


Introduction
Habitat loss is a threat to biodiversity (Brooks et al., Reference Brooks, Mittermeier, Mittermeier, Rylands, Konstant and Flick2002) and is the leading cause of population decline and species extinction in birds (Stattersfield & Capper, Reference Stattersfield and Capper2000). This is particularly significant in the tropics, where the highest biodiversity but also the highest rates of land-use change are found (Cincotta et al., Reference Cincotta, Wisnewski and Engelman2000).
The consequences of habitat loss have been studied for a range of taxa (Waltert et al., Reference Waltert, Bobo, Sainge, Fermon and Mühlenberg2005; Norris et al., Reference Norris, Asase, Collen, Gockowski, Mason, Phalan and Wade2010), showing that biodiversity usually declines along a gradient that reflects a decrease in the complexity of the vegetation structure (Schulze et al., Reference Schulze, Waltert, Kessler, Pitopang, Shahabuddin and Veddeler2004; Harvey et al., Reference Harvey, Medina, Sánchez, Vilchez, Hernández and Saenz2006). However, most of our knowledge comes from a few tropical sites that may exhibit context-specific responses (Gibson et al., Reference Gibson, Ming Lee, Pin Koh, Brook, Gardner and Barlow2011). For most areas, including much of Africa and all of Angola, there is limited or no information (Gardner et al., Reference Gardner, Barlow, Chazdon, Ewers, Harvey, Peres and Sodhi2009; Gibson et al., Reference Gibson, Ming Lee, Pin Koh, Brook, Gardner and Barlow2011).
Angola has high biodiversity as a result of its location at the confluence of six biomes (Huntley, Reference Huntley1974), yet it is also one of the least studied areas because of a succession of armed conflicts over almost 30 years. One of the most important areas for biodiversity is the Central Escarpment as it holds forests that are an evolutionary hotspot for birds (Hall, Reference Hall1960). These forests constitute the core habitat of the Western Angola Endemic Bird Area (Stattersfield et al., Reference Stattersfield, Crosby, Long and Wege1998), are a priority for global conservation (Dean, Reference Dean, Fishpool and Evans2001; BirdLife International, 2013a) and are some of the most important areas for bird conservation in Africa (Collar & Stuart, Reference Collar and Stuart1988).
Although there have been proposals for the establishment of conservation areas within this bioregion, none of the forests of the Central Escarpment fall within protected areas (Huntley, Reference Huntley1974). Moreover, during the 1950s and 1960s an estimated 95% of these forests were converted to large-scale coffee farming, although large-canopy trees were mostly left intact to provide shade (Hawkins, Reference Hawkins and Wilson1993). During the civil war (1974–2002) these farms were abandoned and some native vegetation recovered. However, more recently slash-and-burn agriculture has become widespread and represents a threat to most of the forest in this region (Ryan et al., Reference Ryan, Sinclair, Cohen, Mills, Spottiswoode and Cassidy2004; Mills, Reference Mills2010). The consequences of these activities for bird communities and threatened species remain unknown.
Kumbira Forest has been identified as the most important site for the conservation of threatened forest endemics of the Central Escarpment because it is the only known site to hold significant populations of four of the five threatened endemic birds (Gabela bushshrike Laniarius amboimensis, Gabela akalat Sheppardia gabela, Pulitzer's longbill Macrosphenus pulitzeri and Monteiro's bushshrike Malaconotus monteiri; Mills, Reference Mills2010). The aims of this study were to determine how habitats have changed within Kumbira Forest and the effects of these changes on the bird community and on endemic species. Specifically we assessed the changes in land use and land cover that have occurred in the past 2 decades, identified the most important regions of Kumbira for the endemic species, and evaluated the composition of the bird community in different land-use types.
Study area
Kumbira Forest is situated in the western Angolan province of Cuanza Sul, in the municipality of Conda (Fig. 1). The terrain varies from relatively flat at the bottom of the valley to steep on the slopes of Njelo Mountain. It is difficult to define exactly the limits of the forest because forest habitats gradually merge into thickets and other dense habitats associated with the escarpment. The eastern limits of the forest are most clearly delimited by the grasslands of Njelo Mountain. Here we define the northern limit of the forest as Cassungo village (11.104°S 14.311°E) and the southern limit as 11.230°S. Within this area, altitude varies from c. 680 m in the south to 1,160 m at the forest border on the slopes of Njelo Mountain.

