


Introduction
Geographical range size is a basic criterion for determining if a species faces a heightened risk of extinction (IUCN 2001). With large-scale changes in land use, it is generally easier to eliminate a species with a narrow range than a species with a large range (Sekercioglu et al. Reference Sekercioglu, Schneider, Fay and Loarie2008). The distribution of Jankowski’s Bunting Emberiza jankowskii is restricted to China, Russia and North Korea, and it has been listed as ‘Endangered’ on the IUCN Red List of Threatened Species since 2010 due to its small distribution and rapid population decline (BirdLife International 2012). The main threat to this species appears to be the conversion of its habitat into agricultural land, pasture and sometimes even forestry (BirdLife International 2016). Nevertheless, recent information on its population status is lacking and scattered in the literature (e.g. Jiang et al. Reference Jiang, Gao, Lei, Wan, Zhao and Wang2008, Wang et al. Reference Wang, Jiang and Gao2010, BirdLife International 2016).
Historically, Jankowski’s Bunting bred in Inner Mongolia, Heilongjiang, western and eastern Jilin Province in China, extreme north-eastern North Korea and southern Primorye in Russia (BirdLife International 2001). However, the range of Jankowski’s Bunting has been drastically contracting for many years. Before the early 1970s, Jankowski’s Bunting previously bred in the extreme southern regions of the Russian Far East (Vorob’ev Reference Vorob’ev1954, Litvinenko and Shibaev Reference Litvinenko and Shibaev1966, Panov Reference Panov1973), but there has been no record of the species in Russia for more than 40 years. Similarly, there has been no information reported on the species in North Korea for many years (BirdLife International 2001).
In China, Jankowski’s Bunting has disappeared from a number of breeding sites, such as Dongning (site 32 in Figure 1), Tumen (site 52) and Tuquan (site 14). A small population at Dagang in Jilin was reported in 2010 (Wang et al. Reference Wang, Jiang and Gao2010). Gao (Reference Gao2002) reported that a small population bred at Tumuji and Maanshan of Zhalaite Qi in Inner Mongolia, though fewer than 30 individuals were detected (Wang et al. Reference Wang, Jiang and Gao2010). Breeding populations at Xiergen and Xinjiamu of Keerqinyouyizhong Qi in Inner Mongolia were found in 2008, and the population sizes reportedly fluctuated around approximately 50 individuals (Wang et al. Reference Wang, Jiang and Gao2010).
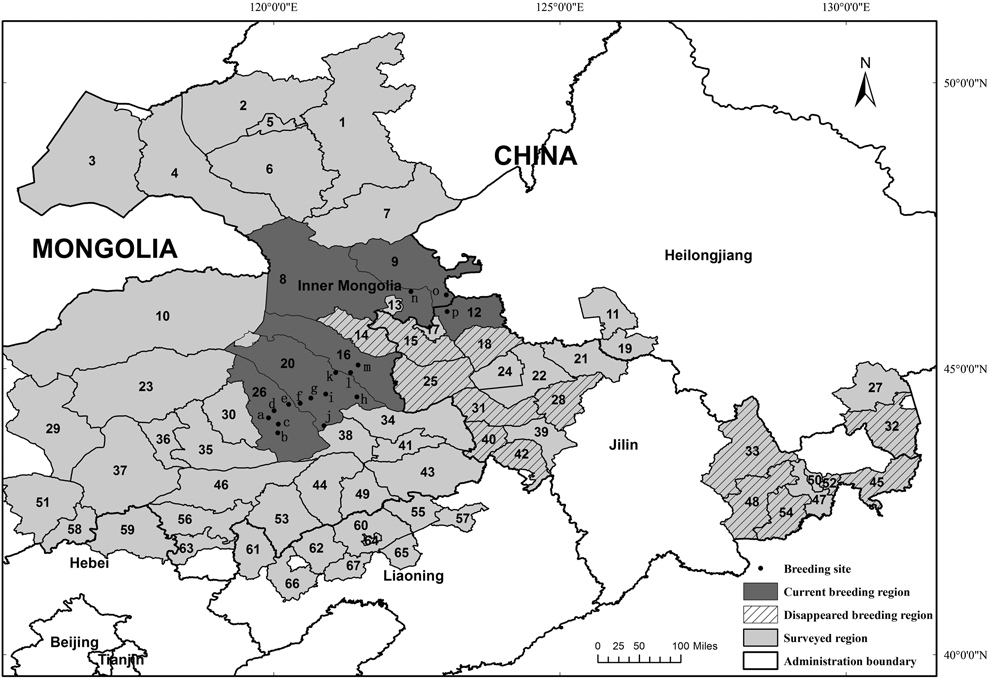
Figure 1. The current and historical breeding distribution of Jankowski’s Bunting Emberiza jankowskii in China.
