


Introduction
Safeguarding and breeding threatened species in captivity is a promising management approach that has been successfully employed for a variety of species threatened with imminent extinction (Conde et al. Reference Conde, Flesness, Colchero, Jones and Scheuerlein2011, Conway Reference Conway2011). However, captive husbandry is expensive, requires appropriate facilities and expertise, and may not always be the most effective conservation option (Dolman et al. Reference Dolman, Collar, Scotland and Burnside2015). Evaluating the performance of captive breeding programmes is therefore essential to make efficient use of sparse conservation resources.
One Critically Endangered species that benefitted greatly from a captive breeding programme is the Northern Bald Ibis Geronticus eremita, an extremely rare colonial cliff-nesting bird that used to occur throughout much of southern Europe, North Africa and the Middle East (Hirsch Reference Hirsch1979, Kumerloeve Reference Kumerloeve1984). Major population declines in the past 150 years resulted in only two isolated subpopulations persisting today - a sedentary population in coastal Morocco, and a migratory population in southern Turkey and Syria (Bowden et al. Reference Bowden, Aghnaj, Smith and Ribi2003, Reference Bowden, Smith, El Bekkay, Oubrou, Aghnaj and Jimenez-Armesto2008, Serra et al. Reference Serra, Abdallah, Assaed, Abdallah, al Qaim, Fayad and Williamson2004). The breeding colony in southern Turkey was first recorded in 1879, and contained about 600–800 pairs in 1954 before a dramatic population decline started in 1959 (Hirsch Reference Hirsch1979, Kumerloeve Reference Kumerloeve1984). The intensive application of the agrochemical DDT by Turkish authorities to suppress both malaria and locusts led to the death of > 600 birds in 1959 and 1960, and was followed by years of almost complete reproductive failure. As a consequence, the Northern Bald Ibis population in southern Turkey declined to only five pairs in 1986 – a decline of 99% in 30–35 years (Hirsch Reference Hirsch1979, Akçakaya Reference Akçakaya1990).
To halt the decline and to safeguard the species in Turkey, a captive breeding station was established in 1977 in close proximity to the original site of the former colony along the river Euphrates, near the town of Birecik in southern Turkey (Hirsch Reference Hirsch1979). Between 1977 and 1988 at least 41 birds were caught, and the project succeeded in attracting remaining wild birds to cliff and artificial breeding sites less vulnerable to disturbance. Unfortunately the captive breeding in adjacent aviaries at the site was not successful (Akçakaya Reference Akçakaya1990). In 1989 only three adult birds returned from migration, of which only one survived to the end of the breeding season and the wild population was considered effectively extinct (Akçakaya Reference Akçakaya1990, Baris Reference Baris1992). Since the 1990s, the captive breeding station was therefore the sole remnant of the Turkish Northern Bald Ibis population. The current population is maintained as a semi-wild population, with birds being allowed to roam freely between March and mid-July, but held in enclosures during autumn and winter to eliminate mortality on migration.
In contrast to the surviving sedentary population in Morocco, the population in Syria and Turkey is migratory and spends the winter in Ethiopia (Serra et al. Reference Serra, Bruschini, Peske, Kubsa, Wondafrash and Lindsell2013). However, of 21 birds tracked with satellite transmitters from Turkey since 2005, none returned to breeding grounds (Taner Hatipoğlu pers. comm.), and persecution along the migratory flyway is the most likely cause of mortality (Serra et al. Reference Serra, Lindsell, Peske, Fritz, Bowden, Bruschini, Welch, Tavares and Wondafrash2015). Since the civil war in Syria began in 2011, the remaining breeding population may have become extinct, but monitoring there is no longer feasible. The poor survival prospects of the Northern Bald Ibis in Syria and along its flyway elevate the importance of the captive breeding station in Turkey. The breeding station in Birecik enables birds to winter in Turkey, and thus functions as a critical lifeline for the last remnant of the eastern population of the species which is known to have distinct genetic characteristics (Broderick et al. Reference Broderick, Korrida and Idaghdour2001, Pegoraro et al. Reference Pegoraro, Föger and Parson2001). The reintroduction of Northern Bald Ibis to areas from where it became extinct > 300 years ago in Austria and Germany is showing promising signs of success (http://waldrapp.eu/index.php/en/project/project-info, accessed 5 Dec 2015), and reintroduction and the creation of alternative semi-wild populations was agreed in the recently updated Action Plan (Bowden Reference Bowden2015) as a beneficial step to reduce the risk of extinction in Turkey. However, such programmes require the selection of suitable habitat, and so far very little information on habitat use and habitat requirements of Northern Bald Ibis in Turkey is available.
