


Palmer amaranth, a dioecious weed species native to the southwestern desert region of the United States, has been listed as one of the most troublesome weed species with rapid expansion throughout the midwestern and southern regions of the United States (Butts et al. Reference Butts, Norsworthy, Kruger, Sandell, Young, Steckel, Loux, Bradley, Conley, Stoltenberg and Arrriaga2016; Webster and Coble Reference Webster and Coble1997). Abundant seed production and high interference potential make this weed species a serious threat to crop production (Webster and Coble Reference Webster and Coble1997; Wiggins et al. Reference Wiggins, McClure, Hayes and Steckel2015). Most non-legume agronomic crops will regularly receive nitrogen fertilization to maximize yield. However, many studies have also shown that weed species take advantage of crop nitrogen fertilization by physiologically increasing seed production, germination rate, growth rate, and biomass partitioning (Sweeney et al. Reference Sweeney, Renner, Laboski and Davis2008; Wortman et al. Reference Wortman, Davis, Schutte and Lindquist2011). This is especially important in Palmer amaranth; previous studies have reported it as a species that is highly responsive to nitrogen fertilization (Ruf-Pachta et al. Reference Ruf-Pachta, Rule and Dille2013).
The control of Palmer amaranth populations has become a challenge due to the evolution of resistance to different herbicides, including triazine, acetolactate synthase (ALS) inhibitors, dinitroaniline, hydroxyphenylpyruvate dioxygenase inhibitors, and protoporphyrinogen oxidase–inhibiting herbicides, during the last two decades (Gaeddert et al. Reference Gaeddert, Peterson and Horak1997; Gossett et al. Reference Gossett, Murdock and Toler1992; Heap Reference Heap2017; Jhala et al. Reference Jhala, Sandell, Rana, Kruger and Knezevic2014; Ward et al. Reference Ward, Webster and Steckel2013). Palmer amaranth has also evolved resistance to glyphosate, the most widely used herbicide in agronomic crops in the United States. Although the first report of glyphosate-resistant (GR) Palmer amaranth originated in the state of Georgia (Culpepper et al. Reference Culpepper, Grey, Vencill, Kichler, Webster, Brown, York, Davis and Hanna2006), since 2008 there have been confirmed cases of GR populations in Alabama, Arkansas, Florida, Mississippi, Missouri, New Mexico, North Carolina, South Carolina, and Tennessee (Berger et al. Reference Berger, Ferrell, Dittmar and Leon2015; Heap Reference Heap2017; Mohseni-Moghadam et al. Reference Mohseni-Moghadam, Schroeder and Ashigh2013; Norsworthy et al. Reference Norsworthy, Griffith, Scott, Smith and Oliver2008).
Because of its dioecious reproductive system, Palmer amaranth has high genetic variability, which is predominantly found within populations and to a lesser extent among populations (Chandi et al. Reference Chandi, Milla-Lewis, Jordan, Burton, Zuleta, Whitaker and Culpepper2013b). In other words, differentiation among populations (measured with neutral molecular markers) seems to be low due to gene flow, but individuals within populations have marked differences in their genetic makeup. However, because selection forces in agricultural fields are considerably high, it is possible that key life-history traits might be under selection. If this selection increases weed survival, populations could eventually differentiate for these traits despite the high levels of gene flow. Ignoring such differentiation processes can result in increased weediness.
Bravo et al. (Reference Bravo, Leon, Ferrell, Mulvaney and Wood2017) reported differentiation of several life-history traits among 10 Palmer amaranth populations, and the differences seem to be related to particular characteristics of the cropping systems where the populations were found. Moreover, it was reported that GR populations produced more biomass and were taller than glyphosate-susceptible (GS) populations, although previous reports found no differences in fitness between GR and GS Palmer amaranth populations (Giacomini et al. Reference Giacomini, Westra and Ward2014; Vila-Aiub et al. Reference Vila-Aiub, Goh, Gaines, Han, Busi, Yu and Powles2014).