(1) Yakeshi City; (2) Chenbaerhu Qi; (3) Xinbaerhuyou Qi; (4) Xinbaerhuzuo Qi; (5) Hailaer City; (6) Ewenkezuzizhi Qi; (7) Zhalantun City; (8) Keerqinyouyiqian Qi; (9) Zhalaite Qi; (10) Dongwuzhumuqin Qi; (11) Zhaodong City; (12) Zhenlai County; (13) Wulanhaote City; (14) Tuquan County; (15) Taonan City; (16) Keerqinyouyizhong Qi; (17) Baicheng City; (18) Da’an City; (19) Shuangcheng City; (20) Zhalute Qi; (21) Songyuan City; (22) Qianguoerluosi County; (23) Xiwuzhumuqin Qi; (24) Qian’an County; (25) Tongyu County; (26) Alukeerqin Qi; (27) Muling City; (28) Nong’an County; (29) Xilinhaote City; (30) Balinzuo Qi; (31) Changling County; (32) Dongning County; (33) Dunhua City; (34) Keerqinzouyizhong Qi; (35) Balinyou Qi; (36) Linxi County; (37) Keshiketeng Qi; (38) Kailu County; (39) Gongzhuling City; (40) Shuangliao City; (41) Tongliao City; (42) Lishu County; (43) Keerqinzuoyihou Qi; (44) Naiman Qi; (45) Huichun City; (46) Wengniute Qi; (47) Longjing City; (48) Antu County; (49) Kulun Qi; (50) Yanji City; (51) Zhenglan Qi; (52) Tumen City; (53) Aohan Qi; (54) Helong City; (55) Zhangwu County; (56) Chifeng City; (57) Faku County; (58) Duolun County; (59) Weichang County; (60) Fuxinmengguzuzizhi County; (61) Jianping County; (62) Beipiao City; (63) Kalaqin Qi; (64) Fuxin City; (65) Heishan County; (66) Chaoyang County; (67) Yi County. Breeding Site: a) Bayanhua; b) Tianshan; c) Xinmin; d) Kundu; e) Saihantala; f) Bayantala; g) Xiangshan; h) Daolaodu; i) Lubei; j) Wulijimuren; k) Gahaitu; l) Xiergen; m) Bayanhushu; n) Eergetu; o) Tumuji; p) Dagang.
Jankowski’s Bunting breeds in a variety of open habitats at low elevations. Historically, it bred in three different habitat types, defined by their plant assemblages. The first habitat consisted of low hills, meadows or coastal plains with xerophilous grass species and scattered young oak bushes; this habitat was found in eastern Jilin Province and the surrounding areas (Yamashina Reference Yamashina1957, Stresemann and Portenko Reference Stresemann and Portenko1981). The second consisted of steppe-like habitats in Xianghai that were composed of mixed shrubs of Prunus sibirica, Ulmus macrocarpa and Ulmus pumila (Gao Reference Gao2002); this also included afforested habitats with young Pinus sylvestris mongolica and Populus spp. in Changling (Zhao et al. Reference Zhao, Nickel and Groh1994). The third habitat consisted of Mongolian steppes or low hills dominated by grass species, such as Filifolium sibiricum, Stipa baicalensis and Spodiopogon sibiricus, interspersed with Prunus sibirica (Gao Reference Gao2002, Gao et al. Reference Gao, Wang and Sun2003). Currently, known populations are all found in the Mongolian steppes. Such habitat specialisation limits the distribution range of Jankowski’s Bunting and makes it vulnerable to habitat loss caused by human activities. For example, the habitat in Inner Mongolia is increasingly used for pasture and crops, so the bunting’s population is likely to decrease even further (BirdLife International 2001).