Here we evaluate whether the breeding success of the semi-wild population in southern Turkey has improved since the late 1980s (Akçakaya Reference Akçakaya1990), and whether factors associated with wild foraging behaviour, use of supplementary food, and habitat use influence the productivity of individuals. We used monitoring data from the breeding station between 2009 and 2015 to calculate the average number of fledglings produced by each breeding pair in each year. Food is a critical determinant of breeding success, and individuals with unlimited food supply (via supplementary feeding) or foraging in areas where obtaining prey is easier should have an advantage and raise more offspring (Drent and Daan Reference Drent and Daan1980, Martin Reference Martin1987). To test this hypothesis, we used intensive field observations of individually colour-ringed individuals to assess whether breeding productivity in 2013 and 2014 was related to habitat use and foraging success in the wild, and to assess in which habitat foraging success was highest. This information is critical to identify and maintain important habitats around the breeding station, and to explore other areas in southern Turkey with similar habitat configuration that may be suitable for the establishment of a second breeding station. This assessment will therefore be beneficial to reduce the extinction risk of the species in Turkey and will benefit other re-introduction programmes throughout the range of the Northern Bald Ibis.
Methods
Study area
The Northern Bald Ibis breeding station is located along the river Euphrates 3 km north of the town of Birecik in southern Turkey (37°02’N, 37°59’E). The limestone cliffs along the river banks form the natural nesting habitat of Northern Bald Ibis, and the breeding station was constructed around 50 natural caves and ledges in the cliffs (Hirsch Reference Hirsch1979, Akçakaya Reference Akçakaya1990). At least 10 natural and 50 semi-natural (wooden platforms installed on smaller cliff ledges) nest cavities are available within the breeding station, but the natural cliffs provide further suitable breeding habitat adjacent to the fenced breeding station.
During the breeding season between March and mid-July every year the Northern Bald Ibis population is allowed to roam freely. During this time the enclosure fence is opened, and birds can leave and enter the breeding station and forage and roost in surrounding habitats. While the birds breed mostly inside and adjacent to the breeding station, the principal habitats that are regularly used by free-roaming Northern Bald Ibis are: (i) short grasslands and marsh lands along the banks of the river Euphrates, which are grazed by sheep and cattle and used to dispose of human and animal excrement from nearby sewage treatment plants and poultry farms. These grasslands are similar to the habitat used by the species at breeding grounds in Syria (Serra et al. Reference Serra, Abdallah and al Qaim2008) and at wintering grounds in Ethiopia (Serra et al. Reference Serra, Bruschini, Peske, Kubsa, Wondafrash and Lindsell2013), and have generally loose soil, with very short vegetation (< 20 cm) and often extensive areas of bare ground. (ii) A tree nursery with tall shady trees of various native and exotic species, interspersed with irrigated fields of small tree seedlings on fertile dark soil. The seedlings are grown in rows that are spaced 70 cm apart, and allow the birds to forage among rows of seedlings despite the seedlings ranging in height from 10 to 80 cm. (iii) An area of cultivation with arid and irrigated cropland adjacent to embankments of the river Euphrates (hereafter ‘poldered cultivation’); the most common crops are mint, legumes, lettuce, and eggplant. This area has fertile soil and vegetation height and density varies among different crop types, but vegetation height is usually <50 cm in the frequently visited areas.
In 2013, we discovered an additional area in wet meadows adjacent to oxbow lakes on the west bank of the river Euphrates approximately 2 km south-west of Birecik. These meadows are used as pastures and to grow mint and eggplant, and Northern Bald Ibis were regularly observed foraging in this area in 2014.
Supplementary feeding
Throughout the year, including the period when the birds from the breeding station are allowed to roam freely, supplementary food is provided twice a day (at 06h00 and 15h00 hrs local time) at the breeding station. The supplementary food is provided in a large communal area and the amount provided is based on the number of birds in a given year and is composed of 100 g minced beef, 44 g poultry feed, 0.13 boiled eggs (including shell) and 14 g unsalted cheese per bird.
Attendance at supplementary feeding events
Since 2010 all young and adult birds in the population have been marked with coloured rings with a unique alphanumeric inscription allowing individual identification with optical equipment from as far as 200 m away. During each morning and afternoon feeding event, we counted the number of birds attending and recorded the presence of each individual by identifying its unique colour-ring number.
