




Introduction
Opinions differ between those who consider archaic hominin cognition to be comparable to that of anatomically modern humans (e.g. Hovers & Belfer-Cohen Reference Hovers, Belfer-Cohen, Hovers and Kuhn2006; Nowell Reference Nowell2010; d’Errico & Stringer Reference d'Errico and Stringer2011; Zilhão Reference Zilhão, Condemi and Weniger2011; Villa & Roebroeks Reference Villa and Roebroeks2014), and those who consider morphological, physiological, ontogenetic and behavioural differences to suggest different cognitive frameworks (e.g. Spoor et al. Reference Spoor, Hublin, Braun and Zonneveld2003; Lieberman Reference Lieberman2007; Gunz et al. Reference Gunz, Neubauer, Maureille and Hublin2010; Junker Reference Junker2010; Mellars Reference Mellars2010; Benazzi et al. Reference Benazzi, Douka, Fornai, Bauer, Kullmer, Svoboda, Pap, Mallegni, Bayle, Coquerelle, Condemi, Ronchitelli, Harvati and Weber2011; de Boer Reference de Boer2012; Pearce et al. Reference Pearce, Stringer and Dunbar2013; Bruner Reference Bruner, Akazawa, Ogihara, Tanabe and Terashima2014; Neubauer Reference Neubauer and Bruner2015). The first viewpoint is supported by the fact that the emergence of our species in Africa, c. 300 kya (Grün et al. Reference Grün, Brink, Spooner, Taylor, Stringer, Franciscus and Murray1996; Stringer Reference Stringer2016; Hublin et al. Reference Hublin, Ben-Ncer, Bailey, Freidline, Neubauer, Skinner, Bergmann, Cabec, Benazzi, Harvati and Gunz2017), was not accompanied by the immediate development of behaviours characteristic of historically documented populations. For tens of thousands of years after their emergence, anatomically modern human populations in Africa continued to use technologies that differed little from those of the non-modern populations preceding them, or from those contemporaneous populations inhabiting other regions, both inside and outside the African continent. It is also supported by the discovery of cultural innovations generally considered to be hallmarks of modern cognition among archaic populations of Eurasia. Burials, the collection of fossils and other rare objects, the production of personal ornaments and engraved patterns, pigment use, cave painting and the extraction of bird feathers and claws are all interpreted as proof that Neanderthals in Europe and the Near East engaged in symbolically mediated behaviour (e.g. Pettitt Reference Pettitt2002; Zilhão et al. Reference Zilhão2010; Peresani et al. Reference Peresani, Fiore, Gala, Romandini and Tagliacozzo2011; Finlayson et al. Reference Finlayson, Brown, Blasco, Rosell, Negro, Bortolotti, Finlayson, Sánchez Marco, Giles Pacheco, Rodríguez Vidal, Carrión, Fa and Rodríguez Llanes2012; Rodríguez-Vidal et al. Reference Rodríguez-Vidal2014; Romandini et al. Reference Romandini, Peresani, Laroulandie, Metz, Pastoors, Vaquero and Slimak2014; Radovčić et al. Reference Radovčić, Sršen, Radovčić and Frayer2015; Hoffmann et al. Reference Hoffmann, Standish, García-Diez, Pettitt, Milton, Zilhão, Alcolea-González, Cantalejo-Duarte, Collado, de Balbín, Lorblanchet, Ramos-Muñoz, Weniger and Pike2018). Most of these innovative cultural adaptations, however, are virtually unknown in vast regions inhabited by archaic hominins prior to the arrival of modern human populations—particularly in East Asia. A putative engraving on a bone flake from the Shiyu site, Shanxi Province, dated to 32–28 kya (You Reference You1984), has been interpreted as resulting from post-depositional surface modifications (Bednarick Reference Bednarik1994).
