



Introduction
Outcrops of Laurentian lower Cambrian (Cambrian Series 2) strata extend from Alabama and Tennessee to North-East Greenland along the Eastern Seaboard of North America (Palmer, Reference Palmer and Holland1971; Hatcher et al., Reference Hatcher, Thomas and Viele1989; Williams, Reference Williams1995; Derby et al., Reference Derby, Fritz, Longacre, Morgan and Sternbach2012; Torsvik and Cocks, Reference Torsvik and Cocks2016; Fig. 1). Traditionally, their age has been determined by occurrences of trilobites indicative of the Dyeran Stage of North American usage (Cambrian Stage 4), with widespread records of Olenellus Hall in Billings, Reference Billings, Hitchcock, Hitchcock, Hager and Hitchcock1861 and related trilobites in the literature (Resser, Reference Resser1938a; Resser and Howell, Reference Resser and Howell1938; Bird and Rasetti, Reference Bird and Rasetti1968; Palmer, Reference Palmer and Holland1971; Skovsted, Reference Skovsted2006; Stein, Reference Stein2008; Webster and Hageman, Reference Webster and Hageman2018). By contrast, extensive faunas of older Cambrian trilobite faunas indicative of the Montezuman Stage (Cambrian Stage 3, but see Geyer, Reference Geyer2019), are well known from the opposite side of Laurentia, in the western United States (Hollingsworth, Reference Hollingsworth2011). However, fossiliferous Avalonian successions in Rhode Island and Massachusetts, New Brunswick, Nova Scotia, and eastern Newfoundland (Fig. 1.2), comprising the Terreneuvian (Cambrian Series 1) of Landing et al. (Reference Landing, Peng, Babcock, Geyer and Moczydlowska-Vidal2007) and Cambrian Series 2 to Lower Ordovician, are juxtaposed against the Laurentian of the Eastern Seaboard of present-day North America.
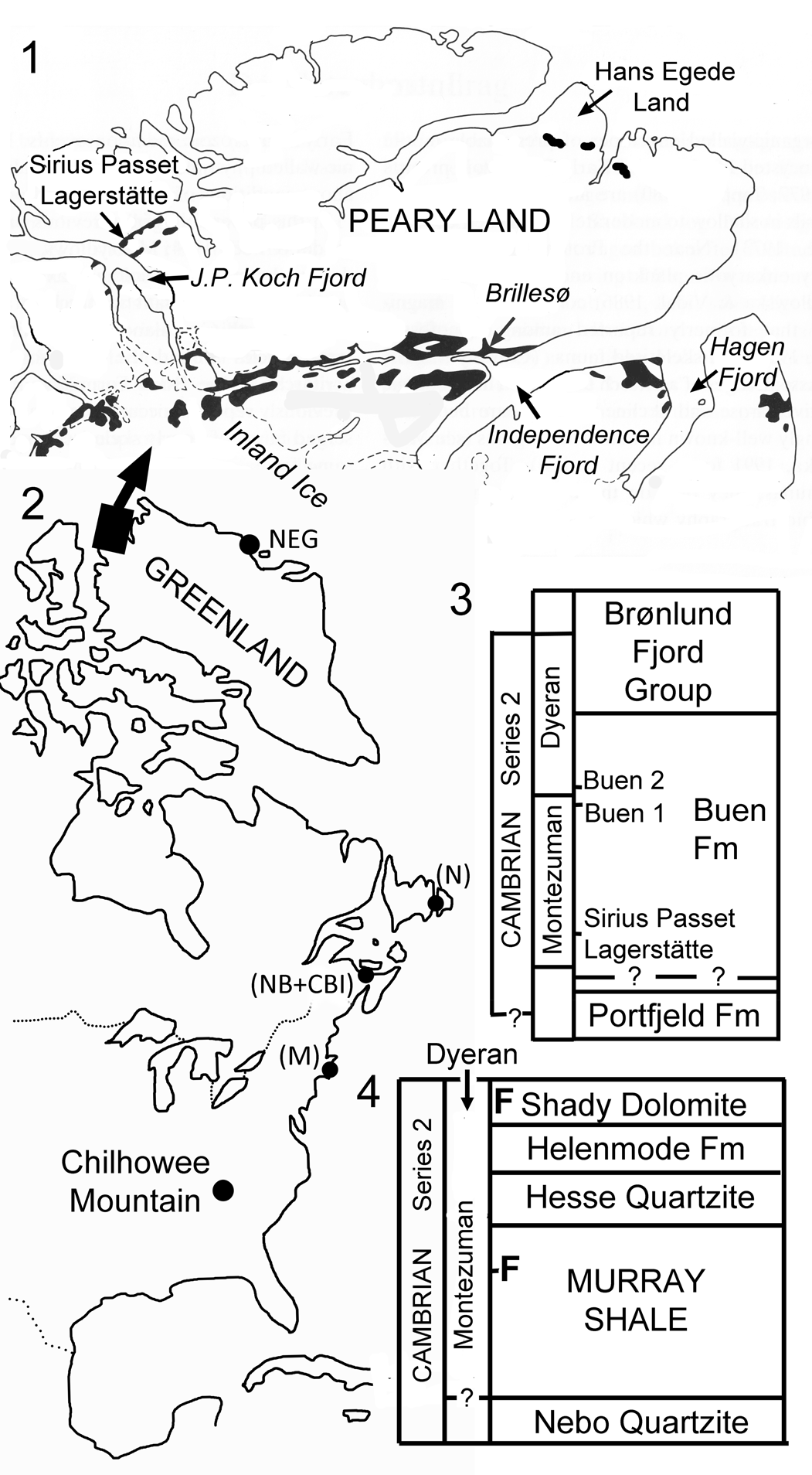
Figure 1. Localities and stratigraphy. (1) Peary Land region of North Greenland showing outcrops of the Buen Formation (black) and localities with Montezuman Stage fossil assemblages; (2) Eastern Seaboard of North America with Greenland displaced southward to its approximate position in the Cambrian. NEG indicates Dyeran occurrences in North-East Greenland. (M), (NB + CBI), and (N) locate Avalonian successions in Massachusetts, New Brunswick and Cape Breton Island, and eastern Newfoundland, respectively; (3) early Cambrian stratigraphy in southern Peary Land showing derivation of fossil assemblages with the Buen Formation; (4) early Cambrian stratigraphy at Chilhowee Mountain, Tennessee, indicating location of fossiliferous samples (F) in the Murray Shale (Montezuman Stage) and the established Dyeran Stage faunas of the Shady Dolomite in Virginia (Byrd et al., Reference Byrd, Weinberg and Yochelson1973; Fritz and Yochelson, Reference Fritz and Yochelson1988; McMenamin et al., Reference McMenamin, Debrenne and Zhuravlev2000).
The recent description of the nevadioid trilobite Buenellus Blaker, Reference Blaker1988 from the upper Murray Shale of Chilhowee Mountain, eastern Tennessee (Fig. 1.2, 1.4) is significant in providing evidence of the Montezuman Stage in the Laurentian terrane of the eastern United States (Webster and Hageman, Reference Webster and Hageman2018). Buenellus is otherwise known only from its type locality in the Sirius Passet Lagerstätte (Fig. 1.1) of Peary Land, North Greenland (Blaker and Peel, Reference Blaker and Peel1997; Babcock and Peel, Reference Babcock and Peel2007; Ineson and Peel, Reference Ineson and Peel2011; Peel and Willman, Reference Peel and Willman2018).