Because some birds may have attended a supplementary feeding event without being recorded, we conducted a repeated observation trial in 2014 during which all individuals were recorded during five independent scans of the foraging flock, and the probability of not detecting a present bird was estimated using an occupancy modelling approach. Because this trial indicated that the probability of not detecting a bird that was actually present was extremely small (< 0.001%), we assumed that attendance at supplementary feeding events was recorded perfectly. For each bird we calculated the average attendance rate as the number of feeding events attended divided by all observed feeding events in a season (n = 63 in 2013 and 50 in 2014).
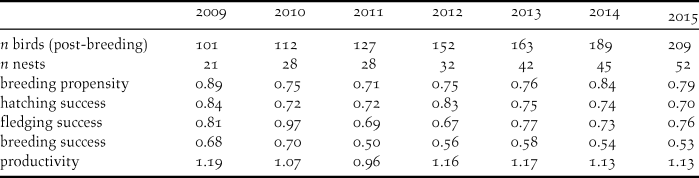
Monitoring of breeding propensity and productivity
Each year between early March and mid-July we inspected all natural and semi-natural nest cavities with binoculars and spotting scopes to assess whether they were occupied by a pair, and whether the pair initiated a nest and laid eggs. We then monitored each recorded breeding attempt at least twice a week to assess how many chicks hatched, and how many fledglings were produced per nest. We calculated breeding propensity as the proportion of nesting pairs that laid an egg, hatching success as the proportion of eggs that hatched, fledging success as the proportion of hatched chicks that fledged, and breeding success as the proportion of breeding pairs that fledged at least one young. Overall productivity was calculated as the average number of fledged young that a pair produced, including pairs that built a nest but did not lay any eggs (Bowden et al. Reference Bowden, Aghnaj, Smith and Ribi2003, Serra et al. Reference Serra, Peske, Abdallah, al Qaim and Kanani2009). We also recorded the identity of the individuals in a pair based on their colour rings, which allowed us to relate breeding performance to individual behaviour observed in the field.
Field observations of habitat use and foraging success
To understand how wild foraging habitat use affects productivity of Northern Bald Ibis, we conducted intensive observations of foraging between March and June 2013 and 2014. Direct observations of birds in the main wild habitats around Birecik (grasslands, tree nursery, poldered cultivation) were carried out with telescope and binoculars three times a week at all times of the day during the whole breeding period.
We first searched for flocks along a fixed route traversing all habitats where the birds had been observed in previous years. Each time a flock was spotted, the size of the flock was recorded along with the individual colour ring codes of all birds. In 2014, we also recorded the specific habitat the flock was using at the time (pasture, manure, fallow, tree seedling, eggplant, mint, lettuce). This information allowed us to assess the relative frequency of occurrence of individual birds at each site to explore whether productivity was affected by more frequent use of certain sites or crop types.
To assess whether certain habitats are used primarily for certain behaviours, we employed instantaneous scan sampling to quantify the behaviour of Northern Bald Ibis in the various wild habitats around Birecik (Altmann Reference Altmann1974). At each scan, the number of individuals in each of five behavioural categories was counted, based on what an individual was doing instantaneously at the moment of observation. We distinguished between foraging, locomotion, comfort behaviour (preening, bathing), resting, and other behaviour (including social interactions, scanning, nest building etc.). To evaluate whether foraging success differed between habitats, we observed the foraging behaviour of two randomly selected birds in each flock for 15 consecutive minutes, and counted the number of unsuccessful bill-probes until successful prey capture, which was discerned by observing swallowing behaviour. Northern Bald Ibis feed on a wide variety of large invertebrates (Coleoptera, Orthoptera, especially Gryllotalpa gryllotalpa, scorpions) frogs, and lizards (Lacerta cappodocia and Acanthodactylus boscianus), and capture of these prey items is generally observable from a distance, but identification of prey items is usually not possible.
Data analysis
We examined whether breeding propensity and productivity could be explained by attendance at feeding events at the breeding station, by the frequency of observation in various habitats, and by individual foraging success observed in wild habitats with suites of generalised linear mixed models (GLMM) using year and nest as random factors to account for environmental stochasticity and serial autocorrelation of the two partners in a nest (Bolker et al. Reference Bolker, Brooks, Clark, Geange, Poulsen, Stevens and White2009). We used a similar approach to examine whether habitat type affected individual foraging success. For each specific question, we constructed a series of plausible candidate models and used an information-theoretic approach to identify the covariates explaining the most variation in the response variable (Burnham and Anderson Reference Burnham and Anderson2002).
To assess whether the attendance at feeding events affected any of the breeding parameters, we fit similar GLMMs with breeding propensity, number of eggs laid, number of eggs hatched, and number of chicks fledged as response variables, and we present the parameter estimates (± 1 standard error) for the proportion of feeding events attended.
