



Along the Pacific west coast of South America, a long Middle Preceramic periodFootnote 1 (ca. 7000–5000 BP) bears many Mesolithic-like hallmarks, including a broadening resource base and increasing sedentism, population density, and social complexity. Here, as elsewhere, logistically organized, complex hunter-gatherers reduced their mobility by locating themselves in so-called ecotonally diverse settings: places where different habitats conjoined, offering access to diverse resources according to seasonal rounds (Beresford-Jones et al. Reference Beresford-Jones, Pullen, Whaley, Moat, Chauca, Cadwallader, Arce, Orellana, Alarcón, Gorriti, Maita, Sturt, Dupeyron, Huaman, Lane and French2015; Standen et al. Reference Standen, Santoro, Arriaza and Colleman2017).
Coasts and estuaries are quintessential ecotonally diverse environments, providing rich, sustainable, and easily seen and harvested resources. In many places worldwide, Mesolithic broad-spectrum economies seem to have relied on the systematic exploitation of marine resources (Stiner Reference Stiner2001). Nowhere, however, is the connection between the development of sedentism and coastal productivity more relevant than the Pacific coast of South America. Although the littoral itself is arid desert, it is traversed by numerous riverine oases along the watercourses that rise in its Andean hinterlands, and its offshore marine resources are prodigious, sustained by nutrients in cold upwellings that today make the seas off the coast of Peru the world's richest fishery (FAO 2014).
Moseley's (Reference Moseley1975) Maritime Foundations of Andean Civilization (MFAC) hypothesis suggested that Late Preceramic civilization first arose along this coast based on the exploitation of marine resources, challenging the axiom that agriculture is necessary for the rise of complex societies. However, terrestrial resources—including gathered and, increasingly, cultivated plant foods—are also evident in the Preceramic archaeological record, so that precisely how subsistence patterns unfolded here as agriculture emerged continues to be debated (Prieto and Sandweiss Reference Prieto and Sandweiss2020). Although many sites are reported dating to the end of the Middle Preceramic when settled villages first started to coalesce along this Pacific coast, the “type-site” for this period has long been Paloma on the south-central coast of Peru (Figure 1).

Figure 1. The Pacific coast of South America showing the Preceramic sites discussed in the text.
In this article we present a refined chronological context for three of the first villages on the Peruvian coast—Paloma, Chilca I, and La Yerba III—by calibration and Bayesian modeling of their radiocarbon dates. We then present the results of stable isotope analyses of 33 individuals—29 interred at Paloma, 1 from Chilca I, and 3 from La Yerba III—alongside osteological analyses of 62 individuals interred at Paloma and 28 at Morro I on the north coast of Chile. Together these data offer direct evidence of diet and lifestyle for the critical period foreshadowing the emergence of Late Preceramic civilization on this Pacific coast.
The Archaeological Context: The First Villages of the Middle Preceramic
The early village sites of Paloma and Chilca I both lie in the Chilca Valley on the central coast of Peru, 3.5 and 4.5 km from the Pacific Ocean, respectively, whereas La Yerba III is on the Río Ica on the south coast of Peru, 2.5 km from the sea (Figure 1). Morro 1, meanwhile, is not a habitation site but a cemetery at Arica on the north coast of Chile that contains multiple “Chinchorro” inhumations over many millennia, although habitation sites in its vicinity provide further insight into lifestyles persisting over that time. All these sites enjoy remarkable preservation conditions in an arid climate.
Although the archaeological records of Paloma, Chilca I, La Yerba III, and Morro I therefore entail some differences, not least in their respective time depths, they each have contemporaneous components dating to around 6000 cal BP, as shown in Figure 2, with broadly comparable material cultures and human ecologies.
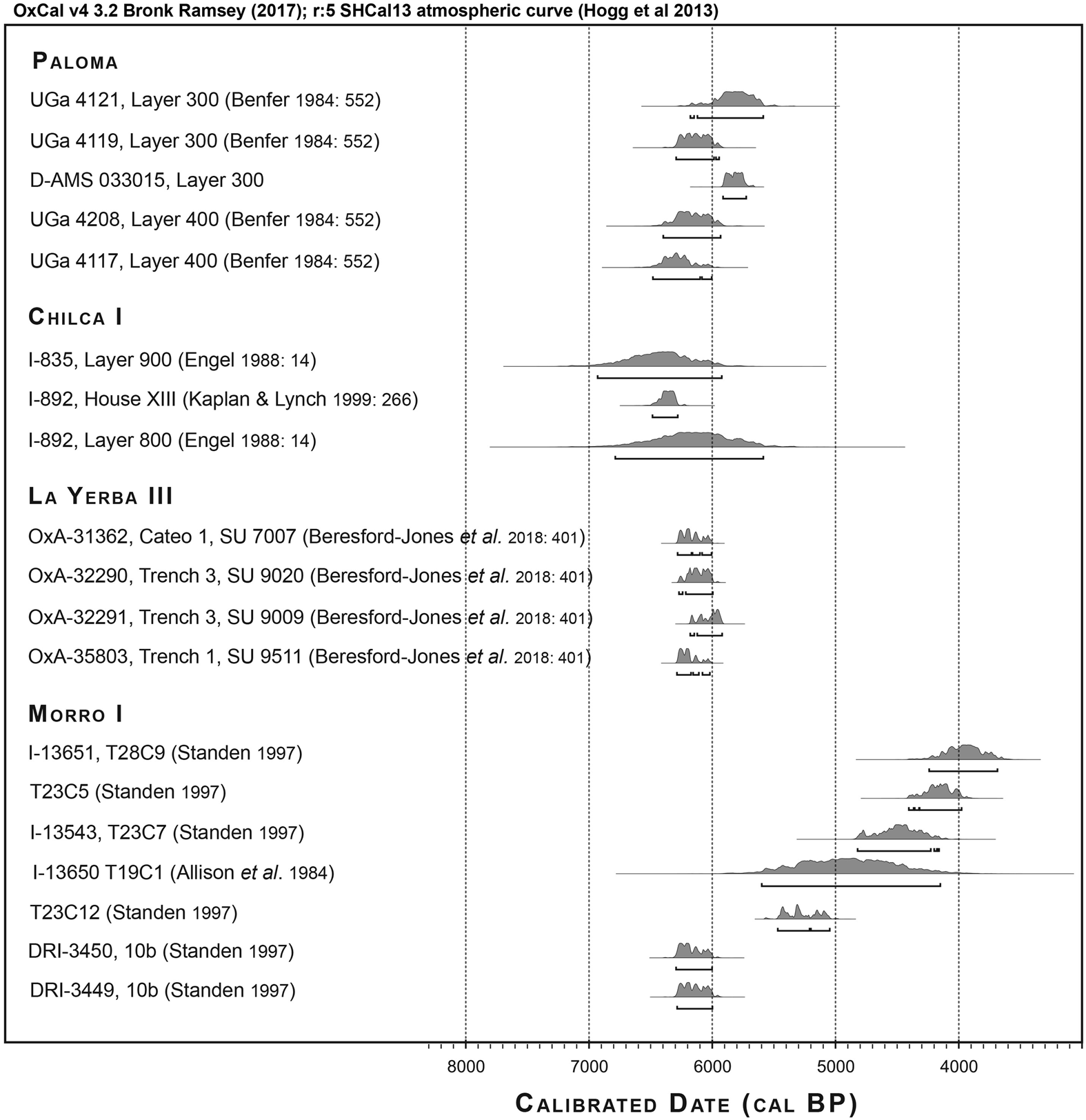
Figure 2. Radiocarbon dates for Paloma, Chilca I, La Yerba III, and Morro I, all calibrated using ShCal13 (Hogg et al. Reference Hogg, Hua, Blackwell, Niu, Buck, Guilderson, Heaton, Palmer, Reimer, Reimer, Turney and Zimmerman2013).