Abstract engravings are reported from almost 40 African and Eurasian sites dated prior to 40 kya (see online supplementary material (OSM) 1). The earliest examples are the engraved patterns on a freshwater shell from Trinil, Indonesia, dating to 540 kya; a bone from Bilzingsleben in Germany (370 kya; see, however, Müller & Pasda Reference Müller and Pasda2011), ochre fragments from the South African sites of Klasies River, Pinnacle Point and Blombos Cave (110–73 kya); and an antler from Vaufrey in France, dating to 120 kya (Vincent Reference Vincent, Otte, Binford and Rigaud1988; Henshilwood et al. Reference Henshilwood, d'Errico and Watts2009; Watts Reference Watts2010; d’Errico et al. Reference d'Errico, Moreno and Rifkin2012). In China, engraved objects from Pleistocene contexts are rare. The earliest possible example is a Stegodon ivory tusk found in a layer dated to 150–120 kya at Xinglongdong Cave, south China. The piece displays a few longitudinal incisions close to the tip of the tusk. Both the site's chronology and the anthropogenic nature of the incisions have been questioned, as elephants use their tusks for a variety of activities that could result in their breakage or the development of ground facets and incisions (Haynes Reference Haynes1991; Villa & d’Errico Reference Villa and d'Errico2001; Norton & Jin Reference Norton and Jin2009). Elsewhere, a pebble from a context dated to c. 40 kya from the lower cultural unit of Shuidonggou locality 1 exhibits a set of sub-parallel and intersecting lines (Peng et al. Reference Peng, Gao, Wang, Chen, Liu and Pei2012). Pei (Reference Pei1934) describes an engraved antler from Zhoukoudian Upper Cave, now dated to 34–29 kya (Chen et al. Reference Chen, Hedges and Yuan1992). A putative engraved bone flake recovered from the Shiyu site, Shanxi Province, dated to 32–28 kya (You Reference You1984) has been questioned on the basis of post-depositional surface modifications (Bednarik Reference Bednarik1994). Finally, an antler fragment with an engraved pattern comprising sinuous parallel lines is reported from Longgu Cave, Hebei Province, dated to 13 kya (Bednarik & You Reference Bednarik and You1991).
Here, we report the discovery of weathered bone fragments exhibiting deliberately engraved lines, one of which is filled with ochre, found at the 125–105 kya site of Lingjing in China. The fragments were found in the same stratigraphic layer that yielded hominin remains attributed to an archaic population exhibiting a mosaic of anatomical features suggestive of complex population dynamics between Eastern and Western Eurasia (Li et al. Reference Li, Wu, Zhou, Liu, Gao, Nian and Trinkaus2017). Recently, it has been suggested that the Lingjing hominins were Denisovans (Martinón-Torres et al. Reference Martinón-Torres, Wu, Bermúdez de Castro, Xing and Liu2017: 444), although there is currently insufficient palaeogenetic evidence to corroborate this hypothesis.
Archaeological context
Lingjing (34° 04’ 08.6” north, 113° 40’ 47.5” east; 117m asl) is an open-air site located in Henan Province, northern China, approximately 120km south of the Yellow River (Figure 1). The site was discovered in 1965. Since 2005, excavations under the supervision of the first author have uncovered 551m2 of the site. The 9m-deep, water-lain sedimentary sequence comprises 11 geological layers (for details, see OSM 2). Three main archaeological horizons attest to human occupation of the site during the Holocene (layers 1–4), the Late Glacial Maximum to the Younger Dryas (layer 5), and the early Late Pleistocene (layers 10–11). Optically stimulated luminescence (OSL) dates from layer 11 indicate that deposition took place c. 125–105 kya (Nian et al. Reference Nian, Zhou and Qin2009), during the early phases of Marine Isotope Stage 5. The faunal assemblage from layer 11 is dominated by horse (Equus caballus), onager (Equus hemionus) and auroch (Bos primigenius). Skeletal elements from late Middle and early Late Pleistocene fauna were also identified in this layer (Li & Dong Reference Li and Dong2007). The high proportion of limb elements (>60 per cent) and high frequency of cut marks observed on midshafts (approximately 34 per cent) suggest that Lingjing layer 11 was a kill-butchery site (Zhang et al. Reference Zhang, Li, Zhang and Gao2012).
