman, Reference Jurkowska and Uchman2013; Bieńkowska-Wasiluk et al., Reference Bieńkowska-Wasiluk, Uchman, Jurkowska and Świerczewska-Gładysz2015; Niebuhr & Wilmsen, Reference Niebuhr and Wilmsen2016; Jagt, Reference Jagt2019), containing numerous teleost remains, including those of dercetids. More or less complete fish or associated remains of skull, fins and body outside burrows, as is the case here, are very rare (Friedman, Reference Friedman, Jagt, Donovan and Jagt-Yazykova2012; Taverne & Goolaerts, Reference Taverne and Goolaerts2015).
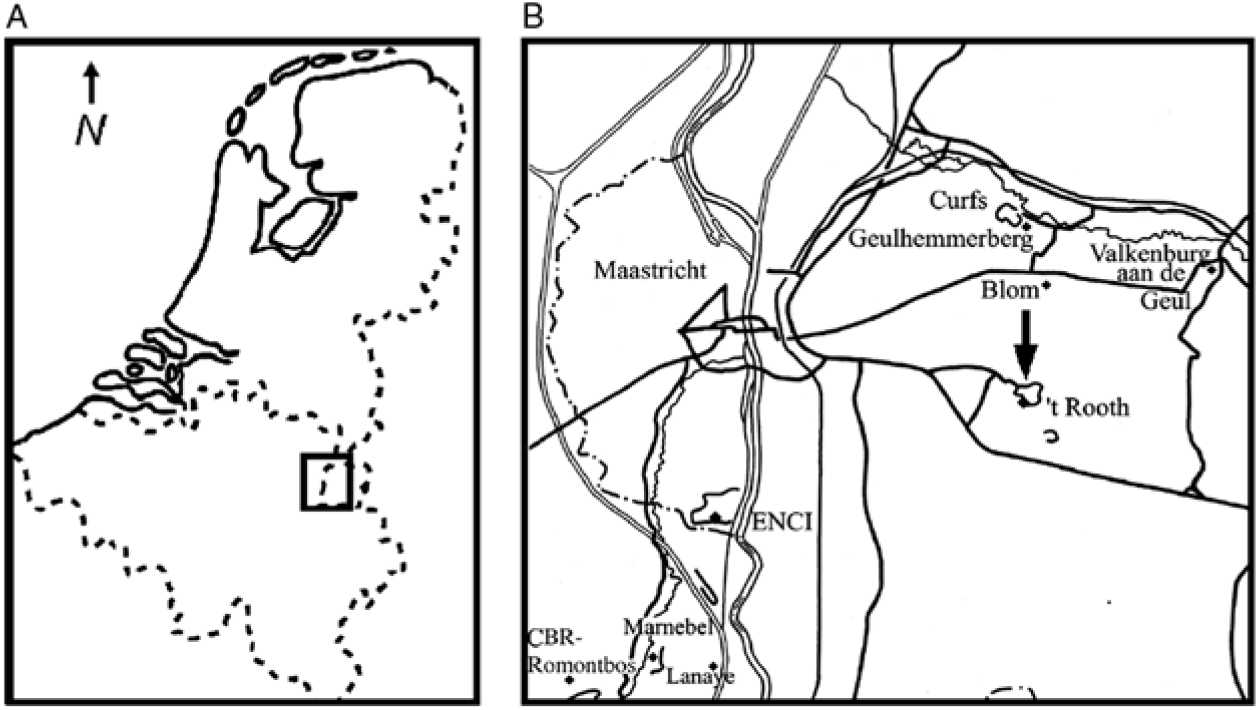
Fig. 1. (A) Map of the Netherlands and Belgium, with indication of the type area of the Maastrichtian Stage (inset). (B) Part of southern Limburg (the Netherlands) and contiguous areas in Belgium (provinces of Limburg and Liège), with indication (arrow) of ’t Rooth quarry (formerly Nekami), near Bemelen (southern Limburg), at which the new dercetid was collected (modified after Jagt & Jagt-Yazykova, Reference Jagt, Jagt-Yazykova, Jagt, Donovan and Jagt-Yazykova2012; maps.google.com).
