



Introduction
Coralline algae, a group of highly calcified marine red algae present in most euphotic zones, have received much attention because of their important role in marine ecosystems (Lei et al., Reference Lei, Huang and Huang2012) and biogeochemical cycling (Burdett et al., Reference Burdett, Hatton and Kamenos2015). They constitute the subclass Corallinophycidae Le Gall and Saunders, Reference Le Gall and Saunders2007, and are subdivided into four orders, i.e., Corallinales, Hapalidiales, Rhodogorgonales, and Sporolithales (Nelson et al., Reference Nelson, Sutherland, Farr, Hart, Neill, Kim and Yoon2015; Rösler et al., Reference Rösler, Perfectti, Pena and Braga2016, Reference Rösler, Aguirre and Braga2017). However, uncertainty besets their origin and early evolution, because fossil records of early ancestral-form (or stem-group) coralline algae are scarce, discontinuous, and are difficult to precisely assign within modern taxonomic frameworks based merely on limited morphological characters.
In South China, the Ediacaran Doushantuo phosphorous deposits yield numerous and diverse phosphatized specimens of multicellular algae (Zhang, Reference Zhang1989; Zhang and Yuan, Reference Zhang and Yuan1992; Yuan et al., Reference Yuan, Wang and Zhang1993; Zhang, Reference Zhang1996; Yuan and Hofmann, Reference Yuan and Hofmann1998; Zhang et al., Reference Zhang, Yin, Xiao and Knoll1998; Xiao et al., Reference Xiao, Knoll, Zhang and Hua1999; Zhou et al., Reference Zhou, Yuan, Xiao, Chen and Xue2004). These specimens, preserved with complex pseudoparenchymatous construction and possible reproductive structures, were interpreted as uncalcified stem-group coralline algae (Xiao et al., Reference Xiao, Knoll, Yuan and Pueschel2004), which was of great significance in indicating a soft-bodied (uncalcified) evolutionary history in the Neoproterozoic, given that Phanerozoic and living coralline algae were almost all calcified (Johansen, Reference Johansen1981; Brooke and Riding, Reference Brooke and Riding1998, Reference Brooke and Riding2000; Riding et al., Reference Riding, Cope and Taylor1998; Luchinina and Terleev, Reference Luchinina and Terleev2008; Chatalov et al., Reference Chatalov, Bonev and Ivanova2015; Teichert et al., Reference Teichert, Woelkerling and Munnecke2019). The Doushantuo specimens also demonstrate the significant potential of Ediacaran phosphatized fossil assemblages to explore the origins and early evolution of coralline algae.
Prior to this study, fossil records of erect coralline algae are all Phanerozoic in age (Johnson, Reference Johnson1960; Yang and Zhao, Reference Yang and Zhao2000; Luchinina and Terleev, Reference Luchinina and Terleev2008; Kundal, Reference Kundal2011). In this paper, a new taxon of phosphatized columnar microfossils is reported from the base of the Ediacaran Dengying Formation in China. Morphologically, the new taxon is similar to present-day erect coralline algae in both thallus morphology and internal microstructures, and hence is interpreted as a possible ancestral form of erect coralline algae in the terminal Neoproterozoic, providing a significant new insight into the origin and early evolution of coralline algae.
Materials and methods
The study area is located in Zhenba County, southern Shaanxi Province, China, tectonically placed at the northwestern margin of the Yangtze Platform (Fig. 1.1). The base of the Ediacaran Dengying Formation (ca. 551 Ma) in this area contains abundant phosphatic deposits. In the Lianghekou section, the Dengying dolostone conformably overlies the Doushantuo silty shale (Fig. 1.2, 1.3), and unconformably underlies the Cambrian Xihaoping limestone. Three lithological members of the Dengying Formation are identified (Chen et al., Reference Chen, Chu, Zhang and Zhai2015; Zhang and Zhang, Reference Zhang and Zhang2017). The lower member (6.3 m thick) consists of thin–medium bedded silty dolostone intercalated with thin bedded dolomite belts. The middle member (1.8 m thick) is characterized by interbedded medium bedded dolostone and sandstone. The upper member (> 100 m) is massive dolostone with abundant chert belts.

Figure 1. Geological setting of the studied section: (1) geographic and tectonic location of studied area; (2) lithostratigraphic column of Lianghekou section; (3, 4) outcrops: (3) boundary between the lower Doushantuo formation and upper Dengying formation (at the middle of the hammer); (4) phosphatic intraclasts embedded within the fossiliferous silty dolostone (scale bar in centimeters).
Abundant poorly sorted phosphatic intraclasts are embedded in patches (Fig. 1.4) within a 4.3 m thick layer of silty dolostone, situated 2 m above the base of the Dengying Formation. Phosphatized spherical microfossils including Megasphaera inornata Chen and Liu, Reference Chen and Liu1986, Megasphaera ornata Xiao and Knoll, Reference Xiao and Knoll2000, and Spiralicellula bulbifera Xue et al., Reference Xue, Tang, Yu and Zhou1995 have been reported from this phosphoric deposit, named the Zhenba microfossil assemblage (Zhang and Zhang, Reference Zhang and Zhang2017). Integrated stratigraphic correlation suggests a broad age constraint, which falls in the Ediacaran, but is no older than the ca. 609 Ma Weng'an biota (Zhang and Zhang, Reference Zhang and Zhang2017; Zhou et al., Reference Zhou, Li, Xiao, Lan and Ouyang2017).
