


Reports linking certain types of dietary fat to serum lipid levels have often been interpreted to mean that the general public, especially those at risk for coronary vascular disease (CVD), should consume diets containing little or no red meat. Early research concluded that dietary SFA such as palmitic acid (16 : 0) elevate serum cholesterol concentrations, PUFA, especially linoleic acid (18 : 2n-6), reduce serum cholesterol concentrations, and MUFA have little or no effect(Reference Hegsted, McGandy and Myers1–Reference Keys, Anderson and Grande3). The major MUFA in beef, oleic acid (18 : 1n-9), has been studied in more detail and found to lower LDL cholesterol without affecting the beneficial HDL cholesterol (HDL-C)(Reference Baggio, Pagnan and Muraca4, Reference Grundy, Florentin and Nix5). This effect is most convincing in studies in which natural foods were used to supplement diets with oleic acid(Reference Mensink and Katan6–Reference Berry, Eisenberg and Haratz8). In addition, different SFA have been found to have different effects on serum cholesterol concentrations, as stearic acid (18 : 0) was shown to have no effect or even to lower serum cholesterol(Reference Bonanome and Grundy9, Reference Grande, Anderson and Keys10).
Some beef products have been shown to decrease(Reference O'Dea, Traianedes and Chisholm11) or have no effect(Reference Scott, Kimball and Wittels12, Reference Smith, Wood and Tseng13) on serum cholesterol in free-living individuals. These earlier studies of the effects of beef consumption on serum cholesterol concentration did not consider that beef products can vary in their MUFA:SFA ratio, or take advantage of beef products with widely differing MUFA:SFA ratios within the context of total beef fat intake. Fat from pasture- or hay-fed cattle contains a high proportion of SFA, and this beef fat also is higher in trans-fatty acids (TFA)(Reference Chung, Lunt and Choi14). Conversely, high-MUFA beef fat with very low concentrations of TFA can be obtained from cattle that have been grain fed for extended periods (i.e. ‘long fed’)(Reference Chung, Lunt and Choi14, Reference Sturdivant, Lunt and Smith15). Certain breed types such as American Wagyu (derived from crossing Japanese Black and Japanese Red bulls on Angus cows) have a genetic propensity to accumulate MUFA in muscle and adipose tissue, and ground beef especially enriched with MUFA can be obtained from Wagyu steers, although feeding practices markedly affect the degree of enrichment with MUFA(Reference Chung, Lunt and Choi14) (the present study).
In the present study, we compared several risks factors for CVD in mildly hypercholesterolaemic male subjects after consumption of either high-SFA, high-TFA hamburger or high-MUFA, low-TFA hamburger for 5 weeks with a 3-week washout period. This experiment tested the hypothesis that risk factors for CVD would be higher in mildly hypercholesterolaemic men after consumption of hamburger enriched with SFA and TFA than after consumption of hamburger enriched with MUFA.
Materials and methods
Experimental design
Texas A&M University faculty and staff (n 10) were recruited for the present study. Normal, healthy and non-smoking males between the ages of 30 and 60 years were screened with a battery of blood chemistry tests by a local physician (S. T.). Subjects with total serum cholesterol values between 5 and 6·5 mmol/l and not on restrictive diets or medications were selected and given a complete physical examination, including an electrocardiogram and a family history. All participants provided informed consent and were free living. Exercise and physical activities were not restricted, but participants were requested not to change their habitual level of physical activity in order to maintain body weight ( ± 2·2 kg of entry weight). Subject characteristics and baseline lipid and dietary profiles are shown in Table 1.
Table 1 Baseline characteristics for subjects
(Mean values with their standard errors for ten men)
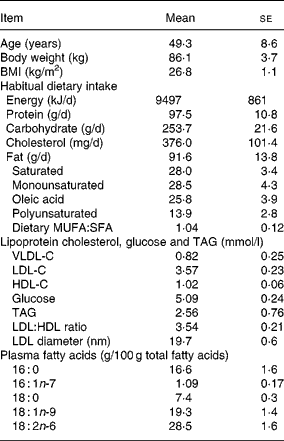
VLDL-C, VLDL cholesterol; LDL-C, LDL cholesterol; HDL-C, HDL cholesterol.