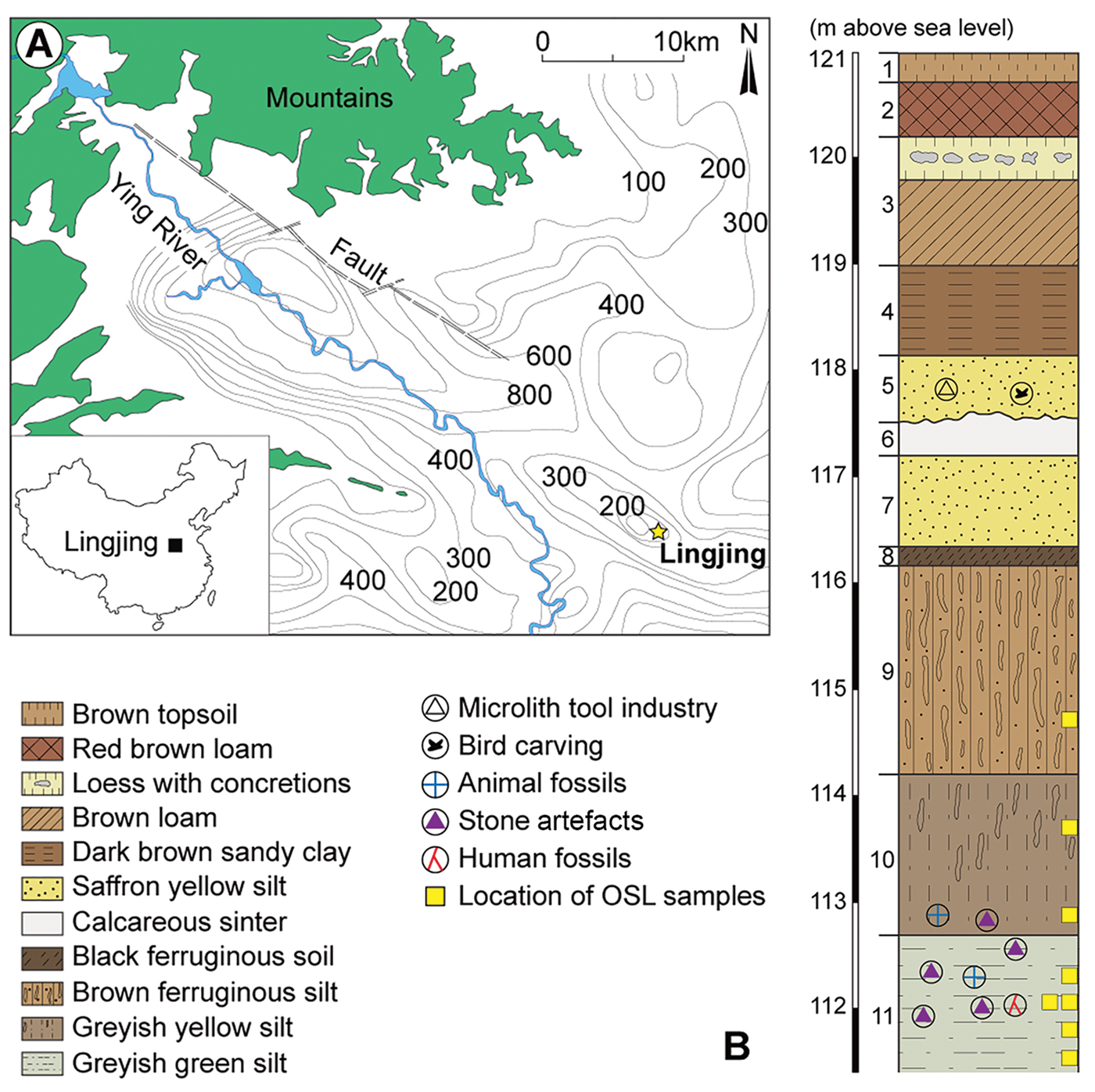
Figure 1. A) Location of Lingjing (Henan Province, China); B) stratigraphy indicating the geological and cultural layers (after Doyon et al. Reference Doyon, Li, Li and d'Errico2018).
