


It is important to develop effective strategies to reduce energy intake and increase energy expenditure (EE) to fight the growing obesity epidemic. An intriguing strategy to increase EE is facultative thermogenesis induced by cold exposure(Reference Yoneshiro, Aita and Kawai1). Cold-induced shivering (ST) and non-shivering thermogenic (NST) processes produce heat as a result of sympathetic nervous system induction and the release of its neurotransmitter, noradrenalin (NA)(Reference Morrison and Nakamura2). In 2009, evidence showing the presence of brown adipose tissue (BAT) in adult human subjects renewed interest in the role of this tissue to contribute to total NST(Reference van Marken Lichtenbelt, Vanhommerig and Smulders3–Reference Cypess, Lehman and Williams5). In this context, we showed for the first time that BAT is not only metabolically active, but can also have whole-body effects through modulating shivering activity(Reference Ouellet, Labbe and Blondin6). Consequently, the question begs: could ingested agents increase the activation of NST in order to increase overall EE and/or reduce the contribution of ST?
In an attempt to stimulate thermogenesis, several studies have been conducted using green tea and its components(Reference Berube-Parent, Pelletier and Dore7–Reference Rudelle, Ferruzzi and Cristiani9). One such study(Reference Dulloo, Duret and Rohrer8) showed that green tea extract (90 mg of epigallocatechin-3-gallate (EGCG) and 50 mg of caffeine) increased EE at rest over a 24 h period through sympathetic activation. The thermogenic properties of green tea reside primarily in the interaction between its high content in catechin polyphenols and caffeine and their effect on sympathetically released NA. When co-ingested in basal conditions, EGCG and caffeine synergistically stimulate thermogenesis by 4–8 %(Reference Dulloo, Duret and Rohrer8, Reference Rudelle, Ferruzzi and Cristiani9). In contrast, during cold exposure, where BAT activity would be greatly increased, only one study has reported the effects of caffeine on thermogenic responses. MacNaughton et al. (Reference MacNaughton, Sathasivam and Vallerand10) reported a 30 % increase in EE when cold-exposed men (2 h in air at 5°C) ingested a gel capsule containing 385 mg (5 mg/kg) of caffeine. While this effect alone is significant, the synergistical effect of a combined EGCG/caffeine ingestion on thermogenic processes have never been quantified in cold conditions.
Therefore, the purpose of the present study was to quantify the combined effects of EGCG and caffeine on processes of thermogenesis (NST and ST) during mild cold exposure. Using a combination of indirect calorimetry and electromyography (EMG), changes in total EE and muscle shivering intensity were quantified in non-acclimatised young men ingesting 1600 mg of EGCG and 600 mg of caffeine. It was hypothesised that, when given at the onset of cold exposure, these compounds would increase NST, thus reducing overall shivering intensity in large torso muscles known to contribute significantly to ST during cold exposure(Reference Ouellet, Labbe and Blondin6, Reference Haman, Legault and Rakobowchuk11, Reference Bell, Tikuisis and Jacobs12). The stimulation of NST will be assumed if, when compared with a placebo: (1) EE increases when given EGCG+caffeine, despite a similar shivering intensity or (2) shivering intensity decreases when given EGCG+caffeine for a similar EE.
Materials and methods
Subjects
A total of eight healthy, non-cold acclimatised men volunteered for the present study conducted according to the guidelines laid down in the Declaration of Helsinki All procedures involving human subjects/patients were approved by the Faculty of Health Sciences ethics committee at the University of Ottawa. Written informed consent was obtained from all subjects. Exclusion criteria included the following: percentage body fat >15 %, smokers, consumption of >200 mg/d of caffeine and use of dietary supplements or stimulants. Anthropometric measurements (height, weight and percentage body fat) and peak oxygen consumption (VO2max) were estimated 5–7 d before the first experimental session (Table 1).
Table 1 Characteristics of male participants (Mean values with their standard errors, n 8)

* Underwater weighing; Brosek et al. (1945).
† Canadian Society for Exercise Physiology incremental treadmill exercise test.