The ten men were fed high-SFA hamburger for a 5-week period and, following a 3-week habitual diet washout period, were rotated to high-MUFA hamburger. The subjects were contacted weekly to ensure that all five beef patties were consumed during each weekly test period. The test subjects were not informed as to which type beef they had been assigned. The beef was supplied to the participants in the form of 114 g hamburger patties (5 patties/week). The frozen, vacuum-packaged hamburger patties for an entire diet period were delivered to the participants on or before the first day. No restrictions were placed on how the beef was to be prepared other than that all of the beef be consumed at each sitting.
Preparation of hamburgers
Hamburgers were prepared at the Texas A&M Rosenthal Meat Science & Technology Center, Texas A&M University. By definition, ground beef contains only fat associated with the lean trim from which it is grounded(16). Because fat trim from other parts of the carcass and/or from different cattle was added to the source of lean trim, the term ground beef cannot be used and so the term hamburger is used.
High-SFA hamburger was formulated from lean and fat trims from domestic cattle and from Wagyu steers-fed pasture-based diets. The high-MUFA hamburger was formulated from lean and fat trims of domestic cattle and Wagyu steers fed a maize-based diet for an extended period of time (a minimum 8 month after weaning). The Wagyu fat trims were obtained from a local producer of genetically similar full-blood Japanese Black (Wagyu) cattle. The domestic fat trim and all lean trims were obtained from the Rosenthal Meat Science and Technology Center. Hamburger patties were formulated to achieve 35 % targeted total fat, so that each 114 g patty contained approximately 40 g total fat. Patties were individually vacuum-packed, quick-frozen and boxed by diet type.
The high-SFA hamburger contained over 2 g more stearic acid per patty than the high-MUFA hamburger (6·14 g v. 4·01 g; Table 2), and the high-MUFA hamburger contained over 2 g more oleic acid per patty (17·2 g v. 15·0 g). Each hamburger type provided a similar amount of palmitic acid (approximately 9·4 g/patty). The high-SFA hamburger also contained 0·48 g more total TFA and 0·014 g more α-linolenic acid (18 : 3n-3) than the high-MUFA hamburger.
Table 2 Fatty acid composition of hamburger high in SFA (high SFA) and hamburger enriched in MUFA (high MUFA)†
(Mean values with their standard errors)

Mean values were significantly different: *P ≤ 0·05, **P ≤ 0·01, ***P ≤ 0·001.
† Data were analysed by Student's t test.
‡ MUFA: 14 : 1n-5+16 : 1n-7+18 : 1n-9+18 : 1n-7+18 : 2(cis-9, trans-11). SFA: 14 : 0+16 : 0+18 : 0+18 : 1(trans-10)+18 : 1(trans-11). Total PUFA = 18 : 2n-6+18 : 3n-3. Total trans-fatty acids = 18 : 1(trans-11)+18 : 1(trans-10).
Survey of area ground beef
In order to empirically determine the range or MUFA:SFA ratio in commercially available products, we conducted a survey of ground beef from retail outlets within the College Station area and Wagyu ground beef that was purchased from an internet vendor. Determined fatty acid compositions were used to calculate amounts of individual fatty acids in 114 g patties containing 20 % fat. This level of fat was selected as it is the most frequently purchased form of hamburger.
Determination of cholesterol fractions, TAG and glucose
Blood was collected from an arm vein before initiation of the dietary treatments and at the end of treatment. Plasma was harvested from blood collected with EDTA and lipoproteins preserved(Reference Walzem, Watkins and Frankel17) before lipoprotein separation, using density gradient ultracentrifugation employing human density intervals(Reference Walzem, Davis and Hansen18)and determination of LDL lipoprotein diameter(Reference Walzem, Davis and Hansen18, Reference Havel, Eder and Bragdon19).
Plasma total lipoproteins isolated as the d < 1·2 g/ml fraction of plasma were separated on the basis of diameter with a gel filtration chromatographic system(Reference Wang, Yu and Walzem20) in order to determine the relative distribution of plasma total cholesterol and TAG among VLDL, LDL and HDL lipoprotein classes. Separate analyses were made for cholesterol and TAG and, in each, the eluting lipids were continuously monitored at 505 nm following enzymatic chromophore development within an in-line post-column reactor(Reference Wang, Yu and Walzem20). Plasma total cholesterol, TAG and glucose were determined by separate enzymatic assays (Sigma Chemical Co., St Louis, MO, USA).
