


Introduction
The earliest putative corals occur in the early Cambrian. Although many of these generally coralline forms (e.g., the Coralomorpha of Jell, Reference Jell1984) are of uncertain affinities, among them are undoubted corals (Scrutton, Reference Scrutton1997, Reference Scrutton1999). The oldest definite tabulate coral, Lichenaria Winchell and Schuchert, Reference Winchell and Schuchert1895, occurs in the Early Ordovician (Scrutton Reference Scrutton1979, Reference Scrutton1999; Webby et al., Reference Webby, Elias, Young, Neuman and Kaljo2004). The scarcity of coralomorphs reported from the Cambrian and Early Ordovician contrasts with the diversity of undoubted coral genera recorded after the Middle Ordovician. In this context any finding of coral-like forms from this critical interval significantly contributes to the global diversity, biogeography, and early evolutionary knowledge of corals and coral-like organisms.
Previous works on Paleozoic corals in the Argentine Precordillera (Rodriguez et al., Reference Rodríguez, Carrera and Fernández-Martínez2002; Fernandez-Martinez et al., Reference Fernandez-Martinez, Carrera and Rodriguez2004; Carrera et al., Reference Carrera, Montoya, Rustan and Halpern2013, and references therein) show records of paleofavositids and sarcinulids from the Middle–Upper Ordovician units, and favositids, pleurodictyids, and rugosans from Silurian and Lower Devonian rocks. The aim of this contribution is to report and describe a tabulate-like coralomorph forming part of boundstones developed within the uppermost Cambrian–lowermost Ordovician that were found in the lower section of the La Silla Formation, Precordillera, western Argentina. This finding represents the oldest record of the genus Amsassia Sokolov and Mironova, Reference Sokolov and Mironova1959. Its association with boundstones and reef-mounds represents the earliest occurrence of this reef-related metazoan found in the gap of reef metazoan constructors after the demise of archaeocyaths in the late early Cambrian. The paleobiogeographic distribution and significance of this new record are discussed.
Geological setting
The new tabulate-like coralomorph is included within shallow-water subtidal limestones of the La Silla Formation in the Argentine Precordillera (Fig. 1). The La Silla Formation (Keller et al., Reference Keller, Cañas, Lehnert and Vaccari1994) is considered to be an ancient tropical Bahamian-type platform (Cañas, Reference Cañas1999; Keller, Reference Keller1999; Pratt et al., Reference Pratt, Raviolo and Bordonaro2012) that accumulated on a uniformly subsiding passive margin within the exotic Precordillera terrane (Astini et al., Reference Astini, Benedetto and Vaccari1995; Gomez and Astini, Reference Gomez and Astini2015). Paleogeographically, by this time, the Precordillera terrane was drifting within the southern Iapetus Ocean (Astini et al., Reference Astini, Benedetto and Vaccari1995; Benedetto, Reference Benedetto2004; Keller, Reference Keller2012). From a stratigraphic viewpoint, it represents the transition from the late Cambrian largely fossil-barren cyclic peritidal dolomites (Zonda and La Flecha formations) into fossiliferous open-shelf pure limestones of the Ordovician San Juan Formation (Fig. 1.2). However, the high-frequency stacking pattern in the La Silla Formation is more similar to the underlying peritidal La Flecha Formation than to the overlying dominantly subtidal San Juan Formation.

Figure 1 (1) Location map of the Precordillera, showing the major two-fold division in eastern and western tectofacies (Astini, Reference Astini1992) and the location of the section under study in the Cerro La Silla, type area for the La Silla Formation. The eastern tectofacies are represented by the largely continuous Cambrian to Ordovician passive-margin carbonate platform and overlying black shales and clastic facies that crop out in the central and eastern Precordillera. The western tectofacies is a thick clastic wedge and related mafic volcanic rocks of Middle and Late Ordovician age and mostly represented in the western Precordillera region. (2) Chart of the general stratigraphy of the Precordillera carbonate platform and the evolutive stages for the Cambrian–Ordovician interval. Colored area highlights the unit considered in this study.
Although there is a clear faunal turnover starting from the La Silla Formation onwards (Cañas and Carrera, Reference Cañas and Carrera2003), the major change to truly open-marine settings seems to occur at the boundary between the La Silla and San Juan formations (Cañas, Reference Cañas1999; Buggisch et al., Reference Buggisch, Keller and Lenhert2003; Thompson et al., Reference Thompson, Kah, Astini, Bowring and Buchwaldt2012). Major type 1 sequence boundaries separate the La Silla Formation from the under- and overlying units (Cañas, Reference Cañas1999; Keller, Reference Keller1999). Accommodation within this largely detached passive-margin setting is interpreted to relate to global sea-level fluctuations rather than with local subsidence (see Astini and Thomas, Reference Astini and Thomas1999; Thomas and Astini, Reference Thomas and Astini1999).
The La Silla Formation is exposed in several thrust sheets over a distance of more than 350 km along the Precordillera fold and thrust belt in the San Juan Province of western Argentina (Fig. 1). Its massive appearance is in part due to pervasive recrystallization that affects the entire unit. It is chiefly composed of medium- to thick-bedded peloidal-intraclastic-ooidal limestones and thrombolitic biostromes (Cañas and Carrera, Reference Cañas and Carrera2003), with subordinate amounts of lime mudstones, laminated stromatolitic dolostones, and rare chert. Within its 325 m of thickness (Fig. 2), three major lower-rank cycles have been recognized (Keller, Reference Keller1999, Reference Keller2012; Raviolo et al., Reference Raviolo, Bordonaro and Pratt2007), each containing higher-frequency meter-scale cycles that record upward-shallowing trends.