Over 300 kg of phosphatic rocks were sampled from the fossiliferous horizon within the basal Dengying Formation at the Lianghekou section. Half of these samples (~150 kg) were used to make petrological thin sections. The remaining samples were dissolved via acetic acid digestion. Residues were washed in distilled water, then air dried and handpicked under a stereomicroscope. Approximately 1,200 three-dimensional (3D) columnar specimens were recovered.
Integrated morphological analyses of the Zhenba columnar fossils were supported by techniques including light and fluorescence microscopic imaging, secondary electron imaging, backscattered electron imaging, and X-ray microtomography. Compositional analysis was supported by energy disperse spectroscopy. The diameter and length of studied fossils was measured based on 3D specimens, and the thickness of internal cross laminae within the specimens was measured from thin sections, using the software Image J (https://imagej.nih.gov/ij/).
Repository and institutional abbreviation
All specimens presented herein are stored in the Shaanxi Key Laboratory of Early Life and Environments (LELE), Northwest University, Xi'an, China (with specimen accession numbers ranging from ZBDY02A03 to ZBDY44H13 for 3D specimens, and ZBDY-TS-02-01 to ZBDY-TS-563-16 for thin sections).
Morphological structure of the columnar microfossils
The phosphatized columnar microfossils from Zhenba County, Shaanxi Province, are all preserved as fragments, to 1,200 μm in length. They are commonly presented as isolated phosphatic intraclasts or preserved as concentrated accumulations within individual phosphatic intraclasts (Fig. 2.1). Most specimens are straight, although some show curved columns. The maximum diameter ranges 100–1,300 μm (Fig. 3, based on 3D specimens).

Figure 2. Concentrating preservation of the Zhenba columnar specimens in thin section: (1) phosphatic intraclast consisting of concentrated fragments of columnar specimens; (2) longitudinal section of one specimen, showing internal structures of cross laminae; (3–5) cross sections of several specimens, showing varied and irregularly shaped cross sections.

Figure 3. Size statistics of the Zhenba columnar specimens.
A total of 550 specimens were observed under SEM, varying greatly in column morphology. In cross section, they are mostly irregularly shaped (Fig. 4), although partial specimens show subrounded (Fig. 4.1), subreniform (Fig. 4.2), or subtriangular (Fig. 4.3) cross sections. Almost every specimen is unique in column morphology. Collectively, the phenomenon reveals unconstrained column morphology. It is difficult to explain such morphological variation citing differences in preservation because the cross-sectional shape remains stable from the bottom to the top in every specimen. We consider the great majority of morphological variation among Zhenba columar microfossils as a result of their nondirectional lateral growth. Irregularly spaced, nearly transverse cracks are frequently seen on the surface of specimens. In broken specimens, internal structures of closely stacked cross laminae are well exposed (Fig. 5.7, 5.9). A contracted end (Fig. 5.2, 5.3, 5.4) was observed in 35 specimens, either rounded or tapered in morphology. Five specimens show structures of an external ‘wall.’ The ‘wall’ that frequently envelopes specimens entirely is either smooth (Fig. 5.11) or transversely cracked (Fig. 5.5) and varies 3–23 μm in thickness. In addition, one specimen shows that the columns can branch into two differently sized subordinate columns at the column apex (Fig. 5.13), indicating that the Zhenba columnar microfossils probably obey a branching growth pattern.

Figure 4. Variation of the Zhenba columnar fossils cross-sectional shapes under SEM, with individual outlines below: (1) subrounded; (2) subtriangular; (3) subreniform; (4) irregularly shaped. Scale bars = 100 μm.
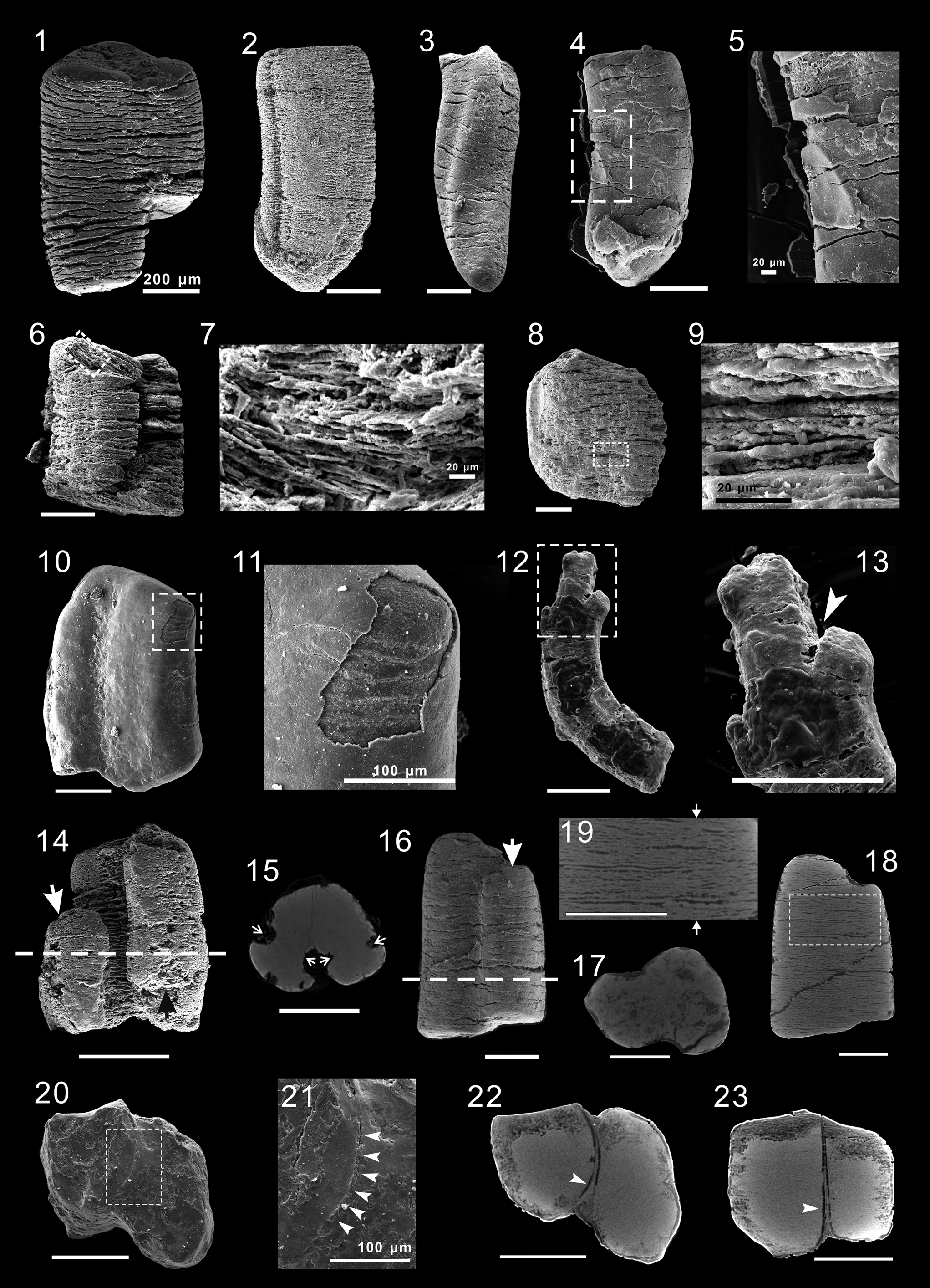