Permanent Settlement, Increasing Population, and Intensifying Networks
The archaeological assemblages at Paloma, Chilca I, La Yerba III, and Morro I reflect a prolongation of marine hunting and gathering lifestyles that had endured along this littoral since its first occupation, although the distribution and availability for human harvesting of marine resources changed over time, with variations both in the El Niño Southern Oscillation (ENSO; Sandweiss et al. Reference Sandweiss, Andrus, Kelley, Maasch, Reitz and Roscoe2020) and in coastal morphology, itself subject to changing eustatic sea levels until around 6000 BP.
Paloma, Chilca I, and La Yerba III show some of the earliest evidence for permanent, sedentary occupation in the Peruvian archaeological record. Their locations were determined, first, by proximity to permanent sources of freshwater. Coastal rivers here have highly seasonal surface flow, but especially in their lower reaches, groundwater supplies are easily accessible year-round (Engel Reference Engel1991:58). Each of these villages comprises sequences of circular or ovoid semi-subterranean houses excavated into middens of previous occupations. The houses were constructed of frames of willow posts and sedge canes covered with finely woven mats of fibrous sedges or cattails; there was a small central hearth, and often they were separate patio areas with other hearths. Interiors were kept clean, with multilayered floor deposits showing continued occupation though time and containing heavy grinding stones (batanes), anvils, other furniture of whalebone, and multiple storage pits. The dead were interred with care and ritual, as individuals or sometimes in groups, in structured pit burials within the houses after their abandonment (Benfer Reference Benfer and Pearsall2008; Beresford-Jones et al. Reference Beresford-Jones, Pullen, Chauca, Cadwallader, García, Salvatierra, Whaley, Vásquez, Arce, Lane and French2018; Engel Reference Engel1980, Reference Engel1988; Gorbahn Reference Gorbahn2020; Quilter Reference Quilter1989; Supplemental Figure 1).
Along with these various traits of sedentism, including early hints of agriculture to be discussed shortly, came significant demographic change and increasing social complexity. Investigations at Paloma and La Yerba suggest population increases of around an order of magnitude through the millennium to 6000 cal BP (Benfer Reference Benfer and Pearsall2008:372; Beresford-Jones et al. Reference Beresford-Jones, Pullen, Chauca, Cadwallader, García, Salvatierra, Whaley, Vásquez, Arce, Lane and French2018; Reitz Reference Reitz2003).
Obsidian lithics at Paloma (Benfer Reference Benfer and Pearsall2008) and La Yerba III (Chauca et al. Reference Chauca, Glascock, Rodríguez, Arce S. and Beresford-Jones2019) derive from the Jichja Parco quarry at Quispisisa at 4,100 m asl in the Ayacucho highlands, which is 170 km and 310 km distant, respectively, from those sites. They are evidence for wide spheres of interaction, although how these materials arrived at the coast remain unknown.
Previous Dietary Evidence
Well-preserved archaeological assemblages offer copious, albeit indirect, evidence of ancient diet in these early villages. Middens at Paloma (Reitz Reference Reitz2003) and La Yerba III (Beresford-Jones et al. Reference Beresford-Jones, Pullen, Whaley, Moat, Chauca, Cadwallader, Arce, Orellana, Alarcón, Gorriti, Maita, Sturt, Dupeyron, Huaman, Lane and French2015, Reference Beresford-Jones, Pullen, Chauca, Cadwallader, García, Salvatierra, Whaley, Vásquez, Arce, Lane and French2018) are manifestly dominated by a wide spectrum of hunted and gathered marine resources. These include pelagic fish such as sea bass (Cilus gilberti) and smaller net-caught fish such as anchovy (Ergraulis ringens) and sardines (Sardinops sagax). Marine mammals, particularly sea lions (Otaria flavescens); edible kelps (Macrocystis spp.); and shellfish, such as various mussels (Mytilidae) and large Concholepas gastropods, were hunted and gathered from rocky littorals. Resources gathered from sandy beaches include crustaceans such as the mole crab (Emerita analoga) and surf clams (Mesodesma donacium). Proliferating in dense beds in shallow waters along sandy beaches and resilient to overexploitation, the latter offered the perfect protein resource for marine hunter-gatherers: they were easily collected without significant risk by all members of society and easily opened in quantities over small fires. Estuarine resources include summer-spawning mullet (Mugil cephalus), crayfish (Cryphiops caementarius), and diverse aquatic birds and eggs (Benfer Reference Benfer and Pearsall2008; Beresford-Jones et al. Reference Beresford-Jones, Pullen, Whaley, Moat, Chauca, Cadwallader, Arce, Orellana, Alarcón, Gorriti, Maita, Sturt, Dupeyron, Huaman, Lane and French2015, Reference Beresford-Jones, Pullen, Chauca, Cadwallader, García, Salvatierra, Whaley, Vásquez, Arce, Lane and French2018; Reitz Reference Reitz2003; Weir and Derring Reference Weir, Phillip Derring, Matos-Mendieta, Turpin and Eling1986).
Habitation sites on the north coast of Chile, broadly contemporaneous with the burials at Morro I, are also dominated by marine resources, including mollusks, crustaceans, fish, birds, and marine mammals (Bearez et al. Reference Bearez, Fuentes-Mucherl, Rebolledo, Salazar and Olguin2016; Salazar et al. Reference Salazar, Camila Arenas, Laura Olguin, Carola Flores, Sandra Rebolledo, Consuelo Sandoval, Ayelen Delgado and Robles2018; Santoro et al. Reference Santoro, Gayo, Carter, Standen, Castro, Valenzuela, De Pol-Holz, Marquet and Latorre2017; Standen et al. Reference Standen, Santoro, Arriaza and Colleman2017). Middens at sites such as Caleta Vitor suggest a long-term reliance on relatively few species of easily seen and collected mollusks and echinoderms, including urchins (Loxechinus sp.), chitons (Acanthopleura echinata), limpets (Fissurella sp.), abalones (Concholepas sp.), and mussels (Santoro et al. Reference Santoro, Gayo, Carter, Standen, Castro, Valenzuela, De Pol-Holz, Marquet and Latorre2017).
