


A number of studies dating back to the early years of the 20th century have examined the role of genetic factors in Criminal behavior (CB) and found heritable influences (Christiansen, Reference Christiansen and Hood1974; Dalgard & Kringlen, Reference Dalgard and Kringlen1976; Lange, Reference Lange1929; Rosanoff et al., Reference Rosanoff, Handy and Plesset1934). A broader literature, including self-report measures of antisocial behavior, has been summarized in two meta-analyses (Ferguson, Reference Ferguson2010; Rhee & Waldman, Reference Rhee and Waldman2002), which concur in showing strong effects of genetic factors (estimates of heritability of 41% and 56% respectively) and significant but less prominent shared environmental influences accounting for 16% (Rhee & Waldman, Reference Rhee and Waldman2002) and 11% (Ferguson, Reference Ferguson2010) of the variance in liability.
During this same time period, intense interest has been focused on developmental theories of crime (Loeber & LeBlanc, Reference Loeber, LeBlanc, Tonry and Morris1990; Moffitt, Reference Moffitt1993; Piquero, Reference Piquero and Liberman2008). One of the most prominent of these theories has been articulated by Moffitt and colleagues (Kark et al., Reference Kark, Shemi, Friedlander, Martin, Manor and Blondheim1996), who argue for two distinct types of offenders: adolescent-limited and life-course persistent. These two groups are distinguished by their level and pattern of offending over time, as well as by the causes and predictors of their offending. The life-course persistent offenders initiate antisocial and CB early in life and persist in offending into adulthood. The adolescent-limited group of offenders, by contrast, is characterized by frequent offending in adolescence and a decline in offending in adulthood. Adolescent-limited offenders, thus, are a less serious, chronic group of offenders as compared to the life-course persistent offenders.
There have, however, been surprisingly few studies that have attempted to bridge the gap between the static twin and adoption studies of CB –– nearly all of which have examined one measure of lifetime prevalence of CB –– and the temporally dynamic developmental theories of criminality. A few studies have applied longitudinal twin models to self-report measures of externalizing disorders to twin samples, but these do not extend past young adulthood (Blonigen et al., Reference Blonigen, Hicks, Krueger, Patrick and Iacono2006; Hicks et al., Reference Hicks, Blonigen, Kramer, Krueger, Patrick, Iacono and McGue2007; Wichers et al., Reference Wichers, Gardner, Maes, Lichtenstein, Larsson and Kendler2013). One of these reports (Wichers et al., Reference Wichers, Gardner, Maes, Lichtenstein, Larsson and Kendler2013) well illustrates the two major kinds of changes that might be seen in genetic and environmental risk factors over development: (1) innovation –– new genetic or environmental factors ‘coming on line’ in later developmental periods, and (2) attenuation – a decline, in subsequent developmental periods, of the impact of genetic or environmental factors.
In this report, we seek to fill this gap in the literature by applying a longitudinal model to registry-based criminal convictions in Swedish male–male twins and sibling pairs. We examine three age periods (15–19, 20–24, and 25–29) over which we had sufficient number of subjects with CB. We seek, in these analyses, to address the following three questions:
-
1. is there evidence for genetic or shared environmental innovation or attenuation for CB over this 15-year period? In particular, we were interested in testing Moffitt's theory (Kark et al., Reference Kark, Shemi, Friedlander, Martin, Manor and Blondheim1996) that predicts the existence of one set of genetic influences that are operative in adolescence and another which ‘comes on line’ during early adulthood.
-
2. does the heritability of CB change over age? An increase might be expected if active gene-environment correlation is occurring over this time period so that individuals at high genetic risk seek out high-risk environments, which in turn feed back to increase their rates of offending (Caspi et al., Reference Caspi, Elder and Bem1987).
-
3. do we see changes in the impact of shared environmental influences on CB? We might expect a decline as individuals leave their home of rearing and sometimes move to different communities.
Methods
Sample and Measures
We linked nationwide Swedish registers via the unique 10-digit identification number assigned at birth or immigration to all Swedish residents. The identification number was replaced by a serial number to ensure anonymity.