Figure 5. Zhenba columnar specimens under SEM: (1) ZBDY024F13, densely cracked specimen; (2) ZBDY03E012, specimen with a rounded end; (3) ZBDY024G15, specimen with a tapered end; (4) ZBDY032C01, specimen with cracked external ‘wall’ structure; (5) detail of inset in (4); (6, 8) ZBDY031D12 and ZBDY02B10, respectively; (7, 9) details of insets in (6) and (8), respectively, showing closely stacked laminae inside column; (10) ZBDY028B13, specimen with a smooth external ‘wall’; (11) detail of inset in (10); (12) ZBDY44F05, specimen with branched columns; (13) detail of inset in (12), indicating branched column (arrow); (14) ZBDY024E27, specimen with three subordinated columns, showing two smaller branches (arrows); (15) transverse virtual slide at dotted line in (14), showing contraction at the joints between subordinate columns (arrows); (16) ZBDY035A16, specimen with two subordinate columns, indicating smaller branch (arrow); (17) transverse virtual slide at dotted line in (16); (18) longitudinal virtual slide of (16); (19) detail of inset in (18), showing structure of internal cross laminae, with position marked (arrows) of the joint between the two subordinate columns; (20) ZBDY02A06, specimen resulting from adhesion of two isolated columnar individuals; (21) detail of inset in (20), with boundary marked (arrows) between two individual columns; (22, 23) transverse and longitudinal virtual slides of specimen in (20), showing the irregular cross sections of the two individual columns and the boundary between them (arrows). Unlabelled scale bars = 200 μm.
Eight 3D specimens revealed that the entire column consists of differently sized subordinate columns that are longitudinally connected. Two of them were analyzed using micro computed tomography (Micro-CT). The results showed that one specimen contains three subordinate columns (Fig. 5.14) whereas the other contains only two (Fig. 5.16). Virtual slides reveal that the subordinate columns are interconnected (Fig. 5.18, 5.19), and the joints between them show distinct contraction (Fig. 5.15, 5.17). Internal structures of cross laminae are well exhibited in longitudinal virtual slides (Fig. 5.18, 5.19) and their construction is highly consistent with those specimens seen in thin sections. These morphological features collectively indicate that the specimens were not formed by coincident adhesion of isolated columns, but probably were undergoing longitudinal fission. There do exist a small number of specimens that each consist of two isolated columns (Fig. 5.20) with both columns exhibiting irregular cross sections (Fig. 5.22). A boundary structure separating the two columns can be distinctly observed in both SEM and Micro-CT images (Fig. 5.21–5.23, arrows). They are typically formed by coincident adhesion of isolated columnar individuals.
Over 300 specimens in thin sections were identified and are consistent with observations from 3D specimens from acidic resistant residues in showing irregularly shaped cross sections (Fig. 2.3, 2.4, 2.5). An external wall-like structure was found preserved in 15 specimens, with their thickness ranging 6–37 μm, and appearing at different positions in the same specimen. The ‘wall’ commonly shows concentric layers (Fig. 6.4) and can envelope specimens entirely (Fig. 6.1). In other cases, the wall is partially peeled off, leaving remnants attached to specimens (Fig. 6.2, 6.3). However, the ‘wall’ might not be a biological structure and might be better interpreted as a phosphatic coating formed during diagenesis. Evidence comes from at least three aspects: (1) fragmentary specimens are entirely enveloped in such a ‘wall’ structure; (2) there is no diagnostic feature, e.g., plastic deformation or ornamentation, that can be used to identify the wall structure as a biological structure; and (3) the same ‘wall’-like structure is frequently observed in coexisting clastic particles (Fig. 6.4–6.8).