Fig. 1 The study area in Kumbira Forest (a), with the land uses in 2010 and the seven sectors where the changes in forest cover were analysed (b). The bird community composition was assessed in sectors 3 and 6. The rectangle on the inset shows the location of Kumbira Forest in Angola.
Methods
To assess changes in land use and land cover we used Landsat 5 TM and 7 ETM+ satellite scenes (WRS-2 path 181 row 68) made available by the Earth Resources Observation & Science Center of the U.S. Geological Survey via the EarthExplorer interface (USGS, 2014). Such studies often benefit from using scenes from the same dates in different years (Jensen, Reference Jensen2005) but in this case scenes for similar dates were unavailable because of a failure on Landsat 7 and high cloud cover, and therefore we used a broader array of scenes from the dry season (May–September), when cloud cover was low (preferentially < 10%). Scenes meeting these requirements were from 1989, 1990, 1993, 1995, 1997, 2001, 2006, 2009 and 2010 (Supplementary Table S1).
To deal with heterogeneity of images individual scenes were radiometrically normalized and an atmospheric correction was applied, using image-based methods. We applied modified dark object subtraction (DOS4) as proposed by Chavez (Reference Chavez1996). No geometric correction was applied because the root mean square error was always a fraction of pixel dimension (< 23% for all scenes). Pre-processing analysis and calibrations were carried out in R v. 3.0.0 (R Development Core Team, 2013), with Landsat (Goslee, Reference Goslee2011) and Raster (Hijmans & van Etten, Reference Hijmans and van Etten2013). No correction was performed on the Landsat 7 scenes and the empty lines produced by the scan line failure were treated as no data.
Unsupervised ISOCLUST classification excluding thermal bands was performed on each scene, using IDRISI Selva (Eastman, Reference Eastman2012). Clusters were then reclassified to pre-defined classes (or regions of interest), which were established during a visit to the study area in 2010: bare ground and crop field, sparse/open forest, and dense forest. Accuracy of land cover data for each date was assessed by comparing the resulting classification with our 2010 ground sample points, high-resolution images from the QGIS OpenLayers plugin (QGIS Development Team, 2013), false colour composite images, and scatterplots from bare soil line/full canopy points from pseudoinvariant feature analysis (Maas & Rajan, Reference Maas and Rajan2010). We also compared the land cover spectral signatures, using the 2010 classification as a reference. Percentage cover of each class was obtained for every image to evaluate temporal changes in land use and land cover.
We also evaluated changes in dense forest cover at seven sectors of the study area (300 m radius, 28.3 ha each sector). Sectors were selected to evaluate how changes took place in three context-specific scenarios: (1) on privately owned land, (2) where there was evidence of significant forest degradation and (3) where endemic species were present.
The age of forest stands was assessed for the 2010 classification by evaluating pixel history. For each image sparse and dense forests were clumped and the reclassified images were then overlaid. To assess the areas of old forest (≥ 22 years old) we compared the fraction of unchanged forest patches in 2010 with forest area in 1989.
Endemic bird species distribution
During 13 September–2 October 2010 the first visit to Kumbira Forest took place to determine the distribution of the endemic species. Bird surveys were conducted daily at 203 sample points covering the entire study area. Sample points were spaced at least 150 m apart and located within relatively uniform habitat.
At each sample point a 10-minute point count (Bibby et al., Reference Bibby, Burgess, Hill and Mustoe2000) was conducted between sunrise (c. 05.45) and 10.30, always in good weather conditions (in the absence of rain, strong winds or high temperatures). Each morning 8–16 point counts were conducted, depending on accessibility and weather. Each point count was divided into two 5-minute periods. Between these periods we used a portable music player and amplifier to play a track of 30-second snippets of vocalizations of each of the endemic species: Monteiro's bushshrike, red-crested turaco Tauraco erythrolophus, Gabela bushshrike, Gabela akalat and Pulitzer's longbill.
All individuals of these species were recorded, irrespective of their distance from the sample point, to produce a more complete map of their distribution. Because playback violates one of the assumptions of point surveys, that birds do not approach the observer, only presence–absence data were used in the analysis. Where there was a possibility that individual birds had already been sampled during the sample period these were excluded to avoid double-counting.