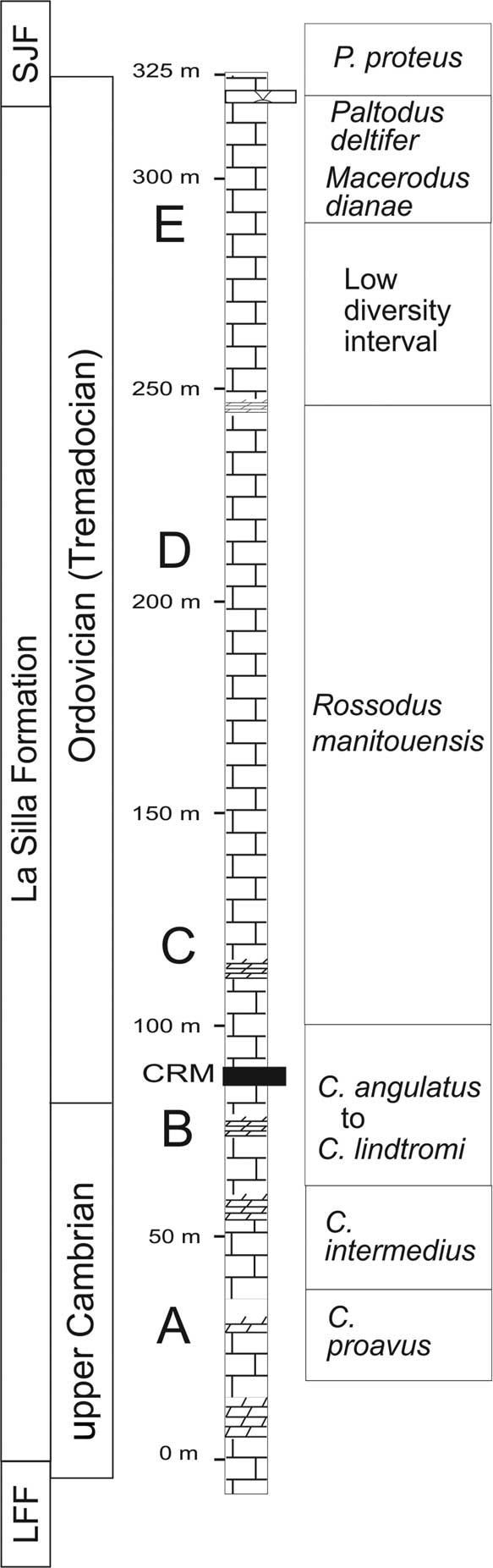
Figure 2 General stratigraphy and conodont biostratigraphy (from Buggish et al., 2003) of the La Silla Formation. CRM: stratigraphic position of the coralomorph reef-mounds; SJF: San Juan Formation; LFF: La Flecha Formation; A-E: main section points for conodont biozones.
The dominant lithologies in the La Silla Formation (Fig. 3) are peloidal and intraclastic grainstones and packstones, and fewer peloidal wackestones, all of which have been pervasively recrystallized and affected by neomorphism. Pressure-solution seams and stylolites commonly enhance primary facies contacts and bedding pattern. Silicified Thalassinoides paradoxicus (Woodward, Reference Woodward1830) are exceptionally preserved in some beds (Mángano and Buatois, Reference Mángano and Buatois2003), mostly within the middle and upper section. Facies associations in the La Silla Formation represent persistently high-energy shallow subtidal marine conditions. However, different platform-interior and shelf-lagoonal to tidal-flat subenvironments have been recognized (Cañas, Reference Cañas1999; Keller, Reference Keller1999; Pratt et al., Reference Pratt, Raviolo and Bordonaro2012) within the three major subdivisions.

Figure 3 Stratigraphy and facies of the Early Ordovician La Silla Formation, Argentine Precordillera, including location of the coralomorph reef-mound interval (arrow).
Stromatolites, and particularly thrombolites, are ubiquitous and very common components of the higher-frequency cycles (Armella, Reference Armella1994; Cañas and Carrera, Reference Cañas and Carrera2003). Thrombolites vary in diameter from meter scale to several meters, and can be isolated, coalescent, distributed in clusters or patches, and forming laterally persistent biostromes remarkably linked to shallow subtidal high-energy grainy facies (intraclastic to peloidal grainstones). Calcarenites are mainly composed of peloids, well-rounded intraclasts, aggregate lime grains, micritized ooids, and a few bioclasts. Bioclasts are very scarce and include rare calcareous algae such as Nuia sp. and scattered fragments of gastropods, trilobites, and nautiloids within the cross-bedded subtidal grainstone-packstone intervals. Abundant in situ gastropods seem intimately related with domical thrombolites also are common components in associated storm layers and within intercolumnar spaces between domical thrombolites.