The crime register covers all convictions in lower court from 1973 to 2011 and the following criminal conviction types were used to define CB, law and chapter in parentheses: (aggravated) assault (3:5, 3:6); illegal threat (4:5); threats and violence against an officer (17:1, 17:2); intimidation (4:7); [Gross] violation of a person’s/woman's integrity (4:4a); kidnapping (4:1); illegal confinement or restraint (4:2); (aggravated) robbery (8:5, 8:6); illegal coercion (4:4); (aggravated) arson (13:1, 13:2); murder, manslaughter or filicide (3:1, 3:2, 3:3); sexual crimes (excluding prostitution and the buying of sexual services but including child pornography; 6:1–6:10, 6:12, 16:10A); theft of a vehicle (8:1–2, 8:4, 8:7–8); theft (including burglary; 8:1–2, 8:4); vandalism (12:1–4); vandalism causing danger to the public, sabotage, hijacking (13:3–10; 5a-b); unlawful entering of a person's home, trespassing (4:6); fraud (9:1–10); embezzlement (10:1–8; 5a-e); dishonesty/crime towards a creditor (includes forged book-keeping in companies; 11:1–5); and forgery (14:1–10). Individuals’ CB were assessed in the three age periods –– 15–19, 20–24, and 25–29 –– and based on year of the crime if possible and if this was missing, year of the conviction.
Our measure of CB is from officially recorded data on criminal convictions and is therefore biased toward the most severe types of offending and does not reflect criminal offending that goes undetected by the criminal justice system. For the purposes of simplicity, we refer to criminal convictions as CB throughout the article.
We identified all twins and full-siblings born in Sweden within two years of each other. monozygotic (MZ) and dizygotic (DZ) twins were identified from the Swedish Twin Registry and full-siblings from the Multi-Generation Register. So as to be able to follow the occurrence of CB from ages 15 to 29, we included pairs where the oldest was born in 1958 and the youngest in 1976. We censored for death and emigration. The crime registry does not contain information about the start and end dates for incarceration. Consistent with the documented low incarceration rates and short prison terms in Sweden (Mauer, Reference Mauer1995, Reference Mauer2003; Rouse, Reference Rouse1985), only 5.4% of our sample with CB was subject to any incarceration. In less than 1% of our sample did this period exceed one year. Therefore, our inability to censor subjects during their time in prison is unlikely to have a substantial effect upon the results presented. We assumed all twin pairs were reared together while for full-siblings we constrained the population to siblings living together for at least 80% of the possible years together before the oldest turned 18.
As detailed elsewhere (Lichtenstein et al., Reference Lichtenstein, de Faire, Floderus, Svartengren, Svedberg and Pedersen2002), zygosity in the same-sex pairs from the twin registry was assigned using standard self-report items from mailed questionnaires which, when validated against biological markers, were 95–99% accurate. We have previously noted that the prevalence of CB is lower in same-sex MZ and DZ twin pairs than in opposite-sex twin pairs (Kendler et al., Reference Kendler, Maes, Lönn, Morris, Lichtenstein, Sundquist and Sundquist2015). This is almost surely because the former but not the latter were screened for level of cooperation, because at least one of the pair had to return a questionnaire to the twin registry and cooperation was lower in subjects with CB.
Statistical Methods
We utilized, with one exception, a classical twin model applied to male–male twins assuming three sources of liability to CB: additive genetic (A), shared environment (C), and unique environment (E). The exception arose because the number of DZ twin pairs concordant for CB in these age periods was insufficient to produce stable statistical estimates. Therefore, we added to these DZ pairs full-sibling pairs who were ≤2 years difference in age. We were also unable to obtain stable estimates for parallel analyses in female twins and siblings due to the low prevalence of CB, especially at the older ages.
The model assumes that MZ twins share all their genes while DZ twins and siblings share on average half of their genes identical by descent, and that the shared environment, reflecting family and community experiences, increases similarity to the same extent for twin or sibling pairs. Unique environment includes stochastic developmental effects, environmental experiences not shared by siblings, and random error.
We assume the same thresholds for CB for MZ and DZ twins –– given they both were weakly screened for cooperation by returning zygosity questionnaires –– and permitted a separate threshold for full-siblings who did not undergo a parallel screening.
Developmental changes in the genetic and environmental influences on CB over the three time periods (ages 15–19, 20–24, and 25–29) were modeled as a Cholesky decomposition. This developmentally informative approach divides genetic risk into three factors (A1 through A3), the first of which begins in adolescence (ages 15–19) and is continually active over the entire developmental period. The strength of its effect at each age is reflected in the path coefficients from this factor to CB at ages 15–19, 20–24, and 25–29. The second factor begins in early adulthood (ages 20–24) and impacts on CB at ages 20–24 and 25–29. The third and final factor begins at ages 25–29 and acts only at that age. A developmentally stable hypothesis for CB predicts that genetic liability to CB originates solely in the first factor with no later genetic innovation. The developmentally dynamic hypothesis predicts both genetic innovation (new genetic variation impacting on CB emerging later in development) and genetic attenuation (declining impact over time of the genetic factors acting earlier in development).