The lithic assemblage from layer 11 comprises mostly quartz and quartzite artefacts. The presence of cores, flakes, formal tools and debris, plus the identification of use wear on some artefacts (Li Reference Li2007; Li et al. Reference Li, Li, Gao, Kuman and Sumner2019), of bone retouchers, organic soft hammer and pressure flakers (Doyon et al. Reference Doyon, Li, Li and d'Errico2018, Reference Doyon, Li, Li, Wang and Zhao2019), suggest that knapping activities, including tool manufacture and use, occurred at the site. The two engraved bone fragments described in this study also derive from layer 11. Discovered in 2009, their engravings were later identified during analysis of the faunal assemblage conducted in 2016.
Methods
Excavation methods at Lingjing involved the careful removal of sediments using curved-tipped trowels to avoid damaging finds, the 3D plotting of bone remains and lithic artefacts, and sediment sieving through 2mm mesh. Lithic and faunal remains are cleaned using soft brushes under running water. When present, sediment concretions are not removed from the faunal remains.
The artefacts described in this study are curated at the Henan Provincial Institute for Cultural Relics and Archaeology, Zhengzhou, China. Permission was granted to analyse one object (9L0141) at the Raman and Scanning Electron Microscopy Facilities of the Shandong University's Institute of Measurement and Testing. The specimens were examined visually, and then photographed with Canon PowerShot 100 and Nikon D300 AF Micro Nikkor 60 mm f/2.8D cameras. Microscopic analysis was conducted using a Leica Wild M3C stereomicroscope equipped with a Nikon CoolPix 900 digital camera at magnifications ranging from 4–40×. Data on the morphology and features of the engraved lines were recorded, along with the presence of a red residue. Identification of the cause of the modifications on the bone's surface is based on criteria inferred from the experimental reproduction and subsequent microscopic analysis of sequential marks produced on bone using different tools and motions (d’Errico Reference d'Errico1995; Fritz Reference Fritz1999).
A total of 227 bone fragments from the 2005–2015 excavations exhibiting few or no concretions were examined microscopically. The following variables were recorded for those specimens that exhibit indisputable cut marks (Noe-Nygaard Reference Noe-Nygaard1987; Lyman Reference Lyman1994; Fisher Reference Fisher1995; Fernández-Jalvo & Andrews Reference Fernández-Jalvo and Andrews2016): number, arrangement (divergent, overlapping, parallel or sub-parallel), morphology (curved, sinuous or straight), edge morphology (clean or fringed), and the occurrence of side striations, conspicuous changes in direction, surface micro-breaks and the presence of red residue.
A selected area of the engraved part of specimen 9L0141 was replicated using Coltene President regular body dental elastomer (Coltène, Switzerland). The negative replica was analysed with a Sensofar S-Neox confocal microscope driven by the SensoScan 6 software (Sensofar, Barcelona). The data was processed with SensoMap 7.3 software. Specimen 9L0141 and a sample of sediment from the cancellous bone were analysed with SEM-EDS and Raman spectroscopy. Sediment from layer 11 was analysed with ED-XRF (for details, see OSM 3).
Results
Taphonomic analysis
The surfaces of the analysed faunal remains are, when not affected by taphonomic processes or coated by concretions, exceptionally well preserved. Cut marks are the main anthropogenic modification observed on the faunal assemblage (23.79 per cent; Figure 2; OSM 4). They are present on all skeletal elements, including ribs (12.96 per cent). They show morphological traits typical of cut marks on fresh bone (Fisher Reference Fisher1995) and very rare changes in direction (OSM 4). Root etching affects 22.03 per cent of the analysed sample, while gnawing and etching resulting from consumption by carnivores affect 3.96 and 2.20 per cent of the sample, respectively. Other modifications include percussion marks possibly associated with marrow extraction (3.52 per cent), traces of use as retouchers (3.08 per cent) and staining produced by heat (1.32 per cent). Other than on specimen 9L0141, no red residues were detected on the 227 sampled faunal bone fragments.