Stratigraphic position and age
The tabulate-like form is included in patchy, mud-rich microbial mounds that grew immediately above the major flooding surface that initiated the second accommodation-related shallowing-upward cycle, ~95 m above the base of the La Silla Formation (Fig. 2). The fossiliferous level is within the Cordylodus angulatus Conodont Zone (lower Tremadocian), slightly above the Cambrian-Ordovician boundary, indicated by the presence of the Clavohamulus hintzei and Cordylodus lindstromi zones (Buggisch et al., Reference Buggisch, Keller and Lenhert2003).
The age of the La Silla Formation has been established by combining information from conodonts and sparse trilobites (Lehnert and Keller, Reference Lehnert and Keller1993; Keller et al., Reference Keller, Cañas, Lehnert and Vaccari1994; Lehnert, Reference Lehnert1995; Lehnert et al., Reference Lehnert, Miller and Repetski1997; Albanesi and Ortega, Reference Albanesi and Ortega2002). The trilobite Plethopeltis obtusus (Rasetti, Reference Rasetti1945), which in North America ranges from the Saukia to the Missisquoia trilobite zones, has been found near the base of the unit (Vaccari, Reference Vaccari1994), indicating a late Sunwaptan (late Furongian) age for this part of the section. Four conodont associations were described from intervals B through E within the La Silla Formation (Lehnert, Reference Lehnert1995; Buggish et al., Reference Buggisch, Keller and Lenhert2003).
The oldest recorded conodont fauna of the unit occurs ~55 m from its base (point B in the section, Fig. 2), containing the species Clavohamulus hintzei Miller, Reference Miller1969, which is diagnostic of the Clavohamulus hintzei Conodont Subzone of the Cordylodus intermedius Zone. The next conodont fauna (point C in the section, Fig. 2) was correlated with the Rossodus manitouensis Conodont Zone of the Skullrockian Stage (early Tremadocian) (Ibexian Series) of North America. Point D conodonts were correlated with the ‘Low Diversity Interval’ and with the lower Macerodus dianae Conodont Zone in North America, whereas point E near the top of the section is correlated with the Paltodus deltifer Conodont Zone, indicating a late Tremadocian age for the top of the La Silla Formation. According to this scheme, the base of the Tremadocian Stage (and the Cambrian-Ordovician boundary) lies in the section between points B and C at the Cerro La Silla type section, where our fauna was collected (Fig. 2). A detailed carbon isotope record (controlled by conodont assemblages) of the La Silla and San Juan formations was published by Buggisch et al. (Reference Buggisch, Keller and Lenhert2003), presenting an intercontinental correlation of the δ13Ccarb curve with biostratigraphically well-constrained records of the U.S.A. and Australia. The close match of the curves enabled the authors to place the Cambrian-Ordovician boundary between points B and C, ~80–90 m above the base of the La Silla Formation and immediately below our faunas.
Hence, the material under study may come from the lowermost Ordovician levels of the La Silla Formation, belonging to the Cordylodus angulatus Conodont Zone (lower Tremadocian), ~20 m above the Cambrian-Ordovician boundary. These levels probably correlate in time with those of the St. George Group in Newfoundland, but could be slightly older than the thrombolites-Lichenaria mud-mounds of Pratt and James (Reference Pratt and James1982).
Reef-mound interval
Within the La Silla Formation, benthic biota seems both sparse and low diversity in composition, especially in lacking pelmatozoan ossicles, as Pratt et al. (Reference Pratt, Raviolo and Bordonaro2012) have pointed out. However, complex microbial consortiums forming thrombolites, the new coralomorph described herein, and calcareous algae such as Nuia sp. begin to occur in these limestones at the lower interval of the unit (uppermost Cambrian–lowermost Ordovician).
The coralomorphs are restricted to a 2m thick massive, matrix-rich interval (Fig. 3), where they seem to be in life position. The outcrop is partially covered by debris and vegetation, making it difficult to precisely track the coralomorph-bearing boundstone interval laterally. However, this interval completely disappears laterally at a scale of tens of meters, suggesting low-relief mound geometry.