Dercetids are highly specialised Late Cretaceous marine teleosts (Taverne & Goolaerts, Reference Taverne and Goolaerts2015), occasionally referred to as ‘needle fish’, that are characterised by a long pointed rostrum (i.e. an extension of the upper and lower jaws), one or two pairs of transverse processes on each abdominal vertebral centrum, and an eel-shaped body covered with cordiform or tripartite scutes (Silva & Gallo, Reference Silva and Gallo2011). Sixteen genera of dercetid, comprising 26 species, are known to date, the oldest one being of Cenomanian, the youngest of Early Palaeocene (Danian) age, with records from Europe, North, Central and South America, Africa, the Middle East and Far East of Asia (Siegfried, Reference Siegfried1966; Longbottom & Patterson, Reference Longbottom, Patterson, Owen and Smith1987; Taverne, Reference Taverne1987, Reference Taverne2005a, Reference Taverne2005b, Reference Taverne2006a, Reference Taverne2006b, Reference Taverne2008, Reference Taverne2013; Blanco & Alvarado-Ortega, Reference Blanco and Alvarado-Ortega2006; De Figueiredo & Gallo, Reference De Figueiredo and Gallo2006; Ekrt et al., Reference Ekrt, Košťák, Mazuch, Voigt and Wiese2008; Taverne & Goolaerts, Reference Taverne and Goolaerts2015; Licht et al., Reference Licht, Kogan, Fischer and Reiss2016; Vernygora et al., Reference Vernygora, Murray, Luque, Ruge and Fonseca2017).
Until publications by Lambers (Reference Lambers, Jagt, Leloux and Dhondt1998) and Friedman (Reference Friedman, Jagt, Donovan and Jagt-Yazykova2012), dercetids were unknown from upper Maastrichtian strata of southern Limburg and Liège (northeast Belgium), but there was a single record (under the name of Leptotrachelus sp., later placed in the genus Dercetis by Siegfried (Reference Siegfried1966)) from the lower Campanian Vaals Formation at Aachen–Linzenhäuschen (Germany) by Albers & Weiler (Reference Albers and Weiler1964, p. 26, figs 44, 45a, b, 46a, b). More recently, Taverne & Goolaerts (Reference Taverne and Goolaerts2015) have recorded four genera and four species of dercetids, two of them new, on the basis of incomplete, albeit associated remains (Fig. 2).
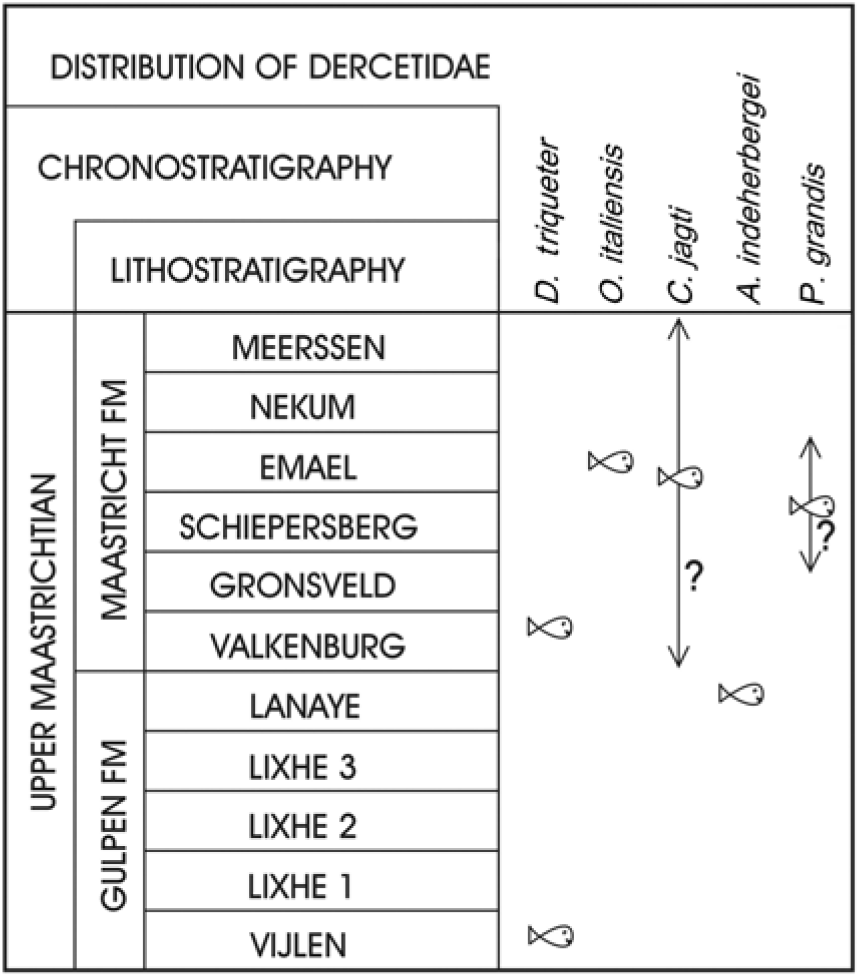
Fig. 2. Stratigraphical provenance of all latest Cretaceous dercetid taxa (Dercetis triqueter, Ophidercetis italiensis, Apuliadercetis indeherbergei, Cyranichthys jagti and Pelargorhynchus grandis n. sp.) known to date from the Maastrichtian type area (modified after Taverne & Goolaerts, Reference Taverne and Goolaerts2015).