Figure 6. ‘Wall’-like structure outlining columnar microfossils and coeval clasts of Zhenba columnar fossils in thin section: (1–3) specimens with external ‘wall’-like structures: (1) ZBDY-TS-107-01; (2) ZBDY-TS-82-01; (3) ZBDY-TS-304-05; (4) detail of inset in (3), showing concentric layers (arrowhead); (5) ZBSPDY-626, quartz grains with phosphatic coating, showing concentric layers (arrowhead); (6) image in (5) under cross-polarized light; (7) ZBSPDY-626, phosphatic intraclast with phosphatic coating; (8) image in (7) under cross-polarized light.
In specimens with high fidelity, the internal structures of closely stacked cross laminae are well exhibited. In longitudinal section, these cross laminae are thin and flat, and vary in morphology from lenses to nonisopachous belts (Fig. 7.2, 7.3, 7.5). These lenses and/or belts vary 1.5–12 μm in maximum thickness, and their diameter ranges 2.4–84% of the column width. The thickness has no correlation with the column width. In construction, these lenses/belts are rarely horizontally continuous inside columns and are imbricated and compressed. The space between adjacent laminae ranges 0–5 μm depending on specimen and position, probably reflecting different preservation.

Figure 7. Cross laminae structures of Zhenba columnar fossils in thin section: (1) ZBDY-TS-323-03; (2, 3) detail of inset in (1), under polarized-light and autofluorescence, respectively, showing closely stacked cross-laminae structures; (4) ZBDY-TS-540-11; (5) autofluorescence image of inset in (4), with white dotted lines marking the outlines of cross laminae inside the column; (6) ZBDY-TS-435-07, under autofluorescence, showing brittle fractures of cross laminae; (7) ZBDY-TS-563-07, showing plastic deformation of cross laminae; (8) detail of inset in (7), under autofluorescence; (9) ZBDY-TS-333-03, under autofluorescence, showing irregularly shaped cavity (arrow) resulting from decomposition; (10) ZBDY-TS-220-02; (11) detail of inset in (10), under autofluorescence, showing partial cross laminae enveloped by membrane-like structure (arrows).
Both plastic deformation (Fig. 7.8) and brittle fractures (Fig. 7.6) are commonly observed on cross laminae. Some poorly preserved specimens show irregularly shaped cavities (Fig. 7.9) probably resulting from decomposition. Such taphonomic features together suggest a relatively hard skeleton for the Zhenba columns, possibly a mixed composition of soft tissues alongside calcified components. Because it is difficult to exclude the possibility that the brittle fractures can form after mineralization, a soft-body explanation for the Zhenba columns remains viable.
Fine structures of cross laminae can be seen in rare specimens. As shown in Figure 8, the cross laminae each consist of two distinct microstructures including micron-grade grains (Fig. 8.3, 8.4) and cuticle-like fabric (Fig. 8.3, arrows). In construction, the grains are arranged either tightly or loosely in spaces sandwiched by adjacent cuticle-like fabrics. Element mapping (Fig. 8.5–8.14) shows that these fine grains and cuticle-like fabrics are rich in phosphorus and calcium, whereas spaces between these microstructures are rich in silicon, aluminium, potassium, magnesium, oxygenium, and iron. Accordingly, we infer that the grains and cuticle-like fabrics are apatite in composition, whereas the spaces between them are comprised of clay. The fine grains are solid and vary from rectangular to polygonal in morphology. The polygonal grains are dominated by diverse patterns including regular hexagons, irregular pentagons, and triangles. Their size ranges 1–3 μm in width and 1–4.4 μm in length. In size and morphology, these grains are highly consistent with automorphic crystals of apatite. Additionally, their solid textures and disordered arrangements do not support a cellular interpretation. We propose that these grains probably represent diagenetic artifacts rather than biological structures. The cuticle-like fabrics are horizontally discontinuous, and commonly show plastic deformation. They probably represent original boundaries of the cross-laminae structures.

Figure 8. Fine microstructure of Zhenba columnar fossil ZBDY-XY-TS-Yao-63-01 in thin section: (1) light microscopic image; (2) backscattered electron image of (1); (3) detail of inset in (2), showing cuticle-like structures (arrows); (4) detail of inset in (1), under autofluorescence; (5–14) elemental mapping of inset in (3).
Novel taxon for the Zhenba columnar microfossils
The phosphatized preservation style, fragmental nature, and columnar morphology of these Zhenba specimens are reminiscent of tubular microfossils represented by Ramitubus Liu et al., Reference Liu, Xiao, Yin, Zhou, Gao and Tang2008, Sinocyclocyclicus Xue, Tang, and Yu, Reference Xue, Tang and Yu1992, Quadratitubus Xue, Tang, and Yu, Reference Xue, Tang and Yu1992, and Crassitubus Liu et al., Reference Liu, Xiao, Yin, Zhou, Gao and Tang2008 from the Weng'an biota (Xue et al., Reference Xue, Tang and Yu1992; Liu et al., Reference Liu, Xiao, Yin, Zhou, Gao and Tang2008). However, morphologically the Zhenba columnar fossils substantially differ from those Weng'an tubular fossils in a number of aspects.
First, although there exists partial overlap in size between the two populations, the Zhenba population columns are much larger than the tubes of the Weng'an population in both size range and typical size (the Weng'an population tubes vary 90–250 μm in maximum width, with typical diameter 146–209 μm [Liu et al., Reference Liu, Xiao, Yin, Zhou, Gao and Tang2008], whereas the Zhenba population columns vary 100–1,300 μm in maximum width, with typical diameter 300–400 μm). Such a significant difference in size should not be neglected when comparing the two populations.