The individual coralomorphs may be grouped in low-relief (up to 10 cm) radial clusters or alternatively may be isolated crusts and slightly broken fragments embedded in peloidal micrograinstones that are intimately associated with various microbial communities. In polished slabs (Fig. 4), radial clusters seem to be passively covered by laminated peloidal muds or abruptly truncated by rugged surfaces capped by peloidal grainstones (Fig. 5). Within the grainstones, the calcareous alga Nuia sp. is present (Fig. 5.3). Radial clusters seem to be nucleated on the peloidal intraclastic grainstones (hardgrounds) or directly stacked on small-diameter domical microbial heads. Intercolumnar spaces between coral clusters are alternatively filled by peloidal muds, peloidal grainstones, or complex calcified microbial consortiums (Fig. 5). This allows for the interpretation that colonial growth occurred synchronous with energy fluctuations and somehow colonies were interacting with microbial communities and periodically interrupted by higher-energy events, developing erosion surfaces, and grainy facies. Microscopic observations in thin sections seldom show terminal module walls protruding out of these irregular surfaces, indicating that not all of these surfaces are strictly erosive, but depositional. Irregular micritic to micropeloidal laminae, occasionally with filamentous textures, point to a stromatolite-like habit and trapping and binding processes. Microscopically clotted-peloidal dense micrite with diverse shapes and growth patterns suggest pervasive bacterially induced precipitation within microbial biofilms, as well as cryptic microbialites (Chafetz, Reference Chafetz1986; Reitner, Reference Reitner1993; Riding, Reference Riding2002; Adachi et al., Reference Adachi, Ezaki and Liu2004; Flügel, Reference Flügel2004; Chen and Lee, Reference Chen and Lee2014). As pointed out in other examples (e.g., Sun and Wright, Reference Sun and Wright1989; Riding and Tomas, Reference Riding and Tomas2006), microbial textures occur both as irregularly laminated crusts on framework elements, as complex open-cavity fills within the boundstone framework, and as internal fills in cavities (intraskeletal and boring). Among distinct cavity-filling microbial consortiums, we recognized Renalcis-like chamber arrays (Fig. 5.1) (e.g., Riding, Reference Riding1991; Chafetz and Guidry, Reference Chafetz and Guidry1999; Riding and Fan, Reference Riding and Fan2001; Stephens and Sumner, Reference Stephens and Sumner2002), as well as dendritic forms (Fig. 5.5) (e.g., Pratt and James, Reference Pratt and James1982; Pratt, Reference Pratt1984; Riding, Reference Riding1991; Shen et al., Reference Shen, Yu and Bao1997) and clustered globous forms (Fig. 5.4). Internal fills have a patchy intraskeletal clotted homogeneous pattern (Figs. 5, 6) and may correspond to later eodiagenetic non-photosynthetic cryptic bacteria that grew within available pore spaces, largely represented by decaying of coralomorph colonies.

Figure 4 Vertically oriented polished rock slab (CEGH-UNC 26127) from the studied carbonate reef-mounds in La Silla Formation (Lower Ordovician) showing details of main components and their distribution. C: coralomorph (Amsassia colonies, see Fig. 6); S: stromatolite crusts (see Fig. 5.6); T: thrombolite microbialites (see Fig. 5.1, 5.5). Note intimate scaffold development and voids (V) preserved within framework. Also note stylolite development throughout the sample.

Figure 5 Thin section photomicrographs and boundstone petrography showing textures and distinct microbial components, including Amsassia argentina n. sp. colonies; scale bars=1 mm, except in Fig. 5.3: (1) Renalcis-like chambers (white arrows) isolated and forming arrays within the microbial consortium growing towards center of cavity (filled with sparite); (2) view of the colony in sharp contact with a peloidal grainstone through a stylolite (white arrow); (3) peloidal grainstone overlying a cemented erosive surface (hardground). Note oblique section of Nuia sp. (black arrow) within grainstone, scale bar=0.5 mm; (4) detail of termination of a colony showing a complex microbial consortium with chambered, dentritic, filamentous, and globous forms growing toward a framework reef cavity; (5) similar to 4. Note the contrast between cryptic microbial communities (represented by homogeneous clotted peloidal automicrite) growing within modules (black arrow) and those more oriented forms colonizing the surface (white arrow); (6) finely laminated peloidal micrograinstone (stromatolite crust, black arrow) draping the termination of a colony. Note trilobite section (white arrow) and other bioclasts within upper part of photograph and some infiltration within the upper modules of the underlying colony. 1, 2, 4, 5 and 6 show partial views of longitudinal to transverse sections of Amsassia colonies. Note that in most cases, individual modules are partly occluded by homogeneous clotted automicrite of bacterial origin.

Figure 6 Thin section photomicrographs of Amsassia argentina n. sp.: (1) a colony in longitudinal section CEGH-UNC 26120, scale bar=5 mm; (2) complete view of the transversal section of holotype CEGH-UNC 26119, scale bar=5 mm; (3) transversal section of a colony showing the conspicuous infilling of peloidal grains, CEGH-UNC 26121, scale bar=2 mm; (4) close up of holotype CEGH-UNC 26119 showing arrangements of the polygonal modules and diagenetically dissolved areas, scale bar=1 mm; (5) close up of Fig. 6.4 showing module wall with a thick outer wall on both sides and fibrous structures perpendicular to the median line, scale bar=0.5 mm; (6) transverse section showing wall discontinuities and the ceroid to phaceloid structure, CEGH-UNC 26122, scale bar=1 mm; (7) longitudinal section of a colony showing long slender and slightly sinuousmodules, without (or missing) tabulae, with axial longitudinal increase (arrow), CEGH-UNC 26123, scale bar=2 mm; (8, 9) longitudinal views of modules with regularly distributed tabulae, CEGH-UNC 26124, both scale bars=1 mm; (10) longitudinal section showing the flexuous development of modules and scarcity of tabulae, CEGH-UNC 26125, scale bar=5 mm; (11) longitudinal section in holotype CEGH-UNC 26119 showing development of longitudinal increase (arrow), scale bar=2 mm; (12) transversal section of holotype CEGH-UNC 26119 showing polygonal modules, scale bar=1 mm.