Bird community in different habitats
During a second visit to Kumbira Forest during 11–29 October 2012 sampling effort was focused in the two sectors where more endemic species had been recorded in 2010. Sample points were distributed over four previously defined land-use types: (1) slash-and-burn, (2) mixed (abandoned coffee plantations or farms), (3) secondary forest (forest patches next to roads, rivers and agricultural plots) and (4) forest (the oldest forest available in the area). To examine the differences between these types we measured habitat variables at each point. We estimated canopy height and cover within 5 m of the sample point in each cardinal direction, and understorey vegetation cover within 10 m. We calculated the mean of the four measurements of each variable for each sample point. Canopy height was measured as the maximum visible height of the canopy (Dallimer et al., Reference Dallimer, King and Atkinson2009), using a laser rangefinder. Canopy cover was measured using a convex spherical densitometer and understorey vegetation was measured using a 2 m graduated white pole, marked every 10 cm with red tape. The 10-cm sections that were clearly visible from a 10 m distance were counted and converted to a percentage (Barlow et al., Reference Barlow, Haugaasen and Peres2002). Tree density per plot was calculated by counting all the trees with diameter at breast height (DBH) > 10 cm within a 20 m radius of each sample point. The distances from the observer to the nearest tree (dist1) and the nearest neighbour to this tree (dist2) were recorded to calculate a tree density index, using the formula 1/((dist1+dist2)/2) (Catry et al., Reference Catry, Mellanby, Suleiman, Salim, Hughes and McKean2000).
Data for all bird species were collected using 10-minute point counts within a 50 m radius, with no playbacks for endemics. A total of 24 points were sampled, with three repetitions for each point at different times (early morning 05.45–07.00; mid-morning 07.00–08.15 and late morning 08.15–09.30).
Data analysis
We used non-parametric Kruskal–Wallis and one-way permutation tests to evaluate differences in habitat variables between land-use types. Individual-based rarefaction curves were used to compare bird species richness across different land-use types (Gotelli & Colwell, Reference Gotelli and Colwell2001). Differences in bird communities were assessed using non-metric multidimensional scaling plots based on the Bray–Curtis similarity index. Overall difference of bird communities and pairwise difference in land-use types were measured and tested using ADONIS. All analyses were performed using R v. 3.0.0 (R Development Core Team, 2013) and vegan (Oksanen et al., Reference Oksanen, Blanchet, Kindt, Legendre, Minchin and O'Hara2012).
Results
During 1989–2010 the cover of dense forest varied in the different sectors assessed (Supplementary Fig. S1). In 1989, 53.8% (13.4 km2) of the study area was covered by forests; by 2010 this had increased to 65.2% (15.5 km2). This represents an increase of 11.4% (2.1 km2) in total forest area. However, original forest stands (present since 1989) covered only 30.8% (7.3 km2) in 2010, which represents a 45.5% loss of original forest stands (Supplementary Fig. S2).
A total of 100 bird species were recorded in 2010, including the five endemic species (Supplementary Table S2). Red-crested turaco (Fig. 2a) and Gabela akalat (Fig. 2b) were the most widely distributed, recorded at 137 (67%) and 93 (46%) sample points, respectively. Gabela bushshrike (Fig. 2c) was the third most recorded endemic, present at 42 points (21%); it was present in two well-defined zones but absent from most of the study area. Monteiro's bushshrike (Fig. 2d; 16 points, 8%) and Pulitzer's longbill (Fig. 2e; 11 points, 5%) were the least prevalent. Monteiro's bushshrike occurred almost exclusively in the upper third of the study area, with a few records further south, and Pulitzer's longbill occurred almost exclusively in the upper reaches of the valley. Combining all species (excluding the non-threatened red-crested turaco; Fig. 2f), the upper third and part of the medium third of the valley were highlighted as the most important areas for the endemics and were selected for a detailed survey.