Yet, even though marine resource exploitation is certainly highly visible in the Middle Preceramic middens along this Pacific coast, most, if not all, of these middens also include significant evidence of terrestrial resources, hunted and gathered according to seasonal rounds from adjacent riparian and lomas fog oasis habitats. These resources include guanaco (Lama guanicoe) and deer (Odocoileus virginianus) at Paloma (Reitz Reference Reitz2003:78) and La Yerba III (Beresford-Jones et al. Reference Beresford-Jones, Pullen, Whaley, Moat, Chauca, Cadwallader, Arce, Orellana, Alarcón, Gorriti, Maita, Sturt, Dupeyron, Huaman, Lane and French2015) and vizcachas (Lagidium viscacia), guinea pigs (Cavia porcellus), and dogs (Canis familiaris) at La Yerba. Land snails (Bostryx sp.) proliferate in winter lomas and comprise 14% of the total mollusk assemblage at La Yerba III (Beresford-Jones et al. Reference Beresford-Jones, Pullen, Whaley, Moat, Chauca, Cadwallader, Arce, Orellana, Alarcón, Gorriti, Maita, Sturt, Dupeyron, Huaman, Lane and French2015) and are found in smaller proportions at Paloma (Reitz Reference Reitz2003).
Gathered and, increasingly, cultivated plant foods are also evident. Wild roots and tubers such as the rhizomes of Cyperaceae esculentus provided sources of starch at Paloma (Benfer Reference Benfer and Pearsall2008; Weir and Derring Reference Weir, Phillip Derring, Matos-Mendieta, Turpin and Eling1986), La Yerba III (Beresford-Jones et al. Reference Beresford-Jones, Pullen, Whaley, Moat, Chauca, Cadwallader, Arce, Orellana, Alarcón, Gorriti, Maita, Sturt, Dupeyron, Huaman, Lane and French2015), Huaca Prieta (Bonavia et al. Reference Bonavia, Vásquez, Tham, Dillehay, Netherly, Benson and Dillehay2017), and sites on the north coast of Chile (Ugalde et al. Reference Ugalde, McRostie, Gayo, Garcia, Latorre and Santoro2021). Cultivated food plants at Paloma, Chilca I, and La Yerba III include the earliest directly dated lima beans (Phaseolus lunatus) at Chilca I (6485–6280 cal BP), La Yerba III (6270–5996 cal BP), and Paloma; jack beans (Canavalia ensiformis) at Chilca I and La Yerba III; and guava (Psidium guajava) at Paloma and La Yerba III (Beresford-Jones et al. Reference Beresford-Jones, Pullen, Chauca, Cadwallader, García, Salvatierra, Whaley, Vásquez, Arce, Lane and French2018; Kaplan and Lynch Reference Kaplan and Lynch1999; Weir and Derring Reference Weir, Phillip Derring, Matos-Mendieta, Turpin and Eling1986). At Huaca Prieta's Phase 3, corresponding to this period (Bonavia et al. Reference Bonavia, Vásquez, Tham, Dillehay, Netherly, Benson and Dillehay2017:368; Grobman et al. Reference Grobman, Bonavia, Dillehay, Piperno, Iriarte and Holst2012), and at Paredones, these and other food crops are reported, including avocado (Persea americana), chili peppers (Capsicum spp.), and maize (Zea mays). At sites along the north coast of Chile, achira (Canna sp.), sweet potato (Ipomoea batatas), manioc (Manihot esculenta), and perhaps maize are reported (Ugalde et al. Reference Ugalde, McRostie, Gayo, Garcia, Latorre and Santoro2021). By the end of the Middle Preceramic after 5300 cal BP, subsistence at Pernil Alto, located some 50 km inland from La Yerba, was predominantly based on food crops (Gorbahn Reference Gorbahn2020), including beans (Phaseolus and Canavalia spp.), sweet potato, achira, and pumpkin (Cucurbita sp.), supplemented by gathered Prosopis pods and hunting. By the Late Preceramic, food cultivars proliferate at many sites along the coast of Peru (Pearsall Reference Pearsall, Silverman and Isbell2008).
Yet for all the richness of these data, quantified comparisons of the relative significance of these foodstuffs to the ancient diet are compromised by differential taphonomy, sampling, and analyses. Tiny schooling fish such as anchovy are underrepresented in most practical recovery methods, whereas durable shellfish remains may well be overrepresented. Plant foods are typically inconspicuous for various taphonomic reasons (Reinhard et al. Reference Reinhard, LeRoy-Toren and Arriaza2011; Weir and Derring Reference Weir, Phillip Derring, Matos-Mendieta, Turpin and Eling1986). So, whereas faunal remains suggest that fish dominated Paloman diets (Reitz Reference Reitz2003:71), coprolite analyses suggest that they were instead composed largely of shellfish, with significant plant components (Weir and Derring Reference Weir, Phillip Derring, Matos-Mendieta, Turpin and Eling1986:39). Coprolite analyses of burials at Morro I (Reinhard et al. Reference Reinhard, LeRoy-Toren and Arriaza2011:Table 5) and quantified archaeobotanical analyses of many sites along the north coast of Chile (Ugalde et al. Reference Ugalde, McRostie, Gayo, Garcia, Latorre and Santoro2021: Figure 3) also suggest more significant plant components to diets than hitherto widely recognized. Indeed, pace Moseley's (Reference Moseley1975) MFAC hypothesis, archaeological orthodoxy now ascribes earlier and greater significance to cultivated food plants in coastal Preceramic diets (Prieto and Sandweiss Reference Prieto and Sandweiss2020), albeit often underpinned by microfossil data with their own limitations of context and dating (Mercader et al. Reference Mercader, Bundala, Collins, Copeland, Crowther, Henry, Itambu, Larter, Longo, Patalano, Sammynaiken, Tyler and Xhauflair2018). For all these reasons then, questions remain about the trajectory of subsistence change that underpinned the increases in population foreshadowing the emergence of Late Preceramic social complexity in the wider Andean region. Here we contribute some direct evidence of diet and lifestyle from the first sedentary villages of the Middle Preceramic, epitomized by Paloma.