Most of the microfacies, including the microbial peloidal-clotted matrix, the grainstones, and the coralomorph clusters themselves, are sparsely bioturbated. Field, slab-, and thin-section analysis is consistent with a combination of processes typical of microbial boundstones and microbial-coralomorph boundstones. Due to their relative abundance, the complex microbial consortium, together with the coralomorphs, seems to represent the major framework builders in our case. During their growth and metabolic activities, microbes and cyanobacteria may have induced calcite precipitation in cyanobacterial filaments, algal sheaths, and extra-polymeric substances (EPS) within the boundstone and/or periodically helped trap and bind lime peloidal muds that in some cases seem to drape coralomorph colonies (Fig. 4). Together with early cementation, microbial activity may have contributed to substrate stabilization, encrustation, and development of the metazoan (coralomorph) skeletal frameworks. It is therefore evident that microbes may have played roles as binders and substrate stabilizers, further allowing encrustation by coralomorphs. It is also clear that erosion and fragmentation periodically affected these low-relief reef-mounds, as suggested by the presence of irregular erosive surfaces (Fig. 5.3) and interbedded peloidal intraclastic grainstones with fragmented and disrupted colonies.
Repository and institutional abbreviation
The specimens are housed under the prefix CEGH-UNC in the paleontological collection of Centro de Investigaciones Paleobiológicas (CIPAL), at the building of Centro de Investigaciones en Ciencias de la Tierra (CICTERRA), Universidad Nacional de Córdoba, Argentina.
Systematic paleontology
“Coralomorpha” (Jell, Reference Jell1984; Scrutton, Reference Scrutton1997)
Remarks
The calcified modular skeletons of several organisms have similarities with coralline organization. Most of these forms were recorded in the Cambrian (e.g., Rosellatana Kobluk, Reference Kobluk1984; Flindersipora Lafuste et al., Reference Lafuste, Debrenne, Gandin and Gravestock1991; Moorowipora Fuller and Jenkins, Reference Fuller and Jenkins1994 ; Tabulaconus Handfield, Reference Handfield1969; Harklessia Hicks, Reference Hicks2006). All these coral-like forms were grouped under the informal name Coralomorpha, or the more formal Tabulaconida Scrutton, Reference Scrutton1997 for tabulate-like forms (Jell, Reference Jell1984; Debrenne et al., Reference Debrenne, Gangloff and Lafuste1987; Zhuravlev et al., Reference Zhuravlev, Debrenne and Lafuste1993; Scrutton, Reference Scrutton1997). Tabulata Milne-Edwards and Haime, Reference Milne-Edwards and Haime1850 is now regarded as a cohesive group (Scrutton, Reference Scrutton1979, Reference Scrutton1984; Hill, Reference Hill1981; Pandolfi, Reference Pandolfi1989), with Lichenaria as the oldest ancestral genus. However, some uncertainties as to the taxonomic position still exist among the stock of early basal coralomorphs and tabulate-like forms recorded from the lower Cambrian to the Lower Ordovician.
We use the term Coralomorpha as an informal name with no phylogenetic implications. The genus Amsassia is provisionally included in this morphologic group after the comprehensive reevaluation performed by Sun et al. (Reference Sun, Elias and Lee2014) of this problematic organism with uncertain affinities.
Genus Amsassia Sokolov and Mironova, Reference Sokolov and Mironova1959
Type species
Amsassia raduguini Mironova in Sokolov and Mironova, Reference Sokolov and Mironova1959 from the Amsass Suite, Upper Ordovician of Shoria Mountains, western Siberia.
Amsassia argentina new species
Holotype and other material
Holotype CEGH-UNC 26119. Other material: CEGH-UNC 26120-26127 (8 complete and fragmentary colonies in thin sections and polished slabs). All specimens are from the base of the La Silla Formation, Lower Ordovician, Cordylodus angulatus Conodont Zone (lower Tremadocian), San Juan Province, western Argentina.
Diagnosis
Growth form massive, tabular, domical to foliaceous or irregular. Structure mainly cerioid to phacelocerioid in some areas. Transverse shape of modules rounded to subpolygonal in loosely packed areas (module diameter 0.4–0.8 mm) to polygonal in densely packed areas (module diameter 0.5–1.1 mm). Module wall fibrous, with median line between adjoining modules. Module increase by longitudinal fission. Tabulae rare, irregularly spaced, bowed downward.
Description
Growth form massive, tabular, domical to foliaceous or irregular. Structure mainly cerioid to phacelocerioid; size up to 170 mm across, 70 mm high. In transverse section, modules generally polygonal to subpolygonal and closely packed, ranging from quadrate through circular to irregular in loosely packed areas. Maximum diameter 0.5–1.1 mm in areas of polygonal modules, 0.4–0.8 mm in areas of subpolygonal to rounded modules (Table 1). In longitudinal section, module walls relatively straight to gently undulate. Tabulae rare, very thin, complete or partially aborted, usually concave. Incomplete tabulae short and thick, or long with sharp tips pointed downwards (Fig. 6.8). When tabulae present, commonly bowed (downward), most of them complete, <0.01 mm thick and irregularly spaced, ~0.8–2 mm apart.
Table 1 Module diameter of Amsassia argentina n. sp. For seven specimens from La Silla Formation, two distinct areas within a transverse thin section were sampled, characterized by polygonal and subpolygonal to rounded modules. Polygonal module diameter=(long axis + short axis)/2. Abbreviations: Max., maximum diameter (mm); Min., minimum diameter (mm); Avg., average diameter (mm); SD, standard deviation; n, number of modules.