Experimental protocol
Each subject participated in two experimental trials, separated by at least 7 d. The order of the trials was randomly assigned and followed by a single-blind, balanced, cross-over design. Participants ingested a gel capsule of green tea extracts consisting of 1600 mg of EGCG and 600 mg of caffeine (Green tea) or a placebo containing all-purpose white and bran flour (Placebo). Each trial consisted of a 60 min baseline period at ambient temperature (22·7 ± 0·4°C), followed by 180 min of cold exposure. Experiments were conducted from 07.30 to 14.30 hours. Participants refrained from consuming caffeine, alcohol and avoided physical activity 24 h prior to the trials. The last evening meal, ingested between 18.00 and 20.00 hours, was standardised (3220 kJ or 770 kcal, 42 % carbohydrates (CHO), 28 % fat and 30 % protein) and subjects were asked to report to the laboratory at 07.30 hours the next morning after a 12–14 h fast.
Upon their arrival to the laboratory, subjects were asked to empty their bladder and then, wearing only shorts, were fitted with a liquid-conditioned suit (three-piece high density; Allen-Vanguard, Inc.). They were instrumented with thermal probes, a heart rate monitor and EMG electrodes. Maximal voluntary contraction (MVC) measurements were carried out for each recorded muscle. Volunteers then remained seated for 60 min at ambient conditions (22·7 ± 0·4°C) under a canopy. Following the baseline period, subjects were asked to empty their bladder for a second time and ingested either the green tea extracts or placebo containing capsule. The liquid-conditioned suit was then perfused with 15°C water using a temperature-controlled circulation bath (Thermoscientific Neslab Refrigirated RTE-7 Bath Circulator). Previous work has shown that perfusion of water from approximately 4–18°C only results in a grade decrease in average skin temperature without any modification of core temperature, indicating that thermal balance is achieved; increases in heat loss are compensated by increases in metabolic heat production(Reference Haman, Legault and Rakobowchuk11, Reference Blondin, Depault and Imbeault13–Reference Blondin, Peronnet and Haman19). Thermal response, metabolic rate and muscle activity were measured continuously during baseline and during the 180 min cold exposure.
Thermal response
Rectal (T rec) and mean skin temperature (T skin) were monitored continuously prior to and during cold exposure using peadiatric rectal (Mon-a-therm general purpose; Mallinckrodt Medical, Inc.) and heat flux transducers (area-weighed equation from twelve sites: forehead, chest, biceps, forearm, abdomen, lower and upper back, front and back calf, quadriceps, hamstrings and hand(Reference Hardy and Dubois20)). Heart rate and thermal comfort were measured every 15 min throughout the experiments. Heart rate was measured using a Polar heart rate monitor (Polar FS2C Fitness Heart Rate Monitor System) and thermal comfort was monitored using a thermal comfort scale, ranging from 5 (being the hottest) to − 5 (being the coldest).
Metabolic measurements
Changes in whole-body EE and fuel selection were quantified by indirect calorimetry (Flow Generator/Controller, FoxBox Field Gas Analysis System, RH-300 Water Vapor Analyser; Sable systems). Total protein (PROTox), CHO (CHOox) and lipid (Lipidsox) oxidation rates (in g/min) were derived from oxygen consumption rate (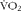 $$V\dot O_{2} $$) and CO2 production (
$$V\dot O_{2} $$) and CO2 production (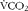 $$V\dot CO_{2} $$) measurements corrected for the contribution of proteins, as determined from urinary urea excretion (UREAurine). Calculations were made using the following equations(Reference Haman, Peronnet and Kenny14–Reference Haman, Peronnet and Kenny16, Reference Blondin, Peronnet and Haman18, Reference Blondin, Depault and Imbeault21, Reference Haman, Legault and Weber22):
$$V\dot CO_{2} $$) measurements corrected for the contribution of proteins, as determined from urinary urea excretion (UREAurine). Calculations were made using the following equations(Reference Haman, Peronnet and Kenny14–Reference Haman, Peronnet and Kenny16, Reference Blondin, Peronnet and Haman18, Reference Blondin, Depault and Imbeault21, Reference Haman, Legault and Weber22):
 $$PROT_{ox}\,(g/min) = 2\cdot 9\times UREA_{urine}\,(g/min), $$
$$PROT_{ox}\,(g/min) = 2\cdot 9\times UREA_{urine}\,(g/min), $$ $$CHO_{ox}\,(g/min) = 4\cdot 59\,\times V\dot CO_{2}-(litres/min) - 3.23\times V\dot O_{2}(litres/min), $$