Fatty acid composition of plasma and test hamburger
Fatty acids were measured in the baseline whole-plasma samples and from whole-plasma samples taken after 5 weeks of each test hamburger treatment. Additionally, fatty acid concentrations and concentrations of fat and moisture(Reference Helrich21) of the test hamburgers were measured for every batch of product (a minimum of three batches per beef fat combination). Total lipid was extracted and methylated as described(Reference Folch, Lees and Sloane Stanley22, Reference Morrison and Smith23), and fatty acid methyl esters were analysed with a Varian gas chromatograph (model CP-3800 fixed with a CP-8200 autosampler; Varian Inc., Walnut Creek, CA, USA).
Separation of fatty acid methyl esters was accomplished on a fused silica capillary column CP-Sil 88 (100 m × 0·25 mm internal diameter) (Chrompack Inc., Middleburg, The Netherlands) with He as the carrier gas (1·2 ml/min). After 32 min at 180°C, oven temperature was increased at 20°C/min to 225°C and held for 14·15 min. Injector and detector temperatures were at 270 and 300°C, respectively. Individual fatty acid methyl esters were identified using genuine standards (Nu-Chek Prep, Inc., Elysian, MN, USA) and expressed as a g/100 g total fatty acid methyl esters analysed or as g/114 g hamburger patty.
Diet records
Before each diet phase, and once during each phase, participants completed a 3 d record (to include 1 weekend day). The diet records were analysed for nutrient composition by a registered dietitian to establish baseline observations, and encourage compliance with the requirement of total patty consumption. The diets records were analysed using Nutrient Calc version 1.1 (University of Minnesota, St Paul, MN, USA). Plasma fatty acid compositions were used to verify recorded patterns of fatty acid intake.
Statistical analyses
Retail ground beef fatty acid composition was analysed by ANOVA (SuperAnova; Abacus Concepts, Inc., Berkeley, CA, USA). When the ground beef type was significant (P ≤ 0·05), means were separated by the Fisher's protected LSD method. Fatty acid composition of the test hamburgers was tested by the Student's t test and, after the test, plasma concentrations of lipoprotein cholesterol, glucose, TAG and fatty acids were compared by a paired t test.
The present study was conducted according to the guidelines laid down in the Declaration of Helsinki, and all procedures involving human subjects were approved by the Texas A&M University Institutional Review Board for use of human subjects in research (Protocol Number 2004-0026). Written informed consent was obtained from all subjects.
Results
Fatty acid composition of retail ground beef
Chub pack, ground chuck and ground round all had MUFA:SFA ratio less than 1·0 (Table 3). The lowest MUFA:SFA ratio in ground beef was observed in chub pack ground beef (0·84), and the highest ratio (1·46) was measured in a branded ground beef from maize-fed Wagyu cattle. There was no difference in the amount of palmitic acid per 114 g serving across retail ground beef types. The chub pack ground beef contained more stearic acid and TFA, and less oleic acid, than the branded Angus and Wagyu ground beeves.
Table 3 Fatty acid composition of ground beef purchased from local retailers*
(Mean values with their standard errors for a minimum of three samples per ground beef type)
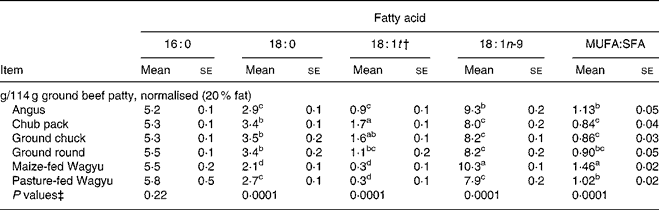
Mean values within a column with common superscripts or no superscripts are not different (P>0·05).