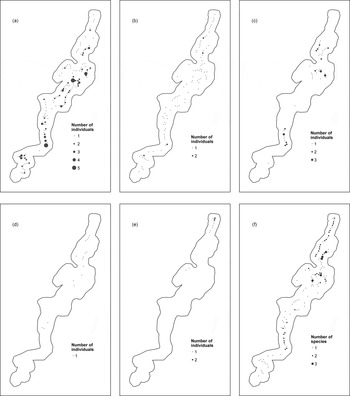
Fig. 2 Distribution of (a) red-crested turaco Tauraco erythrolophus, (b) Gabela akalat Sheppardia gabela, (c) Gabela bushshrike Laniarius amboimensis, (d) Monteiro's bushshrike Malaconotus monteiri, and (e) Pulitzer's longbill Macrosphenus pulitzeri, and number of individuals registered at each sample point. The distribution of all threatened endemic species (except the red-crested turaco) is shown in (f).
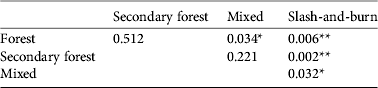
Our visual assessment of land-use types was supported by structural measurements, as there were significant differences between them (Fig. 3). We recorded 73 bird species during the survey (Supplementary Table S3). Individual-based species accumulation curves did not reach an asymptote (Supplementary Fig. S3) and we did not observe any difference in the species richness between forest classes. Bird community composition was significantly different between land-use types (P < 0.01, Fig. 4) although pairwise analysis did not show a significant difference between the bird community composition of forest and secondary forest, or between secondary forest and mixed (Table 1). However, of the threatened endemics only Gabela bushshrike and Gabela akalat were recorded, with the former apparently restricted to forest.

Fig. 3 % canopy cover, canopy height, % understorey vegetation, number of trees per plot, and vegetation index for the different land-use types: forest, secondary forest, mixed and slash-and-burn. χ2 and P values are given for each habitat variable. There were significant differences among land-use types for all habitat variables except canopy height. Lower-case letters indicate significantly different groups and black dots indicate outliers.

Fig. 4 Two axes of the non-metric multidimensional scaling (NMDS) ordination plot for bird communities in forest, secondary forest, mixed (abandoned coffee plantations or farms) and slash-and-burn. The stress value of the NMDS is 0.186, indicating that the data are well represented in the dimensions of this analysis.
Table 1 Comparison of the bird community composition for different land-use types in Kumbira Forest, Angola (Fig. 1). The values were obtained by Adonis analysis, using R v. 3.0.0.
* P < 0.05; **P < 0.01
Discussion
Remote-sensing results indicate that forest cover in Kumbira has been maintained since 1989. However, the relative stability of total forest cover masks a dynamic landscape where 34.4% of the study area was covered by forests that have regenerated since the beginning of our remote-sensing sequence in 1989. This indicates a loss of older-growth forests and their replacement with younger forests, which may have a lower conservation value (Gabela bushshrike was not present in secondary forest). Many of these younger forests are dominated by the non-native evergreen Bersama abyssinica, which is planted and used for construction and charcoal production (Schmitt, Reference Schmitt2006).
One of the limitations to assessing changes in land use and land cover in Kumbira is the lack of validation means for image classification, such as the lack of historical aerial photography. At present it is not possible using Landsat images to differentiate natural old-growth dense forests from secondary growth. Further ground surveys are needed to identify characteristic features of secondary forests and those dominated by B. abyssinica. These may be distinguishable if high variation in biomass corresponds to native deciduous species.
Red-crested turaco and Gabela akalat were the most frequently recorded endemic species. Although the global population of red-crested turaco has not been quantified and is suspected to be declining, the species has a large range and is regarded as common locally. In contrast, Gabela akalat is the most range-restricted of the escarpment's endemics (BirdLife International, 2013b). Although this species may be common locally only c. 10% of its range has suitable habitat (Mills, Reference Mills2010). Monteiro's bushshrike and Pulitzer's longbill were the least recorded endemics (< 10%) and no analyses were performed to assess their habitat preferences. Both species occur within a length of c. 370–400 km and width of c. 10–20 km of the escarpment and are common in drier forests (Mills, Reference Mills2010). They may be relatively rare in the more humid forests of the study site.
No significant relationships were found between the presence of endemics and variables describing local condition (e.g. canopy cover, understorey vegetation) or the condition of the landscape (percentage of forest cover). These non-significant results may be explained by the sampling strategy used to maximize the recording of endemic species, as points were placed to cover as much of the study site as possible instead of in specific well-defined land-use types. As the landscape in Kumbira is fragmented and dynamic, many of the sample points were influenced by edge effects and the surrounding matrix. This will have been compounded by the use of playback, which may have attracted birds into suboptimal forest types. Furthermore, some endemic species were not recorded sufficiently to evaluate their habitat use.