Our candidate models for productivity included the relative frequency with which individual birds were observed at the feeding station, the tree nursery, the poldered cultivation, or the grasslands. We also included models that combined the frequency of observation in the three wild habitats, and a null model that only included the random effects.
Detailed crop information was only available from 2014, and we therefore used the subset of data collected in 2014 to examine whether productivity was related to the frequency with which individuals were observed in different crop types. For this analysis we fitted eight candidate models exploring the effects of pasture, manure, fallow fields, eggplant, mint, lettuce, and tree seedlings on productivity. All breeding propensity models used a logit link and a binomial error structure with the incidence of breeding (yes/no) as response variable, all productivity models were fitted using the number of fledged young as response variable with a log link and a Poisson error structure.
We used a similar Poisson model structure to relate the number of pecks necessary before prey capture to habitat and crop type (foraging success). For this analysis we first examined whether foraging success differed among the four main habitats in our study area using data from 2013 and 2014. We then used only data from 2014 for which more detailed crop type data were available to compare four candidate models that assumed that foraging success was either constant or varied by crop type, or by increasingly coarser classifications, namely habitat (marsh land, tree nursery, poldered cultivation, meadow), and land use (grassland, cultivated land). We included the individual bird as random intercept to account for serial autocorrelation of repeated observations of the same individual (Bolker et al. Reference Bolker, Brooks, Clark, Geange, Poulsen, Stevens and White2009).
Finally, we examined whether the proportion of birds observed foraging varied among the wild habitat and land-use types. For each flock encountered in 2014 we used the number of birds observed foraging during instantaneous scan sampling as a binomial response variable with the total number of birds in the flock as the number of trials. We fitted 10 competing GLMs (without random effects) which assumed that the proportion of birds foraging varied by month, by crop type, habitat, land use, site, or by both temporal (month) and spatial (crop type, habitat, land use or site) variables.
For all analyses, we present a model selection table showing the amount of support for each fitted model, and we present the parameter estimates and standard errors for the most parsimonious models based on the second-order Akaike’s Information Criterion (AIC c ).
Results
The Northern Bald Ibis population in Birecik increased from 40 birds in 2000 to 209 in 2015 (Table 1). We recorded a total of 248 breeding attempts in Birecik between 2009 and 2015, and found that on average breeding productivity was 1.12 fledglings/pair (including pairs that did not breed; Table 1), but breeding pairs raised on average 1.42 fledglings per pair (range 1.34–1.54; excluding pairs that did not breed). Across the seven years, breeding propensity was on average 78.5%, and 58.3% of pairs that laid eggs succeeded in raising at least one offspring (breeding success; Table 1). Both hatching and fledging success were > 75% (Table 1). Despite being provided with food, breeding individuals attended on average only 35.6% of supplementary feeding events in 2013 and 2014 (range 0.8–70%, n = 155).
Table 1. Summary of breeding parameters of the semi-wild Northern Bald Ibis population at the breeding station in Birecik, southern Turkey, from 2009 to 2015. See text for definition of demographic parameters. Note that the total number of birds is based on the post-breeding recapture and includes juvenile birds from the respective year.
Non-breeding adults had a slightly higher but also hugely variable attendance rate at feeding events (mean attendance: 44.6%, range 0–90%; P < 0.001), but there was no correlation between the proportion of feeding events attended at the breeding station and productivity for the 98 individuals for which information was available. Attendance at supplementary feeding events at the breeding station had no effect on the number of eggs laid (b = -0.07, SE = 0.51, P = 0.88), the number of eggs that hatched (b = 0.37, SE = 0.60, P = 0.53), or the number of fledglings produced (b = 0.06, SE = 0.69, P = 0.93).
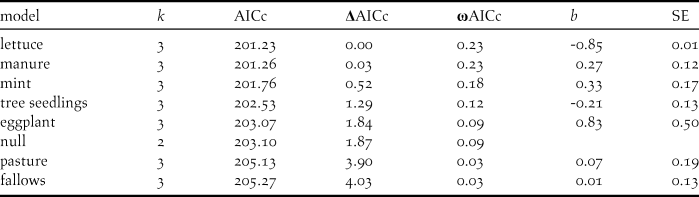
Among our seven candidate models relating productivity to the frequency of occurrence in various wild habitats, there was strong support for all models that included the number of sightings at the tree nursery and in poldered cultivation (Table 2). Birds that were frequently observed at the tree nursery appeared to raise fewer offspring (b = -0.25, SE = 0.07, P < 0.001), while birds that were more frequently observed in the poldered cultivation appeared to raise more offspring (b = 0.25, SE = 0.10, P = 0.009). Breeding propensity followed a similar pattern, with birds that were frequently observed at the tree nursery having a lower probability to breed (b = -0.27, SE = 0.09, P = 0.002), while birds that were more frequently observed in the poldered cultivation (b = 1.35, SE = 0.36, P < 0.001) and in croplands (b = 0.24, SE = 0.10, P = 0.01) were more likely to breed.