Jankowski’s Bunting has vanished from most of its historic breeding sites during the past 40 years (Gao Reference Gao2002, Jiang et al. Reference Jiang, Gao, Lei, Wan, Zhao and Wang2008, Wang et al. Reference Wang, Jiang and Gao2010). This bird was once abundant in the border areas between Jilin Province and Inner Mongolia, and some sites had encounter rates higher than 10 birds per hour. Fu and Chen (Reference Fu and Chen1966) suggested the areas surrounding the Daxingan Mountains (Greater Khingan Mountains) should support many Jankowski’s Buntings given the similar vegetation and climatic elements. With small, discrete sites (i.e. Xiergen and Xinjiamu) found as suitable habitats in 2008 (Wang et al. Reference Wang, Jiang and Gao2010), we suspect that undiscovered populations may exist elsewhere. However, no systematic surveys have been conducted, so the population status and any trends related to this species cannot be accurately assessed (BirdLife International 2016). Therefore, we developed this study with the objective of assessing the current breeding distribution and population size of Jankowski’s Bunting within China, which would allow us to provide a more accurate assessment of this species’ conservation status.
Methods
Field investigations
The breeding habitat of Jankowski’s Bunting was surveyed between late April and early September, from 2011 to 2016. The survey covered Inner Mongolia and Jilin, Heilongjiang, Liaoning and Hebei Provinces (Fig. 1), which included historical breeding distribution areas, wintering sites, and regions adjacent to historical breeding distribution areas.
First, we visited local forestry bureaus or consulted with local residents for information on the distribution of habitats with similar vegetation structure to the historical breeding sites of the species reported in the literature (Yamashina Reference Yamashina1957, Stresemann and Portenko Reference Stresemann and Portenko1981, Zhao et al. Reference Zhao, Nickel and Groh1994, Gao Reference Gao2002, Gao et al. Reference Gao, Wang and Sun2003). If detailed information on potential habitats (such as location, patch size, natural or artificial) was obtained, we visited the sites to verify whether Jankowski’s Buntings were breeding there. A site was classified as a breeding ground strictly based on the presence of Jankowski’s Bunting (determined via call or nest), and each site was checked several times in different breeding seasons. Once a breeding site was identified, we conducted an extended survey centred on the site. If a region had a similar vegetation structure as that of historical breeding sites but no Jankowski’s Bunting were found during the first investigation, we repeated the survey at that site at least once more in subsequent years. Precise locations of breeding sites were recorded using a hand-held GPS (Garmin eTrex Summit HC, Garmin Int. Inc., Kansas). For small suitable habitats, such as Dagang and Eertuge, which are surrounded by farmland or forest, the boundary of the patch was distinct, we directly measured the patch size using GPS.
Range size estimation
In China, the distribution of Jankowski’s Bunting is currently restricted to Mongolian steppes, where Siberian apricot Prunus sibirica and grass density are important vegetation characteristics that could be helpful as reference signatures for classification of remote sensing-derived images. Therefore, we applied visual interpretation of Landsat imagery, a widely used method in land-cover and vegetation mapping (Wilson and Sader Reference Wilson and Sader2002, Lillesand et al. Reference Lillesand, Kiefer and Chipman2014), to evaluate breeding areas of Jankowski’s Bunting. First, we acquired Landsat 7 Enhanced TM plus (ETM+) and panchromatic Landsat 8 OLI satellite images (path/row 121/28, 121/29) from May and September of 2014, with 30-m and 15-m resolution, respectively. After georeferencing, original DN values of these images were converted into reflectance ratios, and panchromatic and multispectral data were merged in ENVI 5.0 to yield a pan-sharpened false colour composite for high-resolution image interpretation (Jiang et al. Reference Jiang, Strittholt, Frost and Slosser2004, Thomas et al. Reference Thomas, Ranchin, Wald and Chanussot2008). Then we performed a Gaussian contrast transformation during the image enhancement process, along with different RGB band combinations (bands 4, 3 and 2; bands 3, 4 and 5; bands 5, 4 and 1), to further delineate vegetation cover and highlight forest, shrub and grass. These steps were repeated as many times as required to assign a final classification. Additionally, we obtained a GIS map of land-use cover (1:100,000 scale; 2012) from the National Science & Technology Infrastructure Center (http://www.geodata.cn) to help discern differences in plant structure at a coarser level, and we combined field GPS sites and trajectories of line transects to create ancillary datasets that improved the accuracy and quality of the final land-cover classification. The above work was carried out in ENVI 5.0 (ENVI 2011), and habitat areas were calculated in ArcGIS 10.1 (ESRI 2012).