* Data are normalised to 20 % total fat (22·8 g fat/114 g ground beef patty). Chub pack, ground round, ground chuck and Angus ground beeves were purchased from three major retailers in the vicinity of Texas A&M University. Wagyu ground beef (grain- and pasture-fed) was purchased from a web-based supplier. Lipids were extracted and fatty acids analysed as described in the text. A minimum of three samples of each type of product were analysed.
† Sum of 18 : 1(trans-10) plus 18 : 1(trans-11). Not all fatty acids present in the ground beef are listed in the table.
‡ Data were analysed by ANOVA with ground beef type as the main effect.
Nutrient intake, body weights and plasma glucose concentrations
The intakes of total fat, SFA, MUFA and oleic acid were greater during consumption of the test hamburgers than for the habitual diets (all P values ≤ 0·05; Tables 1 and 4). During the high-MUFA phase, participants consumed less SFA and more MUFA than during the high-SFA hamburger phase. Participants consumed approximately 40 g/d more fat during the test phases than during their habitual intake, indicating that most participants ate the beef patty in addition to their habitual meals. In spite of the greater daily fat intake, initial (86·1 (sd 3·7) kg) and final (85·9 (sd 3·8) kg) mean body weights were not different.
Table 4 Daily intake of nutrients for test diets of men rotated from hamburger containing fat trim high in SFA (high SFA) to hamburger containing fat trim high in MUFA (high MUFA)†
(Mean values with their standard errors for three diet records from ten men per test hamburger)
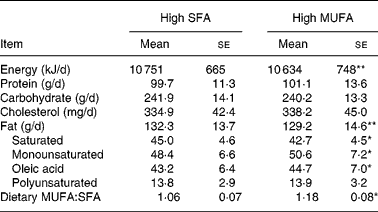
Mean values were significantly different: *P ≤ 0·05, **P ≤ 0·01.
† Data were analysed by paired t test.
Participants consumed 117 fewer kJ/d during the high-MUFA phase than during the high-SFA phase (Table 4). This difference, although small (1 % of total energy intake), was statistically significant, and was caused by lesser intake of total fat during the high-MUFA phase than during the high-SFA phase. Daily intakes of protein, carbohydrate, cholesterol and PUFA were not different between the test phases.
Plasma lipoprotein cholesterol concentrations, plasma fatty acid concentrations and LDL particle diameters
The concentration of plasma TAG and the LDL:HDL ratio were greater (P < 0·05) after the high-SFA hamburger phase than after the high-MUFA hamburger phase (Table 5). Conversely, HDL-C was greater after consumption of the high-MUFA hamburger than after consumption of the high-SFA hamburger.
Table 5 Lipoprotein cholesterol, glucose, TAG and plasma fatty acid concentrations for mildly hypercholesterolaemic men-fed hamburger containing fat trim high in SFA (high SFA) or fat trim high in MUFA (high MUFA)†
(Mean values with their standard errors for ten men per test hamburger)
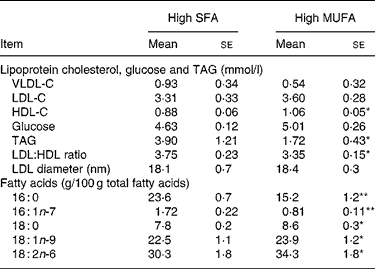
VLDL-C, VLDL cholesterol; LDL-C, LDL cholesterol; HDL-C, HDL cholesterol.
Mean values were significantly different: *P ≤ 0·05, **P ≤ 0·001.
† Data were analysed by paired t test.
Mean LDL particle diameters were 19·7 (sd 0·6) nm at baseline, and LDL particle diameters ranged from 13·4 (sd 0·4) nm at the 10th percentile to 25·4 (sd 0·7) nm at the 90th percentile of the overall particle population diameter distribution. Percentage baseline particle diameter was significantly different between the high-SFA and high-MUFA treatment periods beyond the 50th percentile of the LDL particle population (Fig. 1), reflecting the depression in particle diameter caused by the high-SFA hamburger (to 18·1 (sd 0·7) nm; Tables 1 and 5). LDL particle diameters did not increase significantly during the 3-week washout period (18·3 (sd 0·2) nm; Fig. 1) or during the high-MUFA hamburger phase (18·4 (sd 0·3) nm; Table 5).