Pores or wall discontinuities related with dissolution or with different types of longitudinal fission. Module increase by longitudinal fission, in most cases bipartite, but tripartite observed in some sections.
Module wall thickness fairly uniform, ~0.09–0.1 mm, walls generally fibrous or have microgranular areas. Microstructure variously preserved; observed clearly where module wall is well preserved. Dark median line visible where adjoining modules are in contact, with a thick fibrous outer wall on both sides, perpendicular to median line (fibro-normal coating) (Fig. 6.5).
Etymology.— The new species is named for Argentina.
Remarks
Amsassia argentina n. sp. shows cerioid to phacelocerioid massive colonies, polygonal, subpolygonal, to rounded modules, scarce tabulae when present, and axial increase by longitudinal fission. All these characteristics are found in the coralomorph Amsassia, (Hill, Reference Hill1981; revised in Sun et al., Reference Sun, Elias and Lee2014; M. Lee et al., Reference Lee, Park, Tien, Choh, Elias and Lee2016), which is diagnosed by the co-occurrence of cerioid and phaceloid growth forms, with the transverse shape of modules rounded in loosely packed areas to polygonal in densely packed areas. The diameter of modules in the genus ranges from 0.06 to 1.7 mm. Module increase by longitudinal fission may be bipartite, tripartite, or quadripartite. Tabulae are rare.
Although size and shape of modules are highly variable in Amsassia, in the Argentinean material the polygonal module diameter in cerioid areas is considerably bigger than the rounded module diameter. This is uncommon in the other species of Amsassia. The size of the modules (between 0.4 and 1.1 mm) distinguishes our material from the ranges of size variation recorded in other species of the genus (reviewed in Sun et al., Reference Sun, Elias and Lee2014; M. Lee et al., Reference Lee, Park, Tien, Choh, Elias and Lee2016). The Argentinean material is assigned to a new species, A. argentina.
Species of Amsassia reported from northern Kazakhstan and western Siberia have module diameters that are less variable in size than A. argentina n. sp.: A. princeps (0.73–1.0 mm; Sokolov and Mironova, Reference Sokolov and Mironova1959), A. chaetetoides (1.0–1.4 mm; Sokolov and Mironova, Reference Sokolov and Mironova1959), and the type species A. raduguini (1.1–1.4 mm; Sokolov and Mironova, Reference Sokolov and Mironova1959).
Amsassia shaanxiensis from northern China (Sun et al., Reference Sun, Elias and Lee2014) has modules generally polygonal to subpolygonal and closely packed, rarely rounded, and partially to entirely separated, with module diameter ranging from 0.46 to 1.66 mm, which is larger than in A. argentina (0.41–1.11 mm).
Amsassia koreanensis M. Lee et al., Reference Lee, Park, Tien, Choh, Elias and Lee2016 was described from Korea and China. This form has an exceptionally small module diameter (0.06–0.28 mm), which is different from the size range observed in A. argentina.
Other species reported from China and Mongolia, include A. minima (0.2–0.4 mm; Yang et al., Reference Yang, Jin and Zhou1978), A. tudiaoensis (0.3–0.6 mm; Yang et al., Reference Yang, Jin and Zhou1978), A.(?) minima (0.3–0.5 mm; Lin and Chow, Reference Lin and Chow1980), and A. sheshanensis (Deng, Reference Deng1984; Ye et al., Reference Ye, Yang, Xu, Zheng, Zuo, Zhou, Li, Li, Song, Yong, Zhang and Zhang1995), all with smaller module diameters and narrower size ranges than A. argentina n. sp. Amsassia soluta and A. abnormis (Yu, Reference Yu1962) have a range of variation (0.5–1.1 mm; M. Lee et al., Reference Lee, Park, Tien, Choh, Elias and Lee2016) that almost overlaps the range observed in A. argentina n. sp. However, module diameters of 0.3–0.4 mm are found in our material, measured in loosely packed areas.
The dark median line separating adjacent modules, visible in some sections, could be related with early diagenesis, and apparently not related with the presence of a true medial plate or fused epithecae of adjacent modules. Unfortunately, poor preservation prevents a complete analysis of this aspect.
Amsassia is a problematic modular organism with a coral-like skeleton that recently was reevaluated by Sun et al. (Reference Sun, Elias and Lee2014) and M. Lee et al. (Reference Lee, Park, Tien, Choh, Elias and Lee2016). After a comprehensive analysis, Amsassia was compared with different types of corals, tetradiids, chaetetid sponges, and algae, those authors suggested that Amsassia may represent an extinct group of calcareous algae.
Lichenaria is a common Lower and Middle Ordovician tabulate, considered the basal genus in the early radiation of tabulates (Scrutton, Reference Scrutton1984, Reference Scrutton1997; Pandolfi, Reference Pandolfi1989). In species of Lichenaria, the corallites exclusively show a cerioid structure and are separated by fibrous, continuously fused common walls. The septal structures are absent and it has abundant tabulae (Elias et al., Reference Elias, Lee and Woo2008, and references therein). Lichenaria grew exclusively through lateral corallite increase, which is considered to be the key feature for identifying the genus (Elias et al., Reference Elias, Lee and Woo2008). All these characteristics clearly separate our material from the lichenariid stock.