Population size estimation
We estimated the population density of Jankowski’s Bunting using line transects, a commonly used method in bird abundance assessments (Bibby et al. Reference Bibby, Burgess, Hill and Mustoe2000). Bird surveys were conducted from late May to early June in 2014 and 2016, covering currently known breeding ranges. A total of 99 line transects, covering a total of 474.13 km, were established in 2014. In 2016, we resurveyed another six line transects, where bird density fluctuated heavily due to habitat degradation caused by grazing. We established paralleled line transects in intervals of 500 to 1,000 m at each patch, the lengths of which were roughly determined in proportion to the actual patch size. All surveys were completed by six experienced observers who used binoculars for visual observations. All surveys were carried out between 05h00 and 09h00 and 15h00–18h00 and only when light and weather conditions were favourable for observation. The observers moved at a speed of 1–1.5 km hr-1 and stopped occasionally to watch and listen. Since Jankowski’s Buntings already bred by the time of the surveys, the males were usually singing on perches, and the females were incubating and were not easily found. At each patch, we recorded the number of male individuals detected by sight or call within a fixed distance of 0.05 km on either side of the central transect line, and the population size was estimated by multiplying the number of observations by two (Jiang et al. Reference Jiang, Gao, Lei, Wan, Zhao and Wang2008). In addition to males that were seen or heard, occasional flocks with three or more Jankowski’s Buntings were also recorded during monitoring visits.
To estimate the population size, we first estimated bird density in each patch as follows: D=n/2ωL, where D is population density, n is the count number, L is the length of the transect, and ω is 0.05 km (Buckland et al. Reference Buckland, Marsden and Green2008). The number of birds in the population was then calculated by multiplying bird density (mean ± standard deviation) by the total size (A) of the breeding area: N =A*D.
We reported the results of our investigation at the town level. A schematic plot on the survey sites and distribution ranges of Jankowski’s Bunting was created in ArcGIS 10.1. To clearly indicate the current distribution boundary of the Jankowski’s Bunting, the 17 previously surveyed counties/cities that were identified as unsuitable for the species were also shown on the map.
Results
In the study, we confirmed that Jankowski’s Bunting has disappeared from most of its historical breeding regions in China (Figure 1), which included Dongning of Heilongjiang, Yanbian, Siping, Taonan, Tongyu and Songyuan of Jilin, and Tuquan of Inner Mongolia. For the remaining populations identified prior to 2010, no breeding individuals were detected at Xinjiamu or Maanshan, and the population at Tumiji was unstable (only one pair was observed in 2012, and no birds were recorded in 2014); however, populations at Dagang and Xiergen still persisted. Encouragingly, 13 new breeding localities were discovered in Inner Mongolia during this survey (Figure 2).
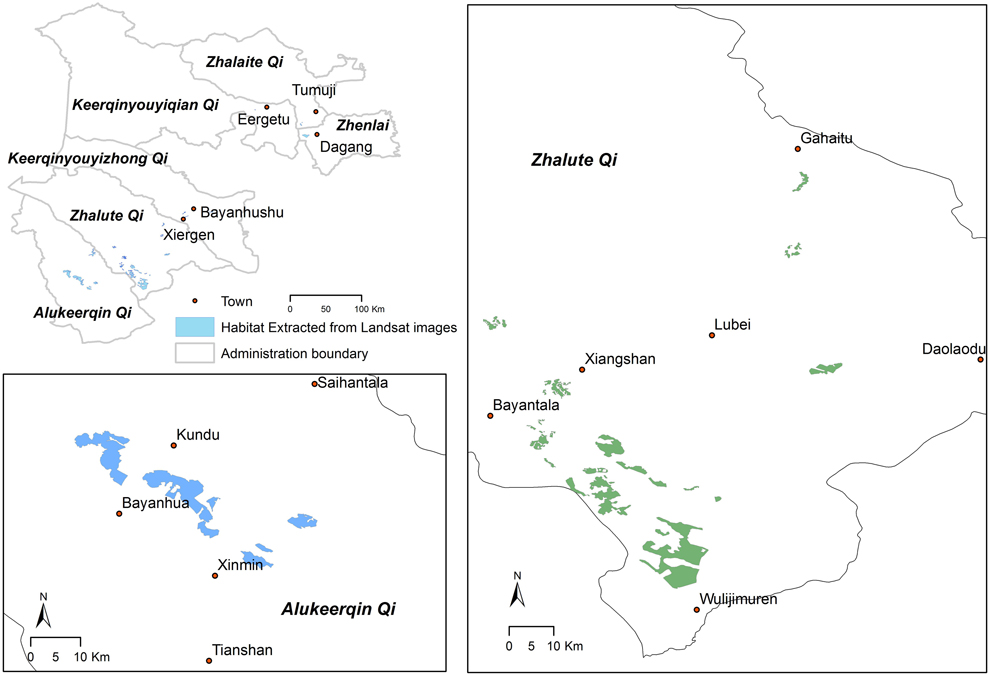
Figure 2. Breeding habitat distribution of Jankowski’s Bunting (up to 2014) derived from Landsat imagery. The upper-left indicates its entire breeding range in China. Suitable habitats are shown in Zhalute Qi (right) and Alukeerqin Qi (lower left). Among the 16 towns, 13 of them were discovered as new breeding sites for the Jankowski’s Bunting, with the exception of Tumuji, Dagang and Xiergen.