Although our sample size is considerable, the prevalence of CB in our older age groups becomes relatively rare, resulting in limited statistical power. As the objective is to quantify the nature and magnitude of developmental changes in genetic and environmental risk factors for CB, we chose to present measures of accuracy of the estimates and avoid hypothesis testing. This is in line with the recommendations based on simulations that show that in such situations, parameter estimates from the full model are typically more accurate than those from submodels, even if the latter provide a better model fit (Sullivan & Eaves, Reference Sullivan and Eaves2002). Models were fit in the OpenMx software (Boker et al., Reference Boker, Neale, Maes, Wilde, Spiegel, Brick and Fox2011).
Results
We studied a total of 69,767 twin and sibling pairs. The prevalences of CB in our three twin-sibling groups across the three age periods are outlined in Table 1. All three groups demonstrate a substantial and progressive decline in the rates of registration for CB from ages 15–19 through to ages 25–29. Rates of CB are comparable in the MZ and DZ twins but moderately higher in the full-siblings, which we capture in our modeling by allowing siblings to have a distinct threshold.
TABLE 1 The Sample Size and Prevalence of Criminal Behavior in the Sample Used –– Monozygotic and Dizygotic Twins and Full Siblings 0–2 Years Apart in Age, Born 1958–1976
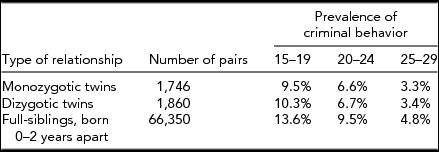
For our entire twin-sibling sample, the within-individual cross-time tetrachoric correlations (±SE) for CB from ages 15–19 to 20–24, 15–19 to 25–29, and 20–24 to 25–29 were, respectively, +0.58 (0.01), +0.53 (0.01), and +0.63 (0.01). We see substantial within-individual stability in CB over these 15 years.
Table 2 shows the within-twin tetrachoric correlations for CB in our MZ and DZ-sib samples for each of the three time periods. The MZ correlation consistently exceeds that seen in the DZ-sibs, suggesting the importance of genetic factors. However, the DZ-sib correlation exceeds half of the MZ twin correlation in all three time periods suggesting the importance of shared environmental effects. The twin correlations in both relative groups were highest at ages 15–19 and then declined, and were relatively stable from ages 20–24 to 25–29.
TABLE 2 Tetrachoric Correlations (± SE) for Criminal Behavior Within Relative Pairs by Age Period

Table 3 presents the cross-twin/sib cross-time correlations in the MZ and DZ-Sib samples. The cross-time correlations within MZ twin pairs is only modestly lower than that observed within individuals and is consistently higher than that seen with the DZ-Sib sample.
TABLE 3 Tetrachoric Correlations for Criminal Behavior Across Age Periods in Monozygotic Twins and DZ Twins/Siblings

Monozygotic twins above the diagonal, DZ twins/siblings below the diagonal.
The parameter estimates and SEs for the genetic effects from our full Cholesky model are depicted in Figure 1. As illustrated in Figure 2, a robust first genetic factor impacts strongly on liability to CB at ages 15–19 but also has a sustained but declining influence at ages 20–24 and 25–29. A second major genetic factor begins at ages 20–24 which also has essentially the same impact at ages 25–29. By contrast, the third genetic factor is very modest in its effect, accounting for only 4% of the variable in liability to CB at ages 25–29. We see, in aggregate, evidence for both genetic innovation and attenuation for CB.
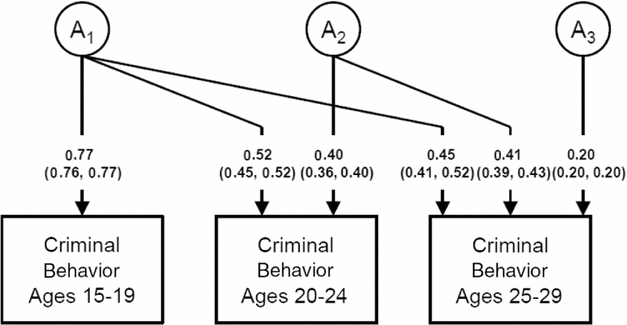
FIGURE 1 Parameter estimates (and SEs) for the genetic effects from the full Cholesky Model for Criminal Behavior at ages 15–19, 20–24, and 25–29 in Swedish male–male twin and near-aged sibling pairs. ‘A’ refers to additive genetic factors, with the subscripts 1, 2, and 3 indicating those effects that come ‘online’ at ages 15–19, 20–24, and 25–29, respectively.