The Sirius Passet Lagerstätte is the oldest known fossil assemblage within a succession of Montezuman–early Dyeran (Cambrian Stages 3–4) age assigned to the Buen Formation (Ineson and Peel, Reference Ineson and Peel1997; Peel and Willman, Reference Peel and Willman2018; Fig. 1.3). The Lagerstätte is known only from a single locality, but the siliciclastic sediments of the Buen Formation are otherwise widely distributed in eastern North Greenland (Higgins et al., Reference Higgins, Ineson, Peel, Surlyk and Sønderholm1991a, Reference Higgins, Ineson, Peel, Surlyk, Sønderholm and Trettinb; Ineson and Peel, Reference Ineson and Peel1997, Reference Ineson and Peel2011; Peel and Willman, Reference Peel and Willman2018; Fig. 1.1). The Buen Formation lies within the transarctic Innuitian Orogen, facing the Arctic Ocean (Higgins et al., Reference Higgins, Ineson, Peel, Surlyk and Sønderholm1991a, Reference Higgins, Ineson, Peel, Surlyk, Sønderholm and Trettinb). By contrast, the Murray Shale at Chilhowee Mountain, and other Laurentian outcrops along the Eastern Seaboard between Alabama and North-East Greenland (Fig. 1.2, 1.4), accumulated along the shore of the former Iapetus Ocean (Torsvik and Cocks, Reference Torsvik and Cocks2016). In Cambrian times, however, Laurentia occupied a tropical position, and this Iapetan margin faced to the south (Torsvik and Cocks, Reference Torsvik and Cocks2016).
The description of Buenellus chilhoweensis brings into focus other faunal elements that have been known from the Murray Shale since the late nineteenth century. The history of research into these, and the geological setting of the Murray Shale, were described in detail by Hageman and Miller (Reference Hageman and Miller2016) and Webster and Hageman (Reference Webster and Hageman2018), the former providing a detailed description of the succession of trace fossil assemblages. In addition to Buenellus, the bivalved arthropod Isoxys Walcott, Reference Walcott1890 was proposed by Walcott (Reference Walcott1890) with Isoxys chilhoweanus Walcott, Reference Walcott1890 as its type species, the latter illustrated also by Williams et al. (Reference Williams, Siveter and Peel1996). The bradoriid Indota tennesseensis (Resser, Reference Resser1938a) was established on the basis of material from Chilhowee Mountain (Resser, Reference Resser1938a), and it has been revised subsequently by Laurence and Palmer (Reference Laurence and Palmer1963), Siveter and Williams (Reference Siveter and Williams1997), and Streng and Geyer (Reference Streng and Geyer2019). Walcott (Reference Walcott1890) first noted the occurrence of hyoliths, and Resser (Reference Resser1938a, pl. 4, figs. 30, 31) illustrated two specimens identified as Hyolithes sp. indet. Webster and Hageman (Reference Webster and Hageman2018) reported numerous trace fossils from the fossiliferous samples (Hageman and Miller, Reference Hageman and Miller2016). Undescribed elements in the fauna include a possible pelagiellid mollusk, while numerous small (0.5 mm) circular tablets of unknown affinity that resemble specimens described by Peel and Willman (Reference Peel and Willman2018, fig. 16A–D) from Buen Assemblage 2 at Brillesø, southern Peary Land, North Greenland, have been observed on several shale surfaces.