Fig. 1 Changes from baseline values for LDL particle diameter percentiles for men rotated from hamburger high in SFA (●, high SFA) to hamburger enriched in MUFA (○, high MUFA). Baseline LDL particle diameters were 19·1 (sd 0·7) nm (before high-SFA phase) and 18·3 (sd 0·2) nm (before high-MUFA phase). Data are population percentiles for ten men for each hamburger type. Pooled sem is affixed to the symbols. Diameters at each decile were compared by a paired t test. *P ≤ 0·05, **P ≤ 0·01.
Plasma concentrations of palmitic and palmitoleic acids were higher after the high-SFA phase than after high-MUFA phase (Table 5). Conversely, plasma stearic, oleic and linoleic acid concentrations were higher after consumption of the high-MUFA hamburger than after consumption of the high-SFA hamburger. Unlike other plasma fatty acids, palmitic acid did not return to pre-treatment values after the 3-week washout period, but remained elevated (20·5 (sd 0·7) g/100 g plasma fatty acids; data not shown in Table 5).
Plasma palmitic, palmitoleic and oleic acids were positively correlated with plasma TAG and VLDL cholesterol concentrations (Table 6). The highest correlation was between palmitoleic acid and TAG (r 0·90). Palmitic and palmitoleic acids were negatively correlated with HDL-C and positively correlated with the LDL:HDL ratio (all r ≥ 0·34). Palmitic, stearic and oleic acids were negatively correlated with LDL particle diameters. Linoleic acid was negatively associated with TAG, VLDL cholesterol and the LDL:HDL ratio.
Table 6 Simple correlation coefficients between plasma fatty acids and lipoprotein cholesterol measures for mildly hypercholesterolaemic men-fed hamburger containing fat trim high in SFA or fat trim high in MUFA†
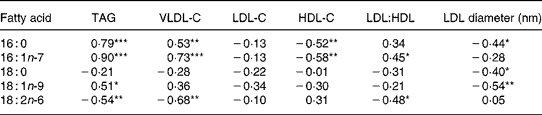
VLDL-C, VLDL cholesterol; LDL-C, LDL cholesterol; HDL-C, HDL cholesterol.
Mean values were significantly different: *P ≤ 0·05, **P ≤ 0·01, ***P ≤ 0·001.
† Data are from baseline and final samples.
Discussion
In 2000, the Nutrition Committee of the American Heart Association moved away from its former insistence on low-fat diets and concluded that diets providing up to 40 % of dietary energy as primarily unsaturated fat (20 % MUFA, < 10 % SFA, 10 % PUFA and 1 % TFA) were as heart healthy as low-fat diets(Reference Krauss, Eckel and Howard24). The recommendations were revised down to accommodate 35 % total fat and < 7 % SFA. An outcome of this official opinion has been the re-evaluation of the nutritional properties of a number of higher fat foods such as dairy, nuts and dietary oils such as olive oil, which are rich in oleic acid(Reference Kris-Etherton, Etherton and Carlson25). It is of interest to note that, during consumption of their habitual diets, total fat constituted approximately 35 % of total dietary energy, with 11 % from SFA, 11 % from MUFA and 5 % from PUFA. The diet records indicated that the hamburger patties were added onto the habitual diets, rather than replacing a portion of the meat of their habitual diets, so the participants consumed an additional 40 g/patty during the test phases (45–46 % total dietary energy from fat). Participants consumed as much as 2·5 g more TFA, 12 g more SFA and 15·5 g less MUFA each week during the high-SFA hamburger phase than they consumed during the high-MUFA hamburger phase.
Beef or beef products that vary widely in fatty acid composition have not yet been evaluated with regards to their effects on risk factors for CVD, perhaps because the fatty acid composition of beef was considered to be constant. Our survey of retail ground beef indicated that the MUFA:SFA ratios tested in the present study were reflective of the variation present in the available food supply. Of the ground beef types evaluated, most contained approximately 20 % total fat, but the chub pack ground beef contained considerably more fat (28 %) than the other ground beef types. Both the chub pack and ground chuck ground beef from local retail outlets contained more total TFA than the high-SFA hamburger used in the present study, even though the high-SFA hamburger contained more total fat. These data indicate that habitual consumption of the relatively inexpensive high-fat chub pack ground beef potentially could cause some of the same effects caused by the high-SFA test hamburger.