Second, in contrast to the Weng'an tubes, which are either cylindrical or tetragonal, the Zhenba columns show variable, irregular, frequently asymmetric shapes in cross section. Partial specimens with subreniform (Fig. 4.2) or subtriangular (Fig. 4.3) cross sections might lead one to consider whether they can be explained as two or three individuals that are subrounded in cross section. However, such suspicions can be removed by the absence of an observed boundary between those subjective subdivisions. So far, no specimen that consist of pairs/groups of regularly cylindrical/tetragonal individuals has been observed among Zhenba columnar microfossils. Additionally, regularly cylindrical/tetragonal specimens within the Zhenba population are extremely rare: the irregular cross sections exhibited by the Zhenba population are less likely a result from linked/adjacent pairs/groups of cylindrical/tetragonal individuals like the Weng'an tubular microfossils, but more likely represent their original morphology throughout a single individual column.
Third, the Zhenba columns fundamentally differ from Weng'an tubes in body construction. All Weng'an tubular specimens consist of parallel, isopachously stacked plates that are horizontally continuous. In contrast, the Zhenba columnar specimens consist of imbricatedly stacked belts that are morphologically different, exhibiting lenses to nonisopachous belts. One could suspect that the discontinuous, imbricatedly stacked belts shown by the Zhenba columns might have been transformed from original continuous, isopachously stacked plates, similar to those of the Weng'an microtubular specimens. However, based on existing fossil materials, it is more likely that the morphology and construction of these lenses/belts represent the original biological structure of the Zhenba columns, because: (1) no specimen that consists mainly of continuous belts has been observed among the Zhenba columnar fossils; (2) the lenses/belts are more frequently observed in those specimens with high fidelity; (3) among the Weng'an microtubular specimens consisting of continuous, isopachously stacked plates, no similar structures of imbricatedly stacked lenses/belts has been reported; and (4) in partial specimens, the belts/lenses are observed completely enveloped by a membrane-like structure (Fig. 7.10, 7.11), which refutes an explanation that the belts/lenses originated from incomplete infilling of plate-like cells. Moreover, even if we suppose that the imbricatedly stacked belts inside the Zhenba columns were transformed from their originally parallel, isopachously stacked plates, they are relatively thinner than those of the Weng'an microtubular specimens in thickness. For the Zhenba columnar specimens, the typical thickness of the ‘plates’ is 3 μm, whereas for the Weng'an microtubular specimens, the plates range 7–12 μm in typical thickness (Liu et al., Reference Liu, Xiao, Yin, Zhou, Gao and Tang2008).
Fourth, unlike the Weng'an population tubes, which are enveloped by single- or multiple-layered walls with/without annulations on external surfaces (Liu et al., Reference Liu, Xiao, Yin, Zhou, Gao and Tang2008), no similar structure has yet been identified within the Zhenba population. This indicates that such a tube wall structure probably did not exist in the Zhenba columnar fossils.
In addition to the Weng'an population, there exist other diverse tubular fossils in late Ediacaran to early Cambrian sedimentary strata, represented by taxa including Conotubus Zhang and Lin in Lin et al., Reference Lin, Zhang, Zhang, Tao and Wang1986, Cloudina Germs, Reference Germs1972, Sinotubulites Chen, Chen, and Qian, Reference Chen, Chen and Qian1981, emend. Chen and Sun, Reference Chen and Sun2001, Shaanxilithes Xing, Yue, and Zhang, Reference Xing, Ding, Luo, He and Wang1984, and Sabellidites Yanishevsky, Reference Yanishevsky1926. Although these taxa overlap partially with the Zhenba columns in diameter, they differ essentially from the Zhenba columns in body construction. Conotubus, Cloudina, and Shaanxilithes are characterized by tube walls that consist of nested cones (Chen and Sun, Reference Chen and Sun2001; Cai et al., Reference Cai, Schiffbauer, Hua and Xiao2011, Reference Cai, Hua, Schiffbauer, Sun and Yuan2014, Reference Cai, Cortijo, Schiffbauer and Hua2017; Wang et al., Reference Wang, Zhang, Zhang, Cui and Li2021). Sinotubulites and Sabellidites are characterized by a multiple-layered tube wall (Chen and Sun, Reference Chen and Sun2001; Chen et al., Reference Chen, Bengtson, Zhou, Hua and Yue2008; Moczydlowska et al., Reference Moczydlowska, Westall and Foucher2014; Cai et al., Reference Cai, Xiao, Hua and Yuan2015). All of these tube-wall structures are absent from the Zhenba columnar fossils. The observed morphological characters, including variable cross sections and imbricated construction of cross belts inside the columns, collectively distinguish the Zhenba columnar fossils from these Ediacaran tubular microfossils and any other tubular forms of other geological periods, and therefore clarify a novel taxon of the Ediacaran. However, it is hazardous to formally establish a new taxon based only on these fragmental columns, and so we here restrict our study of these remains to a general description.
Biological affinity
Determining the biological affinities of a Precambrian taxon based on information from fragmented specimens is challenging. However, despite incomplete preservation, the Zhenba columnar fossils show diverse features in morphology, anatomy, and physiology, which are of great value in constraining their biological affinity. Three distantly related groups of living organisms—(1) filamentous cyanobacteria akin to Oscillatoria Vaucher in Gomont, Reference Gomont1892, (2) cnidarians akin to tabulate corals, and (3) multicellular algae akin to corallines—could be comparable with the Zhenba columns in body structure, and accordingly are considered here as potential candidates for interpreting the Zhenba columns.