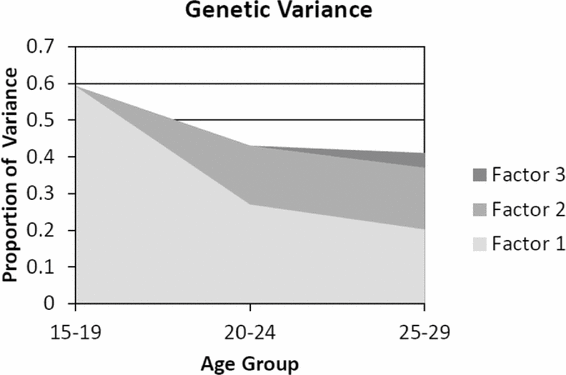
FIGURE 2 The proportion of total variance in criminal behavior accounted for by genetic factors from ages 15 to 29. The y-axis represents the total phenotypic variance so the sum of all the factors equals the total heritability. The first genetic factor which starts at ages 15–19 is represented in light grey. An intermediate grey represents the second genetic factor starting at ages 20–24. Dark grey represents the third genetic factor starting at ages 25–29.
The parameter estimates from our Cholesky model for shared and individual specific environmental risk factors are seen in Figure 3. Almost all the shared environmental effect on CB is captured by the first factor, which slowly became weaker over time. Very small effects, accounting for ≤1.0% of total variance, were seen for the second and third shared environmental factors. The large majority of the individual specific environmental effects were time-specific in their effect.
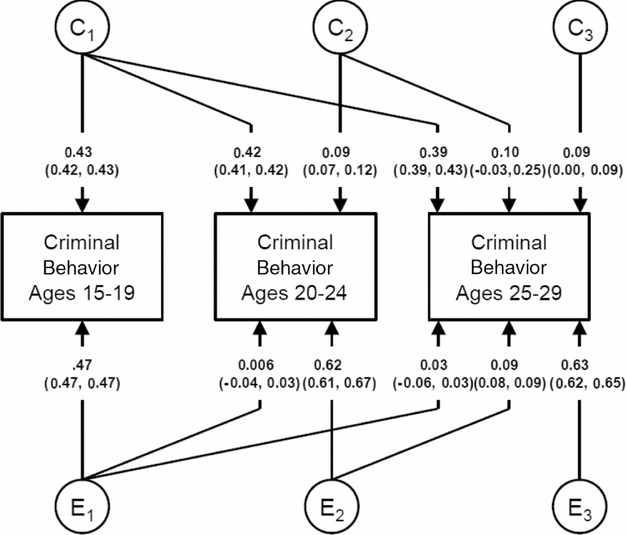
FIGURE 3 Parameter estimates (and SEs) for the shared and individual-specific environmental effects from the full Cholesky model for criminal behavior at ages 15–19, 20–24, and 25–29 in Swedish male–male twin and near-aged sibling pairs. C and E refer, respectively, to shared and individual-specific environmental factors, with the subscripts 1, 2, and 3 indicating those effects that come ‘online’ at ages 15–19, 20–24, and 25–29 respectively.
The estimates and 95% CIs for heritability and shared and specific environmental effects from the full Cholesky model for each of the three time periods are seen in Table 4. The heritability of CB declines monotonically over time from estimates of 59.4% at ages 15–19 to 41.4% at ages 25–29. Shared environmental effects also decline monotonically over time but only very slightly. By contrast, individual-specific environmental effects become progressively more important for CD with age.
TABLE 4 Estimates of Additive Genetic (a2) Shared Environmental (c2) and Individual Specific Environmental Effects (e2) by Age in the Multivariate Cholesky Model of Criminal Behavior*

Model fit -2logL = 222,806.1, ep = 21, df = 417,725 AIC = -612,643.9.