Table 2. Model selection summary of seven candidate models explaining the number of fledglings produced by Northern Bald Ibis at the breeding station in Birecik, southern Turkey, in 2013 and 2014, in relation to individual’s relative frequency of occurrence in different wild foraging habitats or at supplementary feeding events.

In 2014, the subset of data for which we had detailed information on crop type (75 individuals), there was some support for models that included the number of sightings in lettuce fields, manure, and mint fields (Table 3). Birds that were frequently observed in lettuce appeared to raise fewer offspring (Table 3). Conversely, birds that were more frequently observed in mint or manure raised more offspring (Table 3). There was little evidence that presence in certain crop types was associated with breeding propensity, which was best explained by frequency of observation at the tree nursery (see above, all other effects P < 0.05).
Table 3. Model selection summary of eight candidate models explaining the number of fledglings produced by Northern Bald Ibis at the breeding station in Birecik, southern Turkey, in 2014, in relation to individual’s relative frequency of occurrence in different crop types. b indicates the estimated effect size of the crop type on productivity, SE is the standard error of the estimate.
To examine the potential cause of variation in productivity based on variation in frequency of occurrence patterns in different habitats, we tested whether foraging success varied by habitat based on 602 focal observations of 61 individuals. There was strong evidence for differences in foraging success among the main habitats (Figure 1), with the highest number of pecks required in the tree nursery (b = 2.43, SE = 0.13, P < 0.001), followed by poldered cultivation (b = 1.76, SE = 0.14, P < 0.001), and croplands (b = 0.85, SE = 0.13, P < 0.001). Because both the poldered cultivation and the cropland habitats harbour a diverse suite of crop types, we used a subset of 233 observations of 46 individuals in 2014 to examine whether foraging success varied also among crop types. This model received overwhelming support from the data (Table 4), and indicated that foraging success was highest in manure and mint plantations, and very low on fallow grassland (Figure 2). We were not able to quantify foraging effort in lettuce fields, because birds were observed to rip out and consume lettuce leaves, and discerning the capture of animal prey was therefore impossible.
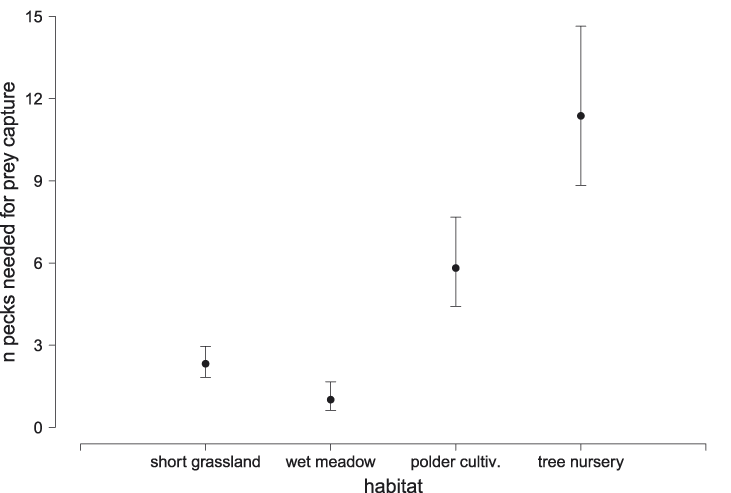
Figure 1. Mean foraging success (number of pecks required for successful prey capture, ± 95% confidence intervals) of Northern Bald Ibis foraging in different habitat types near Birecik in southern Turkey in 2013 and 2014.
Table 4. Model selection summary of four candidate models explaining variation in foraging success by Northern Bald Ibis in different wild habitats near Birecik, southern Turkey, in 2014. Habitats were grouped either by crop type, or by coarser classifications (habitat, land use).