Filamentous cyanobacteria akin to Oscillatoria
Among living cyanobacteria, colonial groups like Oscillatoria are composed of thin discoid cells that are closely stacked to form filaments. They resemble the Zhenba columns in gross morphology as well as laminated inner structure.
However, a filamentous-cyanobacteria interpretation is incompatible with the present columnar fossils in at least four aspects. First, the body size of living filamentous cyanobacteria (< 100 μm in diameter) is much smaller than that of the Zhenba columns (100–1,300 μm diameter). It is also less likely that early filamentous cyanobacteria could reach the equivalent body size of the Zhenba columns, given that early-Cambrian fossils of filamentous cyanobacteria are consistent with modern counterparts in size and structure (Shang et al., Reference Shang, Liu, Yang, Chen, Wang and Kouchinsky2016). Second, filamentous cyanobacteria differ from the Zhenba columns in the shape of their cross section. All living and fossil filamentous cyanobacteria are cylindrical, and their cross section is without exception circular. However, the Zhenba columns have variable and irregular shapes of cross section; only a minority of specimens show a subcircular cross section. Third, cross walls inside filamentous cyanobacteria are horizontally continuous, although incomplete cross walls are frequently observed between adjacent cross walls. In contrast, cross laminae inside the Zhenba columns are frequently discontinuous and are commonly represented by nonisopachous belts in longitudinal sections. Finally, longitudinal fission and upward branching in the Zhenba columns is never seen in living filamentous cyanobacteria.
These significant differences do not support a filamentous-cyanobacteria interpretation. Furthermore, the large size and complex tissue construction of the Zhenba columns is incompatible with any individual and/or colonial unicellular organisms, and therefore strongly suggests an interpretation as a multicellular organism.
Tabulate corals
Among living Metazoa, diverse groups are able to form columnar body plans, although no group exhibits structures of cross laminae inside their bodies. However, the tabulate corals, an extinct group of cnidarians, were able to form ‘cross walls’ inside their tubular skeletons, which are somewhat similar in construction to the ‘cross laminae’ inside the Zhenba columns. Tabulate corals also obey an upward-branching growth pattern, and because of their colonial lifestyle, individuals commonly show variable and irregularly shaped cross sections. However, the tabulate-coral interpretation is incompatible with morphological characters shown by the Zhenba columns in the following two aspects. First, all tabulate corals live in tubes secreted by themselves. Therefore, a tube wall structure is required for their identification. However, no such structure has been recognized among the Zhenba columns (which is why we illustrate them as columnar fossils instead of tubular fossils). Second, the longitudinal fission present in the Zhenba columns implies that the ‘cross laminae’ structures were alive and therefore able to conduct growth and fission. However, such a phenomenon is absent in tabulate corals, in which ‘cross walls’ are nonliving structures that represent abandoned parts of the polyp. Integrated morphological comparison therefore does not support a tabulate-coral interpretation.
Multicellular algae akin to corallines
Morphological convergence is pervasive in living multicellular algae. It is difficult to constrain biological affinities of the Zhenba columns based merely on gross morphology because many distantly related groups have columnar thalli. However, tissue construction can provide valuable information in phylogenetic analyses.
As illustrated above, the columnar fossils from Zhenba exhibit construction of closely stacked ‘cross laminae.’ Such tissue construction is diagnostic. Although some taxa of multicellular green algae, e.g., Cladophora Kützing, Reference Kützing1843, Penicillus Lamarck, Reference Lamarck1813, and Rhipocephalus Kützing, Reference Kützing1843, are also able to form columnar and branched thalli (Graham and Wilcox, Reference Graham and Wilcox2000), they differ from the Zhenba columns in lacking stromatolithic structures inside the thalli. Among living multicellular algae, only coralline red algae are able to form stromatolithic tissues that resemble the construction of cross laminae seen in the Zhenba columns. Physiological features exhibited by the Zhenba columns, including an upward-branching growth pattern and longitudinal fission, are also consistent with some modern coralline algae (Johansen, Reference Johansen1969; Walker et al., Reference Walker, Brodie, Russell and Irvine2009). Collectively we suppose that the Zhenba columns possibly represent fossils of early coralline algae. Considering that the partial Weng'an tubular microfossils were also interpreted as multicellular algae similar to some living coralline algae (Cunningham et al., Reference Cunningham, Vargas, Liu, Belivanova, Marone, Martínez-Pérez, Guizar-Sicairos, Holler, Bengtson and Donoghue2015), it is also possible that the Zhenba columnar fossils might represent relatives of those Weng'an specimens, although they show distinct morphological differences.
The Zhenba columns show a large size range (100–1,300 μm diameter) and morphological variation (varied shapes of cross sections). Interestingly, intergenicula of main branches in several species of living coralline algae range 300–1,000 μm in width (Walker et al., Reference Walker, Brodie, Russell and Irvine2009), and also show thalli of varied and inconsistent forms, ranging from cylindrical to compressed morphologies (Walker et al., Reference Walker, Brodie, Russell and Irvine2009). Morphological variation in marine algae is relatively common and is affected by diverse environmental factors, such as light, temperature, salinity, availability of nutrients, current motion, etc. (Lobban and Harrison, Reference Lobban and Harrison1997). Furthermore, if developmental factors are taken into account, the marked differences in size and morphology among individuals would be amplified significantly.