Discussion
We sought to address three major questions in these analyses, the first of which was the continuity of genetic and shared environmental influences on CB from mid-adolescence to early adulthood. Our results were inconsistent with a developmentally stable hypothesis for genetic effects on CB over the 15-year period examined. We identified one major set of genetic influences on CB, which originated in mid-adolescence and had a persistent but substantially reduced effect through early adulthood (Figure 1). We also found clear evidence for genetic innovation in a substantial second set of genetic risk factors for CB that first became active at ages 20–24. These risk factors were stable in their impact on liability to CB through age 29. After age 24, further genetic innovations were very modest in effect size.
We are aware of two prior developmental twin studies using questionnaire-based assessments of externalizing symptoms that are broadly relevant to our findings. Wichers et al. (Reference Wichers, Gardner, Maes, Lichtenstein, Larsson and Kendler2013) examined self- and parental reports of externalizing behaviors at four time points from ages 8 to 20 in Swedish twins and found evidence for both genetic innovation and attenuation. Hicks et al. (Reference Hicks, Blonigen, Kramer, Krueger, Patrick, Iacono and McGue2007) studied the trait of impulsive antisociality in male twins from Minnesota at ages 17 and 24. Congruent with our findings, 43% of the genetic influences active at age 24 were shared with those active at age 17, while 57% were novel. In aggregate, these results suggest the genetic risk for CB and the associated externalizing behaviors and personality traits are developmentally dynamic from adolescence to early to mid-adulthood.
Our results are broadly consistent with the widely discussed developmental theory of antisocial and CB of Moffitt and colleagues (Kark et al., Reference Kark, Shemi, Friedlander, Martin, Manor and Blondheim1996), which suggests that criminal offenders fall into two main types: adolescent-limited and life-course persistent. To a first approximation, our first genetic and second genetic factors might reflect the genetic liabilities to, respectively, adolescent-limited and life-course persistent CB. However, while the genetic risk factors for CB that ‘come on line’ during adolescence attenuate moderately in early adulthood, they do not disappear and hence are not adolescent-limited in their effect. Further work on criminal trajectories using genetically informative samples would be necessary to test this hypothesis more definitively.
A few studies applying models designed to examine developmental patterns of criminal offending over time have found mixed results when examining familial similarity in assignment to criminal offending groups (Besemer & Farrington, Reference Besemer and Farrington2012; van de Rakt et al., Reference van de Rakt, Nieuwbeerta and Dirk De Graaf2008). Using data from the Cambridge Study in Delinquent Development (CSDD), Besemer and Farrington (Reference Besemer and Farrington2012) examined the similarity in criminal offending trajectories between fathers and male and female offspring. They found substantial increased risk of being assigned to a criminal offending trajectory among offspring who had fathers with a criminal conviction, although the intensity of the father's record (frequency) did not impact risk among offspring. Other studies using a different data set from the Netherlands also found increased risk of criminal offending among offspring with fathers who had convictions (van de Rakt et al., Reference van de Rakt, Nieuwbeerta and Dirk De Graaf2008). They also found evidence suggesting similarity among persistence of offending in fathers and offspring. These studies have examined similarity in long-term developmental offending trajectories between parent and offspring samples; future research should also apply this approach using other types of familial relationships and across shorter developmental time periods.
The development patterns of shared environmental influences on CB differed considerably from those found for genetic factors. We found only one major factor for the shared environment that began at ages 15–19 and attenuated in its effects very slowly over time. No substantial new variation arose at later ages. This pattern of findings would be consistent with enduring effects of shared experiences in the household or community during adolescence as the median age of leaving home was approximately 21 in Sweden during this time period (Yi et al., Reference Yi, Coale, Choe, Zhiwu and Li1994).
The second major aim of this study was to determine if the heritability of CB increased over this time period. This might be predicted if active gene-environment correlation is occurring over this time period. As Caspi et al. (Reference Caspi, Elder and Bem1987) have outlined, individuals with high levels of externalizing traits can create or seek out for themselves high risk environments that can feed back positively on their genetic predisposition, thereby further encouraging deviant behaviors. A developmental twin study of peer deviance showed increasing heritabilities for peer deviance from childhood to early adulthood perhaps as a result of such processes (Kendler et al., Reference Kendler, Jacobson, Gardner, Gillespie, Aggen and Prescott2007). However, we found clear evidence for a decrease in the heritability of CB from adolescence to middle adulthood in our sample rather than the predicted increase. Wichers et al. (Reference Wichers, Gardner, Maes, Lichtenstein, Larsson and Kendler2013) also found no evidence for consistently increasing heritability for externalizing behaviors over the age period 8–20. Hicks et al. (Reference Hicks, Blonigen, Kramer, Krueger, Patrick, Iacono and McGue2007) in Minnesota twins saw a slight decrease in heritability of antisocial behavior from ages 17 to 24. While the hypothesis that risk variants for CB amplify their influence over time through active gene-environment correlation is an intriguing one, it is not supported by the available data.