Considering that Palmer amaranth was adapted to low nitrogen availability, as a native species from desert regions (Ward et al. Reference Ward, Webster and Steckel2013), and that this weed is successfully colonizing agricultural systems with high nitrogen-fertilization inputs, we proposed that Palmer amaranth populations have evolved to take advantage of nitrogen fertilization to increase their growth and competitive ability within crops. Also, we proposed that adaptations to nitrogen fertilization might be related to growth differences between GR and GS populations (Bravo et al. Reference Bravo, Leon, Ferrell, Mulvaney and Wood2017). Therefore, we tested two hypotheses: (1) Palmer amaranth populations originating from cropping systems with high nitrogen-fertilization histories will modify growth more in response to nitrogen fertilization than populations from systems with low nitrogen-fertilization histories, and (2) GR Palmer amaranth populations have higher nitrogen-use efficiency than GS populations. This information is critical to assess how cultural practices not only affect Palmer amaranth success in agricultural fields but also influence GR population dynamics.
Materials and Methods
Seed Collection
The present study used the same Palmer amaranth populations reported by Bravo et al. (Reference Bravo, Leon, Ferrell, Mulvaney and Wood2017). Seeds from 10 populations of Palmer amaranth were collected in Florida and Georgia during the fall 2014 in coordination with local growers and county extension agents who had previously identified fields with a history of issues with this weed species. Field agricultural management history was considered in selecting sampling locations. Factors such as glyphosate sensitivity (determined by Bravo et al. Reference Bravo, Leon, Ferrell, Mulvaney and Wood2017 with dose–response studies), nitrogen-fertilization history, crop rotation, and crop canopy structure were the main selection criteria (Table 1). At every location, Palmer amaranth seeds were collected from plants found inside the crop in the first 25 m from field borders. Seeds were collected from 8 female plants (i.e., seeds from a single female plant were considered a family) within a 3-m radius in heavily infested areas (i.e., 0.5 to 15 females m−2), and sampling locations were at least 2.5 km apart. Once collected, the seeds were labeled and kept separated in paper bags until the initiation of the experiment.
Table 1 Origin and crop history of 10 Palmer amaranth populations.
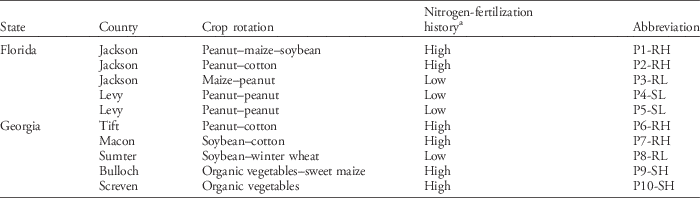
a Populations were classified as high (H) and low (L) according to nitrogen-fertilization history, with high and low receiving an average over 8 yr of >90 and <45 kgNha−1 yr−1, respectively. Populations were also classified as glyphosate resistant (R) and susceptible (S) based on Bravo et al. (Reference Bravo, Leon, Ferrell, Mulvaney and Wood2017).
Outdoor Pot Experiment
Outdoor pot experiments were conducted in the spring–summer season of 2015 at the West Florida Research and Education Center in Jay, FL. Palmer amaranth seeds were germinated on trays under greenhouse conditions. Once plants reached the 3-leaf stage, a single plant was transplanted into 10.7-L pots filled with the Ap horizon of a Red Bay sandy loam soil (Fine-loamy, kaolinitic, thermic Rhodic Kandiudult) containing 3.6 mgNkg−1, 1.6% organic matter, 69% sand, 16% silt, and 15% clay with pH 6 from the West Florida Research and Education Center in Jay, FL, and placed outdoors. Plants were watered on a daily basis with a drip-irrigation system to maintain soil field capacity. Although the soil did not have a natural Palmer amaranth seedbank, hand weeding was conducted as needed to avoid interference from other weed species. The experiment was arranged as a completely randomized design with six replications per treatment. Treatments were combinations of 10 populations and 2 fertilization treatments: 0 and 40 kgNha−1 (i.e., 0 and 1.3 g N pot−1, respectively, based on pot area), and the experiment was conducted twice. This fertilization rate was chosen after conducting preliminary studies. This amount of fertilizer allowed clear detection of fertilization responses without causing root or leaf burning due to overfertilization. The first experiment was initiated in late April and the second one in early June. Families were used as replications, so we could capture intrapopulation variability, but due to limited space, only six families were randomly selected from the eight families collected per population in the field. Ammonium nitrate was used as nitrogen source and was applied by burying it at 3 cm 7 d after transplanting.