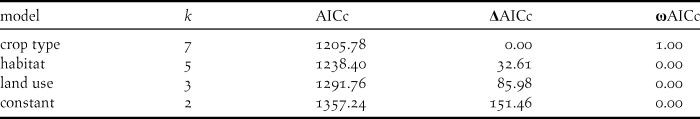
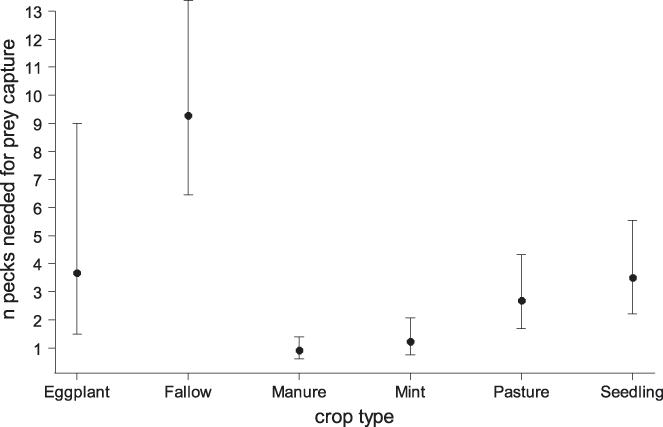
Figure 2. Mean foraging success (number of pecks required for successful prey capture, ± 95% confidence intervals) of Northern Bald Ibis foraging in different crop types near Birecik in southern Turkey in 2014.
We quantified the behaviour of 3,331 birds in 149 flock observations with flocks ranging from one to 140 birds in size to understand whether habitats were actually used for foraging. There was strong evidence that observed behaviour in the wild changed dramatically over the season, with a much higher proportion of birds observed foraging in March and gradually fewer birds as the season progressed (Table 5, Figure 3). Besides the temporal changes, there was also overwhelming support for variation in behaviour among crop types (Table 5): the proportion of birds observed foraging was highest in eggplant, and lowest in mint (Figure 3).
Table 5. Model selection summary of nine candidate models explaining variation in the proportion of foraging behaviour within Northern Bald Ibis flocks in different wild habitats near Birecik, southern Turkey, in 2014. Habitats were grouped either by crop type, or by coarser classifications (habitat, land use).
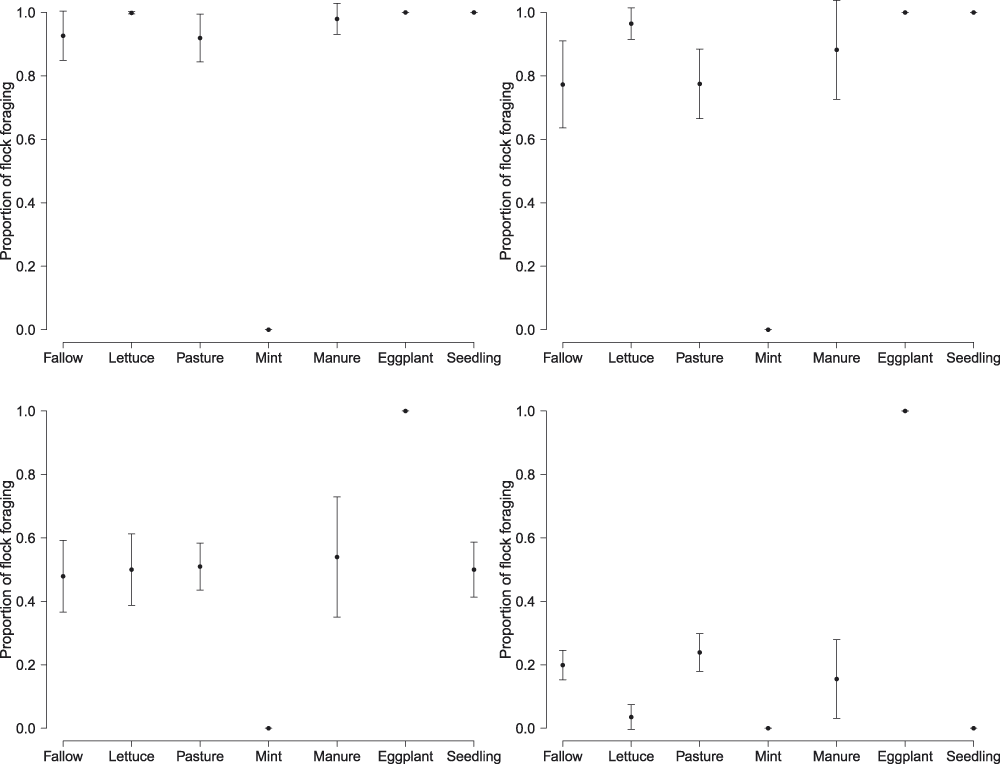
Figure 3. Average proportion of Northern Bald Ibis flocks foraging across the four months of the breeding season and separated by seven different wild crop types.