$$CHO_{ox}\,(g/min) = 4\cdot 59\,\times V\dot CO_{2}-(litres/min) - 3.23\times V\dot O_{2}(litres/min), $$ $$Lipids_{ox}\,(g/min) = - 1\cdot 70\,\times V\dot CO_{2} (litres/min) + 1.70\times V\dot O_{2}(litres/min), $$
$$Lipids_{ox}\,(g/min) = - 1\cdot 70\,\times V\dot CO_{2} (litres/min) + 1.70\times V\dot O_{2}(litres/min), $$where 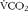 $$V\dot CO_{2} $$ (litres/min) and
$$V\dot CO_{2} $$ (litres/min) and 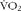 $$V\dot O_{2} $$ (litres/min) were corrected for the volumes of O2 and CO2 corresponding to protein oxidation (1·010 and 0·843 l/g, respectively). UREAurine was measured in urine collected over the entire 4 h of the experiment using a urea assay kit (BioAssay Systems). Energy potentials of 16·3 kJ/g for CHO, 40·8 kJ/g for lipids and 19·7 kJ/g for proteins were used to calculate total EE and the relative contributions of each fuel to total EE(Reference Elia23, Reference Péronnet and Massicotte24). During mild cold exposure, EE relies almost entirely on aerobic metabolism for ATP production, as the relative change in metabolic rate does not exceed twice the RMR (see Haman(Reference Haman25) for a review). AUC was used to represent total EE and fuel selection using middle Riemann sum over 30 min intervals.
$$V\dot O_{2} $$ (litres/min) were corrected for the volumes of O2 and CO2 corresponding to protein oxidation (1·010 and 0·843 l/g, respectively). UREAurine was measured in urine collected over the entire 4 h of the experiment using a urea assay kit (BioAssay Systems). Energy potentials of 16·3 kJ/g for CHO, 40·8 kJ/g for lipids and 19·7 kJ/g for proteins were used to calculate total EE and the relative contributions of each fuel to total EE(Reference Elia23, Reference Péronnet and Massicotte24). During mild cold exposure, EE relies almost entirely on aerobic metabolism for ATP production, as the relative change in metabolic rate does not exceed twice the RMR (see Haman(Reference Haman25) for a review). AUC was used to represent total EE and fuel selection using middle Riemann sum over 30 min intervals.
Determination of shivering intensity
Shivering intensity was measured using surface EMG (Myomonitor, Delsys, Inc.). EMG sites were located on the right side of the upper body on the following muscles: trapezius, pectoralis major and rectus abdominis. These muscles have been shown to contribute significantly to total shivering activity during mild cold exposure(Reference Haman, Legault and Rakobowchuk11, Reference Bell, Tikuisis and Jacobs12). Raw EMG signals were analysed with the use of custom-designed MATLAB algorithms (Mathworks). EMG signals were filtered to remove spectral components below 20 Hz and above 500 Hz, as well as 60 Hz contamination (and associated harmonics). Shivering intensity of individual muscles and their mean (EMGshiv) was determined from root mean square values (RMS) rectified from EMG signals using a 50 ms overlapping window (50 %). Baseline RMS values (RMSbaseline: 5 min RMS average measured before cold exposure) were subtracted from RMS shivering (RMSshiv) as well as RMSmvc values. EMGshiv was then normalised to RMSmvc by using the following equation:
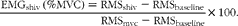 $$\frac {<commr rid="q3" context="main">[Q3]</commr>EMG_{shiv}\,(\%MVC) = RMS_{shiv} - RMS_{baseline}}{RMS_{mvc} - RMS_{baseline}}\times 100. $$
$$\frac {<commr rid="q3" context="main">[Q3]</commr>EMG_{shiv}\,(\%MVC) = RMS_{shiv} - RMS_{baseline}}{RMS_{mvc} - RMS_{baseline}}\times 100. $$AUC was used to represent total shivering intensity and was calculated for all muscles separately. The sum of their mean was used to represent AUC for total shivering intensity.
Statistical analysis
Changes in T rec, T skin, EE and shivering intensity were assessed by a two-way ANOVA for repeated measures to study the effects of time and treatment, as well as their interaction (SPSS for Mac version 18.0; SPSS, Inc.). Significant differences in time were followed up by a Bonferoni post hoc test. Differences in AUC between the two treatments for EE, shivering intensity and fuel selection were determined using a paired t test. The statistical power of our tests was calculated for key parameters (EE, fuel selection and shivering EMG signals), and it ranged from 0·9 to 1·0. Statistical differences were considered significant when P <0·05. All values given are means with their standard errors.