Figure 2. Well-preserved sets of cut marks on faunal remains from Lingjing, layer 11 (photographs by F. d'Errico & L. Doyon).
Microscopic analysis
Although specimen 9L0141 is too fragmentary to propose a firm taxonomic attribution based on morphology (Figure 3), the thickness and flatness of the cortical bone and the morphology of the cancellous bone suggest that 9L0141 is the distal portion of an adult, large-sized mammal rib. The periosteal surface is pronounced and fibrous, comprising natural, alternating sub-parallel grooves and ridges (OSM 6). The four broken edges are ancient, as indicated by their colour and the slight smoothing of their surfaces under the microscope. The morphology of the broken edges and orientation indicate that they occurred on weathered bone (Villa & Mahieu Reference Villa and Mahieu1991; Lyman Reference Lyman1994). Seven sub-parallel lines cross the periosteal surface (Figure 4A). The abrupt terminations of L1 and L2, and the lower terminations of L3–5 indicate that they were interrupted by the adjacent fractures (Figure 5A; OSM 6). The upper terminations of L3–5 were engraved after the piece had already fragmented (OSM 6). L6 and L7 are complete and uninterrupted. The grainy appearance of the surface of all of the lines (Figure 6), their micro-fringed outlines, and the step micro-fractures produced when incising through the natural ridges of the bone surface (see OSM 6), all suggest that they were engraved on weathered rather than fresh bone (OSM 5).

Figure 3. Photographs of the engraved specimens: A) 9L0141; B) 9L0148 (photographs by F. d'Errico & L. Doyon).
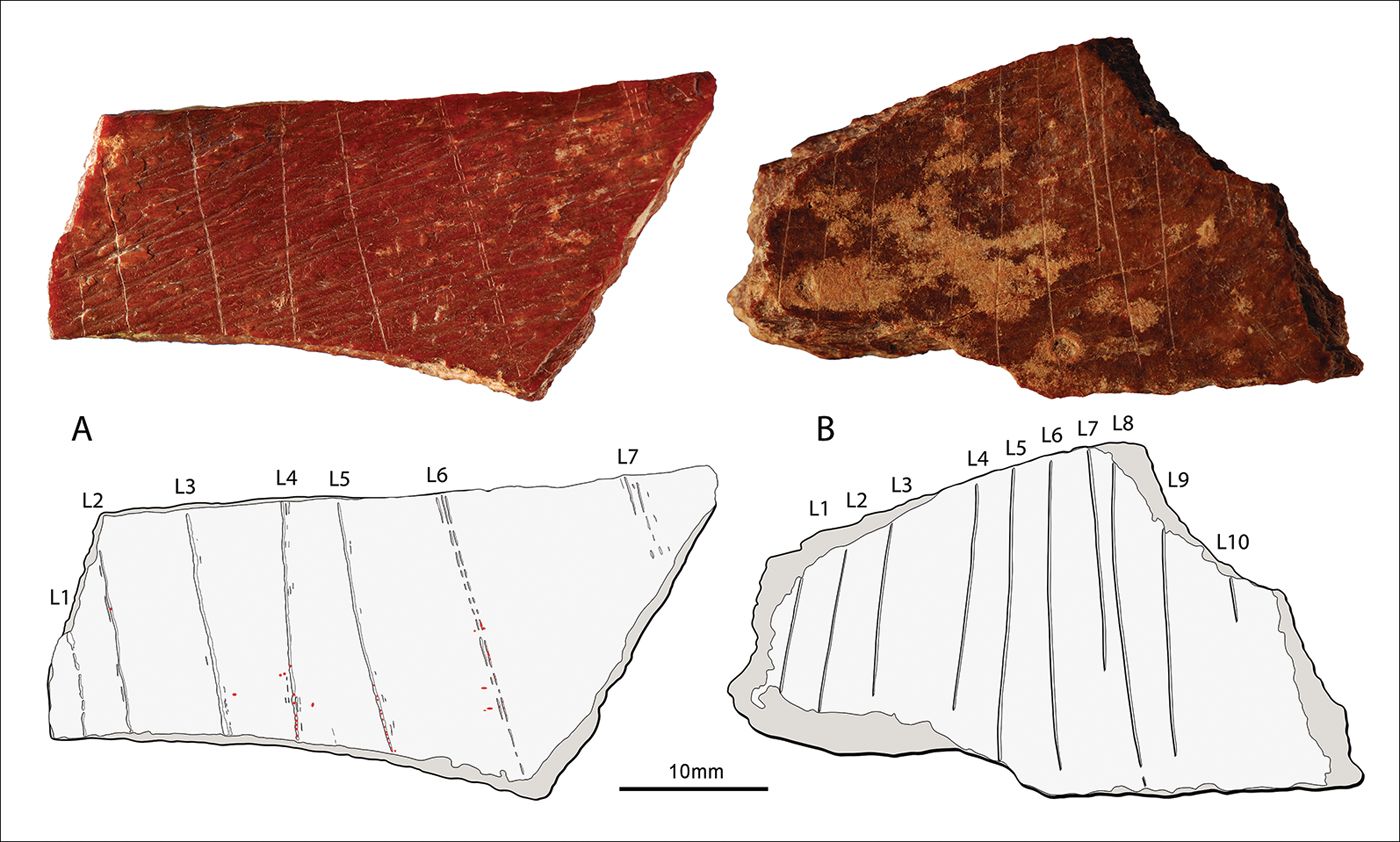
Figure 4. Photographs and tracing of the engraving on specimens: A) 9L0141 (red dots indicate the location of red residues); B) 9L0148 (photographs by F. d'Errico & L. Doyon).

Figure 5. Microscopic photographs of engraved lines on specimen 9L0141. Changes in direction of lines crossing the fibrous structure of the bone indicate the use of a sharp point (A–B). Red residues are present inside the engraved lines (C) (photographs by F. d'Errico & L. Doyon).

Figure 6. SEM photograph of specimen 9L0141 showing the grainy, slightly eroded appearance of L4 (photographs by H. Li, Q. Wang & Z. Zhang).