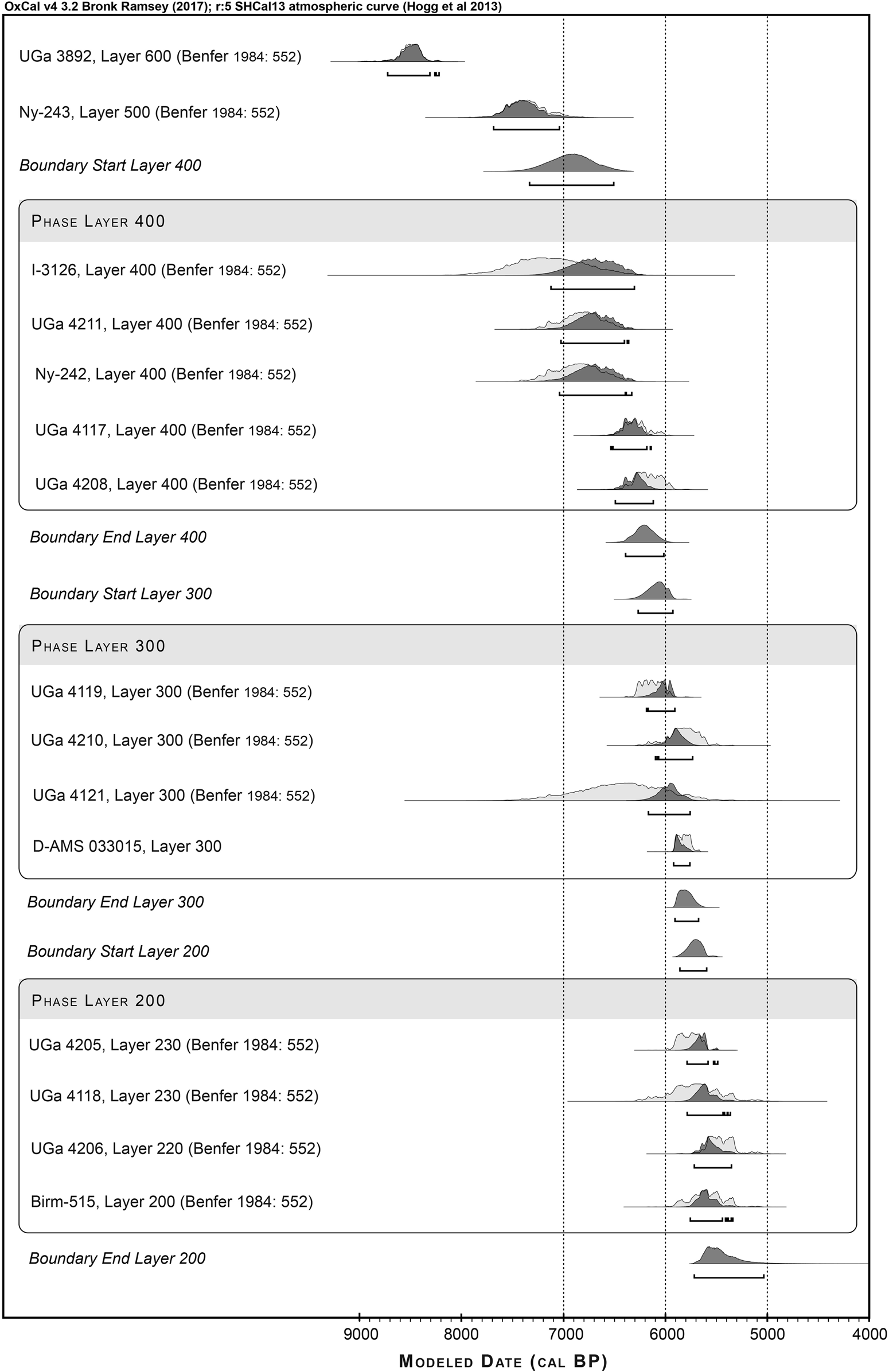
Figure 3. New and published radiocarbon dates for Paloma calibrated using ShCal13 (Hogg et al. Reference Hogg, Hua, Blackwell, Niu, Buck, Guilderson, Heaton, Palmer, Reimer, Reimer, Turney and Zimmerman2013) and then modeled using OxCal's Bayesian analysis following Bronk Ramsey (Reference Bronk Ramsey2009), in a sequence of phases, according to their reported stratigraphic sequence.
Scientific Background
Stable Isotope Analysis
Stable isotope analysis of human remains provides individual, semi-quantitative evidence for past diets because chemical differences in those diets are transmitted through the food chain and recorded in body tissues. Isotopic signatures record general consumption so that at least 20% of protein must derive from a particular isotopically distinct source for it to be detectable (Hedges Reference Hedges2003).
Stable carbon isotope ratios (δ13C) can distinguish between marine and terrestrial food chains because each have carbon sources with different isotope values (Lee-Thorp Reference Lee-Thorp2008). δ13C values can also distinguish between diets based on plants following different C3 and C4 photosynthetic pathways (O'Leary Reference O'Leary1988). Although most economically important plants are of the C3 type, some such as maize use the C4 photosynthetic pathway. Moreover, around one-third of wild edible plants on the south coast of Peru are C4, and these plants may also enter the human food chain after being consumed by animals that in turn are eaten by people (Cadwallader et al. Reference Cadwallader, Beresford-Jones, Whaley and O'Connell2012).
Stable nitrogen isotope ratios (δ15N) provide an indication of the amount of animal protein in diets, because δ15N values increase by 3‰–5‰ with each step in a trophic chain (Bocherens and Drucker Reference Bocherens and Drucker2003; O'Connell et al. Reference O'Connell, Kneale, Tasevska and Kuhnle2012). Food chains in aquatic ecosystems tend to be longer than in terrestrial ecosystems, leading to relatively elevated δ15N values in the former (Lee-Thorp Reference Lee-Thorp2008). People consuming diets high in marine foods are therefore characterized by having both high δ13C and high δ15N values.
In this study, we analyzed bone and dentine collagen (protein). Bone collagen reflects diet over the majority of adult life so that small dietary inputs or changes in diet may not be detectable (Hedges et al. Reference Hedges, Clement, Thomas and O'Connell2007). Dentine collagen, meanwhile, reflects the diet over the time of tooth formation during childhood (Gage et al. Reference Gage, Francis and Triffitt1989). δ15N values reflect only the protein component of the diet, whereas carbon in bone collagen predominantly reflects dietary protein combined with carbon from other dietary fractions (Ambrose and Norr Reference Ambrose, Norr, Lambert and Grupe1993; Tieszen and Fagre Reference Tieszen, Fagre, Lambert and Grupe1993).
Human Osteology
Adult stature is frequently used as a cumulative marker of health and nutrition during growth in studies of health and standards of living in both ancient and modern populations (Bogin and Keep Reference Bogin and Keep1999; Steckel Reference Steckel2009; Tanner Reference Tanner1987). Osteological analyses suggest that in many parts of the world (Cohen and Armelagos Reference Cohen and Armelagos1984; Mummert et al. Reference Mummert, Emily Esche and Armelagos2011), including the Peruvian central coast (Pechenkina et al. Reference Pechenkina, Vradenburg, Benfer, Farnum, Cohen and Crane-Kramer2007), the initial transition to agriculture caused stature and health to decline, attributed in part to an increased disease burden caused by concomitant sedentism (Cohen and Armelagos Reference Cohen and Armelagos1984; Larsen Reference Larsen2006). Data from the north coast of Chile suggest relatively short stature and fairly poor health among the Chinchorro buried at Morro I and other cemeteries (Arriaza and Standen Reference Arriaza and Standen2006, Reference Arriaza and Standen2008; Pomeroy Reference Pomeroy2012) attributed to sedentism facilitated by rich marine resources rather than by agriculture, and therefore persisting after the adoption of agriculture. Previous studies on the Peruvian and Chilean Pacific coasts have, however, varied in their methods used to estimate stature, limiting the comparability of data between sites and regions. Here we estimate stature using a consistent methodology and apply equations specifically derived for South American Andean populations (Pomeroy and Stock Reference Pomeroy and Stock2012) to long bone lengths of adults from Paloma and Morro I.