Wood and Clendening (Reference Wood and Clendening1982) described acritarchs from the lower part of the Murray Shale at Chilhowee Mountain, proposing Medousapalla choanoklosma as a new genus and species for acritarchs with hollow, closed, and distally funnel-shaped processes. The holotype of M. choanoklosma was described from Locality 1 in Wood and Clendening (Reference Wood and Clendening1982), which is geographically closest to the material described herein. In total, Wood and Clendening (Reference Wood and Clendening1982) described a dozen sphaeromorphic and acanthomorphic acritarchs from two localities. The same year, Skiagia was erected by Downie (Reference Downie1982) for acritarchs displaying the same morphology as Medousapalla. Zang (Reference Zang, Alexander, Jago, Rozanov, Zhuravlev and Yu2001) consequently considered Medousapalla to be a junior synonym of Skiagia and included M. choanoklosma in Skiagia ornata (Volkova, Reference Volkova, Volkova, Zhuravleva, Zabrodin and Klinger1968). Skiagia ornata is a geographically widespread early Cambrian taxon potentially marking a stratigraphic level earlier than ca. 531 Ma (Moczydłowska, Reference Moczydłowska2002; see also Moczydłowska and Zang, Reference Moczydłowska and Zang2006 for a longer, up-to-date discussion on the stratigraphic importance of Skiagia and its use in correlation).
This paper describes hyolithids that occur together with Buenellus chilhoweensis in the Murray Shale. Although poorly preserved and not individually age diagnostic, these hyolithids appear to be the most abundant and diverse element of the fauna. As such, they help establish a more complete picture of the Murray Shale biota during the Montezuman Stage. The hyolithid assemblage is compared with the hyolith fauna occurring together with Buenellus higginsi Blaker, Reference Blaker1988 in the Sirius Passet Lagerstätte (Peel, Reference Peel2010; Peel and Ineson, Reference Peel and Ineson2011) and other Montezuman–early Dyeran horizons in North Greenland (Peel and Willman, Reference Peel and Willman2018).
Materials and methods
About 35 hyolithid specimens preserved in pale buff weathering shale were examined from locality CM3 of Webster and Hageman (Reference Webster and Hageman2018) on Chilhowee Mountain (Fig. 1.2, 1.4) where they occur together with Buenellus chilhoweensis in the upper Murray Shale (Webster and Hageman, Reference Webster and Hageman2018, p. 457; 35°44.817′N, 083°48.446′W). The specimens are crushed, but not completely flattened, and some are preserved as external and internal molds. Conchs dominate but most are broken (Fig. 2). Opercula occur as isolated fossils (Fig. 3) and in rare partially articulated associations with conchs (Fig. 2.11). Rare broken fragments of the paired appendages (helens) have been observed as isolated fossils.