The depths of L1–5 remain constant when crossing the micro-grooves and ridges of the fibrous periosteal surface (Figure 5A–B), analysis of which shows that the grooves vary between 50 and 100μm in depth and between 100 and 600μm in width (OSM 6). Evidently, the tool tip was extremely sharp and no more than 50–100μm in width. The frequent, sudden changes in direction of L1–6 (Figure 5B–C) suggest instances of the momentary loss of control in the engraver's motion, due to tool speed changes when crossing the natural undulations of the bone's surface (d'Errico Reference d'Errico1994, Reference d'Errico1995). L1–5 were each engraved by a single passage of the tool. L6–7 comprise, respectively, three and five close, parallel and discontinuous striations each produced by a single passage of the point (OSM 6).
Similarities in section and width indicate that L1–2 were engraved by the same point, which created a narrow groove with a flat bottom (Figure 5A). L3–5 were also produced by the same point, which generated a wider and shallower groove with internal striations similar to each other, and side-striations on both sides of the main line. The multiple striations that comprise L6–7 were probably made by the same tool that was used to engrave L3–5, although the former is more superficial (OSM 6). The morphological differences between L1–2 and L3–7 can either be attributed to a tool change, or to the wear or breakage of the point used to engrave L1–2. The location of the micro-fractures on the periosteal ridges indicates that the lines were engraved from the longer to the shorter edge of the bone (Figure 5A; OSM 7). The sections of well-defined lines are asymmetrical to the right (see OSM 5), which, considering the motion of direction, indicates a right-handed engraver (Bosinski et al. Reference Bosinski, d'Errico and Schiller2002; d’Errico et al. Reference d'Errico, Doyon, Colagé, Queffelec, Vraux, Giacobini, Vandermeersch and Maureille2018). Right-handed engravers generally create sequences of parallel lines by juxtaposing them from left to right (d’Errico Reference d'Errico1992). This would favour the interpretation that the morphological differences between L1–2 and L3–7 are the result of wear or breakage of the engraving tool.