Material and methods
Retained in the collections of the Oertijdmuseum (Boxtel, the Netherlands) is a slab from ’t Rooth quarry at Bemelen (Fig. 1B), the bedding plane of which is replete with large, semi-articulated scales and isolated skull bones. Additional preparation and consolidation (Osteofix) has recently been carried out by the senior author, who also studied it in detail and produced camera-lucida drawings.
Used abbreviations. BRSTG, branchiostegal rays; ECPT, ectopterygoid; FR, frontal; IOP, interoperculum; MC, median crest; MX, maxilla; OP, operculum; PMX, premaxilla; PMX t., premaxilla tip; PS, parasphenoid; Q, quadrate; RI, ridge; Sca, scale; Scu, scute; SOP, suboperculum; SP, spine; UJ, upper jaw.
Systematic palaeontology
Division Teleostei Müller, Reference Müller1846
Subdivision Neoteleostei Nelson, Reference Nelson1969
Order Aulopiformes Rosen, Reference Rosen, Greenwood, Miles and Patterson1973
Family Dercetidae Woodward, Reference Woodward1901
Genus Pelargorhynchus von der Marck, Reference von der Marck1858
Type species. Pelargorhynchus dercetiformis von der Marck, Reference von der Marck1858
Pelargorhynchus grandis n. sp.

Fig. 3. Photograph of the holotype (MAB 3688) of Pelargorhynchus grandis n. sp., with interpretation (labelling) of isolated skeletal elements.
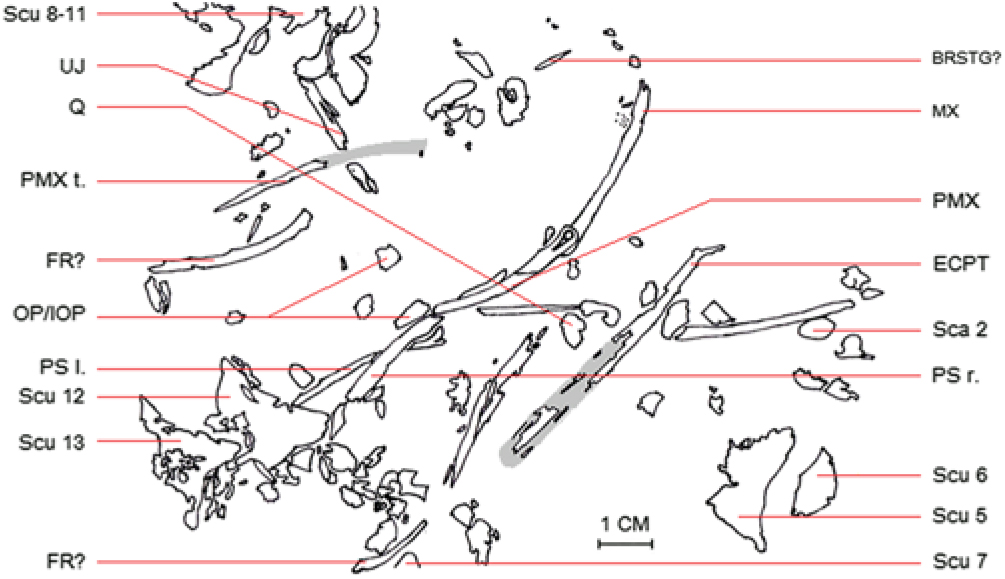
Fig. 4. Line drawing of the different skull fragments (cf. Fig. 3) of the holotype (MAB 3688) of Pelargorhynchus grandis n. sp.