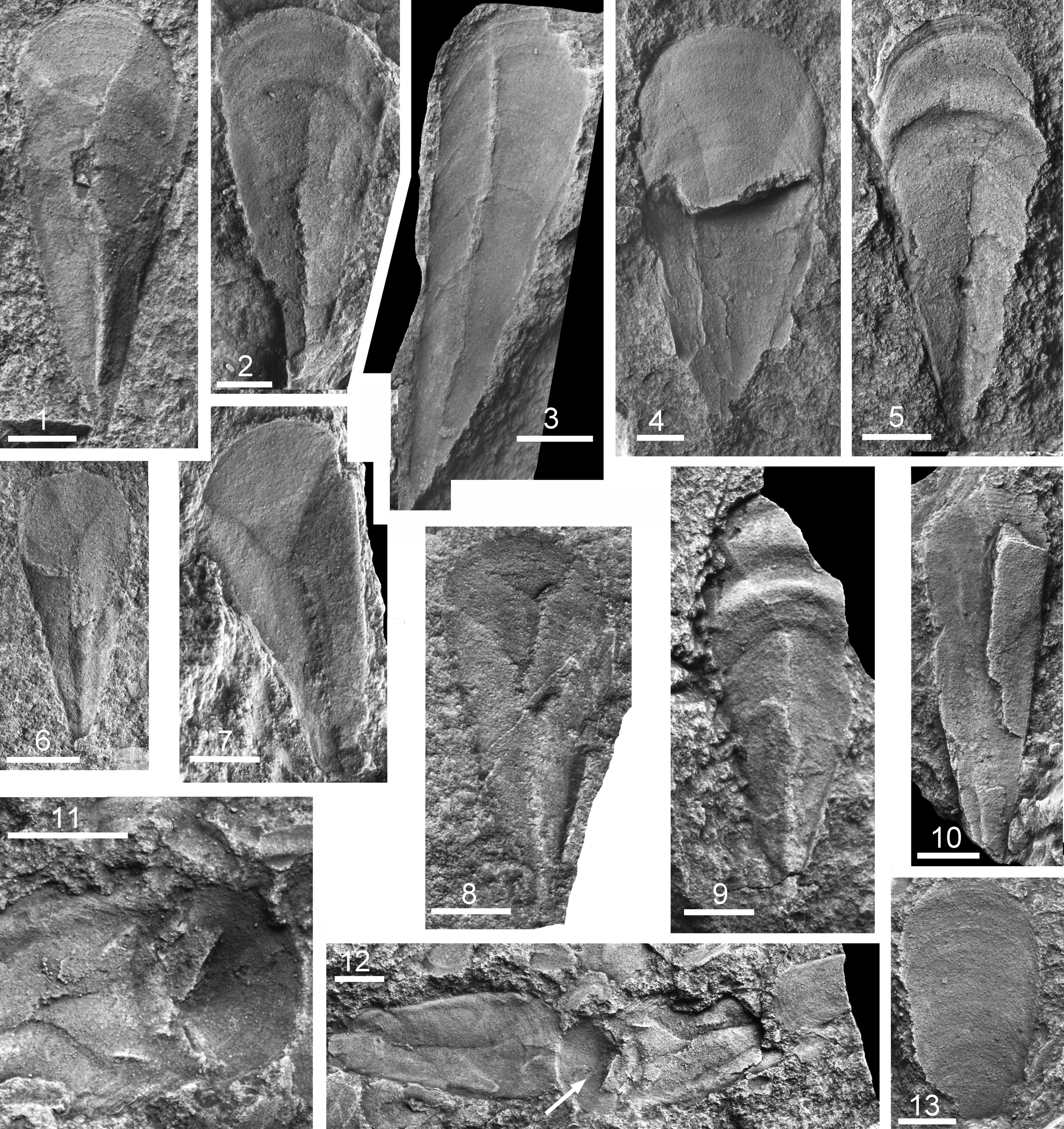
Figure 2. Hyolith conchs from the Murray Shale, Chilhowee Mountain, Tennessee. (1–4, 6–8) Burithes? sp. (1) PMU 35718, ventral surface; (2) PMU 35719, ventral surface; (3) PMU 35720, obliquely crushed ventral surface; (4) PMU 35721, central surface, above, with external mold of dorsal surface, below; (6) PMU 35722, crushed, with dorsal surface overlying internal surface of ventral surface with ligula; (7) PMU 35723, ventral surface with characteristic fractures; (8) PMU 35724, ventral surface. (5, 9) Hyolithid sp. 1, PMU 35725: (5) external and (9) internal molds showing prominent corrugation. (10) Hyolithid sp. 3, PMU 35726, dorsal view. (11–13) Hyolithid sp. 2; (11, 12) PMU 35727 with three fragmentary specimens in ventral aspect, probably within a gut fill, coprolite, or burrow; arrow in (12) locates external mold of operculum shown in detail in (11); (13) PMU 35728, external mold of ventral surface. Scale bars = 2 mm.
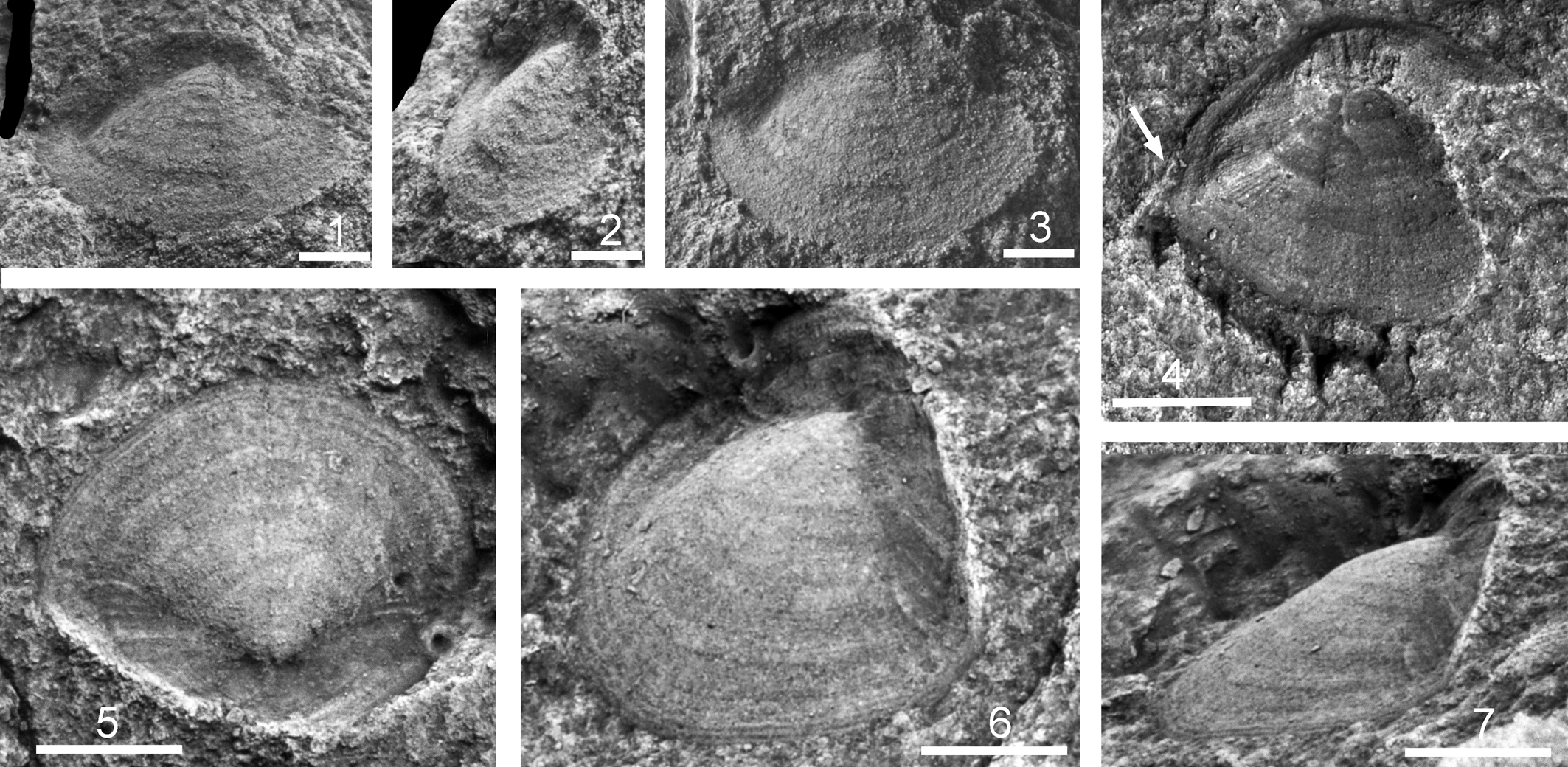
Figure 3. Hyolithid opercula from the Murray Shale, Chilhowee Mountain, Tennessee. (1–3) PMU 35729: (1) oblique apertural, (2) dorso-lateral, and (3) dorsal views. (4) PMU 37730 external mold with arrow indicating lateral sinus for passage of helen; (5–7) PMU 35731: (5) dorsal, (6) dorso-lateral, and (7) lateral views. (1–3) Scale bars = 1 mm; (4–7) scale bars = 2 mm.
After painting with a thin coat of black colloidal carbon, all specimens were whitened with ammonium chloride sublimate before routine examination and photography. Images were captured using a Lumenera Infinity X32 high-resolution USB camera with attached Micro Nikkor 55 mm lens and assembled in Adobe Photoshop CS4.
Several samples were washed and cleaned in distilled water, rinsed in hydrochloric acid (HCl), and finally treated with hydrofluoric acid (HF). Residues were screened for organic-walled microfossils, but deep weathering of these samples has resulted in destruction of original organic material and the introduction of contaminants.
Repositories and institutional abbreviations