Taxonomic and phylogenetic placement of Zhenba columns in coralline algae
Modern coralline algae are traditionally classified into two morphological groups: geniculate forms and nongeniculate forms based on the presence or absence of genicula. Geniculate corallines, which are typically highly branched and segmented, are characterized by the presence of uncalcified genicula between calcified regions known as intergenicula. Nongeniculate forms are mainly encrusting organisms in which the entire lower surface adheres to a substratum; some taxa are also able to form an arborescent canopy with erect stipes, e.g., Mastophoropsis Woelkerling, Reference Woelkerling1978 (order Hapalidiales) and Metamastophora Setchell, Reference Setchell1943 (Corallinales) (Woelkerling, Reference Woelkerling1988), or columnar protuberances, e.g., Neogoniolithon Setchell and Mason, Reference Setchell and Mason1943 (Johansen, Reference Johansen1981).
Without evidence for genicula-intergenicula differentiation, it is difficult to determine whether the Zhenba columnar fossils are nongeniculate or geniculate, because both forms of coralline algae are able to form columnar thalli. Given that the Zhenba specimens show branched, columnar thalli, if they do represent fossils of ancestral coralline algae, then they more likely belong to erect rather than crustose forms.
Tissue stratification is relatively common among living coralline algae. Although both erect and crustose forms are able to form stratified tissues (medulla in erect forms, perithallus in crustose forms), some crustose forms are more comparable with the Zhenba columns in tissue construction than erect forms. Among living erect coralline algae, medullar tissues are highly stratified and commonly coaxial. Meanwhile, in some thin-crust forms of coralline algae, e.g., Tanarea tessellatum (Lemoine, Reference Lemoine1929) Littler, Reference Littler1971, the thalli can be as thin as only two cells thick (Adey et al., Reference Adey, Townsend and Boykins1982); the repeated overgrowth of thalli is able to form structures of horizontally stacked thin layers that more resemble the construction of cross laminae inside the Zhenba columns. Some crustose forms, e.g., Hydrolithon reinboldii Weber-van Bosse and Foslie, Reference Foslie1909, are also able to form nonisopachous lenticular tissues that are several cells thick (Massieux, Reference Massieux and Flügel1977). The superposition of these lenticular zones is astonishingly like the construction of cross laminae inside the Zhenba columns. However, these species are all formless because of their crustose growth form.
Enigmatically, the Zhenba columnar fossils more closely resemble erect coralline algae in gross thalli morphology, but are more comparable with crustose forms in tissue construction. Consequently, no living coralline algae comprehensively matches the Zhenba columns in integrated morphology. The Zhenba columns thus probably represent an extinct group of ancient coralline algae. Given the morphological similarity between stratified tissues in some living coralline algae and the lens/belt structures within the Zhenba columns, we prefer to interpret the lenses/belts as tissue-lever structures whose thickness is controlled by the different numbers of cell layers. Unfortunately, cellular structures were not preserved, leaving no cellular to subcellular diagnostic characters, e.g., pit connections and two-layered calcified cell walls, with which to strengthen a coralline-algae interpretation.
Although morphological features collectively favor an interpretation of coralline algae, the Zhenba columns show distinct differences from present-day coralline algae. First, the thallus composition differs between Zhenba columns and modern coralline algae. As discussed above, taphonomic characteristics collectively support a soft-bodied or weakly calcified nature of the Zhenba columnar specimens, whereas extant erect coralline algae, as well as their fossilized counterparts, are all highly calcified. The second difference lies in tissue/organ differentiation. Modern coralline algae are highly differentiated (Walker et al., Reference Walker, Brodie, Russell and Irvine2009). In geniculate forms, thalli are commonly differentiated into three types of tissues including, from innermost to external, medulla, cortex, and epidermis. In nongeniculate forms, thalli are commonly differentiated into three types including, from ventral to dorsal, hyperthallus, perithallus, and epithallus. Conceptacle structures, used in reproduction, are also important characters for generic identification in modern coralline algae. Neither distinct tissue differentiation nor conceptacle structures has so far been recognized in the Zhenba columnar specimens.
Compared to modern erect coralline algae, the Zhenba columns not only lack highly calcified skeletons, but also show a simpler tissue/organ complexity. However, it is not incompatible to interpret the Zhenba columns as ancestral coralline algae. First, ancestral coralline algae probably had an uncalcified evolutionary history (Johnson, Reference Johnson1956; Xiao et al., Reference Xiao, Knoll, Yuan and Pueschel2004; Smith and Butterfield, Reference Smith and Butterfield2013). Therefore, it makes sense that soft-bodied forms or weakly calcified groups could exist among early ancestral coralline algae. Second, the fossil record illustrates that early coralline algae commonly show simpler thallus construction than modern representatives. For example, those genera suggested as ancestral coralline algae, e.g., Cuneiphycus Johnson, Reference Johnson1960 (late Carboniferous; Johnson, Reference Johnson1960), Tharama Wray, Reference Wray1967 (Devonian; Wray, Reference Wray and Flügel1977), and Solenopora gotalandica Rothpletz, Reference Rothpletz1908 (Ordovician; Brooke and Riding, Reference Brooke and Riding1998), also lack differentiated tissues. Therefore, tissue differentiation probably represents a deuterogenic characteristic in the evolution of coralline algae. Third, the lack of conceptacle structures among the Zhenba columnar specimens might have resulted from preservation bias or limited excavation. Although considering the view that the conceptacle probably represents a derived character of coralline-algae (Johnson, Reference Johnson1956), it is also possible that conceptacle structures were not present in the Zhenba columns. Morphological features and physiological phenomenon seen in the Zhenba columnar specimens collectively support an interpretation of soft or weakly calcified, erect coralline algae. We cannot phylogenetically place the Zhenba columns within the Corallinophycidae because anatomical features that are phylogenetically informative, e.g., cell fusion, secondary pit connections, mode of tetrasporangial conceptacle roof formation, position of spermatangia in conceptacle chambers, etc., are not observed in the Zhenba columns. Because complex tissue-organ differentiation and highly calcified skeletons, which characterize the crown group Corallinophycidae, are lacking from the Zhenba specimens, it is reasonable to interpret the Zhenba columnar specimens as a stem group of the Corallinophycidae.