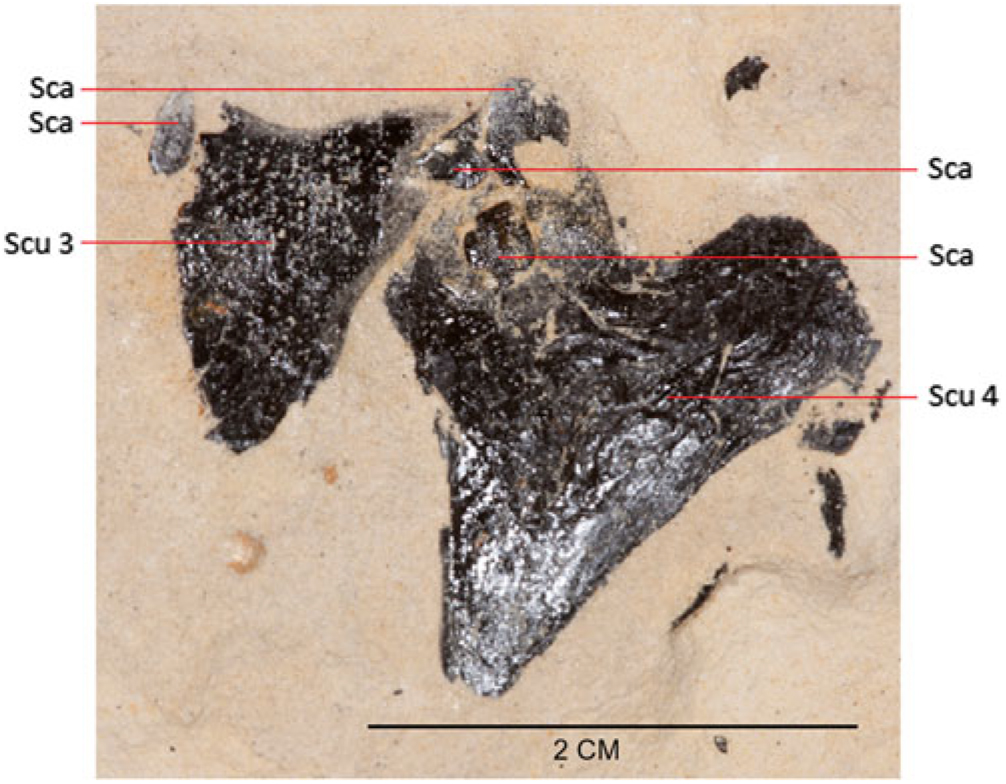
Fig. 5. Photograph of scutes 3 (partially broken) and 4 and several scales in the holotype, MAB 3688.
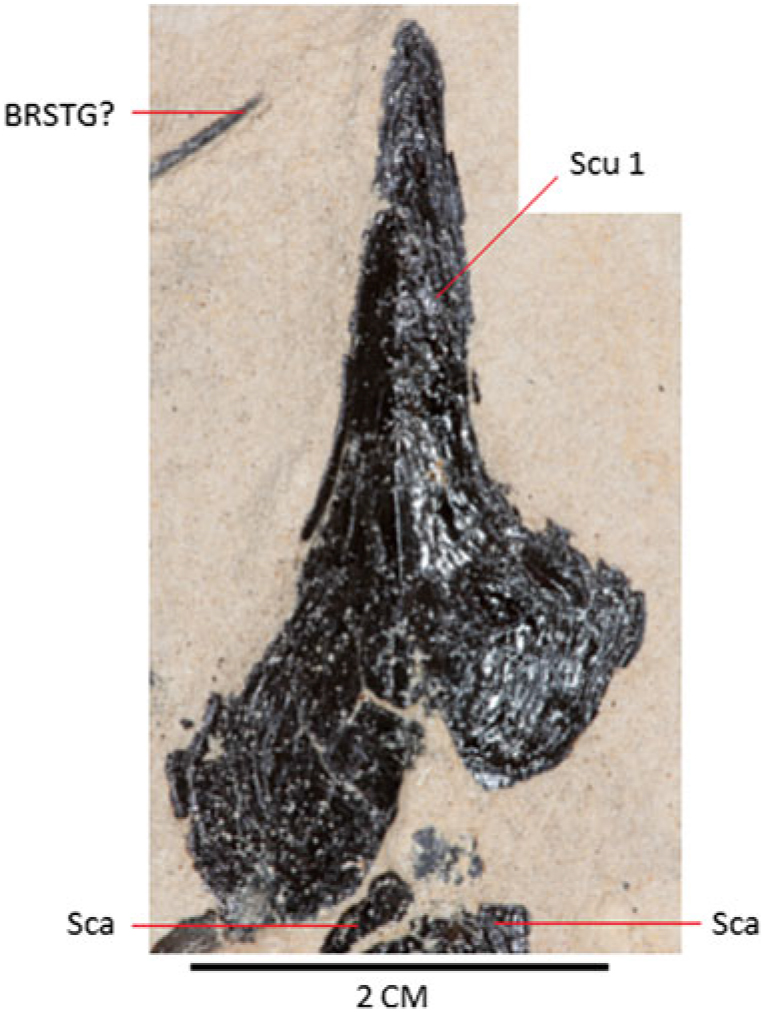
Fig. 6. Photograph of scute 1, several scales and a possible branchiostegal ray in the holotype, MAB 3688.