Specimen 9L0148 is a rib fragment from an adult, large-sized mammal (Figure 3B). The flat periosteal surface displays areas of chemical etching (whitening) (Figure 7A), removal of primary bone lamellae and parallel sets of thin striations, which are attributable to trampling. The top, bottom and left broken edges are eroded and fringed; those on the right side are fresher, suggesting that the fragment experienced at least two distinct breakage events. Ten sub-parallel lines (L1–10), produced before breakage of the rib (see OSM 5), cross the periosteal surface (Figure 4B). Their narrowness, similar internal morphology and the absence of side striations demonstrate that they were engraved using the same sharp lithic point, in a single session (Figure 7B). Slight changes in direction when crossing natural micro-fractures and taphonomically damaged areas, along with their irregular outline, suggest that the lines were engraved on weathered bone. They were cut using a quick motion towards the wider edge of the object, as indicated by the narrow terminations of L6–10.

Figure 7. Close-up views of L6, L7 and L8 on specimen 9L0148: A) chemical etching affecting the bone's surface subsequent to the engraving; B) similar internal morphology indicating the use of the same tool (photographs by F. d'Errico & L. Doyon).
Residue analysis
Abundant red residues were observed microscopically on specimen 9L0141 within lines L2 and L4–6, and adjacent to lines L3–6 (Figure 4A; OSM 7). The whitish sediment from layer 11, which was trapped in the specimen's cancellous bone, showed no such evidence of residue (see OSM 7). Raman analysis of the red residues from within the lines produced a composite spectrum with seven peaks at 225, 293, 411, 497, 612, 1242 and 1271cm−1 (see OSM 7). The five peak values in the low frequencies identify the residue as haematite (de Faria et al. Reference de Faria, Venâncio Silva and de Oliveira1997). Raman analysis of the sediment from the cancellous bone identified only quartz and a single instance of a particle containing haematite (see OSM 7). SEM-EDS analysis reveals that the sediment comprises particles ranging from 20–200μm in size, and is mainly composed of mica—either muscovite or biotite—embedded in a clay matrix, which also contains small quantities of iron (Fe), magnesium (Mg) and calcium (Ca). Single instances of quartz grain, zircon, bone fragment and an iron-rich particle are also detected (see OSM 7). ED-XRF analysis of sediment from layer 11 confirms that silicon (Si) is the predominant chemical element, followed by aluminium (Al) and potassium (K); iron (Fe) accounts only for 2.4–2.6 per cent (see OSM 7).
Discussion and conclusion
The lines on bone specimens 9L0141 and 9L0148 cannot result from excavation, cleaning or curation of the faunal assemblage. They differ in a number of respects from the lines exhibited on cut-marked bones from within layer 11. Cut marks on the latter generally display microscopic features consistent with cuts made on fresh bone. Generally, sets of close cut marks made in rapid succession on fresh bone by the same cutting edge rarely exceed four cuts, and they usually slightly diverge from a focal point, keeping a similar outline. In contrast, the lines on the two specimens—particularly on 9L0141—were made on partially weathered fragments; several lines on this specimen were engraved when the bone was already fragmented. The lines on 9L0141 were produced by an extremely sharp point, and the prehistoric individual was particularly careful when engraving the first five lines, given the rugged topography of the periosteal surface. To increase the visibility of the subsequent lines (L6–7), the engraver marked them using multiple strokes. Combined, this evidence does not support an interpretation of the lines as evidence of butchery activity on 9L0141, but rather, deliberate engraving of the bone. Although less striking, the lines on 9L0148 still differ from fresh cut marks on faunal remains from the same layer in their number, morphology and in the tool used to produce them. Furthermore, damage to some of the lines by chemical alteration of the periosteal surface indicates that they are of ancient origin, attributable to neither excavation nor curation.