Implications for early evolution of coralline algae
Recently, fossils of the order Corallinales were identified from the Silurian of Gotland (Teichert et al., Reference Teichert, Woelkerling and Munnecke2019), over 300 million years earlier than the age previously estimated based on molecular data (Aguirre et al., Reference Aguirre, Perfectti and Braga2010; Yang et al., Reference Yang, Boo, Bhattacharya, Saunders, Knoll, Fredericq, Graf and Yoon2016; Rösler et al., Reference Rösler, Aguirre and Braga2017). The difference in divergence time of coralline algae estimated by molecular data is probably because those putative ancestral coralline fossils were not used as calibration points. The gap in divergence time of coralline algae between estimations made by molecular data and the fossil record can be reconciled by using stem-group fossils as calibration points. Using a seven-gene concatenated dataset and Neoproterozoic ancestral coralline algae fossils as a calibration point, Yang et al. (Reference Yang, Boo, Bhattacharya, Saunders, Knoll, Fredericq, Graf and Yoon2016) estimated that the emergence of the subclass Corallinophycidae occurred at ca. 579 (543–617) Ma. This is quite compatible with the discovery of stem group Corallinophycidae in the Zhenba microfossil assemblage, which was constrained to within the Ediacaran (Zhang and Zhang, Reference Zhang and Zhang2017), no older than Weng'an biota (ca. 609 Ma; Zhou et al., Reference Zhou, Li, Xiao, Lan and Ouyang2017).
Erect coralline algae are relatively abundant and diverse in deposits from the Cretaceous to the present day (Kundal, Reference Kundal2011), and most of them are geniculate in form, but the existence of Paleozoic counterparts (Johnson, Reference Johnson1960; Yang and Zhao, Reference Yang and Zhao2000; Luchinina and Terleev, Reference Luchinina and Terleev2008) implies an early origin for erect coralline algae. These Paleozoic representatives are relatively sparse, probably because the erect coralline algae occupied a smaller range of ecological niches at that time. The discovery of the Zhenba columnar microfossils could extend the origin of erect coralline algae back to the Ediacaran Period. These noncalcified or possibly weakly calcified counterparts along with those highly calcified counterparts in the Paleozoic, together reveal a continuous evolution of erect coralline algae before their large-scale radiation in Mesozoic.
Although there is no evidence for genicula-intergenicula differentiation among the Zhenba erect coralline algae, it cannot be excluded that the Zhenba erect coralline algae could be ancestral to subsequent geniculate forms, because genicula are nonhomologous structures that evolved independently in diverse clades of coralline algae (Bailey and Chapman, Reference Bailey and Chapman1998). The presence of genicula among different lineages of geniculate coralline algae probably represents a phenomenon of convergent evolution for coping with different degrees of current energy (Janot and Martone, Reference Janot and Martone2018). Therefore, geniculate forms must have evolved from nongeniculate forms, which is well supported by the existence of transitional species and evolutionary links between geniculate and nongeniculate coralline algae (Mude and Kundal, Reference Mude and Kundal2011).
The identification of the Zhenba erect Coralline algae as a stem group within the Corallinophycidae implies that the morphological differentiation of crustose and erect forms among coralline algae probably occurred early in their evolutionary history, before large-scale biocalcification in the Paleozoic. Considering that an erect lifestyle makes coralline algae capable of occupying the water column, out of the reach of crustose forms, the emergence of the Zhenba erect forms could also imply an initial ecological expansion of early coralline algae in the Ediacaran.
Conclusions
A new form of columnar microfossil is described from the Ediacaran Zhenba microfossil assemblage. Integrated analyses of internal and external morphology indicate the Zhenba columns resemble some modern coralline algae in thallus morphology and tissue construction but differ from modern counterparts in lacking calcified skeletons and tissue-organ differentiation. Consequently, anatomical comparison supports a phylogenetic interpretation of stem-group Corallinophycidae. The Zhenba columns possibly represent the oldest fossil record of erect coralline algae. The new finding also implies that Ediacaran coralline algae are more diverse than previously thought, and that morphological differentiation and ecological expansion probably occurred early among stem-group coralline algae.
Acknowledgments
We are grateful to H. Gong, Y. Yao, and J. Sun for technical assistance, and to W. Liu, Y. Yang, L. Cui, L. Li, H. Yun, and M. Wang for sampling. This work was financially supported by Natural Science Foundation of China (grants 41890845, 42102012, 41621003, and 41930319), 111 Project (D17013), Strategic Priority Research Program of Chinese Academy of Sciences (grant XDB26000000), and Key Scientific and Technological Innovation Team Project in Shaanxi Province. The authors declare no competing interests.









 Open access
Open access