Plasma palmitoleic acid and apparent hepatic stearoyl-CoA desaturase-1 activity
Warensjo et al. (Reference Warensjo, Sundstrom and Vessby26) evaluated the relationship between serum fatty acids and risk for CVD mortality and total mortality in 1885 men from the Uppsala Longitudinal Study of Adult Men. They reported that, of the individual serum fatty acids, the greatest mortality risk was associated with palmitoleic acid, followed closely by palmitic acid. The serum concentration of linoleic acid was inversely related with CVD and total mortality. Warensjo et al. (Reference Warensjo, Sundstrom and Vessby26) concluded that serum palmitoleic acid and the palmitoleic:palmitic ratio served as indices of hepatic stearoyl-CoA desaturase-1 (SCD1) activity, and that elevated hepatic SCD1 activity was positively associated with CVD mortality. In the present investigation, palmitoleic acid was the plasma fatty acid most highly correlated with changes in TAG, VLDL cholesterol and HDL-C, followed by palmitic acid. The highest plasma palmitoleic acid concentration was observed at the end of the high-SFA phase and the lowest after the high-MUFA phase, even though high-SFA hamburger consumption delivered 29·5 g of palmitoleic acid in the 5-week feeding period, which was much less than the 43·5 g provided by 5 weeks of high-MUFA hamburger consumption. Clearly, the concentration of palmitoleic acid in the test hamburger cannot explain the variation in plasma palmitoleic acid. Therefore, the high-SFA ground beef may have stimulated hepatic SCD1 activity, which was reversed by consumption of the high-MUFA ground beef.
Ntambi and co-workers(Reference Miyazaki, Kim and Gray-Keller27) previously demonstrated that VLDL–TAG were virtually undetectable in mice with a disruption in the SCD1 gene. In livers of SCD1 knockout mice, the concentration of palmitoleic was reduced nearly 50 %. Sampath et al. (Reference Sampath, Miyazaki and Dobrzyn28) reported that Δ9 desaturation of saturated fats such as stearic acid by SCD1 was an essential step in mediating their ability to induce hepatic lipogenesis. Enoch et al. (Reference Enoch, Catala and Strittmatter29) demonstrated that palmitoyl-CoA and stearoyl-CoA have similar substrate properties for SCD1, and that oleoyl-CoA inhibits SCD1, in rat hepatocytes. The high-SFA hamburger provided 15·7 g of SCD1 substrates (palmitic and stearic acid) and 15 g of potentially SCD1-inhibitory oleic acid. In contrast, the high-MUFA hamburger provided 16 % less (13·2 g) SCD1 substrate and 15 % more (17·2 g) inhibitory oleic acid. The marked changes in plasma palmitoleic acid in the present study suggest that hepatic SCD1 activity is sensitive to the composition of ground beef available in retail markets, a proposition that requires direct testing.
LDL particle diameters
LDL particle diameters were reduced by the high-SFA hamburger, and diameters remained depressed even after the 3-week washout period as well as after consumption of the high-MUFA hamburger. Similarly, plasma palmitic acid was elevated by the high-SFA diet and remained elevated thereafter. Differences in LDL particle diameter represent specific metabolic changes that increase the atherogenicity of LDL(Reference Veniant, Sullivan and Kim30). Small, dense LDL particles are recognised as a risk factor for CVD, as this form of LDL is more susceptible to oxidative damage(Reference Chait, Brazg and Tribble31) and promotes vascular inflammation(Reference Libby32). The persistent, high-circulating concentrations of palmitic acid following consumption of the high-SFA hamburger may have depressed LDL clearance. This would have caused the reduced LDL particle diameters we observed following the high-SFA phase, which persisted through the washout period and the high-MUFA phase. This is supported by the negative correlation between plasma palmitic acid and LDL particle diameters. The observation that LDL particle diameters were not affected by the high-MUFA hamburger suggests that the additional oleic acid in the high-MUFA hamburger was unable to offset the depression in LDL diameter caused by the palmitic acid.