Results
Thermal responses
Changes in thermal responses (T rec and T skin) at baseline and during cold exposure are presented in Fig. 1(a). T rec remained constant at 37·1 ± 0·1°C during the first 120 min for both conditions. No significant difference in T rec was found between the two conditions. T skin decreased by 19 % in both conditions during cold exposure (from 34·1 ± 0·2°C to 28·5 ± 0·5°C in Placebo and from 34·1 ± 0·2°C to 28·7 ± 0·4°C in Green tea; P <0·001). Thermal comfort during cold exposure remained constant between treatments averaging − 2·1 (sem 0·3).

Fig. 1 Changes in (a) core temperature and mean skin temperature, (b) energy expenditure and (c) mean shivering intensity measured in men exposed to mild cold exposure following the ingestion of 1600 mg of epigallocatechin-3-gallate and 600 mg of caffeine (Green tea, ![]() ) or a placebo (Placebo,
) or a placebo (Placebo, ![]() ). MVC, maximal voluntary contraction.
). MVC, maximal voluntary contraction.
Metabolic responses
Changes in EE at baseline and during cold exposure are presented in Fig. 1(b). EE increased by 1·8-fold from baseline values in the Placebo treatment (from 63·2 (sem 1·4) to 116·2 (sem 8·3) J/kg per min) and by 2-fold in the Green tea treatment (from 63·5 (sem 1·2) to 123·7 (sem 7·0) J/kg per min) (P =0·001). However, no overall difference in metabolic rate was found between the treatments during the 180 min cold exposure (P =0·16). Mean heart rate remained unchanged in the cold and was not affected significantly by the treatment (64 (sem 4) beats/min for Placebo v. 62 (sem 3) beats/min for Green tea).
Differences in AUC for EE and fuel selection are presented in Fig. 2(a) and (b). AUC for EE was 10 % higher for the Green tea (23·5 (sem 1·4) kJ/kg × 180 min) than for the Placebo treatment (21·5 (sem 1·6) kJ/kg × 180 min) (P= 0·007). In addition, no significant differences in absolute rate or relative contributions of CHO, lipid and protein oxidation to total EE were observed. Fig. 2(b) presents AUC for all metabolic fuels over the 180 min cold exposure period; the relative contributions of CHO (54 (sem 5) %EE for Placebo and 45 (sem 4) %EE for Green tea; P= 0·23), lipids (34 (sem 5) %EE for Placebo and 39 (sem 5) %EE for Green tea; P= 0·39) and proteins (12 (sem 3) %EE for Placebo and 17 (sem 3) %EE for Green tea; P= 0·41) to total EE were also the same between treatments.

Fig. 2 AUC for (a) energy expenditure (EE), (b) relative contribution of carbohydrates, lipids and proteins to total EE, (c) average shivering intensity and (d) the contribution of respective muscles to total shivering intensity measured over 3 h of mild cold exposure in men following the ingestion of 1600 mg of epigallocatechin-3-gallate and 600 mg of caffeine (Green tea, ■ ) or a placebo (Placebo, □). EMG, electromyography; TRA, trapezius; PEC, pectoralis major; RA, rectus abdominis. Values are means, with their standard errors represented by vertical bars. *Mean value significantly different from that of placebo (P< 0.05).
Shivering responses
Changes in shivering intensity (%MVC) at baseline and during cold exposure are presented in Fig. 1(c). ST intensity increased continuously during cold exposure, but no overall difference was observed between the Placebo and the Green tea treatments. However, when values were compared in the last 30 min, shivering intensity was 26 % higher in the Placebo than in the Green tea treatment (2·9 (sem 0·5) v. 2·1 (sem 0·4) %MVC, respectively; P= 0·03).