Palmer amaranth response to nitrogen availability was evaluated when plants reached anthesis. At this point, multiple traits involved in plant architecture, growth rate, and nutrient-use efficiency (NUE) were measured to compare populations. These traits were chosen because they can influence crop interference, light interception, photosynthetic rate, and reproductive success. Traits included number of days to anthesis and greenness (Wood et al. Reference Wood, Reeves, Duffield and Edmisten1992) for each plant, averaging SPAD 502 Plus Chlorophyll Meter (Spectrum Technologies, Lincoln, NE) readings made on the 2nd, 3rd, and 4th fully developed leaves. Furthermore, to explore allometric variation in leaf and canopy morphology, the 2nd and 3rd fully developed leaves were measured for petiole and leaf-blade length. Also, leaf-blade width was measured at three equidistant points (i.e., tip, middle, base); the same approach was followed to determine canopy shape. Thus, plant height was measured, and then the canopy was divided into three sections along the main stem, and canopy width was determined at the top, middle, and base of the plant. After these evaluations, plants were clipped at soil level, and their fresh weight was determined. The apical inflorescence was clipped, and its length and weight were measured. Similarly, axillary inflorescences were removed from the plant, dried, and weighed separately. All leaves were clipped and counted, and total leaf area was determined with a LI-3100 Area Meter (Li-Cor Biosciences, Lincoln, NE).
Fresh weight of aboveground biomass was determined, and plants were dried, weighed, and ground. Total ground-tissue samples were sent to the University of Florida Institute of Food and Agricultural Sciences Analytical Services Laboratories (Gainesville, FL) for nutrient content determination. The estimation of total nitrogen in shoot tissue was performed using total Kjehldahl nitrogen (EPA 1993), and total phosphorus, potassium, magnesium, and zinc content was determined following the ICP emission spectroscopy methodology (EPA 1994).
Data Analysis
Because Bravo et al. (Reference Bravo, Leon, Ferrell, Mulvaney and Wood2017) already demonstrated differences in growth and morphology among the studied Palmer amaranth populations, the analysis of the present study was focused on changes in growth as a result of nitrogen fertilization. For this reason, all traits were converted to a relative response to nitrogen based on the values observed for nonfertilized plants. Therefore, responses of 1, <1, and >1 indicate no change, reduction, and increase under fertilized conditions compared with the nonfertilized control, respectively. Additionally, we quantified tissue content for several nutrients and estimated NUE (Cassman et al. Reference Cassman, Peng, Olk, Ladha, Reichardt, Dobermann and Singh1998; De Datta and Broadbent Reference De Datta and Broadbent1988):
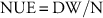 $${\rm NUE}\,{\equals}\,{\rm DW}/{\rm N}$$
$${\rm NUE}\,{\equals}\,{\rm DW}/{\rm N}$$
were DW represents dry weight (g), and N is nutrient content (mg).
The data were analyzed separately in four different (i.e., separate) ways: by sex (female vs. male); by population, by glyphosate sensitivity, and by nitrogen-fertilization history. For glyphosate sensitivity, populations were grouped as GR and GS as reported by Bravo et al. (Reference Bravo, Leon, Ferrell, Mulvaney and Wood2017). Also, populations were grouped based on nitrogen-fertilization history. Populations were classified as high nitrogen history if they received more than 90 kgNha−1 yr−1 from fertilizers during the Palmer amaranth growing season and as low-nitrogen history if they received less than 45 kgNha−1yr−1 from fertilizers during the last 8 yr. The results were subjected to ANOVA (α=0.05) using R (R Core Team 2014). Normality and homoscedasticity assumptions were checked using Q–Q plot and plotting the residuals versus predicted values. Data were square-root transformed for number of branches from main stem, 3rd leaf-tip width, and the ratio of the traits evaluated for the 2nd and 3rd leaves when needed to meet ANOVA assumptions, but nontransformed means are shown in the results. ANOVA models included treatment, experimental run, and their interaction as fixed effects. Mean separation was done using Tukey’s honestly significant difference (HSD; α=0.05). Nutrient content and NUE were evaluated using absolute values, including the fertilized and nonfertilized treatments, so the aforementioned ANOVAs also included the nitrogen-fertilization treatment effect and its interaction with the other factors. One-tailed t-tests (α=0.05) were used to compare different growth, morphology, nutrient content, and NUE between GR versus GS and high- versus low-nitrogen history populations after confirming the absence of significant interactions between experimental factors.
Results and Discussion
No interactions were detected among experimental runs and other effects (i.e., population, nitrogen-fertilization history, glyphosate sensitivity) in the ANOVA model (P≥0.73); consequently, data were pooled over the two experimental runs. Additionally, all populations and treatments had a 1:1 ratio of female:male plants, and no differences due to sex were observed for the different parameters (P>0.42). Therefore, further analyses were conducted pooling data from female and male plants.