Paleogeographic distribution of Amsassia and its role in the early development of metazoan reefs
In the last few years, there have been several new contributions on upper Cambrian to Lower Ordovician reefs and reef-related organisms (Adachi et al., Reference Adachi, Ezaki, Liu and Cao2009, Reference Adachi, Liu and Ezaki2012a, Reference Adachi, Ezaki and Liub, Reference Adachi, Kotani, Ezaki and Liu2015; Hong et al., Reference Hong, Cho, Choh, Woo and Lee2012, Reference Hong, Choh and Lee2015; Wang et al., Reference Wang, Deng, Wang and Li2012; Choh et al., Reference Choh, Hong, Sun, Kwon, Park, Woo, Kwon, Lee and Lee2013; Kruse and Reitner, Reference Kruse and Reitner2014; Lee et al., Reference Lee, Sun, Choh and Lee2014, and references therein). Those studies have begun to fill in the well-known gap in metazoan reef records after the demise of archaeocyaths in the late early Cambrian (Pratt et al., Reference Pratt, Spincer, Wood and Zhuravlev2001; Webby, Reference Webby2002; Rowland and Shapiro, Reference Rowland and Shapiro2002).
After collapse of the early Cambrian calcimicrobial-archaeocyathan reef consortium, metazoans played little part in reef construction for ca. 30 Myr (Rowland and Shapiro, Reference Rowland and Shapiro2002; Adachi et al., Reference Adachi, Kotani, Ezaki and Liu2015). Many reef-mound findings in the post-extinction interval were completely constituted by microbes (Webby, Reference Webby2002; Rowland and Shapiro, Reference Rowland and Shapiro2002; Lee et al., Reference Lee, Chen and Chough2015).
Nevertheless, exceptional metazoan reefs have been documented from the interval commencing around the beginning of the Furongian Epoch, when spiculate sponges began to actively participate in reef construction. Some of these reefs are mainly microbial, but demosponges helped as secondary reef builders (Cañas and Carrera, Reference Cañas and Carrera1993; Riding and Zhuravlev, Reference Riding and Zhuravlev1995; Shapiro and Rigby, Reference Shapiro and Rigby2004; Kruse and Zhuravlev, Reference Kruse and Zhuravlev2008; Wang et al., Reference Wang, Deng, Wang and Li2012; Choh et al., Reference Choh, Hong, Sun, Kwon, Park, Woo, Kwon, Lee and Lee2013; Kruse and Reitner, Reference Kruse and Reitner2014) (mainly summarized in Rowland and Shapiro, Reference Rowland and Shapiro2002; Hong et al., Reference Hong, Cho, Choh, Woo and Lee2012; Adachi et al., Reference Adachi, Kotani, Ezaki and Liu2015).
Orchoclad demosponges diversified in the late Cambrian as the main metazoan constituents of reef settings. Orchoclads, in particular the anthaspidellids, had tough, ladder-like, desma-based skeletal construction that allowed them to occupy high-energy reef environments (Carrera and Botting, Reference Carrera and Botting2008).
Other mainly Early Ordovician reef-related metazoans are the receptaculitid calathids, which commonly are associated with the orchoclad demosponges and recognized as accessory reef constructors (Carrera, Reference Carrera1991; Church, Reference Church1991; Cañas and Carrera, Reference Cañas and Carrera1993; Liu et al., Reference Liu, Zhu and Li2005; Adachi et al., Reference Adachi, Liu and Ezaki2012a; Hong et al., Reference Hong, Cho, Choh, Woo and Lee2012; Wang et al., Reference Wang, Deng, Wang and Li2012; Choh et al., Reference Choh, Hong, Sun, Kwon, Park, Woo, Kwon, Lee and Lee2013; Li et al., Reference Li, Li, Wang and Kiessling2015). Calathids are not as diverse as the orchocladine demosponges, but show a widespread distribution (Rowland and Shapiro, Reference Rowland and Shapiro2002; Webby, Reference Webby2002; Wang et al., Reference Wang, Deng, Wang and Li2012).
The stromatoporoids or stromatoporoid-like organisms, commonly domical or encrusting forms, became frame builders or accessory frame builders in varying degrees by the early Tremadocian (Toomey and Nitecki, Reference Toomey and Nitecki1979; Keller and Flügel, Reference Keller and Flügel1996; Webby, Reference Webby2002; Cañas and Carrera, Reference Cañas and Carrera2003; Zhen and Picket, Reference Zhen and Pickett2008). Pulchrilamina Toomey and Ham, Reference Toomey and Ham1967 from the Lower Ordovician of Utah and New Mexico (Toomey and Nitecki, Reference Toomey and Nitecki1979) was one of the oldest recorded forms. New records of Lower Ordovician reef-related stromatoporoids have been reported from China and Korea (J.-H. Lee et al., Reference Lee, Hong, Woo, Ho, Lee and Choh2016; Li et al., Reference Li, Li and Kiessling2016). Stromatoporoids and stromatoporoid-like organisms were classically recognized as frame builders because of their size and dome shaped or encrusting laminar forms. They are widespread, but mainly well developed in North America and China, and less frequent in South America and Australia. Later, they became the main reef constructors in Silurian and Devonian carbonate platforms.