The prefix MGUH indicates a specimen deposited in the paleontological type collection of the Natural History Museum of Denmark (formerly Geological Museum), Copenhagen. PMU indicates a specimen deposited in the paleontological type collection of the Museum of Evolution, Uppsala University, Sweden.
Systematic paleontology
Order Hyolithida Syssoiev, Reference Syssoiev1957
Family Angusticornidae Syssoiev, Reference Syssoiev1968
Remarks
Angusticornids are characterized by the acutely angular lateral transition between the flattened ventral surface and the convex dorsal surface (Malinky and Geyer, Reference Malinky and Geyer2019).
Genus Burithes Missarzhevsky, Reference Missarzhevsky1969
Type species
Linevitus distortus Syssoiev, Reference Syssoiev1962 from the Cambrian (late Terreneuvian) of the Aldan River, Siberia, by original designation.
Remarks
Missarzhevsky (Reference Missarzhevsky1969, Reference Missarzhevsky and Raaben1981) proposed Burithes from the Tommotian and Atdabanian stages (Cambrian Stage 2 and 3) of Siberia, with type species Linevitus distortus Syssoiev, Reference Syssoiev1962 from the Aldan River. The shallowly convex dorsum, considered ventral by Missarzhevsky (Reference Missarzhevsky1969, Reference Missarzhevsky and Raaben1981), passes by way of acute lateral angulations onto the almost flat ventral surface. Malinky (Reference Malinky2014) noted that all specimens in Syssoiev's (Reference Syssoiev1962) type material were internal molds and that details of ornamentation and the external expression of the lateral edges were not known. Consequently, Malinky (Reference Malinky2014) recommended that the name should be restricted to the type material, and he further established Haydenoconus, in which the lateral edges on the shell exterior are sharp, keel-like. However, the type species, Hyolithes gallatinensis Resser, Reference Resser1938b, was described from the Furongian of Wyoming although a second species, Haydenoconus prolixus (Resser, Reference Resser1939), was described from the Miaolingian of Idaho. Malinky (Reference Malinky2014) did not comment on the status of the other species that Missarzhevsky (Reference Missarzhevsky1969, Reference Missarzhevsky and Raaben1981), Meshkova (Reference Meshkova1974), Meshkova et al. (Reference Meshkova, Missarzhevsky, Sysoiev and Valkov1983), and Rozanov et al. (Reference Rozanov2010) referred to Burithes. While acknowledging Malinky's (Reference Malinky2014) recommendation, Burithes is employed in the present context on the basis of the descriptions and illustrations presented by Missarzhevsky (Reference Missarzhevsky1969, Reference Missarzhevsky and Raaben1981).