The final aim of our project was to determine if shared environmental influences on CB decline with time as might be predicted by developmental theory. Here, our results were consistent with expectation as c2 estimated declined but the effects in our data were very modest, that is, from ~19% at ages 15–19 to ~17% by ages 25–29. A pattern of falling shared environmental effects over the ages 14–40 were recently seen for nicotine, alcohol, and cannabis consumption (Kendler et al., Reference Kendler, Schmitt, Aggen and Prescott2008). Hicks et al. (Reference Hicks, Blonigen, Kramer, Krueger, Patrick, Iacono and McGue2007) found appreciable levels of c2 for their latent construct of externalizing behaviors (which included antisocial behavior and nicotine, alcohol, and drug dependence) but not for antisocial behavior alone. Their estimates declined much more rapidly than we observed –– almost 50% from age 17 to 24. At least two different processes might be involved in the attenuation of shared environmental influences on CB over development. First, the psychological influences of the family environment might be diluted by an increasing accumulation of different adult experiences. Second, peer and neighborhood effects would likely decline as more individuals move away from their home and community of rearing and develop new peer groups.
Limitations
These results should be interpreted in the context of five potential methodological limitations. First, the Swedish Crime Register contains only data on criminal convictions. In Sweden as in most other countries, most crimes are not officially reported or do not result in a conviction. In the 2008 National Swedish Crime Victim Survey, the proportion of crimes reported to the police ranged from 14% for sexual offenses to 55% for serious assaults (Swedish National Council for Crime Prevention, 2008). Bias might arise if the probability that a committed crime is reported, or that a reported crime leads to a conviction, differs across social strata or between members of pairs of MZ twins versus DZ twins or siblings.
Second, we needed to combine male–male DZ twins and close in age full-siblings to obtain stable estimates. Furthermore, the numbers of DZ pairs were inadequate to support a twin-sibling model that would have allowed us to estimate the presence of a ‘special twin environment’. Our approach might have resulted in an upwardly biased estimate of heritability if environmental risk factors for CB are more highly correlated in DZ twins than in close-in-age siblings. This bias is unlikely to be substantial because when we calculated heritability of CB from MZ and DZ pairs alone, we obtained an estimate of 45% (Kendler et al., Reference Kendler, Maes, Lönn, Morris, Lichtenstein, Sundquist and Sundquist2015) in the middle of the estimates obtained here using our DZ-sibling sample.
Third, we were unable to study developmentally CB in males beyond age 29 or in females at all. Despite our substantial sample size, prevalence for CB in males after age 30 and females in even younger age groups were too low to obtain stable statistical results.
Fourth, our modeling was restricted to the twins and siblings and did not include parents. Spouses are substantially correlated for CB in Sweden (Frisell et al., Reference Frisell, Pawitan, Långström and Lichtenstein2012). If we had modeled the sources of spousal resemblance in the parents of our twins, it is possible that some proportion of the detected shared environment for CB might have been shown to result from assortative mating (Neale & Cardon, Reference Neale and Cardon1992).
Fifth, as it is typical for twin studies, we were only able to include same-sex male twins whose zygosity was known as a result of at least one member responding to a mailed questionnaire. As expected, CB was associated with a reduced probability of returning questionnaires, so the rate of CB was lower in MZ twins than in our mixed group of DZ twins and siblings. This is a form of ‘concordance-dependent’ ascertainment where the probability of known zygosity will be lowest in pairs concordant for CB, intermediate in those discordant for CB and highest in those where neither twin has CB. Simulations suggest that with the moderate level of differential ascertainment expected in our data given the observed prevalence differences, biases in parameter estimates are likely to be modest with slight underestimations of a2 and c2 and overestimation of e2 (Kendler & Eaves, Reference Kendler and Eaves1989).
Acknowledgments
This project was supported by the Ellison Medical Foundation, the Swedish Research Council to Kristina Sundquist (K2012-70X-15428-08-3), the Swedish Research Council for Health, Working Life and Welfare (In Swedish: Forte; Reg. nr: 2013-1836) to Kristina Sundquist, the Swedish Research Council to Jan Sundquist (2012-2378) as well as ALF funding from Region Skåne awarded to Jan Sundquist and Kristina Sundquist.