Discussion
The Northern Bald Ibis population in Birecik has recovered from its nadir in the 1980s, and productivity at the breeding station was higher from 2009 to 2015 than when the breeding station was originally founded (Akçakaya Reference Akçakaya1990). The mean productivity of 1.12 fledglings/pair is similar to the productivity found in the wild population in Morocco, where productivity ranged from 0.39 to 2.27 fledglings/pair depending on rainfall and other factors (Bowden et al. Reference Bowden, Smith, El Bekkay, Oubrou, Aghnaj and Jimenez-Armesto2008). The number of fledglings per breeding pair (1.45) was only slightly lower than the past mean of the wild population in Birecik (1.6 young per breeding pair in 1977–1988; Akçakaya Reference Akçakaya1990), or of the population in Syria (1.72 chicks per breeding pair in 2002–2007; Serra et al. Reference Serra, Peske, Abdallah, al Qaim and Kanani2009). Given the reduced mortality of the semi-wild birds, the productivity is so far sufficient to ensure steady population growth (Table 1). The improved productivity compared to the early period of the breeding station is likely due to improved husbandry and supplementary diet over that period, the provision of more protected breeding ledge sites, and the ability for birds to forage freely in surrounding habitats.
We provided the first examination of the wild foraging ecology of the species around Birecik, and demonstrated that the birds breeding at the breeding station attend only a fraction of the supplementary feeding events. We also provide some evidence that the frequency of attendance at feeding events was not a determinant of breeding success. Instead, we found that birds frequently foraging in poldered cultivation and on grasslands covered in manure appeared to have both greater breeding propensity and success. This increase in breeding performance is likely a consequence of the excellent feeding conditions, as birds on mint fields and on grasslands covered in manure required on average many fewer pecks to successfully capture prey than birds foraging in other wild habitats. We therefore speculate that the Northern Bald Ibis population around Birecik does not rely entirely on supplementary food, but that the availability of suitable foraging habitat, such as irrigated mint fields and grasslands covered in manure, allows the population to forage and breed successfully. Despite the use of wild foraging habitats, the supplementary food may still be an important contribution to the basic energy requirements of the semi-wild birds, but more intrusive investigations would be necessary to quantify what proportion of energy is derived from wild versus supplementary food.
Besides the open grasslands, crops, and cultivated fields, Northern Bald Ibis in Birecik also use tree seedling fields at a local tree nursery (Akçakaya Reference Akçakaya1990). However, we found that foraging success in those fields was extremely low, and that birds more frequently observed at the tree nursery had on average lower breeding propensity and productivity than birds more frequently observed elsewhere. We speculate that the tree nursery is not a preferred foraging habitat, but that the tall trees offer shady and moist habitat that allows the birds to rest in cooler conditions during the middle of the day, when average temperatures in Birecik rise to 30°C in the shade. Because our study was correlational, we are unable to determine whether the frequency of occurrence at the tree nursery is a cause or a consequence of low breeding propensity and success: individuals that did not breed or failed in their breeding attempt may have used the tree nursery as a preferred resting habitat once they had been relieved of the energetic demands of breeding.
Although the use of various foraging habitats around Birecik has been known for decades (Hirsch Reference Hirsch1979, Akçakaya Reference Akçakaya1990), our study is the first to indicate that the combination of suitable wild foraging habitat and safe breeding sites contributes to a highly productive breeding environment. Despite being provided with sufficient supplementary food, the Northern Bald Ibises in Birecik appear to acquire much of their nutrition from wild foraging habitats, and the availability of such productive foraging habitat would be essential if new breeding stations are to be created for the species in its former range in southern Turkey. The observed foraging habitat used around Birecik has similar properties to the foraging areas found for the species elsewhere, where sparsely vegetated fallow land, cultivation, or semi-natural steppe grassland were the most preferred foraging habitats due to their higher level of prey availability (Bowden et al. Reference Bowden, Smith, El Bekkay, Oubrou, Aghnaj and Jimenez-Armesto2008, Serra et al. Reference Serra, Abdallah and al Qaim2008, Lindsell et al. Reference Lindsell, Shehab and Anderson2011). Such cultivations and grasslands have been used for foraging around Birecik in the past (Hirsch Reference Hirsch1979, Akçakaya Reference Akçakaya1990), but there is no information on whether these habitats have changed as a consequence of pesticide use and whether they retain similar prey densities today as they did several decades ago. Further work is required to quantify prey availability and identify factors that lead to the observed differences in foraging success among the different foraging habitats around Birecik. An improved understanding of the factors that make foraging sites suitable and increase foraging success and ultimately productivity could further facilitate the identification of alternative suitable sites in Turkey where new Northern Bald Ibis colonies may be established. Similarly, the contamination of natural food with pesticides, and the contribution of the supplementary food to the overall energy balance of the semi-wild Northern Bald Ibis population could be examined to assess whether there is a critical need to provide supplementary food at the breeding station.