Another common observation in osteological analyses of human remains is that the transition to agriculture in many parts of the world is accompanied by decreased levels of habitual mobility—presumably due to the much greater sedentism that accompanies agricultural lifestyles as compared with that of hunter-gatherers (Mummert et al. Reference Mummert, Emily Esche and Armelagos2011; Ruff et al. Reference Ruff, Larsen and Hayes1984). Cross-sectional geometry of the femoral and tibial midshafts provides direct skeletal evidence for habitual mobility levels because the amount and distribution of bone in the cross-section of shafts are influenced by loading patterns. Higher ratios of maximum to minimum bending rigidity (Imax/Imin) of the femur reflect higher habitual mobility levels (Ruff Reference Ruff, Katzenberg and Saunders2008). In this study, we also examined cross-sectional geometric properties of the femoral midshaft to explore potential links between diet, mobility, and stature in skeletal samples from Paloma and Morro I. Examining patterns of habitual mobility may help elucidate whether differences between two main study regions in stature and temporal trends in height may relate to any contrasts in behavior, diet, or both.
Materials and Methods
We calibrated previously published (Benfer Reference Benfer, Cohen and Armelagos1984; Beresford-Jones et al. Reference Beresford-Jones, Pullen, Chauca, Cadwallader, García, Salvatierra, Whaley, Vásquez, Arce, Lane and French2018; Engel Reference Engel1988; Kaplan and Lynch Reference Kaplan and Lynch1999; Standen Reference Standen1997, Reference Standen2003) and new (see Supplemental Table 1) radiocarbon dates for Paloma, Chilca I, La Yerba III, and Morro I using ShCal13 (Hogg et al. Reference Hogg, Hua, Blackwell, Niu, Buck, Guilderson, Heaton, Palmer, Reimer, Reimer, Turney and Zimmerman2013). Twenty-one radiocarbon dates for Paloma were modeled using OxCal v4.3's Bayesian analysis (following Bronk Ramsey Reference Bronk Ramsey2009) as a sequence of phases to define boundary limits for the site's published stratigraphy (Supplemental Table 2). Twenty-five radiocarbon dates for Paloma's Layers 300 and 400, Chilca I, La Yerba III, and the earliest tomb contexts at Morro I were then modeled using OxCal as a single, contiguous phase to test the hypothesis that components of these sites were contemporaneous (Supplemental Table 3).
Collagen was extracted (following Privat et al. Reference Privat, O'Connell and Richards2002) from human bone samples from 29 Palomans, three bone and two dentine samples from three individuals from La Yerba III, and one bone sample from Chilca I. Reliable yields for stable isotopic data were obtained from eight of those from Paloma, two of those from La Yerba III, and the individual from Chilca I (Table 1; Supplemental Text 1).
Table 1. Results of Stable Isotope Analyses of Individuals from Paloma, Chilca I, and La Yerba III.

*Sex and age at death data from Quilter (Reference Quilter1989).
Osteological analyses of stature (following Pomeroy and Stock Reference Pomeroy and Stock2012) and femoral midshaft biomechanics (Imax/Imin) were carried out on human bone samples from 62 individuals from Paloma and 28 from Morro I (Table 2; Supplemental Table 4; see Supplemental Text 1 for full details of all methods, laboratory procedures, and statistical analyses).
Table 2. Comparison of Mean Estimated Adult Statures and Osteological Indicators of Habitual Mobility for Individuals from Paloma and Morro 1.

* Statistically significant values (p < 0.05) are shown in bold.
Results
Chronology
Reliable collagen extractions from Palomans all derive from Layer 300, with the exceptions of one individual from Layer 400 (Burial 84, PAL 10) and one from Layer 100 surface contexts (Burial 2, PAL 01), directly dated to 1182–981 cal BP and therefore not considered further here (Supplemental Tables 1 and 2). OxCal Bayesian modeling of Paloma's radiocarbon dates as a sequence of phases gives median boundary limits of between 6077 and 5802 cal BP for Layer 300 and between 6914 and 6204 cal BP for Layer 400 (Figure 3; Supplemental Table 2). Because of her unusual data and interment, which are discussed later, Burial 110 (PAL 13) was dated directly (D-AMS 033015; Supplemental Tables 1 and 3) to 5913–5725 cal BP and thereby confirmed to be contemporaneous with Layer 300.
Of the 28 Morro I burials analyzed osteologically here, five were directly dated (Figure 2), whereas others can be dated by association with varying degrees of precision (Supplemental Table 5). Seven likely date to Phase 1, 6252–5690 cal BP, which is approximately contemporary with the Middle Preceramic Peruvian villages (Standen Reference Standen2003). Nineteen date to Phases 2 and 3 and postdate the Peruvian villages by one to two millennia. Two remaining can only be broadly assigned to the Preceramic. Four Morro I burials previously analyzed for stable isotope ratios (King et al. Reference King, Millard, Gröcke, Standen, Arriaza and Halcrow2018) likely date to Phase 1, contemporary with the Peruvian villages, whereas a fifth is likely dated to Phase 2, 5185–4415 cal BP (see Supplemental Text 1 for further discussion).
OxCal Bayesian modeling of 25 radiocarbon dates for Paloma's Layers 300 and 400, Chilca I, La Yerba III, and Morro I's T10b tomb context as a single, contiguous phase tests the hypothesis that these specific contexts of these sites were contemporaneous (Supplemental Table 3). Unsurprisingly, a date from Chilca I's surface Layer 600 and four dates from Paloma's Layer 400, each with previously identified contextual uncertainties or very large standard errors (Benfer Reference Benfer, Cohen and Armelagos1984:552), show poor agreement indices (<60) with such a model. Setting these as outliers, however, produces an overall agreement index of 97, supporting the hypothesis that components of these four sites are contemporaneous between 6372 and 5848 cal BP (single-phase boundary median start and end dates, respectively, at 95.4% confidence limit; Figure 4).

Figure 4. Paloma, Chilca I, La Yerba III, and Morro I radiocarbon dates calibrated using ShCal13 (Hogg et al. Reference Hogg, Hua, Blackwell, Niu, Buck, Guilderson, Heaton, Palmer, Reimer, Reimer, Turney and Zimmerman2013) and then modeled as a single phase using OxCal's Bayesian analysis following Bronk Ramsey (Reference Bronk Ramsey2009).
Stable Isotope Analyses
Stable isotope results are shown in Table 1 and Figure 5. Bone collagen data from Palomans have a mean δ13C value of −13.9‰ (±1.9‰, range −17.6 to −11.9‰, n = 8) and a mean δ15N value of 15.8‰ (±2.6‰, 9.9 to 17.9‰, n = 8). One outlier is identified: Burial 110 (PAL13, female) has a δ15N value of 9.9‰ (lying more than 1.5 times the interquartile range below quartile 1). This individual also has the lowest δ13C value (−17.6‰), although this is not identified as a statistical outlier. All reliable collagen extractions from Paloma came from Layer 300, with the exception of one from Layer 400 (Burial 84, PAL10), whose isotopic values are similar to other Palomans.

Figure 5. Stable isotope δ13C and δ15N values for individuals from Paloma, Chilca I, and La Yerba III (see Table 1) compared with other published stable isotope values from the Preceramic period.
Bone collagen from the single individual from Chilca I (PAL19) has a δ13C value of −13.3‰ and a δ15N value of 16.8‰, whereas the two dentine collagen results from La Yerba III have δ13C values of −15.1‰ and −14.1‰, and δ15N values of 15.3 and 16.8‰. Although these sample sizes are too small for statistical comparison, they clearly fall within the range of typical isotopic variation in Layer 300 of Paloma.