Differences in AUC between treatments for overall shivering intensity and for the relative contribution of individual muscles to total ST are presented in Fig. 2(c) and (d), respectively. AUC for overall shivering intensity was 20 % lower for the Green tea (266 (sem 57) %MVC × 180 min) than for the Placebo treatment (332 (sem 69) %MVC × 180 min) (P= 0·01). No significant differences in the AUC was found in the relative contribution of individual muscles to total shivering intensity; it was not different between treatments for trapezius (51 (sem 5) % Shivering for Placebo and 47 (sem 8) % Shivering for Green tea; P= 0·44), pectoralis major (39 (sem 6) % Shivering for Placebo and 43 (sem 8) % Shivering for Green tea; P= 0·55) and rectus abdominis (10 (sem 2) % Shivering for Placebo and 10 (sem 4) % Shivering for Green tea; P= 0·80).
Discussion
The present study is the first to quantify the effects of green tea extracts on changes in shivering intensity and EE during mild cold exposure. When 1600 mg of EGCG and 600 mg of caffeine were ingested at the onset of a 3 h mild exposure to 15°C, total shivering intensity was reduced by approximately 20 %, while EE increased by approximately 10 % compared with the Placebo (Fig. 2). Together, these results confirm both predictions for determining an increase in NST. It also indicates that the contribution of both thermogenic processes to total heat production can be modulated during mild cold exposure. This modulation occurred without modifications in fuel selection and in the relative contribution of individual muscles (Fig. 2).
During cold exposure, the increase in whole-body thermogenic rate is accounted for by the combined activation of NST and ST. Because NST could not be measured directly in the present study, effects of the combined ingestion of EGCG and caffeine on activation of this process were assumed by measuring changes in EE and ST intensity. This indirect methodological approach for quantifying the contribution of NST in cold conditions was suggested almost 40 years ago by Jansky(Reference Jansky26) and recently by Cannon & Nedergaard(Reference Cannon and Nedergaard27). In the present study, the present results showed that during mild cold exposure, the ingestion of a green tea extract not only reduces shivering activity, but also increases overall thermogenic rate (Fig. 2). While this finding clearly indicates an increased contribution of NST to total heat production, this whole-body methodological approach does not allow us to discriminate on the exact pathways involved in providing this additional amount of heat.
Most NST pathways have one commonality; their activation by cold-induced sympathetic stimulation leads to a release in NA, subsequently triggering the activation of cyclic AMP-responsive pathways. Polyphenols found in teas have the potential to inhibit cathecol-O-methyltransferase, the enzyme that breaks down NA in the synaptic cleft(Reference Borchardt and Huber28). Such an increase in synaptic NA creates an overflow into the circulation(Reference Dulloo, Duret and Rohrer8) and the action of NA on target tissues is therefore amplified. Additionally, intracellular cyclic AMP is increased with the inhibition of phosphodiesterase by caffeine(Reference Dulloo, Seydoux and Girardier29). As shown by Dulloo et al. (Reference Dulloo, Seydoux and Girardier30), when NA was released (by adoeralin), EGCG and caffeine increased respiratory rate of in vitro inter-scapular BAT. The effect was even greater when EGCG and caffeine were given together, compared to caffeine alone. Indeed, the thermogenic effect of green tea goes beyond its content in caffeine per se (Reference Dulloo, Duret and Rohrer8). Conversely, EGCG must be given with a minimal amount of caffeine in order to get the desired thermogenic effect(Reference Gregersen, Bitz and Krog-Mikkelsen31). Together, EGCG and caffeine exert a synergistic effect on tissues, such as BAT, contributing to NST.
Even though changes in BAT activity were not measured in the present study, we can still speculate that the combined ingestion of EGCG and caffeine would have stimulated cold-induced NST by increasing the contribution of BAT to total thermogenesis, as suggested by Dulloo et al. (Reference Dulloo, Seydoux and Girardier30). Because BAT is sympathetically innervated(Reference Bartness, Vaughan and Song32, Reference Bartness, Kay Song and Shi33), the inhibition of cathecol-O-methyltransferase by EGCG would increase the action of cold-released NA on adrenergic receptors of BAT. Simultaneously, caffeine would increase intracellular cyclic AMP, therefore activating the release of fatty acids and inducing heat production from activated uncoupling protein 1. Skeletal muscle may also be a viable target for this NST stimulation through Ca2+ cycling. The leakage of Ca2+ in the cytosol (through ryanodine receptors) creates a gradient imbalance and Ca2+-ATPase is activated to restore the equilibrium. The pumping of ions through Ca2+-ATPase requires the hydrolysis of ATP and results in heat dissipation(Reference van Marken Lichtenbelt and Daanen34, Reference Block35). This mechanism could greatly contribute to NST, as skeletal muscle represents approximately 42 % of body mass in adult human subjects(Reference Rolfe and Brown36). Indeed, a study by Astrup et al. (Reference Astrup, Bulow and Madsen37) showed a greater thermogenic contribution from adoeralin-stimulated skeletal muscle compared with BAT. In their study, adoeralin increased oxygen consumption by 40 ml/min in skeletal muscle compared with 10 ml/min in perirenal BAT. Just like cold exposure, adoeralin stimulates the release of NA(Reference Dulloo and Miller38). This thermogenic response could be increased with EGCG by maintaining a higher level of NA in the circulation or by prolonging its action in the synaptic cleft.