We only observed foraging and behaviour within 15 km of the breeding station, which may not adequately cover the entire foraging range of the species. In the 1970s, the birds were assumed to forage on untilled fields up to 70 km from the breeding location (Hirsch Reference Hirsch1979), but it is possible that such long foraging flights may have been due to intraspecific competition of the much larger colony in those years (Ashmole Reference Ashmole1963, Brown and Brown Reference Brown and Brown2001, Oppel et al. Reference Oppel, Beard, Fox, Mackley, Leat, Henry, Clingham, Fowler, Sim, Sommerfeld, Weber, Weber and Bolton2015). However, even relatively small colonies in Morocco and in Syria display regular foraging flights to highly suitable fields up to 20–35 km from the colony (Bowden et al. Reference Bowden, Smith, El Bekkay, Oubrou, Aghnaj and Jimenez-Armesto2008, Serra et al. Reference Serra, Peske, Abdallah, al Qaim and Kanani2009, Lindsell et al. Reference Lindsell, Shehab and Anderson2011). In contrast to the birds in Morocco, the colony at Birecik benefits from a permanent source of fresh water for drinking (river Euphrates), which may permit birds to explore a wider area for foraging within a short travel time from a large water source (Bowden et al. Reference Bowden, Smith, El Bekkay, Oubrou, Aghnaj and Jimenez-Armesto2008, Smith et al. Reference Smith, Aghnaj, El Bekkay, Oubrou, Ribi, Armesto and Bowden2008). Although it is unknown how many birds the wild foraging grounds around Birecik can support, we predict that if the size of the population continues to recover to former levels, the foraging range may expand and the birds may start utilising new and possibly more distant foraging locations. Future monitoring may require tracking birds using GPS or satellite transmitters to identify distant foraging locations (Lindsell et al. Reference Lindsell, Serra, Pëke, Abdullah, al Qaim, Kanani and Wondafrash2009) and ensure that these locations remain suitable.
Based on scan sampling observations of flocks encountered in wild foraging areas we documented a marked decrease in the proportion of time spent foraging throughout the season. This pattern is counterintuitive for two reasons: as chicks hatch and grow, the energetic demands on parents increase, and birds feeding chicks should therefore increase their foraging investment (Drent and Daan Reference Drent and Daan1980); in addition, prey availability may decline sharply during the season (Lindsell et al. Reference Lindsell, Shehab and Anderson2011), which would require an increase in foraging effort to maintain similar levels of energy intake. We speculate that the apparent decrease in foraging behaviour during the season in our data may have been generated by an increasing proportion of failed breeders making up our sample of observed birds later in the season, rather than an actual decline in foraging effort during the breeding season.
In summary, we found that the semi-wild population of Northern Bald Ibis at Birecik has similar productivity as wild populations elsewhere, and that birds frequently visiting highly productive foraging areas are able to raise more offspring. Although the population is currently dependent on captivity for the migration and winter period to avoid the extremely dangerous migration through the Middle East to Africa (Serra et al. Reference Serra, Lindsell, Peske, Fritz, Bowden, Bruschini, Welch, Tavares and Wondafrash2015), the generally low attendance at supplementary feeding events indicates that the population may be self-sustainable if suitable foraging habitat and safe nesting sites are provided during the breeding season and non-natural mortality during migration could be eliminated.
Acknowledgements
This work was supported by generous funding from the Prince Albert II Monaco Foundation and BirdLife International. We appreciate the support of the Ministry of Forestry and Water Affairs, Şanlıurfa Provincial Directorate, Birecik District Governorship, Birecik Directorate of District Food Agriculture and Livestock, Birecik Municipality, Association for Nature, Culture and Life, Ahmet Aloğlu, Ferdi Akarsu, Özge Balkız, İ.Turan Çetin, Mustafa Çulcuoğlu, Taner Hatipoğlu, Nuri Özbağdatlı, Lubomir Peske, Jose Tavares and Abdullah Yaşar. We appreciate the constructive comments of Jeremy Lindsell, Phil Atkinson, Gianluca Serra and an anonymous reviewer on an earlier version of the manuscript.