Multiple life-history traits changed among populations in response to nitrogen fertilization (P<0.01). Traits such as plant height, foliar area, number of branches from the main stem, the 3rd leaf-tip and leaf-base widths, SPAD values, and allometric ratios of the 3rd leaf and canopy architecture exhibited significant differences among populations (Table 2). Overall, no single population exhibited an increase in all traits in response to nitrogen fertilization, so changes depended on specific trait and population combinations (Table 3). Branch number was the morphological trait that varied the most in response to nitrogen fertilization. For example, P7-RH had a 6-fold increment in branch number compared with other populations such as P10-SH, which had almost no change in branch number after nitrogen fertilization (Table 3). This increment in production of branches as a response to nitrogen fertilization could be advantageous when competing with a crop.
Table 2 ANOVA statistical significance of morphological and growth traits in response to nitrogen fertilization comparing 10 Palmer amaranth populations grouped by glyphosate sensitivity (resistant vs. susceptible) and by nitrogen-fertilization history (high vs. low).
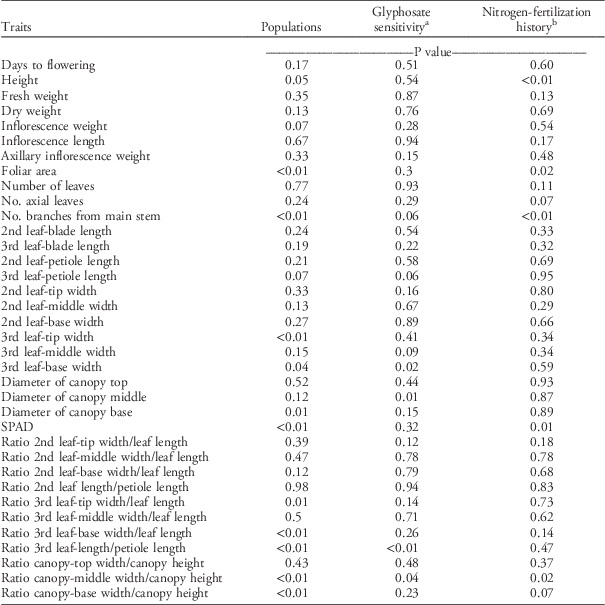
a Four populations were identified as susceptible (n=48) and six as resistant (n=72) to glyphosate.
b Six populations were identified as high (n=72) and four as low (n=48) nitrogen-fertilization history.
Table 3 Relative change in morphological and growth traits in response to nitrogen fertilization among 10 Palmer amaranth populations based on a nonfertilized control.Footnote a
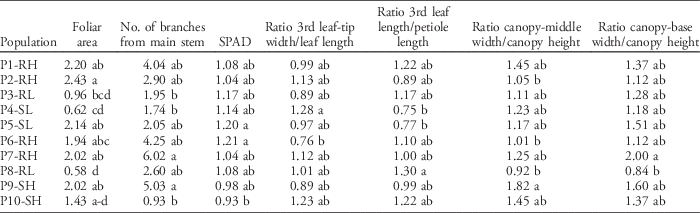
a Means within a column followed by the same letter are not different according to Tukey’s honestly significant difference (α=0.05). Values represent the pooled means of two independent experiments (n=12).
No changes in dry weight in response to the nitrogen fertilization were detected, but differences for height, foliar area, number of branches, and SPAD values varied depending on the nitrogen-fertilization histories of the cropping systems (Table 2). Populations with low nitrogen-fertilization histories were 30% taller than populations with high nitrogen-fertilization histories, but the foliar area was 30% higher for high-nitrogen populations in response to nitrogen fertilization than low-nitrogen populations (Figure 1). Populations with high nitrogen-fertilization histories increased branch number 85% and SPAD values 9% more than populations with low nitrogen-fertilization histories. The capacity of Palmer amaranth to take advantage of the extra nitrogen provided by fertilizers can be associated to its nitrophilous nature, previously reported by Ward et al. (Reference Ward, Webster and Steckel2013) and Ruf-Pachta et al. (Reference Ruf-Pachta, Rule and Dille2013).