Fig. 7. Photograph of scutes 8–11; visible are a part of the upper jaw, as well as different scutes and scales (MAB 3688).
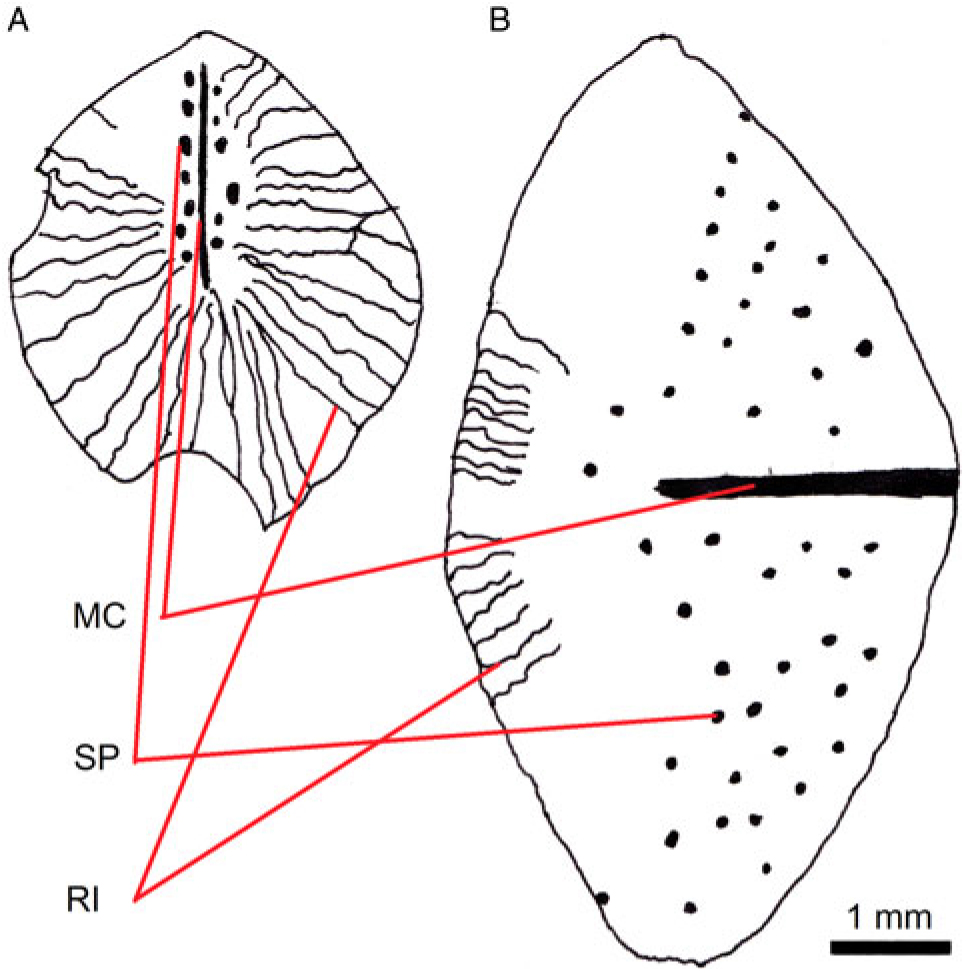
Fig. 8. Line drawing of scales 1 (A) and 2 (B) in the holotype (MAB 3688); recognisable are spines (SP), ridges (RI) and median crest (MC).