Another heat-producing mechanism that could be stimulated by EGCG and caffeine is TAG and NEFA (TAG/NEFA) cycling. The release of NEFA is also stimulated by NA and activated through cyclic AMP. Vallerand et al. (Reference Vallerand, Jacobs and Kavanagh39) speculated that the ingestion of a caffeine–adoeralin mixture in cold conditions (air at 10°C) had an effect on increasing TAG/NEFA cycling. Their results showed an increase in plasma glycerol and TAG in the caffeine–adoeralin condition, with practically unchanged NEFA levels and no increase in lipid oxidation compared with placebo. This goes along the lipolysis-enhancing effect of caffeine in thermoneutral condition(Reference Hetzler, Knowlton and Somani40). Caffeine enhances lipolysis of TAG and the release of NEFA, but not necessarily their oxidation(Reference Acheson, Gremaud and Meirim41). Later, Vallerand et al. (Reference Vallerand, Zamecnik and Jones42) showed that a mild cold exposure increased TAG/NEFA cycling through the activation of the sympathetic nervous system and release of NA. Caffeine given with EGCG has the potential of increasing TAG/NEFA cycling in the cold through the release of NA, thus increasing NST. Possible sites for this to occur would be white adipose tissue(Reference Garofalo, Kettelhut and Roselino43) and the liver(Reference Rolfe and Brown36, Reference Kim, Saidel and Kalhan44). Based on the results obtained in the present study, it is possible to say that NST is potentiated during cold exposure when consuming EGCG and caffeine, but the contributing pathways can only be assumed. When examining the specific mechanisms contributing to cold-induced NST, nuclear imaging techniques as well as indirect calorimetry coupled with stable isotope techniques may provide important estimates. In future studies, it would be of importance to investigate the pathways that contribute to this increase in NST.
In conclusion, the present study shows that the contribution of NST towards total EE can be stimulated by the combined ingestion of EGCG and caffeine in cold-exposed human subjects. When ingested before a mild cold exposure, EGCG and caffeine increased total EE and decreased shivering intensity compared with control. This increase in EE accompanied by a similar decrease in EMG activity demonstrates the replacement of ST by NST without modifying whole-body substrate utilisation. Possible contributing mechanisms to NST could be UCP-1-mediated mitochondrial uncoupling in BAT, Ca2+ cycling in skeletal muscle and TAG/NEFA cycling in white adipose tissue and liver. The ability of EGCG to inhibit cathecol-O-methyltransferase and of caffeine to inhibit phosphodiesterase increases the NA- and cyclic AMP-dependent mechanisms responsible for NST. Although some inter-individual differences were observed, EGCG and caffeine seem to stimulate NST either by a notable increase in EE or a decrease in shivering intensity. The present study provides an experimental approach for human investigations into the potential role of diet and bioactive food ingredients in modulating NST during cold exposure. Stimulating NST pathways in such a manner may not only prove to be important for improving fine motor performance and survival in cold-exposed human subjects, but may also provide important targets in the search of targets for the management of obesity and diabetes.
Acknowledgements
The authors would like to acknowledge the contribution and assistance provided by Marie-Andrée Imbeault, Denis Blondin, Jean-François Mauger and Bernard Pinet. They would also like to thank the subjects of the present study for their participation. The present work was funded by grants and scholarships from the Natural Sciences and Engineering Research Council of Canada (NSERC Canada) to F. H. and C. G. as well as from Ontario Graduate Scholarship (OGS). C. G. conducted the experiments, analysis and participated in the writing of the present paper under the supervision of F. H. The authors have no conflict of interest in relation to the present study.