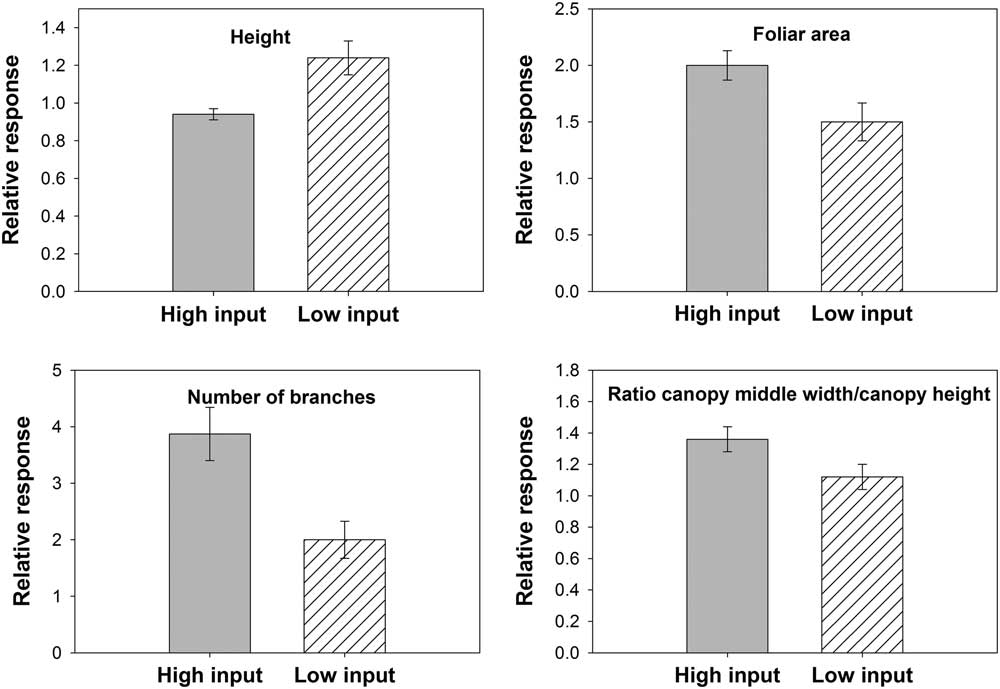
Figure 1 Relative change in morphological traits in response to nitrogen fertilization based on a nonfertilized control for Palmer amaranth populations with high and low nitrogen-fertilization histories. Six populations came from cropping systems with high nitrogen-fertilization histories (n=72) and four from low nitrogen-fertilization histories (n=48). Error bars represent 95% confidence intervals.
When populations were grouped based on glyphosate sensitivity, several differences in the response to nitrogen fertilization were detected for morphological traits. GR populations tended to have a wider base on the 3rd leaf, and the leaf blade was larger with respect to the petiole than GS populations (Figure 2). Furthermore, GR populations exhibited a 36% more-elongated canopy architecture than GS populations, and the latter presented a canopy 16% wider in the middle compared with the former (Figure 2).
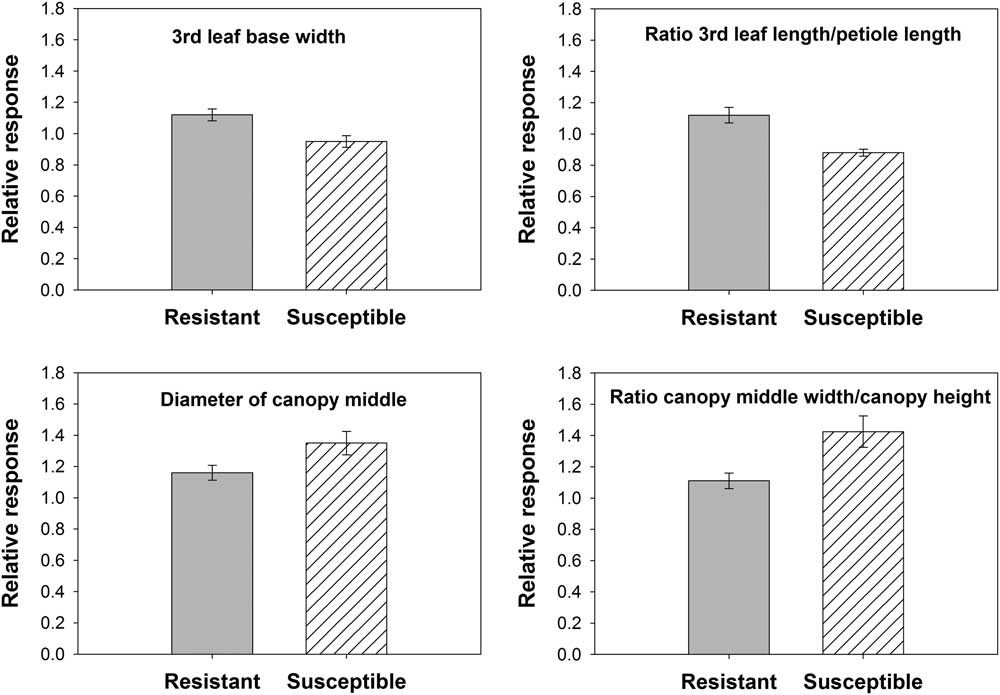
Figure 2 Relative change in morphological traits in response to nitrogen fertilization based on a nonfertilized control for glyphosate-resistant (GR) and glyphosate-susceptible (GS) Palmer amaranth populations. Six populations were identified as GR (n=72) and four as GS (n=48). Error bars represent 95% confidence intervals.