Bryozoans have also been found associated with Ordovician reef mound structures, assuming a constructional role (Adachi et al., Reference Adachi, Ezaki and Liu2012b; Cuffey et al., Reference Cuffey, Chuantao, Zhu, Spjeldnaes and Hu2013). The main framework of these reef structures was shared with demosponges, calcimicrobes (Girvanella Nicholson and Etheridge, Reference Nicholson and Etheridge1878), and the stromatoporoid Pulchrilamina.
Corals were minor components of reefs structures during the Early Ordovician. The Lichenaria specimens, recognized by Pratt and James (Reference Pratt and James1982) in Newfoundland, are included in small thrombolite mounds or small bioherms composed of a microbe-Lichenaria-Renalcis boundstone (Pratt and James, Reference Pratt and James1982).
The coralomorph Amsassia was recently reported from north-central China as an important component of Middle Ordovician reef assemblages (Lee et al., Reference Lee, Sun, Choh and Lee2014). The reef-mound structure is characterized by peloidal masses composed of aggregated peloids, micrites, and bundles of tubular calcimicrobes, with the subordinate Amsassia, the calcimicrobe Ortonella Garwood, Reference Garwood1914, green algae, and tetradiids.
Amsassia argentina n. sp. is abundant, dome shaped, and represents an important proportion, clearly >50%, of the Lower Ordovician reef framework in western Argentina. These characteristics suggest that this form had a dominant role in the skeletal framework construction. It can be considered a main frame builder associated with a complex microbial-algal consortium composed of renalciforms, Nuia Maslov, Reference Maslov1954, and a variety of thrombolites and small stromatolites. This structure differs slightly in composition and proportions from the younger Amsassia reef framework in north-central China (Lee et al., Reference Lee, Sun, Choh and Lee2014).
Amsassia was reported from the western margin of Gondwana around the Sino-Korean and Tarim blocks from the middle part of the Middle Ordovician (M. Lee et al., Reference Lee, Sun, Choh and Lee2014, Reference Lee, Park, Tien, Choh, Elias and Lee2016, and references therein), and wider occurrences have been reported from the early Late Ordovician of Kazakhstan (e.g., Popov et al., Reference Popov, Cocks and Nikitin2002), Siberia (Bondarenko and Ulitina, Reference Bondarenko and Ulitina2009), and a single report from the Late Ordovician of Laurentia (Bolton, Reference Bolton2000).
Discovery of Amsassia in the lowermost Ordovician modifies the paleogeographic scenario envisaged by M. Lee et al. (Reference Lee, Sun, Choh and Lee2014, Reference Lee, Park, Tien, Choh, Elias and Lee2016). These authors suggested that Amsassia emerged in western Gondwana (Sino-Korean and Tarim blocks), where the genus appears to have originated in the Middle Ordovician, and might subsequently have spread into Kazakhstan and Siberia prior to the early Late Ordovician.
With this new discovery in the Lower Ordovician of Argentina, the spatio-temporal distribution of Amsassia is substantially modified: its origination and migration pathway is significantly changed. This new record shows that Amsassia emerged from the eastern margin of Laurentia, since the Argentine Precordillera rifted from this continent by the late Cambrian (Astini et al., Reference Astini, Benedetto and Vaccari1995; Benedetto, Reference Benedetto2004). Subsequent records occur in the Middle Ordovician of the Sino-Korean and Tarim block (Lee et al., Reference Lee, Sun, Choh and Lee2014), and later the expansion continued in Siberia and Kazakhstan. The record in Canada (Bolton, Reference Bolton2000) by the early Late Ordovician can be explained by its proximity with the Siberian block. The presence of Amsassia in the “Laurentian stage” of the Argentine Precordillera suggests that we should expect Lower or Middle Ordovician Laurentian records between the Lower Ordovician occurrence of Amsassia in the Argentine Precordillera and their subsequent presence in the Late Ordovician of Canada. This probably reflects a gap in the sampling record more than a true dispersion pattern. However, as Sun et al. (Reference Sun, Elias and Lee2014) pointed out, there is a critical need to re-examine the species previously referred to Lichenaria or Amsassia, which have been commonly misidentified.
By the late Cambrian and Early Ordovician, a variety of metazoan forms occupied the niches left by archaeocyaths with varying degrees of success. All these metazoan forms were aided in reef or biostrome construction by microbes. Large-sized organisms (orchoclad demosponges, calathids, stromatoporoids) could easily form the scaffold structure of the framework, and usually required a smaller proportion of microbes or other secondary constructors. In our example, it is clear that the high proportion of Amsassia colonies represented in the boundstone, the coalescent arrangements, and the stacking patterns they acquired, enabled them to be the primary scaffolding organism of the reef framestone. The new paleogeographic scenario shows that Amsassia acquired a worldwide distribution with a significant participation in reef construction and represents a successful attempt of skeletal organisms to construct reefs during the Early to Middle Ordovician interval.
Acknowledgments
The authors acknowledge support from the National Research Council of Argentina. MGC acknowledges support from CONICET (Grant PIP 2009- 00861). RAA acknowledges support from SECYT-UNC 213/14. Appreciation is extended to the reviewers, G.A. Young and D.-J. Lee, for their helpful comments and suggestions that significantly improved the final version of the manuscript. We would also like to thank the editorial staff of the Journal for their assistance. This is a contribution to the IGCP 653 project, “The onset of the Great Ordovician Biodiversity Event”.