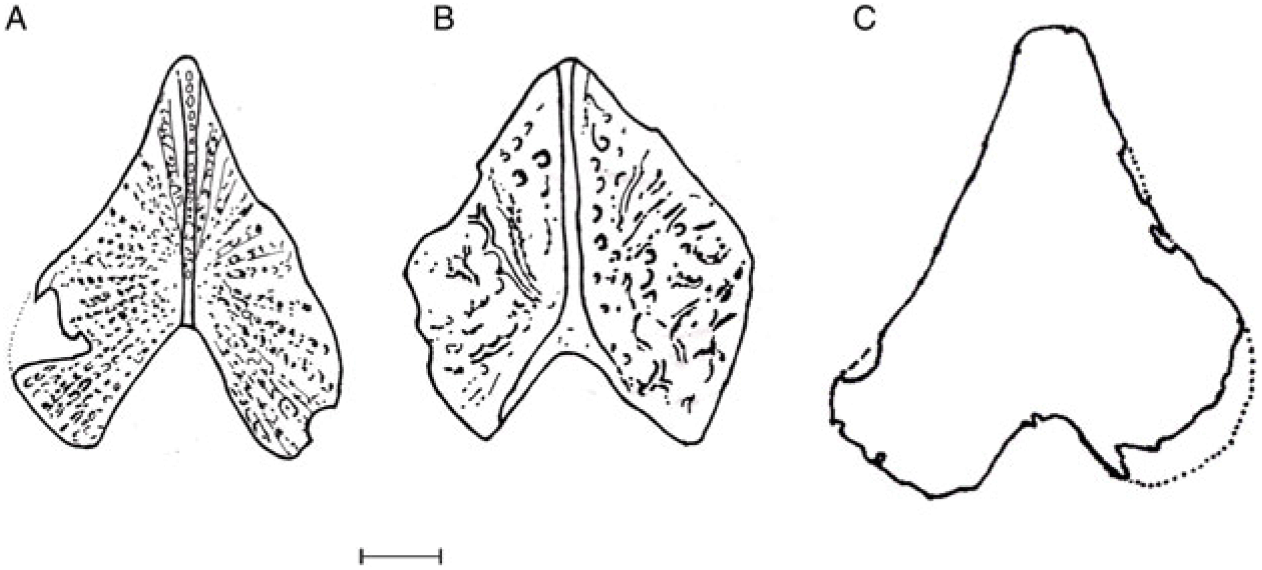
Fig. 9. Line drawings of scutes of Cyranichthys jagti Taverne & Goolaerts, Reference Taverne and Goolaerts2015 (A), Cyranichthys ornatissimus (Casier, Reference Casier1965) (B) and Pelargorhynchus grandis n. sp. (C). Scale bar equals 2 mm (A and B; adapted from Taverne & Goolaerts, Reference Taverne and Goolaerts2015) and 5 mm (C).
Diagnosis
Body squamation consisting of one or more lateral rows of smooth, cordiform scutes, combined with small, ornamented scales; scale ornamentation comprising several small spines and ridges originating from median crest; skull bones with high degree of curvature; premaxillae not fused, maxilla and premaxilla both lacking teeth.
Type material
The holotype is MAB 3688, preserved on the bedding plane of a slab (c. 210 × 290 mm) of fine-grained, homogeneous biocalcarenite, with a few whitish-grey burrow flints (Fig. 3).
Etymology
In allusion to the comparatively large size of the specimen.
Type locality and stratum
’t Rooth quarry (formerly Nekami; outcrop 62A-7, coordinates 182.750/316.300), near Bemelen, southern Limburg (see Felder & Bosch, Reference Felder and Bosch2000, pp. 106–107, fig. 4.4). On an associated label the collector, the late Robert Frijns, mentioned merely that the slab was collected from below the Laumont Horizon, which is the base of the Nekum Member (Maastricht Formation). On account of the fine-grained, even-coloured nature of the matrix, with few whitish-grey burrow flints and the presence of a single, incomplete specimen of the small pectinoid bivalve Syncyclonema haeggi Dhondt, Reference Dhondt1971, the slab must have been recovered from the lower or middle portion of the Maastricht Formation (i.e. upper Gronsveld, Schiepersberg or Emael members).