Considering these results, we investigated whether differences in morphology and growth might be related to differences in nutrient use, since changes in dry weight in response to nitrogen fertilization were not different among populations (Table 2).
No interactions between nitrogen-fertilization treatments and populations were observed for NUE (P>0.15), so data from fertilized and nonfertilized plants were pooled when comparing populations and groups based on glyphosate sensitivity and nitrogen-fertilization history. Palmer amaranth populations had a few differences in nutrient content (0.008≤P≤0.31), but there were clear differences in NUE (P≤0.007; Table 4).
Table 4 ANOVA statistical significance of differences among 10 Palmer amaranth populations for nutrient content and nutrient-use efficiency (NUE).Footnote a

a There were no interactions between population and nitrogen-fertilization treatments (P>0.15), so data of fertilized and nonfertilized plants were pooled for the analysis.
Similar to the response observed for morphology, high variation was observed among populations for nutrient content and NUE, but no trend was observed among populations, and only a few populations differed based on Tukey’s HSD (Table 5).
Table 5 Nutrient use efficiency (NUE) among 10 Palmer amaranth populations.
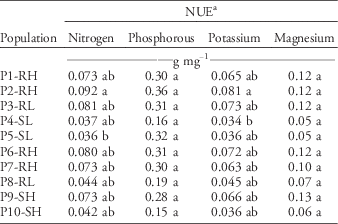
a Means within a column followed by the same letter are not different according to Tukey’s honestly significant difference (α=0.05). Values represent the pooled means of two independent experiments (n=24). There were no interactions between population and nitrogen-fertilization treatments for NUE (P>0.15), so data of fertilized and nonfertilized plants were pooled for the analysis.
No differences in nutrient content were observed when comparing populations with different nitrogen-fertilization histories (P>0.12), but clear differences in NUE were found between populations with high and low nitrogen-input histories (Table 6). In general, populations with high nitrogen-fertilization histories exhibited 43% to 59% higher NUE than populations with low nitrogen-fertilization histories, depending on the nutrient. Also, GR populations had at least 47% higher relative NUE than GS populations (Table 7). The differences in NUE were partially explained by GR populations maintaining lower nitrogen, phosphorus, and magnesium content than GS populations (Table 7) without sacrificing growth.
Table 6 Nutrient use efficiency (NUE) among Palmer amaranth populations with a history of high and low nitrogen fertilization.
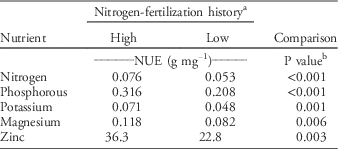
a Populations were classified according to nitrogen-fertilization history with high and low receiving an average over 8 yr of >90 and <45 kgNha−1 yr−1, respectively.
b Within rows and response trait, P values compare high- and low-nitrogen history groups based on a one-tailed t-test (α=0.05). Six populations (24 replications per each population) were identified as high (n=144) and four as low (n=96). There were no interactions between nitrogen history and nitrogen-fertilization treatments for NUE (P>0.15), so data of fertilized and nonfertilized plants were pooled for the analysis.
Table 7 Nutrient content and nutrient-use efficiency (NUE) among glyphosate-resistant (GR) and glyphosate-susceptible (GS) Palmer amaranth populations.Footnote a
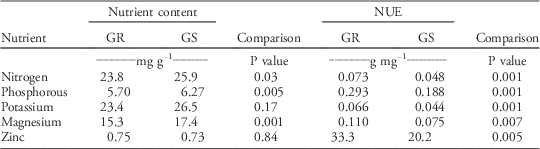
a Within rows and response trait, P values compare resistant and susceptible groups based on a one-tailed t-test (α=0.05). Six populations (24 replications per each population) were identified as GR (n=144) and four as GS (n=96). There were no interactions between glyphosate sensitivity and nitrogen-fertilization treatments (P>0.32), so data of fertilized and nonfertilized plants were pooled for the analysis.