Burithes? sp.
Figure 2.1–2.4, 2.6–2.8
Description
Conch with incremental angle 20°–25° and ligula about one-sixth of total length (Fig. 2.1), with slight longitudinal curvature such that the ventral surface may have been shallowly convex (Fig. 2.3). Width of ligula about half its length, but length increasing proportionately with growth (Fig. 2.4). Shallow lateral sinuses for the likely passage of helens lie on the dorsal side of the angular transition from the convex dorsal surface to the almost flat ventral surface. Dorsal surface seemingly uniformly shallowly convex, but degree of inflation uncertain due to crushing. Ornamentation on ventral surface of fine comarginal growth lines with occasional growth halts that may appear periodic (Fig. 2.3). Ornamentation on dorsal surface poorly known, seemingly almost orthocline. Operculum and helens not certainly known, but associated isolated opercula are wider than long (Fig. 3), supporting the interpretation that the dorsal surface of the conch was not strongly inflated.
Material
PMU 35719–PMU 35724, Murray Shale, Montezuman Stage (Cambrian Stage 3), Chilhowee Mountain, Tennessee.
Remarks
All specimens are compressed, although a degree of separation between the dorsal and ventral surfaces may be maintained (Fig. 2.4). Crushing has often produced Y–shaped cracks that extend from the ligual margin down the median line as a raised, irregular ridge or angulation to near the apex (Fig. 2.1). The median fracture is commonly expressed as a ridge on the ventral surface, but this is a preservational artifact. However, a broad, rounded ridge in some specimens (Fig. 2.1) likely represents compaction around a solid object within the conch interior, possibly an early mineralized burrow or sediment-infilled section of gut (Devaere et al., Reference Devaere, Clausen, Álvaro, Peel and Vachard2014).
In terms of its overall shape, the Murray Shale conchs are similar to Burithes erum Missarzhevsky, Reference Missarzhevsky1969 from the Tommotian of the Anabar Massif of Siberia (Missarzhevsky, Reference Missarzhevsky1969, Reference Missarzhevsky and Raaben1981). Nevadotheca whitei (Resser, Reference Resser1938b) from the Pioche Shale (Cambrian Series 2) of Nevada, the type species of Nevadotheca Malinky, Reference Malinky1988, differs in having a high, inflated dorsum and narrowly rounded lateral margins. This is also the case in Nevadotheca boerglumensis Peel and Willman, Reference Peel and Willman2018 and Kalaallitia myliuserichseni Peel and Willman, Reference Peel and Willman2018, described from the early Olenellus Biozone (Dyeran, Cambrian Stage 4) of the Buen Formation of southern Peary Land, North Greenland, but Kalaallitia is distinguished by its fine longitudinal lirae and longer ligula. Opercula referred to these Peary Land species are proportionately longer than Murray Shale specimens, suggesting that the dorsal surface of their conchs was more strongly inflated than in Burithes? sp.
Malinky and Skovsted (Reference Malinky and Skovsted2004) noted some similarity between specimens from the Dyeran of North-East Greenland and Burithes, but the acute dorsum of their material suggested assignment to Grantitheca Malinky, Reference Malinky1989 by Peel and Willman (Reference Peel and Willman2018). In contrast to material from the Murray Shale and Buen Formation, hyoliths from North-East Greenland are preserved mainly as internal molds in limestone or as phosphatic residues from limestones (Malinky and Skovsted, Reference Malinky and Skovsted2004; Skovsted, Reference Skovsted2006).
Family uncertain
Hyolithid sp. 1
Figure 2.5, 2.9
Material
PMU 35725, internal mold and corresponding external mold of the ventral surface, Murray Shale, Montezuman Stage (Cambrian Stage 3), Chilhowee Mountain, Tennessee.
Remarks
This species, known from the illustrated specimens and two additional fragments, is characterized by two or three prominent transverse folds or corrugations on the adapertural part of the shallowly convex ventral surface of the conch and ligula (Fig. 2.5, 2.9). The incremental angle is about 30°, and the length of the ligula is slightly more than half its width. Lateral sinuses are present at the transition from the shallowly convex ventral surface to the dorsal surface, but the degree of inflation of the latter is not known. Ornamentation consists of fine comarginal growth lines, although these are more strongly developed at the preserved aperture (Fig. 2.5).