The area of suitable breeding habitat for Jankowski’s Bunting is approximately 280 km2, as determined by remote sensing (Figure 2). The estimated population size in China ranges between 9,800 and 12,500 individuals. The currently known populations of Jankowski’s Bunting are scattered within the scope of 16 towns in six counties/cities. The population and habitat sizes at Zhalute Qi were the largest, followed by Alukeerqin Qi and Keerqinyouyizhong Qi (Table 1).
Table 1. The population status of Jankowski’s Bunting in China.
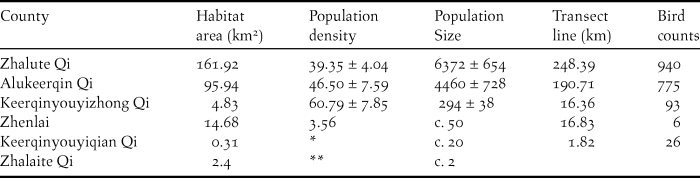
* Only a small flock of Jankowski’s Bunting exists there.
** One pair was recorded in 2012, but no birds were recorded in 2014.
Discussion
Our results show that the population size of Jankowski’s Bunting in China ranges between 9,800 and 12,500 individuals, and the suitable breeding area covers approximately 280 km2, which is mainly located in north-eastern Inner Mongolia but is scattered across different counties. The currently known populations in China are the most significant known globally, especially when the limited information from other historical distribution regions is considered.
Previously, the total population of Jankowski’s Bunting was anecdotally estimated to be fewer than 500 pairs (e.g. Wang and Li Reference Wang and Li2008). Brazil (Reference Brazil2009) assumed that there were approximately 100–10,000 breeding pairs, 50–1,000 individuals in migration and 50–1,000 individuals wintering in China. Having discovered 13 new breeding sites during this survey, declines in the population and the breeding range may be less serious than previously suggested by non-systematic surveys and casual sightings by birdwatchers. However, the situation cannot be considered optimistic. Our findings showed that the distribution range of Jankowski’s Bunting has been substantially reduced from its historical breeding range. In fact, several sites no longer appear to support the species, such as Xinjiamu and Maanshan. The main reason for this decline is habitat loss due to agricultural development, human colonisation, tree planting and overgrazing (Zhao et al. Reference Zhao, Nickel and Groh1994, Gao Reference Gao2002, Jiang et al. Reference Jiang, Gao, Lei, Wan, Zhao and Wang2008, Wang et al. Reference Wang, Jiang and Gao2010, BirdLife International 2012). The current breeding distribution of Jankowski’s Bunting is highly scattered (Figure 2). Habitat isolation and fragmentation may drive population declines. For example, Tumuji, Dagang and Eergetu were isolated from other breeding sites, and the remaining population in Dagang has undergone a dramatic decline from about 60 pairs in 1999 to about 10 pairs in 2010. The population in Tumuji has remained relatively constant at about three (ranging from one to five) pairs (Wang et al. Reference Wang, Jiang and Gao2010). Zhalute Qi and Alukeerqin Qi are newly discovered breeding areas during this study, and they support major populations of Jankowski’s Bunting. However, suitable habitat is severely fragmented here. This fragmentation may affect population persistence by reducing connectivity and restricting dispersal across breeding sites (Fahrig and Merriam Reference Fahrig and Merriam1994). For example, the population severely fluctuates in Xiangshan, where habitat is heavily fragmented, and some isolated habitat patches were recorded as unoccupied during our survey. Jankowski’s Bunting was historically described as a breeding and wintering bird in western Jilin (Fu and Chen Reference Fu and Chen1966), and we also found this species wintering at its current breeding sites in Inner Mongolia. Thus, as a poor disperser, Jankowski’s Bunting is more vulnerable to fragmentation. Genetic studies would be necessary to examine the impact of fragmentation on each of these populations.