Our results showed that populations collected from systems with high nitrogen-fertilization histories have adapted to take advantage of increased nitrogen availability provided by fertilizer inputs, changing their morphology by allocating resources to the production of branches, foliar area, reproductive tissue, and wider leaves, and ultimately increasing NUE. However, since no single population exhibited consistent changes across all evaluated traits in response to nitrogen fertilization, we think that a combination of cropping-system characteristics and not only nitrogen-input history are responsible for the differences observed among populations. Nitrogen fertilization can also modify crop–weed interactions (Blackshaw and Brandt Reference Blackshaw and Brandt2008), so the response of the crop and the structure of the weed community could have influenced the importance of nitrogen-fertilization history on Palmer amaranth populations. Harbur and Owen (Reference Harbur and Owen2004) found that nitrogen response of competitive weed species is affected by photosynthetically active radiation and the maximum potential growth rate of the plant, factors that vary greatly depending on crop canopy. Therefore, it would be beneficial for Palmer amaranth plants to take advantage of nitrogen fertilization to optimize canopy structure and maximize light interception under crop competition (Barnes et al. Reference Barnes, Flint and Caldwell1990). However, the changes in canopy structure will also depend on the characteristics of the canopy of the cropping system. Bravo et al. (Reference Bravo, Leon, Ferrell, Mulvaney and Wood2017) found that crop species and canopy height explained differences in growth and morphological traits among Palmer amaranth populations. The present study underlines the importance of fertilization as a key component of integrated weed management strategies not only to avoid favoring weeds over crops in competition interactions (Ampong-Nyarko and De Datta Reference Ampong-Nyarko and De Datta1993), but also to prevent evolutionary adaptations increasing weediness.
Morphological differences related to herbicide resistance have been previously reported for other weed species (Hall and Romano Reference Hall and Romano1995; Kumar and Jha Reference Kumar and Jha2016), which in the case of Palmer amaranth populations indicate a possible competitive advantage for GR individuals. Since resistant individuals can survive herbicide applications, competition with the crop becomes an important selection force. For instance, the more-elongated canopy, with branching patterns that separate leaves, reducing “self-shading,” enables GR populations to maximize light interception and better compete against the crop canopy (Roig-Villanova and Martinez-Garcia Reference Roig-Villanova and Martinez-Garcia2016).
Our data suggest that the increased growth in GR Palmer amaranth populations compared with GS populations (Bravo et al. Reference Bravo, Leon, Ferrell, Mulvaney and Wood2017) might be partially due to adaptations in metabolic processes favoring higher NUE. These adaptations could have occurred before or after GR trait evolution and spread, and the explanation of why the GR and NUE traits seem to be related in the present study remains elusive. Although previous studies have suggested no fitness differences between GR and GS Palmer amaranth populations (Chandi et al. Reference Chandi, Jordan, York, Milla-Lewis, Burton, Culpepper and Whitaker2013a; Giacomini et al. Reference Giacomini, Westra and Ward2014; Vila-Aiub et al. Reference Vila-Aiub, Goh, Gaines, Han, Busi, Yu and Powles2014), the clear differences in growth reported by Bravo et al. (Reference Bravo, Leon, Ferrell, Mulvaney and Wood2017) indicate that GR populations might be capable of evolving greater growth potential than GS populations depending on selection pressures exerted by the cropping system and its production practices.