Overall, our a priori disturbance classes differed considerably in characteristics and quality (Fig. 3). Although the two forest classes had a similar vertical structure (canopy cover and understorey vegetation) they differed in tree cover.
Bird species richness was similar between the different land-use types, as has been found in other African forests, where bird species richness was maintained or even increased after human disturbance (Plumptre, Reference Plumptre1997; Sekercioglu, Reference Sekercioglu2002; Waltert et al., Reference Waltert, Bobo, Sainge, Fermon and Mühlenberg2005). In part, these non-significant results could be attributable to a limited sample size (particularly as some trends are apparent) but further sampling is needed to consider whether spill-over effects and landscape context could have affected our observations (Barlow et al., Reference Barlow, Gardner, Louzada and Peres2010). Species richness is an unreliable index of conservation value along disturbance gradients and other African studies have shown that a decline in forest-dependant species can be offset by an increase of non-forest and generalist species (Blankespoor, Reference Blankespoor1991; Lawton et al., Reference Lawton, Bignell, Bolton, Bloemers, Eggleton and Hammond1998; Fjeldså, Reference Fjeldså1999). There is some evidence for this in our study, as community composition varied between habitats (Fig. 4; Table 1) and slash-and-burn areas held a unique community, different from the other land-use types.
It is important to note that secondary forest held a similar bird community to the older-growth forest, because our remote-sensing analysis shows that old-growth forest is disappearing and being replaced by secondary forests across the Kumbira region. However, secondary forests are not always adequate replacements of primary forests for conserving tropical biodiversity (Barlow et al., Reference Barlow, Mestre, Gardner and Peres2007; Gibson et al., Reference Gibson, Ming Lee, Pin Koh, Brook, Gardner and Barlow2011) and their capacity to support primary forest species is poorly understood and depends on context-specific characteristics (Chazdon et al., Reference Chazdon, Peres, Dent, Sheil, Lugo and Lamb2009; Dent & Wright, Reference Dent and Wright2009). Although secondary forests are undoubtedly important components of the landscape for bird conservation, more work is needed to assess their role for endemic species.
The avifauna of biodiversity-rich forests of Kumbira, especially the endemics, remain poorly known and it is difficult to predict how species will react to further habitat changes or whether the secondary forests are being subsidized by the remaining patches of primary forest.
Further research is needed to better understand the responses of endemic birds to ongoing land-use change but there is also a need to focus research on other taxa that are less studied yet may have even more sensitive responses. Research should also examine the influence of the surrounding landscape matrix on biodiversity. Although it was not possible to demonstrate these effects in this study, landscape context may explain the patchy distribution of some of the endemic species. We hope that further conservation research can be translated into more effective policies and practices to ensure the preservation of these forests.
Acknowledgements
This research was funded by the Conservation Leadership Programme (Grant 0109710), Mr A. P. Leventis of the A. P. Leventis Ornithological Research Institute, Nigeria, and the International Turaco Society. AC is funded by a PhD grant from the Portuguese Science and Technology Foundation (SFRH/BD/78778/2011). We thank Michael Rogerson, Jose Dala and Feliciano Tchalo for their assistance in the field, and Phil Hall, Nigel Collar, Brian Huntley, Pedro Vaz Pinto, Juliet Mills and Fernanda Lages for their support.
Biographical sketches
Aimy Cáceres is interested in the effects of human activities on biodiversity, especially tropical birds, and in how to implement conservation measures. Martim Melo is interested in the processes driving the diversification of birds in Africa's endemic centres, and in the conservation action needed to safeguard these hotspots. Jos Barlow’s research focuses on assessing and improving the value of human-modified tropical forests for biodiversity. Paulo E. Cardoso is particularly interested in geographical information systems and remote sensing tools applied to habitat and species mapping. Francisco Maiato’s research interests include the vegetation structure and dynamics of Angolan miombo and escarpment forest. Michael Mills’s research interests include species identification and taxonomy and the natural history and conservation of African birds, with a particular focus on Angola.