The population of Jankowski’s Bunting continues to be vulnerable to a number of threats. First, low reproductive success would result in population declines. Fledging success is a reliable index of recruitment at the population level (Weatherhead and Dufour Reference Weatherhead and Dufour2000); however, the reproductive success of Jankowski’s Bunting in Dagang was very low (23% in Gahaitu, Lubei of Zhalute Qi and Taohai; unpubl. data). Several causes have been identified, including interspecific parasitism by cuckoos, nest abandonment due to grazing disturbance, nest-robbing, predation and other anthropogenic disturbances. Other environmental factors, such as drought and wildfires, also pose severe threats to the breeding and survival of this species (Jiang et al. Reference Jiang, Gao, Lei, Wan, Zhao and Wang2008).
Second, the suitable breeding habitats of Jankowski’s Bunting are vulnerable to degradation due to grazing. Jankowski’s Bunting is strictly selective in its breeding habitat (Gao et al. Reference Gao, Wang and Sun2003). The occurrences of Siberian apricot and higher and dense grasses were crucial factors in its selection of nesting sites (Gao et al. Reference Gao, Wang and Sun2003). Its nest is placed amongst sparse grass on the ground, sometimes at the base of a small tree, and is lined with grass and sometimes horse hair (Fu and Chen Reference Fu and Chen1966). Siberian apricot provides perches for the buntings, which may play an important role in territory defence and predator detection (Tong et al. Reference Tong, Xiao, Bai, Sun, Gao and Wang2002, Bai et al. Reference Bai, Gao and Zhou2003). Higher and dense grasses, especially dry grasses, not only provide nesting materials but also provide shelter for the nests and incubating individuals (Gao Reference Gao2002). The nearby plant assemblages generally consist of Stipa baicalensis and Spodiopogon sibiricus, which are favoured by livestock as well. In addition to causing physical disturbance, grazing generally reduces the availability of plant biomass and alters plant composition, which may reduce the availability of specific food items (i.e. grass seed and insects) for Jankowski’s Bunting. Grazing also reduces overall vegetation height and cover and increases the risk of predation associated with inter-patch movements, reducing the species’ survival. Other human activities, such as digging up medicinal plants and picking the fruits of Siberian apricot, changed vegetation structure and increased nest-robbing probability (Jiang et al. Reference Jiang, Gao, Lei, Wan, Zhao and Wang2008). Fruit-pickers and shepherds, with poor conservation awareness, usually remove the nests out of curiosity.
All currently known Jankowski’s Bunting populations breed in Mongolian steppe-vegetation zones with shrubs dominated by natural Siberian apricots and its suitable breeding habitat is only about 280 km2. Among all breeding areas surveyed, only two of the known sites (Tumuji and Alukeerqin) are officially protected as National Nature Reserves. Most remnant habitats highly benefit from the implementation of the National Key Public Forest Protection Project, which fences protected areas and excludes these habitats from grazing and other agricultural activities. However, the project will be terminated at several sites in 2017, according to China’s State Forestry Administration (http://www.forestry.gov.cn/main/4818/content-797309.html). At other sites, this project was not being strictly implemented, which resulted in portions of suitable breeding habitats being destroyed by overgrazing. In sites where fences have been gradually demolished, such as Saihantala of Alukeerqin Qi and Xiangshan, Daolaodu and Wulijimuren of Zhalute Qi, the local populations declined rapidly. Breeding populations at Xinjiamu and Maanshan have vanished due to habitat degradation. In fact, degraded habitats could be restored if grazing can be managed effectively, which could be done by reducing grazing intensity or by local administrations establishing protected areas.