Corrugation of the latest growth stage of the ventral surface and ligula is common in hyolithids, but not with the high degree of emphasis seen in the Murray Shale specimen (Fig. 2.9). Examples were illustrated by Missarzhevsky (Reference Missarzhevsky1969, Reference Missarzhevsky and Raaben1981) in specimens from Siberia referred to Trapezovitus sinscus (Syssoiev, Reference Syssoiev, Orlov, Luppov and Drushchits1958) and Burithes cuneatus Missarzhevsky, Reference Missarzhevsky1969, by Qian et al. (Reference Qian, Li, Zhu and Yin2003) in Nitoricornus wushiensis Qian et al., Reference Qian, Li, Zhu and Yin2003 from the lower Cambrian of Xinjiang, China, and by Malinky (Reference Malinky1990, fig. 1.6) in specimens from New Brunswick.
Simpson and Sundberg (Reference Simpson and Sundberg1987) interpreted a specimen from the Hampton Formation of southwestern Virginia as a rugose hyolith compared to Tuojdachithes? biconvexus (Cobbold, Reference Cobbold1919), but Hageman and Miller (Reference Hageman and Miller2016) considered that it was not a fossil. Peel and Willman (Reference Peel and Willman2018, fig. 13G) illustrated a more-slender, unnamed hyolithid from Buen Assemblage 1 at Brillesø, southern Peary Land (Fig. 1.1) with rugose comarginal ornamentation on the ventral surface of the ligula, but this is much less prominent than in the Murray Shale specimens.
Hyolithid sp. 2
Figure 2.11–2.13
Material
PMU 35727–PMU 35728, Murray Shale, Montezuman Stage (Cambrian Stage 3), Chilhowee Mountain, Tennessee.
Remarks
The coincidence of the long axes of three overlapping incomplete specimens in ventral aspect (Fig. 2.12) suggests that they may form part of a gut fill or coprolite (Vannier, Reference Vannier2012). One of the specimens preserves the operculum in place (Fig. 2.11, arrow in 2.12). The incremental angle is about 20°, and the ligula is short. The ventral surface is shallowly convex, but the lateral edges are rounded and delimited on their axial edge by a shallow longitudinal furrow. The dorsal surface is not known, but lateral sinuses seem to be well developed (Fig. 2.11). The impression of the dorsal exterior of the operculum shows comarginal growth lines and a suggestion of a radial furrow. While similar in shape to that illustrated in Figure 3.5–3.7, it is too poorly preserved for closer comparison.
Hyolithid sp. 3
Figure 2.10
Material
PMU 35726, Murray Shale, Montezuman Stage (Cambrian Stage 3), Chilhowee Mountain, Tennessee.
Remarks
This poorly preserved specimen has an incremental angle of 15° and appears to have an oxygonal aperture. Adaperturally, shallowly convex comarginal growth lines on the ventral surface are cord-like and laterally discontinuous, resembling the pattern illustrated by Malinky (Reference Malinky1990) as Nitoricornus danianum (Matthew in Walcott, Reference Walcott1884) and by Kruse and Hughes (Reference Kruse and Hughes2016) as ?Crestjahitus danianus from New Brunswick, Canada, whereas the dorsal surface is ornamented with barely discernible fine growth lines.
Hyolithid opercula
Figure 3
Material
PMU 35729–PMU 35731, Murray Shale, Montezuman Stage (Cambrian Stage 3), Chilhowee Mountain, Tennessee.
Remarks
Three poorly preserved specimens of partially articulated hyolithid skeletons occur (Fig. 2.11, 2.12), but several isolated opercula are known (Fig. 3). The length of the best-preserved specimen is about four-fifths of its width (Fig. 3.5). Following terminology employed by Malinky and Berg-Madsen (Reference Malinky and Berg-Madsen1999, text-fig. 2), its conical shield is hemispherical in plan view, and the summit lies at about one-quarter of the distance from the adapical margin to the adapertural margin (Fig. 3.5). The folds separating the conical and cardinal surfaces delimit an angle of about 130° in plan view (Fig. 3.5). In lateral perspective (Fig. 3.7), the cardinal shield rises high above the summit of the operculum, with the inclination of its adapical margin (right in Fig. 3.7) suggesting that the conch had an amblyogonal margin. The cardinal area is ornamented with radial ridges transverse to the comarginal growth lines present over the entire conch. The latter are most conspicuous on the conical shield where radial ornamentation consists of fine lines (Fig. 3.5, 3.6).
A second specimen, preserved as an external mold, has a well-defined convex adapical border with prominent coarse ridges between the summit and the adapical margin (Fig. 3.4). A broad shallow sinus in the margin (Fig. 3.4, arrow) marks the exit point for the helen. A third specimen (Fig. 3.1–3.3) has a more elliptical shape than the other two specimens and develops a series of short comarginal rugae located medially on the conical shield.