Human Osteology
Osteological results are shown in Table 2 and Supplemental Figure 2. No significant differences in stature or Imax/Imin were found between layers 200, 300, and 400 for either sex at Paloma using two-way ANOVA (see Supplemental Text 1 for further discussion; Supplemental Figure 2). Measurements of Palomans from all layers are therefore combined in comparisons with those at Morro I (see Supplemental Text 1 for further details). For both stature and femoral midshaft Imax/Imin, the two-way ANOVA indicated significant differences between sexes and sites (p < 0.05), although the interaction effect was not significant (p < 0.05; Table 2). These results show that males were significantly taller than females in both samples and that Palomans were significantly taller than those from Morro I. Paloman adults show significantly higher femoral midshaft Imax/Imin ratios and, therefore, significantly higher levels of habitual mobility than those from Morro 1 (Table 2).
Discussion
The isotopic and osteological data presented here for Paloma, Chilca I, La Yerba III, and Morro I come from a limited number of individuals but contribute to a larger body of gradually emerging direct evidence for diet and lifestyle in the first villages that proliferated along this Pacific coast around 6000 cal BP toward the end of the Middle Preceramic period: changes that foreshadowed the emergence of sedentary complex society in the wider Andean region. Calibration and Bayesian modeling of their new and existing radiocarbon dates support the hypothesis that components of these sites were contemporaneous between 6372 and 5848 cal BP (Figure 4; Supplemental Table 3), providing a refined chronological context for the data presented.
No faunal samples were available to evaluate background environmental isotopic variations, but it is nonetheless evident from the stable isotope data that almost all the individuals analyzed from Paloma, La Yerba III, Chilca I, and Morro I consumed diets high in marine protein. This evidence corroborates the long-standing presumption, based on multiple lines of archaeological data (Beresford-Jones et al. Reference Beresford-Jones, Pullen, Chauca, Cadwallader, García, Salvatierra, Whaley, Vásquez, Arce, Lane and French2018; Moseley Reference Moseley1975; Prieto and Sandweiss Reference Prieto and Sandweiss2020; Quilter Reference Quilter1991), of the critical role of rich marine resources in promoting sedentism and increasing population and social complexity during the Middle Preceramic along this Pacific coast. As discussed, however, quantifying those other lines of indirect data against one another to determine their relative significance to human diet is complicated by uncertainties of taphonomy and sampling: plant foods, in particular, are likely underrepresented for a variety of taphonomic reasons. It is therefore notable that the direct evidence provided by the isotopic values of the inhabitants of these villages continues to assert the generalized primacy of marine protein sources; this is despite the considerable evidence at these sites for hunted and gathered terrestrial foods and, indeed, for incipient agriculture of, for instance, fully domesticated, high-protein beans and, at La Yerba III, possibly domesticated guinea pigs (Beresford-Jones et al. Reference Beresford-Jones, Pullen, Whaley, Moat, Chauca, Cadwallader, Arce, Orellana, Alarcón, Gorriti, Maita, Sturt, Dupeyron, Huaman, Lane and French2015, Reference Beresford-Jones, Pullen, Chauca, Cadwallader, García, Salvatierra, Whaley, Vásquez, Arce, Lane and French2018; Engel Reference Engel1991; Kaplan and Lynch Reference Kaplan and Lynch1999; Weir and Derring Reference Weir, Phillip Derring, Matos-Mendieta, Turpin and Eling1986).
Burial 110 (PAL13), an adult female around 20 years old at death, stands as an exception to this general pattern of significant marine protein consumption at Paloma. Although directly dated as contemporary with other Layer 300 interments, Burial 110's isotopic data suggest a diet comprising little, if any, marine protein (Table 1; Figure 5). She is exceptional too in her unusual interment. Whereas most people at Paloma (and indeed also at Chilca I and La Yerba III) were buried below the floor of a house, flexed, with knees drawn toward the chest and wrapped in twined reed matting, Burial 110 was interred in an unlined grave, beyond the main area of excavated house structures in a “hearth/trash dump” (Quilter Reference Quilter1989:123; Supplemental Figure 1).
Figure 5 compares the isotopic data from these Middle Preceramic coastal villages with other published data for the wider Preceramic period. Burial 110's isotopic values are most similar to those of the somewhat later, inland sites (Figure 1) of La Galgada, 80 km from the sea at 1,100 m asl (Washburn et al. Reference Washburn, Nesbitt, Burger, Tomasto-Cagigao, Oelze and Fehren-Schmitz2020), and of Kasapata, far inland at 3,400 m asl in the Cuzco region (Turner et al. Reference Turner, Bélisle, Davis, Skidmore, Juengst, Schaefer, Alan Covey and Bauer2018). Her isotopic values, akin to inland terrestrial diets, and the unusual aspects of her burial suggest that Burial 110 was a migrant to the coast. While the mechanisms of trade and exchange by which obsidian lithics arrived on the coast from their highland source at Quispisisa remain unknown, the burial of a highland migrant at Paloma certainly hints at direct access interactions with the highlands at this time.
Even excluding the Burial 110 outlier, the isotopic data also show variations in the marine protein consumed by individuals in these Middle Preceramic villages. Interpreting these variations is complicated by pelagic-benthic coupling in the marine ecosystem along this coast driven by deep upwelling. Pelagic carbon sources typically have significantly lower δ13C values than benthic sources (France Reference France1995), so that when pelagic-derived materials such as phytoplankton are transported by upwelling to be consumed by benthic primary consumers and thereby transferred to higher trophic levels in the food web, they in turn yield complex isotopic patterns in their own consumers. Filter-feeding bivalves from northern Chile, for instance, have carbon isotope values that are on average 4.2‰ lower than those of grazing gastropods collected from the same location, while their nitrogen isotope values are broadly similar (Docmac et al. Reference Docmac, Araya, Hinojosa, Dorador and Harrod2017). This is consistent with those bivalves consuming pelagic-derived phytoplankton, whereas the gastropods consume benthic macroalgae. Benthivorous fish from these locations have carbon isotope values showing diets dominated by pelagic-derived material (and nitrogen isotope values reflective of their higher trophic level than the primary consumers). So, although the isotopic data show differences between, on the one hand, the individuals from La Yerba III and two of the Palomans (Burial 39, PAL06, and Burial 212, PAL21, both adult females) and, on the other hand, the rest of the individuals from Paloma and Chilca I (Figure 5), which may suggest that the former consumed less marine protein than the latter, these data might also reflect a relatively greater consumption of pelagic or pelagic-consuming marine resources. Indeed, this interpretation is suggested by the significant proportion of mollusk remains at La Yerba III that are bivalves (35% by MNI and 42% by weight), in particular Mesodesma surf clams; whereas Mesodesma is apparently absent in Paloma's Layer 400 and then only gradually becomes more common in overlying Layers 300 and 200 (Reitz Reference Reitz2003:71).