Third, scattered and small populations are unstable, and the population fluctuates more severely in smaller patches. For example, the population at Tumuji, which is located at the fringe of the Jankowski’s Bunting distribution, varied between one and five pairs from 2001 to 2010 (Wang et al. Reference Wang, Jiang and Gao2010). One pair was recorded in 2012, but no individuals were recorded during this study. At Eergetu of Keerqinyouyiqian Qi, approximately 40 breeding individuals were found centred on a small habitat patch (less than 35 ha) in 2014, but by 2015, the population declined to approximately 20 individuals due to disturbance from raising domestic fowl. The distance to (and from) other breeding sites poses another barrier, as the breeding population cannot disperse easily to colonise suitable neighbouring habitats. At Xiergen, the population was found to be stable within a fenced, small area (about 33 ha), but this was not the case for the surrounding areas.
Duse to lack of current information, the population status of Jankowski’s Bunting in Russia and North Korea remains unclear, but Litvinenko (Reference Litvinenko1989) reported that the bunting declined from being a locally common breeding bird in the mid-1960s to apparent extinction in Russia. Jankowski’s Bunting is currently breeding close to the Mongolian border, suggesting the possible presence of a nearby, previously unknown breeding population. One male Jankowski’s Bunting was recorded in south-eastern Mongolia (46.8°N 116.2°E) in September 2013 by a local photographer. However, no Jankowski’s Buntings were observed in June 2015 in eastern Mongolia, and the population size is likely to be small if populations do exist within the Numrug Protected Area (Muzika et al. Reference Muzika, Fu, Townshend and Yu2015). More surveys should be conducted to ascertain whether unknown breeding populations exist in Mongolia, Russia and North Korea, to provide a more reliable assessment of the status of the species.
In China, Jankowski’s Bunting was historically described as a “locally common” breeding and wintering bird in Dongning, Tumen and western Jilin (Yamashina Reference Yamashina1957, Fu and Chen Reference Fu and Chen1966). Wintering populations or individuals were occasionally found in winter in several regions of Inner Mongolia, Liaoning, Hebei and Beijing (Seys and Licent Reference Seys and Licent1933, Morrison Reference Morrison1948, Cheng Reference Cheng1987, Huang et al. Reference Huang1989). We also found Jankowski’s Bunting wintering at its current breeding sites in Inner Mongolia. Additionally, the species was recently found in Beijing (Xing 2016), suggesting that it is capable of dispersing to areas far outside of its known breeding range. It is important to study its dispersal behaviour (colour-ringing or data-logger studies), which would determine whether young birds are able to colonise distant habitat islands. This will also help to determine if winter habitat requirements are limiting the populations of Jankowski’s Bunting.
In any case, according to the IUCN Red List Categories and Criteria (IUCN 2001), the current status of Jankowski’s Bunting in China still meets criteria B2ab for an ‘Endangered’ species, as the area of occupancy is less than 500 km2, the current population is confined to Inner Mongolia (B2a), and the number of subpopulations in Dagang, Tumuji, and Eertuge is continuously decreasing (B2b). Conservation efforts are urgently needed based on the continued loss and degradation of the species’ habitat and the fact that only two of the known sites have any form of official protection. New protected areas and/or nature reserves should be considered to protect the main breeding populations in Alukeerqin Qi and Zhalute Qi. Ecological studies should be conducted at the breeding sites to clarify habitat requirements during the breeding season, while detailed management regimes need to be established to manage the remaining habitat. Surveys targeted at identifying the dispersal patterns of Jankowski’s Bunting would help determine its dispersal routes and wintering grounds. Genetic studies would also be necessary to examine the impact of fragmentation on each subpopulation. Finally, it is also critical to develop a conservation management plan for this species.
Acknowledgements
This work was supported by the State Bureau of Forestry (Special Investigation Project on Jankowski’s Bunting), the National Natural Science Foundation of China (No. 31172109, 31670398), the Jilin Provincial Science and Technology Department (No. 20150101067JC), and the Fundamental Research Funds for the Central Universities (2412016KJ043). We thank the “National Earth System Science Data Sharing Infrastructure”, National Science & Technology Infrastructure of China (http://www.geodata.cn) for geographical data support. We thank Gui-quan Xiang, Zhi-jie Yang, Tuo Wang, Ji-Yuan Yao, Hui-Juan Tao, Bo Zhang, Jiang-ping YU, Le-yong Chen, Hong-wei Xu, and Yang-yang Yu for their assistance in the field. We thank Pang Chun Chiu, Terry Townshend, Dave Buckingham and anonymous reviewers for their helpful remarks on this paper.




 Open access
Open access