We previously established the effects of the consumption of high-SFA hamburger (17 % fat; MUFA:SFA = 0·83–0·96) on lipoprotein cholesterol metabolism in free-living men(Reference Smith, Wood and Tseng13). High-SFA hamburger increased the apoB:LDL cholesterol ratio, suggesting that LDL particles became smaller and more dense. This was confirmed by the results of the present study, and indicates that reduction in LDL particle diameters is a consistent effect of high-SFA hamburger. A previous study(Reference Kratz, Gulbahce and von Eckardstein33) concluded that, relative to a high-SFA, habitual diet, consumption of oils enriched in MUFA or PUFA reduced LDL diameter. However, these changes were less than 0·36 nm and the diet highest in MUFA (olive oil) actually increased LDL particle diameter by 0·13 nm(Reference Kratz, Gulbahce and von Eckardstein33). Krauss(Reference Krauss34)previously reported that in approximately 70 % of men (LDL subclass pattern A), reduction in LDL cholesterol in response to low-fat diets is the result of depletion of the cholesterol content of LDL particles; this is accompanied by a shift to smaller LDL particles. Wang et al. (Reference Wang, Yu and Walzem20) later confirmed that high-carbohydrate diets reduce LDL particle diameters in hamsters.
These earlier studies suggest that, in response to a high-fat diet enriched in SFA (and lower in carbohydrate), LDL particle diameters should have increased during the first phase of the present study. However, the change in percentage energy from carbohydrates between the habitual (approximately 45 %) and test hamburger phases (38–39 %) in the present study would not be considered to constitute a shift from a high-carbohydrate to a low-carbohydrate diet; nor would any of these diets be considered as low-fat diets (35–46 % energy from fat). Instead, some component(s) of the high-SFA ground beef interacted with the increase in total fat intake to reduce LDL particle diameter. Potential candidates are 18 : 1trans-10 and trans-vaccenic acid, as TFA have been shown to have adverse effects on measures of CVD(Reference Kris-Etherton and Yu35). trans-Vaccenic acid has been shown to increase the LDL:HDL ratio in hamsters(Reference Meijer, van Tol and van Berkel36), although the effects of TFA on LDL particle diameters have not been reported.
General considerations
Ground beef and hamburger from fast-food outlets are the most common sources of MUFA for adults(Reference Nicklas, Hampl and Taylor37), so production practices that can increase the concentration of oleic acid or, conversely, increase SFA and TFA in beef may differentially affect risk factors for CVD. Cattle with a genetic predisposition to deposit MUFA in their lean and fat tissues, such as Wagyu cattle(Reference Chung, Lunt and Choi14, Reference Sturdivant, Lunt and Smith15), can be used to produce beef products that are especially enriched with oleic acid and lower in SFA and TFA, and feeding practices can further enhance the composition of beef fat. This indicates that ground beef or hamburger products can be produced, which are naturally enriched with oleic acid, and conversely that certain production practices can impair the nutritional quality of beef fat.
Finally, we cannot discern from the present study design whether the high-MUFA hamburger reversed the effects of the high-SFA hamburger, or whether the subjects gradually adapted to the elevated intake of total fat. It is clear, however, that the high-MUFA hamburger did not exacerbate any of the effects of the high-SFA hamburger and can be viewed as at least neutral in its effects on HDL-C and TAG.
Acknowledgements
Technical article, Texas Agricultural Experiment Station. The present study was supported in part by a grant by YamaBeef Inc., Mabank, TX, USA, Texas Agricultural Experiment grant no. 5027-04. YamaBeef Inc. produces Wagyu beef and also provided beef fat trim that was high in saturated fat as well as beef fat trim that was high in monounsaturated fat. T. H. A. (graduate student) had primary responsibility for analysing cholesterol concentrations, particle diameters and fatty acid concentrations; R. L. W. (Associate Professor) provided training and technical support for the measurement of cholesterol concentrations and particle diameters; D. R. S. (registered dietitian) provided dietary instructions and analysed diet records; S. T. (physician) assisted in the screening and selection of participants; S. B. S. (Professor) provided training and technical support for all other biochemical measurements, formulated raw materials for the hamburgers and provided oversight for the design and execution of the present study. There were no personal or financial conflicts of interest by the authors.