Figure 5 also shows comparative isotopic data from Morro I (King et al. Reference King, Millard, Gröcke, Standen, Arriaza and Halcrow2018). Of 18 individuals analyzed, 5 yielded sufficient collagen for reliable isotopic values to be determined. Compared to those of their Middle Preceramic contemporaries at Paloma, Chilca I, and La Yerba III, the isotopic values of four Morro I individuals (T4, T6, T10a, and T12) show higher carbon but generally lower nitrogen isotopic values (Figure 5). This suggests that they consumed greater proportions of marine resources from lower trophic levels, such as shellfish rather than fish, and benthic gastropods rather than pelagic bivalves. The fifth individual (T23C13), meanwhile, likely dates up to two millennia later (Supplemental Table 5) and has isotopic values broadly comparable to those of the Peruvian villages, hinting at greater proportions of marine foods from higher trophic levels over time on the north coast of Chile.
These comparisons seem to be reflected in the wider archaeological record. Even though the archaeological assemblages of Paloma (Benfer Reference Benfer and Pearsall2008; Reitz Reference Reitz2003) and La Yerba III (Beresford-Jones et al. Reference Beresford-Jones, Pullen, Chauca, Cadwallader, García, Salvatierra, Whaley, Vásquez, Arce, Lane and French2018) contain a broader range of fish, mollusk, and crustaceans compared with, for instance, Caleta Vitor on the north coast of Chile, the latter becomes more diverse over time (Santoro et al. Reference Santoro, Gayo, Carter, Standen, Castro, Valenzuela, De Pol-Holz, Marquet and Latorre2017). Meanwhile, sophisticated fishing technologies are evident at Paloma and La Yerba III in the form of gorges, cactus spine fishhooks, fragments of possible bottle gourd floats for beach seine nets, copious forms of plant fiber cordage, lines, knotted net fragments, and net-making needles (Beresford-Jones et al. Reference Beresford-Jones, Pullen, Chauca, Cadwallader, García, Salvatierra, Whaley, Vásquez, Arce, Lane and French2018). By contrast, in the earlier Chinchorro burials at Morro I contemporaneous with the Middle Preceramic Peruvian villages, there are no nets and limited fishing paraphernalia, which mostly comprise looped fiber bags similar to the “chinguillos” bags used today for collecting mollusks.
The osteological data presented here, meanwhile, show that males and females from Paloma were, on average, taller than those from Morro 1 (Table 2; Supplemental Figure 2). As discussed, relatively poor health among the Chinchorro has been attributed to early sedentism enabled by rich marine productivity on the north coast of Chile, rather than by plant or animal domestication: an hypothesis that would also account for the lack of change in stature after agriculture was adopted (Arriaza and Standen Reference Arriaza and Standen2008; Supplemental Table 6; Supplemental Figure 2). Palomans, however, seem to have paid little health penalty for their settled lifestyles, as reflected in taller mean stature and lower indicators of nonspecific “stress” compared with their agricultural successors on the central Peruvian coast right through to AD 1000 (Pechenkina et al. Reference Pechenkina, Vradenburg, Benfer, Farnum, Cohen and Crane-Kramer2007). This suggests that sedentism per se does not account for the differences in stature between Paloma and Morro 1, but rather that these variations may be explained by different patterns of marine resource exploitation, reflected in their isotopic and archaeological records and, indeed, analyses of skeletal biomechanics.
Osteological data in many parts of the world confirm other diverse lines of evidence that hunter-gatherers are generally more highly mobile than agriculturalists (Mummert et al. Reference Mummert, Emily Esche and Armelagos2011). Yet femoral cross-sectional geometry data from Morro 1 suggest that these individuals were no more habitually mobile than their agricultural successors in the Azapa Valley in northern Chile (Pomeroy Reference Pomeroy2012; Supplemental Text 1; Supplemental Table 6; Supplemental Figure 2), as reflected also in 87Sr/86Sr ratios comparable with marine mammals (Standen et al. Reference Standen, Santoro, Arriaza and Colleman2017). Cross-sectional geometry of the femur in both males and females at Paloma suggests greater habitual mobility than their counterparts at Morro I.
Previously, longue durée ENSO conditions (Carré et al. Reference Carré, Sachs, Purca, Schauer, Braconnot, Falcon, Julien and Lavallée2014) underpinning environmental abundance and seasonal predictability have been proposed as enabling Middle Preceramic (fisher) hunter-gatherers to reduce their mobility and settle at locations where different habitats conjoined, such as those of Paloma, Chilca I, and La Yerba III (Beresford-Jones et al. Reference Beresford-Jones, Pullen, Whaley, Moat, Chauca, Cadwallader, Arce, Orellana, Alarcón, Gorriti, Maita, Sturt, Dupeyron, Huaman, Lane and French2015). The archaeological records of these first permanent villages include resources gathered from those habitats according to rounds of logistical mobility (sensu Binford Reference Binford1980): ephemeral lomas fog oases, in-shore marine pelagic and benthic habitats (including estuarine, sandy beach and rocky headland); and riverine oases (Beresford-Jones et al. Reference Beresford-Jones, Pullen, Whaley, Moat, Chauca, Cadwallader, Arce, Orellana, Alarcón, Gorriti, Maita, Sturt, Dupeyron, Huaman, Lane and French2015; Engel Reference Engel1988; Weir and Derring Reference Weir, Phillip Derring, Matos-Mendieta, Turpin and Eling1986). For instance, 85% of guanaco bones recorded at La Yerba III are of meatier hind leg parts, suggesting that they were butchered at distant kill sites for transport and consumption in the village. Before riverine oases were altered entirely by agriculture, guanaco likely migrated between winter lomas and summer Andean footslopes, thereby shaping rounds of hunter transhumance (Custred Reference Custred1979). Such patterns of seasonal logistical mobility are reflected in Palomans’ osteological data.
While all people along this Pacific littoral relied on marine resources during the Middle Preceramic, they exploited the sea in different ways. Whereas the inhabitants of the first Peruvian villages employed diverse technologies to harvest a broad range of marine and terrestrial resources in seasonal rounds of logistical mobility, those on the north coast of Chile at this time consumed a more limited range of marine resources, which constituted an even more significant protein component of their diet and were gathered according to less mobile lifestyles.
Finally, even though comparative isotopic data for the long Preceramic are still limited, that data seem to reflect general trends resonant with the wider archaeological record, including changes in the mix between marine and terrestrial resources, different kinds of marine resources, or both (Figure 5). Isotopic data from five individuals from Huaca Prieta's Middle Preceramic Phase 3 (6538–5308 cal BP) and eight from its Late Preceramic Phase 4 (5308–4107 cal BP) in Chicama (DeSantis et al. Reference DeSantis, Dillehay, Goodbred, Feranec and Dillehay2017), alongside eight individuals from the Late Preceramic (4950–3650 cal BP) site of Bandurria, Norte Chico (Coutts et al. Reference Coutts, Chu and Krigbaum2011), all show lower δ13C and δ15N values than those of Paloma, La Yerba III, Chilca I, and Morro I's older contexts, suggesting that they consumed lower proportions of marine protein, or greater proportions of pelagic or pelagic-consuming marine resources. These changes in the direct signatures of diet likely reflect not only the increasing significance of cultivated food crops into the Late Preceramic (Dillehay Reference Dillehay2017; Pearsall Reference Pearsall, Silverman and Isbell2008) but also the increasing sophistication of fishing technologies. Larger seine nets made from cultivated cotton, for instance, enabled the harvest of much larger quantities of schooling fish such as anchovy (Beresford-Jones et al. Reference Beresford-Jones, Pullen, Chauca, Cadwallader, García, Salvatierra, Whaley, Vásquez, Arce, Lane and French2018; Bird et al. Reference Bird, Hyslop and Skinner1985:225; Moseley Reference Moseley1975), significantly altering the benthic/pelagic balance of marine resources consumed. By the end of this chronological sequence a few individuals from Huaca Prieta's Phase 4 have isotopic values that almost converge with those of individuals with terrestrial diets from the Late Preceramic site of La Galgada, 80 km inland. Aside from individual tooth samples from Paredones in unspecified chronostratigraphic contexts (Figure 5; DeSantis et al. Reference DeSantis, Dillehay, Goodbred, Feranec and Dillehay2017; Tung et al. Reference Tung, Dillehay, Feranec and DeSantis2020), and pace the suggestions of Haas and colleagues (Reference Haas, Creamer, Mesía, Goldstein, Reinhard and Rodríguez2013) and others, there is little evidence in any of these isotopic data for significant maize consumption at these time depths, either on the coast or in the highlands (cf. Washburn et al. Reference Washburn, Nesbitt, Burger, Tomasto-Cagigao, Oelze and Fehren-Schmitz2020).