Description
All elements (scutes, scales and skull bones) stem from the anterior part of the body; no vertebrae preserved. Skull elements dissociated and dispersed (Fig. 4); maxilla and premaxilla overlapping, as preserved; lengths of incomplete maxilla and premaxilla 40 and 32 mm, respectively. Premaxilla ending with a triangular-shaped bulge; no teeth, nor any traces of dentition on premaxilla and maxilla. Tip of premaxilla flattened (2 mm in width); fragment measuring 26 mm in overall length; other side broken but imprint of this part 40 mm in length, indicating an even greater original length. Premaxilla not fused; no trace of a sutured premaxilla. Two curved parts of possible frontals (left and right); first one measuring 38 mm in length, second one 22 mm. In both fragments one side is flattened, the other side is broken off. Right and left parasphenoids preserved and overlapping, their lengths being 53 and 38 mm, respectively. Both parasphenoids have one side broken. They have a surface of up to 3 mm in width, and one side ending in a small bulge.
Ectopterygoid with one side broken up into pieces, which precludes obtaining exact measurements; estimated length 65 mm and estimated width at least 4 mm. One side ends with a bulge. Quadrate fragment measures 6 by 4 mm. Another part of the possible upper jaw is preserved, although further identification is impossible because of the damage on this fragment. Also preserved are two possible branchiostegal rays and parts of the possible operculum and interoperculum. Several smaller and more damaged fragments are found in the specimen, but they are severely damaged and thus impossible to identify. The maxilla, premaxilla, the parasphenoids and the frontals are slightly curved. The curvature as preserved is irregular, which rules out post-mortem deformation. Skull shape and jaw structure (curved upwards, downwards or laterally) cannot be determined because too many parts are missing.
Scutes are typically cordiform, although with small variations in shape which reflect former position on the flank of the body. The symmetrical scutes are cordiform (see Fig. 5; Scu 4, measuring 30 by 23 mm), with smooth outer surface and wider than an asymmetrical scute (see Fig. 6; Scu 1, measuring 22 by 40 mm) that appears to be lying upside down. Asymmetrical scutes have a smooth surface, except for a median crest. Scutes 8–11 are preserved as an overlapping set (Fig. 7). Although the shape of these scutes appears different from the other types mentioned above, this is not the case, the apparent difference being most likely caused by poor preservation (fracturing and exfoliation).
Unlike the scutes, all scales have a cover of spines and small ridges which originate from the median crest and end at scale margin. Apparently, scale 2 (Fig. 8B) differs markedly from scale 1 (Fig. 8A); six of the preserved scales are of the same type as scale 1, three of the scales are from the same type as scale 2. Meagre as these remains may be, the body of this new dercetid was most likely covered by one or more lateral rows of scutes, with other parts having a cover of smaller scales.
Discussion
From the extended type area of the Maastrichtian Stage, four dercetid taxa have been recorded to date: Dercetis triqueter Pictet, Reference Pictet1850, Ophidercetis italiensis Taverne, Reference Taverne2005a, Apuliadercetis indeherbergei Taverne & Goolaerts, Reference Taverne and Goolaerts2015 and Cyranichthys jagti Taverne & Goolaerts, Reference Taverne and Goolaerts2015. All of these occur in upper Maastrichtian strata, and are apparently restricted to the Vijlen and Lanaye members (Gulpen Formation) and the overlying Valkenburg, Gronsveld, Schiepersberg and Emael members (Maastricht Formation; Fig. 2 and J.W.M.J., pers. obs.).
The new species is easily differentiated from other dercetids in having conspicuously large and smooth scutes, in combination with small ornamented scales, and in having curved skull elements. In addition, it lacks teeth.
To assess the generic assignment of the present specimen, we have scored the following five features: (1) smooth, cordiform scutes; (2) presence of smaller, ornamented scales; (3) absence of teeth; (4) curvature of skull fragments; and (5) unfused premaxillae.