The numerous haematite-rich residues identified within four lines on 9L0141 and the virtual absence of iron-rich particles in the clay sediment of layer 11 is puzzling. Considering the chemical composition of layer 11, one would have expected to find white residue composed of clay and micas at the bottom of the engraved lines. The absence of white sediment and the presence instead of haematite-rich deposits in the lines and nearby recessed areas could be explained by the smearing of ochre powder on the bone surface to highlight the engraved pattern and increase its visibility—a common practice in Upper Palaeolithic mobiliary art (Buisson et al. Reference Buisson, Menu, Pinçon and Walter1989; García et al. Reference García, Bonilla, Marín and Mascarós2016). The use of ochre to modify the appearance of personal ornaments is attested at 80 ka BP from Middle Stone Age Moroccan sites, such as Taforalt, Rhafas and Ifri n'Ammar (d’Errico et al. Reference d'Errico, Vanhaeren, Barton, Bouzouggar, Mienis, Richter, Hublin, McPherron and Lozouet2009). The earliest evidence for the use of ochre in China comes from Zhoukoudian Upper Cave and Shuidonggou localities 2 and 8, in the form of modified ochre lumps and ornaments coloured with ochre (Pitarch Martí et al. Reference Pitarch Martí, Wei, Gao, Chen and d'Errico2017).
A growing body of evidence from Europe and Southeast Asia (Table 1; OSM 1) supports the hypothesis that the cultural adaptations of archaic hominins involved symbolically mediated behaviour, thereby challenging the notion that modern cognitive abilities are restricted to Homo sapiens. While many scholars now agree on this hypothesis with regard to Neanderthals, the present article offers the first evidence to suggest that the same may also apply to Denisovans—the probable creators of the engravings described here. The Lingjing engravings represent the first possible example of such behaviour in Eastern Asia to pre-date 40 kya. Results were obtained by combining contextual and taphonomic data, with detailed analyses of the objects. The deliberate nature of the markings on 9L0141—and probably on 9L0148—as well as the application of ochre on the former appear to confirm the intentional character of these practices, and is fully consistent with the hypothesis that meaning may have been attributed to the patterns, to the moment in which the action took place, or to the action itself.
We are still far from understanding the meaning of these engravings for the archaic human groups living in China during the early Late Pleistocene. Doyon et al.’s (Reference Doyon, Li, Li and d'Errico2018, Reference Doyon, Li, Li, Wang and Zhao2019) recent identification of bone and antler fragments from layer 11 at Lingjing that were used to retouch lithics demonstrates that the Lingjing hominins were familiar with the mechanical properties of weathered bone and considered it to be a suitable raw material for producing artefacts. The Lingjing engravings suggest that these populations also saw bone as a medium on which they could permanently record sequential markings and use ochre as a substance to help highlight them. Future research may identify spatiotemporal consistencies that could offer clues to help in fully evaluating the significance of these behaviours.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.15184/aqy.2019.81
Acknowledgements
We wish to thank Alain Queffelec (CNRS—UMR5199 PACEA) and François-Xavier Le Bourdonnec (CNRS—UMR5060 IRAMAT-CRP2A) for their help with the sediment analyses. The scientific research at Lingjing is funded by grants from the National Natural Science Foundation of China (41630102, 41672020) and Shandong University 111 Project (111-2-09). The present study was funded by the Sino-French Cai Yuanpei programme (36707NF). This work was partially supported by the Research Council of Norway through its Centre's of Excellence funding scheme, SFF Centre for Early Sapiens Behaviour (SapienCE), project number 262618. L.H. acknowledges the support from the Strategic Priority Research Program of the Chinese Academy of Sciences, and the Chinese Academy of Sciences Pioneer Hundred Talents Program. L.D. was granted financial support from the China/Shandong University International Postdoctoral Exchange Program, and the Institut écologie et environnement of the Centre National de la Recherche Scientifique (CNRS-InEE). PACEA (UMR5199 CNRS) is a partner team of the Labex LaScArBx-ANR n° ANR-10-LABX-52.