Conclusions
Isotopic and osteological data presented here from the earliest sedentary villages along the Pacific coast of South America at Paloma, Chilca I, La Yerba III, and Morro I contribute to a gradually emerging picture of changing diet and lifestyle toward the end of the Middle Preceramic around 6000 cal BP. Aridity in this area has preserved a remarkable archaeological record, but quantifying the relative significance of varying ancient diet remains will inevitably be compromised by differential taphonomy, sampling, and analyses. Isotopic data from bone collagen and dentine, by contrast, yield direct evidence of ancient diet—but only reflecting generalized consumption over many years and dominated by protein components, so that all these data are best interpreted through synthesis.
The data presented corroborate the long-standing connection between marine productivity and sedentism along this coast. δ13C and δ15N isotopic values affirm the generalized primacy of marine dietary protein sources in the diets of the inhabitants of these villages, despite considerable archaeological evidence for hunted and gathered terrestrial foods and incipient agriculture. Yet, since the δ13C and δ15N values of collagen reflect mostly and entirely, respectively, the protein component of diet, isotopic data in isolation inadequately reflect low-protein dietary components, not least plants, whose significance is evidenced directly in coprolites and also in copious midden remains. Moreover, human nutrition requires the high nitrogen levels of protein-rich foods to be diluted by the consumption of either carbohydrates or fats (Speth and Spielmann Reference Speth and Spielmann1983). Midden remains at Paloma and La Yerba III suggest that this requirement was satisfied by fat in hunted marine mammals and by plant foods in the form of gathered tubers and rhizomes and, increasingly, cultivated plants including high-protein beans.
Although this marine isotopic signature is consistent with the sedentism manifest in the permanent architecture, structured burials, and incipient plant cultivation at these early villages, more subtle implications for mobility may also be discerned. A young woman interred in unusual circumstances at Paloma had apparently come from the Andean hinterlands, hinting that these coastal villages obtained highland resources by direct interaction; for instance, the obsidian procured from Quispisisa, which was 300 km away. Palomans have more marked osteological indicators of habitual mobility than those buried at Morro I and those differences are higher among males, reflecting the rounds of logistical mobility followed by some members of society that are evident in their wider archaeological records. By locating themselves at the confluence of habitats, the inhabitants of these early villages were able to broaden their use of resources and increase their residential settlement. Yet they paid little substantial health penalty for their settled lifestyles, as reflected by their taller mean stature and lower rates of nonspecific “stress” indicators compared with those of their agricultural successors, even more than five millennia later. Contrasting isotopic and osteological patterns in their contemporaries on the north coast of Chile seem to reflect exploitation of a narrower range of marine resources, using different plant fiber fishing technologies and entailing less habitual mobility, in turn resulting in shorter stature and poorer health.
These data contribute to an emerging picture for how increasingly permanent settlement, population density, and social complexity on the Pacific coast of South America laid the foundations for the emergence of Late Preceramic civilization in the wider Andean region. That evidence, now including isotopic data over the longue durée of the Preceramic, suggests that whereas marine protein was always important to coastal diets, through time it was consumed in lower proportions, or included greater proportions of pelagic as opposed to benthic marine resources, each change consistent, respectively, with the increasing cultivation of food crops and of cotton for fishing nets that is manifest in the wider archaeological record of the Late Preceramic.
Acknowledgments
We wish to thank three anonymous reviewers of this article; Catherine Kneale and James Rolfe (University of Cambridge) for their help with mass spectrometry; Vivien Standen, Marietta Ortega, Héctor González, Bernardo Arriaza, and Susanna Monsalve (Museo Arqueológico San Miguel de Azapa, Departamento de Antropología, Universidad de Tarapacá, Chile) and Professor Alex Robling (Indiana University School of Medicine) for their help with osteological sampling and analyses; and the Ministerio de Cultural del Perú (ex-Instituto Nacional de Cultura) for granting permission for fieldwork and the analysis of samples. The work of David G. Beresford-Jones was supported by the Leverhulme Trust (grant number RPG-117) and the late Don Alberto Benavides de la Quintana (grant number RG69428). The work of Emma Lightfoot was supported by the TwoRains project, which was funded by the European Research Council (grant agreement number 648609).
Data Availability Statement
Materials from Paloma and Chilca I are curated at the Museo Nacional de Antropología, Biodiversidad, Agricultura y Alimentación (ex-Centro de Investigaciones de Zonas Áridas), Universidad Nacional Agraria La Molina, Lima, Peru; materials from La Yerba III are curated at the Museo Regional de Ica, Ica, Peru, and from Morro 1 at the Museo Arqueológico San Miguel de Azapa, Departamento de Antropología, Universidad de Tarapacá, Chile. Raw data underlying this article are available at the University of Cambridge's Apollo Repository at https://doi.org/10.17863/CAM.63764.
Supplemental Material
For supplemental material accompanying this article, visit https://doi.org/10.1017/laq.2021.24.
Supplemental Text 1. Methods and Materials.
Supplemental Table 1. New Radiocarbon Dates for Paloma.
Supplemental Table 2. Paloma Radiocarbon Dates Modeled as Phases in Sequence.
Supplemental Table 3. Paloma, Chilca I, La Yerba III, and Morro I Radiocarbon Dates Modeled as a Single Phase.
Supplemental Table 4. Breakdown of the Osteological Sample.
Supplemental Table 5. Chronology of Morro I Burials.
Supplemental Table 6. Comparison of Mean Estimated Adult Statures and Indicators of Habitual Mobility for Morro 1 Hunter-Gatherers and Azapa Valley Agriculturalists.
Supplemental Figure 1. Burials at La Yerba III and Paloma.
Supplemental Figure 2. Box-and-whisker plots of (A) stature and (B) femoral midshaft Imax/Imin ratios across different levels at Paloma and at Morro 1; and (C) stature and (D) femoral midshaft Imax/Imin ratios for Morro 1 hunter-gatherers and Azapa Valley agriculturalists.










 Open access
Open access