Cordiform scutes occur in just four dercetid genera, viz. Cyranichthys Taverne, Reference Taverne1987, Ophidercetis Taverne, Reference Taverne2005a, Pelargorhynchus von der Marck, Reference von der Marck1863 and Brazilodercetis De Figueiredo & Gallo, Reference De Figueiredo and Gallo2006 (see De Figueiredo & Gallo, Reference De Figueiredo and Gallo2006; Taverne & Goolaerts, Reference Taverne and Goolaerts2015), which in total comprise five species: O. italiensis Taverne, Reference Taverne2005a, P. dercetiformis (von der Marck, Reference von der Marck1863), C. ornatissimus (Casier, Reference Casier1965) (see Taverne, Reference Taverne1987), C. jagti Taverne & Goolaerts, Reference Taverne and Goolaerts2015 and Brazilodercetis longirostris (De Figueiredo & Gallo, Reference De Figueiredo and Gallo2006). The last-named species does have cordiform scutes, but because of the presence of different tripartite scutes on the remainder of the body, this taxon can be ruled out for a direct comparison. The other species are shown in Table 1.
Table 1. Display of the four dercetid species that are most closely comparable with P. grandis n. sp., as described here
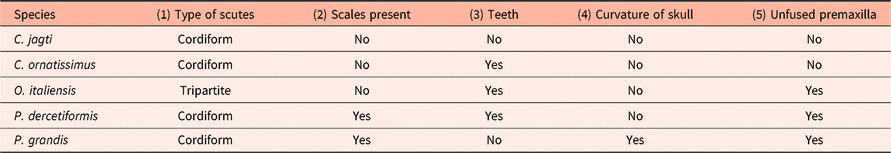
Scute shape in Pelargorhynchus grandis n. sp. is most closely comparable to that of the genera Cyranichthys and Pelargorhynchus, while scutes in Ophidercetis have a more triangular outline. A range of scutes of species of the genus Cyranichthys are illustrated in Figure 9, which demonstrates the similarity between C. ornatissimus, C. jagti and the new species. Feature 2 (see above) is seen in P. dercetiformis, which has a cover of scales and five rows of scutes. The bodies of C. ornatissimus, C. jagti and O. italiensis are covered with a few rows of scutes, but no scales have been described for those species.
Teeth (feature 3) have been recorded in C. ornatissimus, O. italiensis and P. dercetiformis, while C. jagti and P. grandis n. sp. appear to have been edentulate.
Curved skull fragments (feature 4) appear to be unique to the present individual. The other four species do have some curved elements in their skulls, but the curvature is much more modest (von der Marck, Reference von der Marck1858, Reference von der Marck1863; Taverne, Reference Taverne1987, Reference Taverne2005a; Taverne & Goolaerts, Reference Taverne and Goolaerts2015). The premaxillae (feature 5) are firmly fused (sutured) in the genus Cyranichthys, but are unfused in O. italiensis, P. dercetiformis and the new form.
On the basis of features 1 and 3, it would seem that Pelargorhynchus grandis n. sp. fits best within the genus Cyranichthys. Although the presence of teeth (feature 3) is also found in Cyranichthys, features 1, 2 and 5 are comparable with Pelargorhynchus dercetiformis, and based on these features the new species is placed within the genus Pelargorhynchus.
Although features 3 and 4 show a difference between previously described species of the genus Pelargorhynchus and P. grandis n. sp., available data are insufficient to erect a new genus for it. When more complete specimens of P. grandis n. sp. are found, it can be compared in greater detail with other dercetids and possibly be placed in a new genus.
The holotype of Cyranichthys jagti (NHMM 1993 119; see Taverne & Goolaerts, Reference Taverne and Goolaerts2015, figs 11–13) also lacks teeth, but a comparison with its partially preserved and articulated skull with skull elements preserved in P. grandis n. sp. is inconclusive as far as determination of the relationship between these two species is concerned.
Conclusions
Pelargorhynchus grandis n. sp. is the fifth and by far the largest representative of the family Dercetidae that has been recorded from the type Maastrichtian strata to date. A comparison with other dercetid genera shows that the new form is best accommodated, at least for the time being, in Pelargorhynchus when scute shape, the presence of scales, and the unfused premaxilla are considered. The absence of teeth and the marked curvature of skull bones would suggest erection of a new genus to accommodate this form, but the incomplete preservation of the type specimen prevents us from doing so. The new species is characterised by large, smooth and cordiform scutes, small ornamented scales, unfused premaxillae and markedly curved skull bones; in addition, teeth are absent.
Acknowledgements
We wish to thank Dr Louis Taverne (Brussels) and Dr Paul H. Lambers (Utrecht) for supplying items of literature; the senior author is grateful to Dr Wilma Wessels (Utrecht) for supervision during this study and a special word of thanks is extended to Stan van Dongen (Waalwijk) for preparation of photographs.











 Open access
Open access