Once resistance is confirmed, researchers explore the possibility that the resistant trait also causes fitness penalties (Chandi et al. Reference Chandi, Jordan, York, Milla-Lewis, Burton, Culpepper and Whitaker2013a; Giacomini et al. Reference Giacomini, Westra and Ward2014). These are efforts that have a physiological goal, which is to determine whether the mutation has pleotropic effects on the growth of the plant. It is frequently proposed that if the mutation has a fitness penalty, stopping the use of the herbicide in question eliminates the advantage that herbicide-resistant individuals have, which ultimately will allow susceptible individuals (without the fitness penalty) to become predominant again (Diggle et al. Reference Diggle, Neve and Smith2003; Gressel and Segel Reference Gressel and Segel1978). Unless the fitness penalty is extremely severe, it is possible that through the appearance of mutations or gene flow via cross-pollination, individuals carrying the herbicide-resistance trait and suffering the fitness penalty associated with it can acquire modifications in metabolism that might be able to compensate for the aforementioned penalty. Although there is no fitness penalty associated with glyphosate resistance, the present study provides evidence of this last scenario by documenting what seems to be a gain in fitness. The fact that GR populations had bigger individuals than GS populations (Bravo et al. Reference Bravo, Leon, Ferrell, Mulvaney and Wood2017) and that GR populations were able to evolve higher NUE than GS populations clearly indicates that genetic changes in the metabolism of the plant can mitigate for fitness penalties associated with herbicide resistance and even increase the fitness of resistant populations. GR populations collected in Georgia came from areas where EPSPS gene amplification was confirmed (Culpepper et al. Reference Culpepper, Grey, Vencill, Kichler, Webster, Brown, York, Davis and Hanna2006; Gaines et al. Reference Gaines, Zhang, Wang, Bukun, Chisholm, Shaner, Nissen, Patzoldt, Tranel, Culpepper and Grey2010), and Florida GR populations are thought to have been introduced from Georgia. It is possible that the mechanism of gene amplification created genetic variability favoring adaptation. In fact, Molin et al. (Reference Molin, Wright, Lawton-Rauh and Saski2017) described changes in genomic content and structure in Palmer amaranth genome regions containing EPSPS gene amplification (i.e., EPSPS cassette). Some of the putative genes flanking the EPSPS cassette had functions related to stress responses and DNA replication and mobility. Furthermore, GR populations in the southeastern United States have been reported having multiple resistance, especially to ALS-inhibiting herbicides (Berger et al. Reference Berger, Ferrell, Dittmar and Leon2015), so selection forces affecting adaptive traits could have influenced GR populations even when herbicides with other mechanisms of action were used. Although understanding secondary effects of herbicide-resistance traits is important, we must not ignore the fact that evolutionary adaptations might occur rapidly in agroecosystems, and weeds might be able to evolve compensatory mechanisms to disadvantages generated when herbicide resistance first evolves. Darmency et al. (Reference Darmency, Menchari, Le Corre and Délye2015) documented differences in the genetic background of a resistant biotype carrying a Gly-2078 mutation compared with other mutations conferring resistance to acetyl-coenzyme A-carboxylase–inhibiting herbicides in blackgrass (Alopecurus myosuroides Huds.). These researchers proposed that rapid evolution in the genetic background of the Gly-2078 biotype allowed the plant to compensate for the fitness penalty associated with this mutation. Therefore, to properly assess the impact of a herbicide-resistance trait on the fitness of a weed, it is important to assess genetic diversity within and across populations and the ecological context in which those populations exist.
It is not clear why previous studies did not find differences in fitness between GR and GS biotypes, but it is possible that those studies used Palmer amaranth from similar cropping systems (although this is not specified in the articles). Another possibility is that seed collection in those studies was done when glyphosate resistance was just starting, and our study is identifying adaptations in other life-history traits that occurred later. If this is the case, Palmer amaranth exhibits rapid evolution under selection pressure in row-crop systems.
Conclusions
The results of the present study partially confirmed our hypotheses that nitrogen-fertilization history has modified Palmer amaranth populations’ growth and NUE, and that GR Palmer amaranth populations can produce more biomass than GS populations in response to nitrogen fertilization. However, our study also suggests that a combination of selection forces specific to each cropping system might be responsible for differentiation among Palmer amaranth populations in response to nitrogen fertilization. Additionally, GR populations seem to have either coevolved or acquired higher NUE, which could partially explain the increased growth reported by Bravo et al. (Reference Bravo, Leon, Ferrell, Mulvaney and Wood2017) in GR compared with GS populations. These results highlight the need to study the competitive interactions of Palmer amaranth in different cropping systems in more detail, incorporating evolutionary considerations, since selection forces such as nitrogen availability and herbicide resistance might be interacting and increasing the weediness and success of this aggressive weed species in multiple cropping systems.
Acknowledgments
We thank Ted Webster, Carroll Johnson, Tim Grey, Josh Thompson, and William Starr for helpful guidance and assistance during seed collection. We also thank Sharon Howell, Michael Dozier, and Rocio van der Laat for technical support. This research was partially funded by the Institute of Food and Agricultural Sciences, University of Florida.










 Open access
Open access