The cardinal surface is proportionately longer in opercula of Nevadotheca boerglumensis and Kalaallitia myliuserichseni from the early Olenellus Biozone (Buen Assemblage 2) at Brillesø, southern Peary Land (Peel and Willman, Reference Peel and Willman2018, fig. 14), and the folds separating the conical and cardinal surfaces delimit an angle of about 90°, much smaller than in the specimens from the Murray Shale. In this respect, the opercula from the Murray Shale more closely resemble the opercula associated with articulated specimens from the Sirius Passet Lagerstätte (Fig. 4.2, 4.4), but these are too poorly preserved for close comparison. A similar wide angle is seen in Nitoricornus danianum illustrated by Malinky (Reference Malinky1990, fig. 1.1) from New Brunswick and assigned to ?Crestjahitus danianus by Kruse and Hughes (Reference Kruse and Hughes2016).

Figure 4. Articulated hyolithids from the Sirius Passet Lagerstätte, Buen Formation, Peary Land, North Greenland. (1) MGUH 29260; (2, 4) MGUH 29691: (2) ventral surface with operculum displaced and inverted to show conical surface; (4) enlarged view. (3) MGUH 29258, ventral surface. (1–3) Scale bars = 2 mm; (4) scale bar = 1 mm.
Faunal comparison
The Murray Shale hyolithids are the oldest hyoliths known from present–day eastern Laurentia but not from eastern North America. Hyoliths considered to be of Terreneuvian and younger age were reported from Avalonian terranes (Fig. 1.2) by Landing (Reference Landing1988, Reference Landing1991, Reference Landing1995), Landing et al. (Reference Landing, Myrow, Benus and Narbonne1989), Landing and Murphy (Reference Landing and Murphy1991), and Landing and Kröger (Reference Landing and Kröger2012). Malinky and Geyer (Reference Geyer2019, fig. 2) referred some of this material to the Montezuman, citing the occurrence of Aimitus Syssoiev, Reference Syssoiev1966 and Notabilitus Syssoiev, Reference Syssoiev1968.
Peel (Reference Peel2010) and Peel and Ineson (Reference Peel and Ineson2011) described hyolithids and orthothecids from the Sirius Passet Lagertstätte occurring together with Buenellus higginsi. Orthothecids have not been recognized from the Murray Shale. Although several hyolithid specimens from the Sirius Passet Lagerstätte preserve the operculum and paired helens together with the conch, their poor state of preservation precludes more precise identification. However, opercula from the Murray Shale are similar in shape to a Sirius Passet operculum (Fig. 4.2, 4.4) and unlike those occurring in the Dyeran Buen Assemblage 2 from southern Peary Land (Peel and Willman, Reference Peel and Willman2018). Variation in the incremental angle of the conch suggests that several taxa may be present in the Sirius Passet articulated material. Some Sirius Passet specimens have an angular dorsum not seen in Murray Shale material and thereby resemble Grantitheca Malinky, Reference Malinky1989, although this genus was not recognized by Landing and Bartowski (Reference Landing and Bartowski1996, p. 757). The articulated specimens are associated with longitudinally ribbed conchs assigned to Trapezovitus Syssoiev, Reference Syssoiev, Orlov, Luppov and Drushchits1958 and an orthothecid, but specimens similar to these have not been observed from the Murray Shale.
The bradoriid Indota Öpik, Reference Öpik1968, represented by Indota tennesseensis in the Murray Shale, has not been described from the Sirius Passet Lagerstätte. Peel and Willman (Reference Peel and Willman2018) tentatively referred poorly preserved specimens from the upper Buen Formation (Dyeran Stage) to the genus, but the assignment was questioned by Streng and Geyer (Reference Streng and Geyer2019). Isoxys chilhoweanus from the Murray Shale is similar to Isoxys volucris Williams, Siveter, and Peel, Reference Williams, Siveter and Peel1996, which is the most abundant fossil in the Sirius Passet Lagerstätte (Williams et al., Reference Williams, Siveter and Peel1996; Stein et al., Reference Stein, Peel, Siveter and Williams2010; Nielsen et al., Reference Nielsen, Rasmussen and Harper2017).
Peel and Willman (Reference Peel and Willman2018) illustrated two crushed hyolithids from Buen Assemblage 1 at Brillesø, southern Peary Land, in association with the nevadioid Limniphacos perspicullum Blaker and Peel, Reference Blaker and Peel1997 of presumed Montezuman age (Hollingsworth, Reference Hollingsworth2011). The rugose growth ornamentation of one of these (Peel and Willman, Reference Peel and Willman2018, fig. 13G) is reminiscent of Hyolithid sp. 1 from the Murray Shale but is much less strongly expressed. Strata within the Buen Formation of probable Montezuman age occur in Hans Egede Land, eastern Peary Land (Fig. 1.1); they are not well known, but unidentifiable hyolith fragments are associated with poorly preserved trilobites (Peel and Willman, Reference Peel and Willman2018).
Acknowledgments
E. Wallet (Uppsala) helped in the search of HF residues for organic-walled microfossils. E. Landing (Albany) and T. Topper (Stockholm) reviewed the manuscript. J.O.R. Ebbestad (Uppsala) assisted with the curation of specimens.





 Open access
Open access